
国際特許分類[A01H1/00]の内容
生活必需品 (1,310,238) | 農業;林業;畜産;狩猟;捕獲;漁業 (84,932) | 新規植物またはそれらを得るための処理;組織培養技術による植物の増殖 (2,345) | 遺伝子型を改変するための処理 (862)
国際特許分類[A01H1/00]の下位に属する分類
交配の方法または装置;人工授粉 (97)
淘汰の処理方法 (11)
突然変異を起こさしめるための処理,例.化学物質または放射線による処理 (37)
国際特許分類[A01H1/00]に分類される特許
681 - 690 / 717
セリアック病に関連するエピトープ
本明細書に開示される発明は、セリアック病を診断、治療及び予防する方法において有用なエピトープに関する。少なくとも一つのエピトープを含んでなる治療用組成物が、提供される。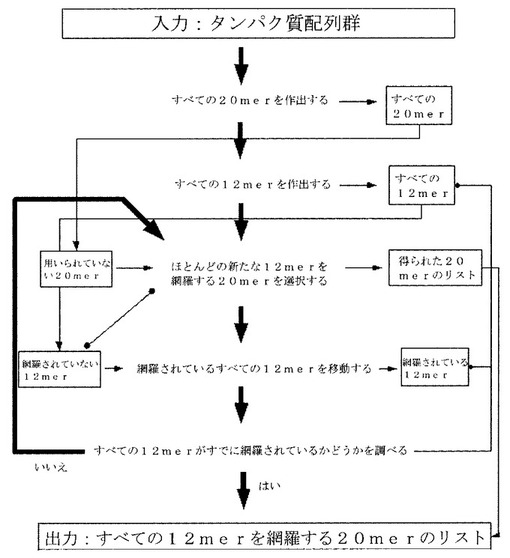
 (もっと読む)
(もっと読む)
改善した生育特性を有する植物及びこれを作製する方法
本発明は、NAP1-様タンパク質をコードする核酸配列の発現をモジュレートすることによる、植物の生育特性を改善する方法に関する。本発明はまた、NAP1-様タンパク質をコードする核酸の発現がモジュレートされている、改善された生育特性を有するトランスジェニック植物に関する。  (もっと読む)
(もっと読む)
増大したデンプンリン酸化酵素活性を有する植物
本発明は、遺伝的に修飾された植物細胞および植物に関し、ここで遺伝的修飾は、遺伝的に修飾されていない対応する野生型植物細胞または野生型植物と比較して、OK1タンパク質のデンプンリン酸化活性の増大をもたらす。さらに本発明は、そのような植物細胞および植物を作製するための手段および方法に関する。これらの型の植物細胞および植物は、修飾デンプンを合成する。したがって、本発明はまた、本発明による植物細胞および植物から合成されたデンプン、これらのデンプンを作製する方法、およびこれらの修飾デンプンのデンプン誘導体の製造、ならびに本発明によるデンプンを含む穀粉に関する。さらに、本発明はまた、デンプンをリン酸化するOK1タンパク質をコードする核酸、そのような核酸分子を含むベクター、宿主細胞、植物細胞、および植物に関する。さらに本発明は、デンプンリン酸化活性を有するOK1タンパク質に関する。 (もっと読む)
改善された成長特性を有する植物とその作製方法

本発明は、S3aタンパク質をコードする核酸を植物に導入し、発現させることにより、植物の成長特性を改善する方法に関する。本発明はまた、S3aタンパク質をコードする核酸を導入したトランスジェニック植物に関し、この植物は、対応する野生型植物と比較して改善された成長特性を有する。本発明はさらに、本発明の方法に有用な構築物に関する。 (もっと読む)
大麦縞萎縮病抵抗性に関与する遺伝子座に連鎖する遺伝マーカーおよびその利用
オオムギの詳細な連鎖地図を作成し、QTL解析を行なうことにより、オオムギ1H染色体上に座乗する大麦縞萎縮病抵抗性に関与する遺伝子座に連鎖する5つの遺伝マーカーと、オオムギ2H染色体上に座乗する大麦縞萎縮病抵抗性に関与する遺伝子座に連鎖する2つの遺伝マーカーと、オオムギ3H染色体上に座乗する大麦縞萎縮病抵抗性に関与する遺伝子座に連鎖する5つの遺伝マーカーと、オオムギ4H染色体上に座乗する大麦縞萎縮病抵抗性に関与する遺伝子座に連鎖する4つの遺伝マーカーと、オオムギ5H染色体上に座乗する大麦縞萎縮病抵抗性に関与する遺伝子座に連鎖する2つの遺伝マーカーとを見出した。 (もっと読む)
収量が向上した植物及び該植物を作出する方法
本発明は、サイクリンD3タンパク質をコードする核酸woシュートで選択的に発現させることができるプロモーターの制御下にある該核酸を植物に導入することにより、植物の収量を向上する方法に関する。また本発明は、シュートでサイクリンD3タンパク質をコードする核酸を選択的に発現させることができるプロモーターの制御下にある該核酸を含むトランスジェニック植物に関し、該植物は、対応する野生型植物と比べて収量が向上する。また本発明は、本発明の方法に有用な構築物に関する。 (もっと読む)
トランスジェニック植物における多価不飽和脂肪酸の製造方法

【課題】 多価不飽和脂肪酸を用いて食物および/または飼料の栄養価を高めるためには、植物系、特にトランスジェニック植物の種子において多価不飽和脂肪酸を簡便で安価に製造する方法が強く切望されている。
【解決手段】 本発明は、トランスジェニック植物の種子中に多価不飽和脂肪酸を産生させる方法に関する。この方法において、ω-3-デサチュラーゼ、Δ-12-デサチュラーゼ、Δ-6-デサチュラーゼ、Δ-6-エロンガーゼ、Δ-5-デサチュラーゼ、Δ-5-エロンガーゼおよび/またはΔ-4-デサチュラーゼ活性を有するポリペプチド、好ましくはΔ-6-デサチュラーゼ、Δ-6-エロンガーゼおよびΔ-5-デサチュラーゼ活性を有するポリペプチドをコードする核酸を生物に導入する。核酸配列は、配列番号11、配列番号27、配列番号193、配列番号197、配列番号199および配列番号201に示されている。有利に、これらの核酸配列は、場合により脂肪酸の生合成または脂質代謝のポリペプチドをコードする他の核酸配列と共に、生物中で発現させることができる。Δ-6-デサチュラーゼ、Δ-5-デサチュラーゼ、Δ-4-デサチュラーゼ、Δ-12-デサチュラーゼおよび/またはΔ-6-エロンガーゼ活性をコードする核酸配列は特に都合がよい。有利に、これらのデサチュラーゼおよびエロンガーゼは、珪藻(Thalassiosira)、ミドリムシ(Euglena)またはオストレオコッカス(Ostreococcus)に由来する。本発明はまた、増加した長鎖多価不飽和脂肪酸含量を有する油および/またはトリアシルグリセリドの製造方法に関する。好ましい実施形態において、本発明はまたアラキドン酸、エイコサペンタエン酸、またはドコサヘキサエン酸の製造方法に関し、増加した不飽和脂肪酸(特にアラキドン酸、エイコサペンタエン酸、および/またはドコサヘキサエン酸)含量を有するトリグリセリドを、トランスジェニック植物中、好ましくはトランスジェニック植物の種子中で産生させる方法に関する。本発明は更に、本発明の方法に用いるエロンガーゼおよびデサチュラーゼの発現に基づいて多価不飽和脂肪酸(特にアラキドン酸、エイコサペンタエン酸、および/またはドコサヘキサエン酸)含量の増加したトランスジェニック植物の製造に関する。本発明はまた、Δ-6-デサチュラーゼ、Δ-6-エロンガーゼ、Δ-5-デサチュラーゼおよびΔ-5-エロンガーゼ活性を有するポリペプチドをコードする核酸配列を一緒にまたは別々に含有する組換え核酸分子、ならびに、これらの組換え核酸分子を含有するトランスジェニック植物に関する。本発明のさらなる態様は、本発明の方法によって産生される油、脂質、および/または脂肪酸、ならびにこれらの使用に関する。さらに本発明は不飽和脂肪酸および不飽和脂肪酸含量の増大したトリグリセリド、ならびにこれらの使用に関する。
(もっと読む)
フィトクロムと相互結合作用する新規蛋白質及びこれの用途(NOVELPHYTOCHROME−INTERACTINGPROTEINANDAUSETHEREOF)
本発明はフィトクロムと相互結合する新規蛋白質及びこれの用途に関し、より詳しくは配列番号4で表示されるアミノ酸配列又は前記アミノ酸配列と少なくとも70%以上の相同性を有するアミノ酸配列を有するポリペプチド及びこれの用途に関するものである。本発明で提供するポリペプチドはフィトクロムA及びBと交叉相互結合し、前記相互結合にはそのN−末端に存在するTPRドメインが関与する。さらに、前記ポリペプチドのC−末端には、脱リン酸化酵素活性を担当するPP2A触媒ドメイン(PP2Ac)が存在する。本発明に伴うポリペプチドは脱リン酸化酵素として利用することができ、光信号伝達に敏感な植物体の製造に有用に利用できる。さらに、本発明のポリペプチド内に存在するTPRドメインは矮性植物体の製造に利用できる。 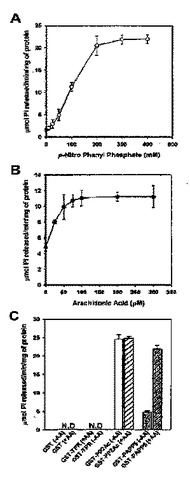 (もっと読む)
(もっと読む)
ウイルス性疾患に耐性がある接木植物及びその生成法
本発明は、ウイルス性疾患に耐性があるトランスジェニック根茎及び敏感な接ぎ穂を含む接木植物に関するものであり、疾患に対する耐性は、接木植物全体がウイルス性疾患に耐性があるように、トランスジェニックウイルス耐性根茎から接ぎ穂に与えられる。本発明はさらに、接木型ウイルス耐性植物の生成法、及びそれによって生成される植物に関する。 (もっと読む)
ピリドキシン生合成関連機能を有するポリペプチド、それを暗号化するポリヌクレオチド、及びそれらの用途
本発明は、ピリドキシン生合成関連機能を有するポリペプチド、これを暗号化するポリヌクレオチド、及び、その用途を開示する。特に、本発明は、ピリドキシン生合成関連機能を有するポリペプチド、それを暗号化するポリヌクレオチド、植物の生長を抑制する方法、植物の生長を抑制する物質のスクリーニング方法、スクリーニング方法により得られた物質を含む植物の生長抑制用組成物を開示する。
 (もっと読む)
(もっと読む)
681 - 690 / 717
[ Back to top ]