
サブユニットとしてOSMRβを含む受容体を介してケラチノサイト移動及び表皮修復を促進するための組成物及びその使用
本発明は、表皮修復の分野に関する。より具体的には、本発明は、サブユニットとしてOSMRを含むヘテロマー受容体を活性化できる分子の、ケラチノサイトによるインビトロ及び/又はインビボのケラチノサイト移動及び/又は抗菌ペプチドの発現を活性化するための組成物を製造するための使用に関する。特に、本発明は、化粧用又は皮膚用組成物の製造のための、OSM及び/又はIL-17及び/又はTNFα及び/又はIL-31或いはこれらのアゴニストの使用に関する。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、表皮修復(repair)の分野に関する。より具体的には、本発明は、インビトロ及び/又はインビボのケラチノサイト移動(migration)を増強するための組成物の製造のための、サブユニットとしてOSMRβを含むヘテロマー受容体を活性化できる分子の使用に関する。
【背景技術】
【0002】
皮膚は、生体とその環境との間の保護的界面を提供する大きく複雑な組織である。表皮は、その外表面を形成し、ケラチノサイトと名付けられた特殊化された上皮細胞の複数の層から主に構成されている。皮膚は、微生物、化学物質、動作、物理的損傷、加齢、U.V.照射、癌、自己免疫疾患又は炎症性疾患を含む多くの異なる原因により損傷され得る。
【0003】
表皮の恒常性は、皮膚の基底層から角化層に分化するケラチノサイトの分化と増殖とのバランスにより調節される。表皮ストレスに応答して、又はある皮膚疾患において、この平衡が破壊される。ケラチノサイトは、表皮成長因子(Epidermal Growth Factor (EGF))ファミリーのメンバーのような可溶性のメディエイタ、並びにさらなる成長因子及びサイトカイン(FGFs、IGF-1、PDGF、HGF、TGFβファミリーのメンバー、GM-CSF、TSLP、IL-1、TNF-α)に対して異なる応答ができるようになる。これらの修飾物質は、ケラチノサイト自体、皮膚繊維芽細胞、ランゲルハンス細胞により産生されるか、又はTリンパ球のような免疫浸潤細胞により産生される。これに応答して、ケラチノサイトはさらなるシグナル伝達分子を放出し、細胞表面受容体の発現レベルを変調し、それらの細胞骨格形態を修飾して、それらの移動、分化及び増殖の能力を変調する。これらの変化は炎症反応に関連し、創傷治癒又は慢性疾患のいずれかに導く。
【0004】
IL-6ファミリーのサイトカインは、多数の生体のシステム、例えば免疫、造血、神経発達、生殖、骨モデリング及び炎症プロセスにおいて細胞の成長及び分化を調節する多機能タンパク質である。このサイトカインファミリーは、9つの異なるメンバーを含む:IL-6、IL-11、IL-27、白血病抑制因子(LIF)、カルジオトロフィン(cardiotrophin)-1、カルジオトロフィン様因子、繊毛様神経栄養因子、ニューロポエチン(neuropoietin)、及びオンコスタチンM (OSM)である。これらのサイトカインの活性は、ダイマー又はトリマーの受容体複合体のリガンド誘発性オリゴマー化を介して媒介される。サイトカインのIL-6ファミリーは、それらの各ヘテロマー受容体の形成においてgp130受容体サブユニットを共有している(Taga及びKishimoto 1997)。IL-31と名付けられた最近記載されたサイトカインは、Dillonらによってgp130-IL6ファミリーの新しいメンバーとして分類された。なぜなら、その受容体は、gp130様I型サイトカイン受容体(GPL)及びOSMRサブユニットを含むヘテロダイマーであるからである(Dillon, Sprecherら 2004)。
【0005】
いくつかの文献は、IL-6ファミリーのいくつかのメンバーがある皮膚疾患及び創傷治癒プロセスに加わっているであろうことを報告している。IL-6、IL-11、LIF及びOSMが、乾癬外傷において増加することが見出されており(Bonifati, Mussiら 1998)、IL-6及びLIFは、精製ケラチノサイトから産生される(Paglia, Kondoら 1996; Sugawara, Gallucciら 2001)。創傷治癒プロセスの欠陥が、IL-6及びSTAT3欠損マウスにおいて報告されている(Sano, Itamiら 1999; Gallucci, Simeonovaら 2000)。しかし、IL-6欠損マウスから単離された培養ケラチノサイトでのさらなる研究は、ケラチノサイト移動に対するIL-6の作用が、皮膚繊維芽細胞により媒介されることを示した。実際に、IL-6単独では、該IL-6欠損ケラチノサイトの増殖又は移動を著しく変調はしなかったが、IL-6は、皮膚繊維芽細胞と同
時培養したときに、その移動を著しく誘発した(Gallucci, Sloanら 2004)。
【0006】
OSMは、活性化されたT細胞、サイトカインにより刺激された単球及び樹状細胞から分泌される。OSMは、炎症誘発性メディエイタであり、肝細胞においてタンパク質合成を強く引き起こす(Benigni, Fantuzziら 1996)。ヒトにおいては、OSM及びLIFは、成長調節の増加、分化、遺伝子発現、細胞生存により、いくつかの組織において重複した生物学的機能を示す。OSMは、LIFと共有していないいくつかの独特の生物学的機能、例えばいくつかの腫瘍細胞系統の成長阻害又はAIDS関連カポジ肉腫細胞の刺激を惹起することも知られている。OSMのこれらの共通及び独特の機能は、OSM受容体複合体が2種類存在することにより説明される。gp130/LIFRβサブユニットで形成される通常のLIF/OSM受容体複合体の他に、OSMは、gp130をOSMRβ(「OSMR」又は「OSM-R」ともよばれる)と結合させるII型受容体を特異的に認識でき、これは内皮細胞、肝細胞、肺細胞、繊維芽細胞、造血細胞及びいくつかの腫瘍細胞系統により発現される。その後のシグナル伝達カスケードは、ヤヌスキナーゼ(JAK 1、JAK 2、Tyk 2)の活性化を含み、その後、シグナル伝達性転写因子(STAT1、STAT3)及びMapキナーゼ経路の活性化が続く。
【0007】
その抗腫瘍性活性及び炎症誘発性反応におけるその役割に加えて(Wahl及びWallace 2001; Shoyabら, US 5,451,506; Richardsら, US 5,744,442)、OSMは、MAPキナーゼ依存性経路により皮膚繊維芽細胞の成長を刺激して、皮膚の創傷治癒を促進すると記載されている(Ihn及びTamaki 2000)。
【0008】
他のサイトカインも、皮膚に対して影響があることが知られている。例えば、Dillonら(上記)は、IL-31の過剰発現が、アレルギー性及び非アレルギー性の疾患を特徴付ける皮膚炎及び上皮反応の促進に関与し得ることを示唆する。これらの著者らは、IL-31を皮膚修復促進のために用いることを示唆していない。
【0009】
皮膚が損傷したときに、その完全な修復は、真皮及び表皮の両方が修復されることを包含する。異なる細胞の種類を含む表皮及び真皮の治癒(healing)は、異なる機構を伴う。
【0010】
現在、皮膚の治癒を改善するための治療は、主に真皮を標的としている。しかし、表皮の再構成が、完全な回復のために必要である。例えば大きい熱傷、潰瘍又は褥瘡の場合は、生理的表皮治癒プロセスは、皮膚の防御機能を回復するために充分に効果的ではない場合がある。このような場合には、損傷を受けた領域を迅速に覆い、感染及び乾燥の可能性を回避することが必要である。表皮の再生を刺激することも必要である。体表面の半分未満の重篤な熱傷の場合には、焼けた皮膚の切除の後に自己移植が行われる。その目的のために、患者から健全な皮膚が取られ、その表面積を増やすために機械的に処理される。この「ウィック(wick)皮膚」は、次いで、損傷の上に移植される。熱傷の表面積が大きすぎる場合(体表面の半分を超える)、このプロセスは実行不可能である。そこで、乾燥及び感染を回避するために創傷を一時的に覆うことが必要である。このことは、現在、死体からの皮膚又は創傷治癒を改善するために成長因子を組み込んでもよい無細胞ドレッシング、例えばツルグラのような皮膚代替物のいずれかを用いて行われている。このような皮膚代替物の例は、US 6,132,759又はWO 01/41820に記載される。並行して、患者からの皮膚細胞を、上皮層を得てこれを次いで移植するためにインビトロで増殖させる。数cm2の健全な皮膚から、1〜2 m2が3週間で得られる。しかし、これらの技術は、長期間で費用がかかり、そして成功して行うためには重大なインフラストラクチャーが必要である。
【発明の開示】
【発明が解決しようとする課題】
【0011】
よって、表皮の修復を改善するための新規な皮膚科学的アプローチが真に必要とされていることが明らかである。ケラチノサイトの求心的移動(centripetal migration)を増強することは、治癒及び再上皮形成を明らかに促進/可能にするであろう。ケラチノサイトの分化及び移動に対して作用することは、表皮水疱症のような特定の疾患の治療のためにも必要である。「表皮水疱症(bullous emolysis)」の用語は、表皮の剥離及び真皮と表皮の間の液体の蓄積に導く、起源が異なる(例えば出血、熱傷、自己免疫疾患など)いくつかの皮膚炎のことをいう。表皮水疱症の具体例は、水疱性類天疱瘡である。
【0012】
最近の小さい創傷又は古い切傷、斑、伸展裂創(stretch marks)などのいずれかに由来する瘢痕の外観(aspect)を改善するための効果的な組成物が存在しない化粧分野においても、表皮修復を改善することは重要である。
【0013】
この関係において、本発明者らは、いくつかのサイトカイン、特にOSM及びIL-31が、ケラチノサイトの移動を増強できることを見出した。興味深いことに、これらの2種のサイトカインは、ともにサブユニットとしてOSMRβを含む異なるヘテロマー受容体に結合する。本発明者らは、通常のヒト表皮ケラチノサイトが、gp130、GPL及びOSMRβを発現することを示している。
【0014】
以下の実験実施例において開示されるように、OSMは、ヒト表皮ケラチノサイトにおいて、STAT3シグナル伝達経路及びMAPキナーゼ経路を補充する(recruit)。OSMは、これらの細胞において、ケモカイン、デフェンシン及びソリアシン(psoriasin)を含む炎症誘発性遺伝子の発現をアップレギュレートする。OSMは、また、再構成されたヒト表皮の厚みを増加させ、一連の分化抗原をダウンレギュレートする。興味深いことに、他のサイトカイン、特にIL-17及びTNFαは、OSMと相乗的に作用し、その効果を強化する。
【0015】
本発明者らが行った実験はまた、IL-31がケラチノサイト移動を媒介することも明らかにした。しかし、本発明者らは、グリア芽腫及び黒色腫の腫瘍細胞において、IL-31の作用が、ヘテロマー受容体の形成におけるOSMRβに含まれるGPLサブユニットの種類に依存することを観察した。特に、本発明者らは、GPL受容体の短い形がIL-31のシグナル伝達に対して著しい阻害効果を発揮し、優性の負の受容体としてふるまうことに注目した。
【0016】
これらの結果を合わせると、OSM及びIL-31が、創傷治癒及びケラチノサイト移動を伴う他のプロセスにおいて重要な役割を演じることが示される。これらの効果は、ヘテロマー受容体のOSMRβサブユニットが参加するシグナル変換を必要とする。
【課題を解決するための手段】
【0017】
本発明の第一の目的は、よって、サブユニットとしてOSMRβを有するヘテロマー受容体の少なくとも1つの活性化物質の、表皮修復を改善するための組成物の製造のための使用である。
【発明を実施するための最良の形態】
【0018】
このような活性化物質の例は、OSMRβサブユニットを含むヘテロマー受容体のリガンド、及びそのアゴニストである。本発明による好ましい活性化物質は、サイトカイン、特にオンコスタチンM (OSM)及びインターロイキン31 (IL-31)、並びにそれらのアゴニストである。以下において、「OSM」及び「IL-31」は、サイトカイン自体、並びにそれらの各アゴニストのことをいうのに用いる。本発明による活性化物質のその他の例は、OSMRβとともにヘテロマー受容体を形成できる受容体サブユニットの発現をアップレギュレートする分子である。
【0019】
OSMの作用を強化するIL-17及びTNFα、並びにIL-31の効果を強化するガンマインターフェロン(IFN-γ)も、本発明による活性化物質と考えることができ、それぞれOSM又はIL-31と組み合わせて、それらの効果を増大し、それにより本発明による組成物中でより低い濃度で用いることを可能にするために、有利に用いることができる。もちろん、これらのサイトカインは、それらのアゴニストで置き換えることができる。変異サイトカイン、サイトカインではない天然又は変異のペプチド、及び非ペプチドの合成又は天然の分子は、本発明による活性化物質として用いることができる。
【0020】
本発明の好ましい実施形態によると、OSM、IL-17及びTNFαは、表皮修復を改善するための組成物の製造のために用いられる。
本発明により得られる組成物は、ケラチノサイト移動の促進、又は表皮治癒の促進のために用いられる。これらの組成物は、大きい創傷の場合に、ケラチノサイトの求心的移動の刺激に有利に用いられる。これらは、表皮水疱症の予防、減弱又は治療のために用いることもできる。実際に、その起源及び段階に応じて、表皮水疱症は、ケラチノサイト移動を増強する分子、又は逆に抗炎症性分子のいずれかを投与することにより治療できる。医師は、状況に応じて、表皮水疱症を罹患した患者にとって、OSM、IL-31及び/又はそれらのアゴニスト並びに強化物質を用いる処置がいつ効果的であり得るか、理解するであろう。
【0021】
本発明による組成物は、例えば局所投与によりインビボで、又はインビトロでのいずれかで表皮の厚みを増加させるために用いて、(ヒトの)再構成される表皮の質及び/又は量を増加させ、例えば移植の必要がある患者のために上皮層の生成を促進することもできる。
【0022】
本発明の特定の実施形態によると、サブユニットとしてOSMRβを有するヘテロマー受容体の活性化物質、又は該受容体を活性化する分子の組み合わせは、手、口唇、顔若しくは体のひび割れを予防するか及び/又は減じるため、或いは伸展裂創を予防するか及び/又は減じるための組成物の製造のために用いられる。本発明により得られる組成物の別の適用は、瘢痕の外観及び快適さ(comfort)の改善、及び/又は表皮創傷の治癒の間の表皮創傷の外観及び快適さの改善である。本発明のこの態様によると、創傷はいずれの起源でもあり得る。創傷は、切傷、熱傷、擦過傷などのような外傷によるもの、外科切開及び皮膚移植のような外科的手法によるもの、並びににきび、アトピー性皮膚炎、湿疹、職業上の皮膚炎、脂漏性皮膚炎、酒さ、紅斑、焼痂、糖尿病(足)、角化上皮症、鱗状化(squama)、潰瘍、魚鱗癬(ichtyosis)、表皮水疱症、足穿孔、疣贅、らい病感染(leprae infection)などのような障害及び疾患によるものを含む。もちろん、炎症性疾患又はケラチノサイト過剰増殖に関連する疾患による創傷の場合、医師は、本発明による組成物を投与する前に該疾患が治療されるように制御するであろう。
【0023】
本発明により製造される組成物は、局所投与用に製剤化されることが好ましい。これらは、例えばクリーム、ローション、軟膏又はドレッシングの形であり得る。
【0024】
本発明による具体的な化粧用及び/又は皮膚用組成物は、OSM又はIL-31を含む。
【0025】
本発明は、また、皮膚用及び/又は化粧用組成物の製造のための、上記の少なくとも2種のサイトカインの使用にも関する。これらの少なくとも2種のサイトカインは、同じ組成物中に混合されるか、又は部分品(parts)のキットで提供されるかのいずれかであり得る。本発明のこの態様によると、好ましい部分品のキット又は組成物は、OSMと、IL-17、TNFα及びこれらのサイトカインのアゴニストから選択される少なくとも1種のサイトカインとを含む。本発明による好ましい化粧用及び/又は皮膚用組成物は、OSM、IL-17及びTNFαを含む。本発明による組成物の別の具体的な実施形態において、上記の組成物は、IL-31及びIFN-γ、及び/又はOSMを含む。
【0026】
OSMを含む本発明による組成物において、OSMは、好ましくは0.1〜100 ng/ml、より好ま
しくは1〜20 ng/mlの濃度である。この濃度は、IL-17及び/又はTNFαの存在下で0.01 ng/mlまで減じることができる。このような複合組成物において、OSM及び/又はIL-17及び/又はTNFαの濃度は、好ましくは0.01 ng/ml〜10 ng/ml、より好ましくは0.1〜2 ng/mlである。IL-31を含む組成物において、後者は、好ましくは1〜500 ng/ml、より好ましくは10〜100 ng/mlの濃度である。
【0027】
上記のように、培養したケラチノサイトに、サブユニットとしてOSMRβを有するヘテロマー受容体の少なくとも1種の活性化物質を加えることにより、(ヒトの)再構成される表皮の質及び/又は量を増加させることができる。よって、本発明は、上記の少なくとも1種の活性化物質を、培養にあるケラチノサイトに加える工程を含む、再構成されたヒト表皮を製造する方法にも関する。例えば、この方法は、OSM及び/又はIL-17及び/又はTNFαを、該ケラチノサイトの培養培地に、各サイトカインについて好ましくは0.01〜10 ng/mlの範囲の濃度で加えることにより行うことができる。
【0028】
本発明は、以下の図面及び実施例によりさらに説明される。
図の説明
図1は、NHEKによるOSM受容体の発現を示す。
(A) トータルRNAを、4人の別個のドナーのNHEKから抽出した。RT-PCRを、OSMR、gp130、LIFR及びGAPDH遺伝子についての特異的プライマーを用いて行った。cDNAの連続希釈を増幅して、転写産物の発現レベルの半定量的分析を行った。PCR産物を、アガロースゲル電気泳動により分析した。(B) NHEK細胞表面の免疫標識及びフラックスサイトメトリー分析。Gp130及びOSMRは、細胞上で明確に検出されるが、LIFRは検出されない。(C) NHEK及びグリア芽腫細胞系統GO-G-UVMからの細胞溶解物20μgを、SDS-PAGE (10%)により分離し、ニトロセルロースメンブレンに移した。ポンソーレッド染色を用いて、ロード量(loading)の均一性を制御した。gp130、OSMR、LIFR及びチューブリンのバンドの検出は、ウェスタンブロットにより評価した。結果は、3つの独立した実験の代表である。
【0029】
図2は、NHEKでのOSMによるSTAT3及びMAPキナーゼのリン酸化の誘導を示す。
(A) NHEKは、LIF若しくはOSM (50 ng/ml)で刺激したか、又はしなかった。(B) NHEKは、15分間、50 ng/mlのIL-5 (ネガティブコントロール)又は100、50、25、12.5、6.25若しくは3.1 ng/mlのOSMで刺激したか、又は刺激せず、ホスホ-STAT3 (P-STAT3)及びSTAT3タンパク質のレベルをウェスタンブロットにより評価した。サイトカインを用いた刺激の前に、細胞を、中和抗体、抗-gp130 (AN-HH1)又は抗-OSMR (XR-M70)モノクローナル抗体の存在下で、又はアイソタイプコントロールmAb MC192 (最終抗体濃度15μg/ml)とともに、2時間インキュベートした(C) 。OSMに応答してのホスホ-MAPK (P-MAPK)及びMAPKタンパク質レベルは、ウェスタンブロットにより評価した(D)。
【0030】
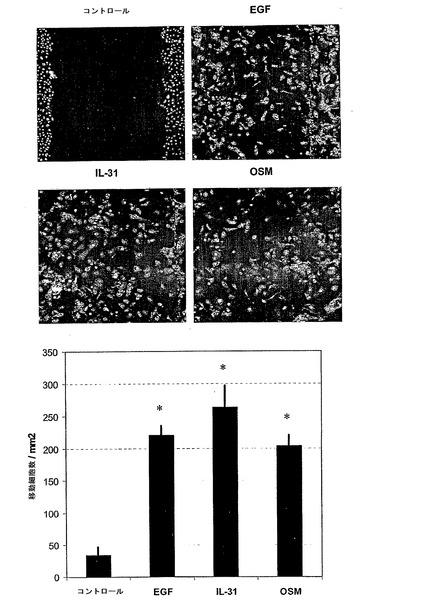
図3は、ケラチノサイト移動に対するOSMの影響を示す。
インビトロ創傷をマイトマイシンで処理した集密なNHEK培養物に導入し、ケラチノサイトを10 ng/mlのEGF若しくはOSMとともに、又は用いずに48時間培養した。無細胞領域への細胞移動を、材料及び方法に記載されるようにして評価した。各バーは、4つの非重複領域で計数された移動ケラチノサイトの平均±SEMを表す。1つの実験は2の代表である。
* スチューデントのt検定に基いて、サイトカインを用いない各コントロールと比較してp<0.001。
【0031】
図4は、OSMで刺激したNHEK及びOSMで処理したRHEから得られる発現プロフィールを示す。
NHEK (A)又はRHE (B)は、10 ng/mlのOSMとともに、又は用いずに24時間培養した。トータルRNAを単離し、Dnase Iで処理し、33P-標識cDNAプローブを作製するのに用い、これをcDNAアレイにハイブリダイズさせた。Molecular Dynamics Storm保存スクリーンに5日間
露光し、さらにスキャンした後に、コンピュータ画像を得た。局所のバックグラウンドを引いた後に、二重のスポットからの平均シグナル強度を、ハウスキーピング遺伝子について得られた値を用いて、プローブ標識における差について標準化した。(C) OSM-誘発変調は、刺激していない細胞についてのシグナル強度に対する、各サイトカインで処理した細胞についてのシグナル強度のパーセンテージ比率として表す。
【0032】
図5は、NHEKによるS100A7〜9合成に対するOSMの影響を示す。
NHEKは、0.4、0.8、1.6、3.1、6.3、12.5若しくは25 ng/mlのOSMとともに、又は用いずに48時間(A)、或いは10ng/mlのOSMとともに、又は用いずに6、12、24、48、72、96時間(B)培養した。トータルRNAを抽出し、逆転写し、S100A7及びHMBS mRNAの相対的発現を、リアルタイムPCRにより定量した。HMBSをハウスキーピング遺伝子として用いて、材料及び方法に説明するようにして遺伝子発現を標準化した。コントロール細胞に対する刺激された細胞の相対的発現として表される結果は、2つの独立した実験の代表である。(C) NHEKは、10 ng/mlのOSMとともに、又は用いずに48時間培養した。S100A8-カルグラヌリンA、S100A9-カルグラヌリンB、β-デフェンシン2及びフィラグリンmRNAの相対的発現を、定量RT-PCRにより定量した。結果は、コントロール細胞に対する刺激された細胞の相対的発現として表す。(D) NHEKを、10 ng/mlのOSMとともに、又は用いずに48及び96時間培養した。細胞溶解物20μgをSDS-PAGE (16%)により分離し、ニトロセルロースメンブレンに移した。ポンソーレッド染色を用いて、ロード量の均一性を制御した。S100A7、S100A8及びS100A9タンパク質レベルは、ウェスタンブロットにより決定した。結果は、3つの独立した実験の代表である。
【0033】
図6は、NHEKによるサイトカイン及びケモカインの産生を示す。IL-1β、IL-6、IL-8、IL-10、IL-12p70、TNFα、ENA-78、MIP 3βは、48時間のNHEK培養上清中で特異的ELISAにより測定した。細胞は、OSMの存在下又は非存在下で培養した。ENA78産生の用量応答(0.4〜25 ng/ml)、及び動態(6〜96h)の研究も行った。
【0034】
図7は、10 ng/mlのOSMを用いて4日間刺激したか又はしなかったRHEの組織学的及び免疫組織化学的分析を示す。RHEは固定し、パラフィンに包埋した。4ミクロンの垂直切片を、ヘマトキシリン/エオシンで染色したか、又は抗-K10ケラチンmAb、抗-フィラグリンmAb又は抗-S100A7 mAbと反応させ、次いで顕微鏡の下で写真撮影した(倍率×200)。
【0035】
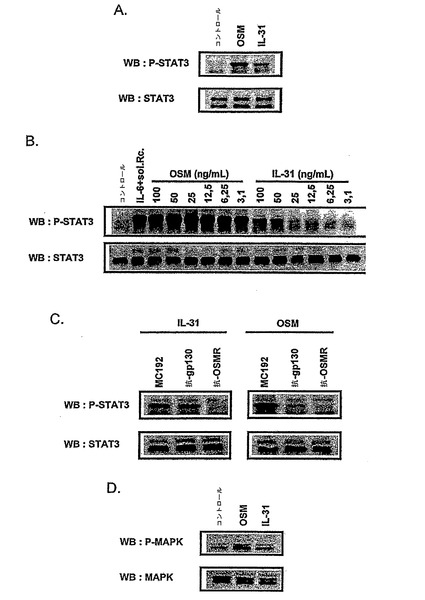
図8は、NHEKにおけるIL-31及びOSMによるSTAT3及びMAPキナーゼリン酸化の誘導を示す。
NHEKは、10 ng/mlのOSM若しくは100 ng/mlのIL-31(A)、又は種々の濃度のこれらのサイトカイン(B)で15分間刺激したか又はせずに、ホスホ-STAT3 (P-STAT3)及びSTAT3タンパク質レベルを、ウェスタンブロットにより評価した。サイトカインでの刺激の前に、細胞を、中和抗体である抗-gp130 (AN-HH1)若しくは抗-OSMR (XR-M70)モノクローナル抗体の存在下で、又はアイソタイプコントロールであるmAb MC192 (最終抗体濃度15μg/ml) (C)と2時間インキュベートした。IL-31及びOSMに応答するホスホ-MAPK (P-MAPK)及びMAPKタンパク質レベルは、ウェスタンブロットにより評価した(D)。
【0036】
図9は、S100A7 mRNA発現に対するIL-31及びOSMの影響を示す。
NHEKは、3.1、6.3、12.5、25、50又は100 ng/mlのIL-31とともに、若しくは用いずに(A)、又は0.4、0.8、1.6、3.1、6.3、12.5又は25 ng/mlのOSMとともに、若しくは用いずに(B) 48時間培養した。100 ng/mlのIL-31 (C)又は10 ng/mlのOSM (D)の存在下又は非存在下でのS100A7 mRNA発現の動態の研究。トータルRNAを抽出し、逆転写し、S100A7及びHMBS mRNAの相対的発現を、リアルタイムPCRにより定量した。HMBSは、材料及び方法に説明するようにして、ハウスキーピング遺伝子として用いて遺伝子発現を標準化した。コントロール細胞に対する刺激された細胞の相対的発現として表される結果は、2つの独立した実験の代表である。
【0037】
図10は、ケラチノサイト移動に対するIL-31及びOSMの影響を示す。
インビトロ創傷は、マイトマイシンで処理した集密なNHEK培養物に導入し、ケラチノサイトは、100 ng/mlのIL-31又は10 ng/mlのEGF若しくはOSMとともに、又は用いずに48時間培養した。無細胞領域への細胞移動は、材料及び方法に記載するようにして評価した。各バーは、4つの非重複領域で計数された移動ケラチノサイトの平均±SEMを表す。1つの実験は、2の代表である。* スチューデントのt検定に基づいて、サイトカインを用いない各コントロールと比較してp<0.001。
【0038】
図11は、S100A7、hBD4/2及びKRT10 mRNA発現に対するいくつかのサイトカインのカクテルの影響を示す。
集密な通常の正常ヒトケラチノサイト(NHEK)を、記載する混合サイトカイン(各サイトカインは1 ng/mlの最終濃度)と24時間処理した。トータルRNAを抽出し、逆転写し、選択した遺伝子の発現を、リアルタイムPCRにより分析した。
【0039】
実施例1:材料及び方法
細胞培養、サイトカイン及び試薬
NHEKは、健康な胸部の皮膚の外科的サンプルから得た。研究実験のためのこれらのサンプルの使用は、ポワチエ病院の倫理委員会(the Ethical Committee of the Poitiers Hospital)により承認された。皮膚サンプルを、ディスパーゼ溶液(25 U/ml;Invitrogen Life Technologies, Cergy Pontoise, France)中で4℃にて一晩インキュベートした。表皮のシートを真皮からはずし、NHEKを、37℃にて15分間のトリプシン消化(トリプシン-EDTA, Invitrogen)により分離した。細胞を、ウシ脳下垂体抽出物(25μg/ml)及び組換え表皮成長因子(0.25 ng/ml)を補った無血清ケラチノサイト培地(Keratinocyte SFM;全てInvitrogenから購入)で培養した。刺激の前に、NHEKは、成長因子を加えないKeratinocyte SFM中で48時間枯渇させた。
【0040】
ヒト組換えOSM、IL-5、IL-6、可溶性IL-6Rは、R&D Systems (Oxon, UK)から購入した。IgG1アイソタイプコントロール(MC192)、抗-gp130 (AN-HH1)、抗-OSMR抗体(AN-A2)及び抗-LIFR (AN-E1)は、実験室で作製した。ホスホ-STAT3、ホスホ-MAPK、MAPKに指向させた抗体は、Upstate Biotechnology (Lake Placid, NY)から購入した。抗-STAT3、抗-S100A8及び抗-S100A9抗体は、Santa Cruz Biotechnology (Santa Cruz, CA, USA)からであった。抗-S100A7抗体は、Imgenex (San Diego, CA, USA)から購入した。ヤギ抗マウス及び抗ウサギペルオキシダーゼ標識免疫グロブリンは、Clinisciences (Montrouge, France)からであり、ウサギ抗ヤギペルオキシダーゼ結合抗体は、Sigma (Amersham Biosciences)からであった。
【0041】
RT-PCR及びRT-リアルタイムPCR分析
トータル細胞RNAは、Trizol試薬(Invitrogen)を用いて単離し、DNase I (0.05 U/μl;Clontech, Palo Alto, CA, USA)で処理した。cDNAを、2μgのトータルRNAから、MMLV逆転写酵素(Promega, Madison, WI)を用いるランダムヘキサマープライマーにより合成した。逆転写産物は、その後、OSMR (フォワード5'-CCTGCCTACCTGAAAACCAG-3' (配列番号1)とリバース5'-ACATTGGTGCCTTCTTCCAC-3' (配列番号2))、gp130 (フォワード5'-GGGCAATATGACTCTTTGAAGG-3' (配列番号3)とリバース5'-TTCCTGTTGATGTTCAGAATGG-3' (配列番号4))、LIFR (フォワード5'-CAGTACAAGAGCAGCGGAAT-3' (配列番号5)とリバース5'-CCAGTCCATAAGGCATGGTT-3' (配列番号6))及びGAPDH (フォワード5'-ACCACAGTCCATGCCATCAC-3' (配列番号7)とリバースTCCACCACCCTGTTGCTGTA (配列番号8))についてのプライマーを用いる25サイクルのPCRにより増幅した。増幅産物は、2%アガロースゲル電気泳動により分析した。
【0042】
定量リアルタイムPCRは、LightCycler-FastStart DNA MasterPLUS SYBR Green Iキット
(Roche, Mannheim, Germany)を用いて行った。反応成分は、1×FastStart DNA MasterPLUS SYBR Green I、並びに0.5μMのS100A7 (フォワード5'-GCATGATCGACATGTTTCACAAATACAC-3' (配列番号9)とリバース5'- TGGTAGTCTGTGGCTATGTCTCCC-3' (配列番号10))、S100A8 (Pattyn, Spelemanら 2003)、S100A9 (フォワード5'-GCTCCTCGGCTTTGACAGAGTGCAAG-3' (配列番号11)とリバース5'-GCATTTGTGTCCAGGTCCTCCATGATGTGT-3' (配列番号12))、hBD2/4 (フォワード5'-GCCATCAGCCATGAGGGTCTTG-3' (配列番号13)とリバース5'-AATCCGCATCAGCCACAGCAG- 3' (配列番号14))、KRT10 (フォワード5'-GCCCGACGGTAGAGTTCTTT-3' (配列番号15)とリバース5'-CAGAAACCACAAAACACCTTG -3' (配列番号16))、ハウスキーピング遺伝子としてのヒドロキシメチル-ビラン合成酵素(HMBS) (Vandesompele, De Preterら 2002)についてのフォワード及びリバースプライマーであった。ヨウ化プロピジウムを用いるcDNAの蛍光定量の後に、250 ng、25 ng及び2.5 ngのcDNAを、LightCyclerのガラスキャピラリにPCR鋳型として加えた。サイクル条件は、95℃にて10分間のポリメラーゼ活性化と、95℃で10秒、64℃で5秒及び72℃で18秒の50サイクルと、単独の蛍光測定とを含んでいた。1秒当たり0.1℃の加熱速度で60℃から95℃まで温度を増加させることにより得られた融解曲線分析、及び連続蛍光測定により、融合温度と思われる狭い単独ピークが明らかになった。数学的モデルを用いて、HMBS参照遺伝子と比較した標的遺伝子の相対的定量を決定した(Pfaffl 2001)。
【0043】
cDNAマクロアレイを用いる遺伝子発現のプロファイリング
トータルRNAを、PCR実験について記載したようにして単離した。DNase処理、ポリA+ RNA富化、33P-標識cDNAプローブ合成、精製及びカスタムAtlasアレイメンブレンへのハイブリダイゼーション(Bernard, Pedrettiら 2002)は、Clontech (Clontech, Palo Alto, USA)の推奨に従って行った。メンブレンは、Molecular Dynamics Storm保存スクリーン(storage screen)に5日間露光し、ホスホリイメージャースキャナ(Molecular Dynamics Stormアナライザ, Amersham Biosciences, Uppsala, Sweden)を用いてスキャンした。局所的バックグラウンドを差し引いた後に、二重のスポットからの平均シグナル強度を、ハウスキーピング遺伝子について得られた値を用いてプローブ標識における差について標準化した(Bernardら, 2002)。各遺伝子について、OSMにより誘発された変調は、刺激したサンプル対コントロールサンプルについての相対的発現値として表した。任意に、2を超える変調は、RT-リアルタイムPCRアッセイを用いる確認について著しいとみなした。
【0044】
ウェスタンブロッティング分析
STAT3及びMAPKのリン酸化のために、NHEKを、記載するサイトカインの存在下で15分間刺激した。細胞を、SDSサンプルバッファー(62.5 mM Tris-HCl pH 6.8、2% SDS、10%グリセロール、50mM DTT、0.1%ブロモフェノールブルー)中で溶解し、超音波破砕し、次いでSDS-PAGEに供してImmobilonメンブレンに移した。メンブレンは、その後、一次抗体と一晩インキュベートした後に適切なペルオキシダーゼ標識二次抗体と60分間インキュベートした。反応は、製造業者の使用説明に従う化学発光により視覚化した。メンブレンを、0.1 Mグリシン、pH 2.8中で抗体をはがし、1 M Tris-HCl、pH 7.6中で中和した後に再ブロットした。中和実験のために、NHEKは、適切な抗体と2時間インキュベートした後に刺激した。
【0045】
gp130、LIFR及びOSMRの発現を決定するために、細胞を10 mM Tris HCl pH 7.6、5 mM EDTA、50 mM NaCl、30 mMピロリン酸ナトリウム、50 mMフッ化ナトリウム、1 mMオルトバナジン酸ナトリウム、プロテイナーゼ阻害剤及び1% Brij 96中で溶解した。溶解及び遠心分離により細胞破片を除去した後に、上清を上記のようにして処理した。
【0046】
S100タンパク質発現のために、NHEKをOSM (10 ng/ml)の存在下に2日間刺激した。細胞溶解は、50 mM Tris HCl pH 7.4、150 mM NaCl、1 mM EDTA、1% Triton、1%デオキシコール酸ナトリウム、0.1 % SDS、1 mM PMSF、1 mMオルトバナジン酸ナトリウム、1%プロテア
ーゼ阻害剤を用いて行った。S100A7、S100A8及びS100A9は、上記のようにして免疫化学により検出した。ポンソーレッド染色を用いて、ロード量の均一性を制御した。
【0047】
インビトロケラチノサイト移動アッセイ
ケラチノサイトを、I型コラーゲン(200μg/ml, Institut Jacques Boy, Reims, France)で予めコートしたウェル中で、80 %の集密性に到達するまで培養した。細胞を、Keratinocyte SFM中で48時間枯渇させ、10μg/mlのマイトマイシンC (Sigma)で2時間処理して細胞増殖を妨げた。無細胞領域は、ケラチノサイトの単層をプラスチックのピペットチップでかきとることにより作製した。無細胞領域へのケラチノサイトの移動は、EGF又はOSMの存在下若しくは非存在下での48時間の培養の後に評価した。倒立位相差顕微鏡を用いる。移動ケラチノサイトの数は、4つの非重複領域において計数した。値は、インビトロ損傷の境界(frontiers)を超えるmm2当たりの細胞の平均±SEMを表す。スチューデントのt検定を、統計学的分析に用いた。
【0048】
再構成されたヒト表皮モデル
組織学的及び免疫組織化学的な研究のために、空気‐培地の界面で12日間増殖させたRHEを、SkinEthic Laboratories (Nice, France)から購入した。これらは、多層表皮からなり、これはヒトの皮膚に類似した形態学的及び成長の特徴を示す(Rosdy, Bertinoら 1997)。推奨されるようにして、RHEをSkinEthic増殖培地中で1日間増殖させ、その後、OSMの存在下又は非存在下に4日間刺激した。次いで、これらを平衡10%ホルマリン溶液中で固定し、パラフィンに包埋した。4ミクロンの垂直切片を、ヘマトキシリン/エオシンで染色するか、又は特異的Abとペルオキシダーゼ結合抗体とで染色し、かつ標準的な方法に従ってヘマトキシリンで対比染色した(Rosdy, Bertinoら 1997)。抗-K10ケラチン及び抗フィラグリンモノクローナル抗体は、Lab Vision Corporation (Fremont, CA, USA)からであった。
【0049】
cDNAマクロアレイを用いる遺伝子発現プロファイリングのために、17日齢のRHEを、SkinEthic維持培地で1日増殖させた後に、OSMの存在下又は非存在下に24時間刺激した。トータルRNAを単離し、上記のようにしてcDNAアレイを行った。
【0050】
実施例2:ヒトケラチノサイトはその表面上にある型のOSMを発現した
正常なヒトケラチノサイトにおけるOSMの潜在的な機能を示すために、本発明者らは、まず、その受容体鎖発現の分析を行った。発現されたI型又はII型の受容体の性質を決定するために、gp130、LIFRβ及びOSMRβについてのRT-PCRを、ケラチノサイトの初代培養から開始して行った。CO-G-UVM及びグリア芽腫細胞は、LIFRについてのコントロールとして用いた。得られた結果は、NHEKはOSMR及びgp130についての転写産物を主に発現するが、LIFR鎖は低いレベルでしか証明できなかった(図1A)。RNA分析を、フローサイトメトリによる対応するタンパク質の発現レベルの測定によりさらに補強した。蛍光分析は、NHEK細胞表面でのgp130及びOSMRβの明確な発現を明らかにした(図1B)。対照的に、膜のLIFRβ発現は検出されなかったが、抗-LIFRβ抗体は、ポジティブコントロールとして用いた細胞系とインキュベートしたときに、予想される結果を示した。このことは、gp130及びOSMRβ鎖の検出、並びにNHEKにおけるLIFRβ発現の不在を示すウェスタンブロット分析によってさらに支持された(図1C)。4人の異なるドナーからのサンプルについて行われた類似の実験は、同様の結果に導き、ドナー間のI型又はII型のOSM受容体発現の多様性の可能性を除外した。これらの第一の結果は、ヒトケラチノサイトが特定のII型のOSM受容体を優先的に発現したことを示す。
【0051】
実施例3:ヒトケラチノサイトにおいて、STAT-3及びMAPキナーゼ経路はOSMにより補充される
本発明者らは、NHEKにおけるOSM誘発シグナル伝達を示す。STAT3は、通常、OSM II型受
容体経路により補充されるので、サイトカインの漸増濃度に応答するシグナル伝達分子のチロシンリン酸化を分析した。チロシンリン酸化の強い誘導がSTAT3について観察され、OSMが3 ng/mlまで下がってもプラトーのレベルの値がまだ存在していた(図2B)。gp130及びOSMRβサブユニットの関与が、OSM接触の前に受容体中和mAbをNHEK培養に加えたときにSTAT3リン酸化がブロックされることによりさらに証明された(図2C)。重要なことには、抗-OSMRβmAbの存在下で観察されたSTAT3リン酸化の完全な中和は、NHEKにおける共有(share) LIF/OSM I型受容体のOSMによる補充が存在しないことをさらに証明した。この観察に合致して、LIF接触後に、STAT3活性化の証明はNHEKにおいて観察できなかった。
【0052】
さらに、II型OSM受容体複合体は、共有LIF/OSM受容体に比べて、Mapキナーゼ経路の補充についてより強力な活性化物質であることも知られている。LIFRbではなくOSMRβを介して媒介されるERK1/ERK2とShcアダプタとの間の協力的な影響は、OSMに応答してのMAPキナーゼ経路のこの強い活性化を説明する(Boulton, Stahlら 1994)。NHEK におけるサイトカインに応答してのERK1/2シグナル伝達を、よって、それらのチロシンリン酸化レベルを決定することにより分析した。予想されたように、OSMを用いるNHEKの刺激は、MAPキナーゼリン酸化を迅速に増加させた(図2D)。まとめると、これらの結果は、ヒトケラチノサイトにおいて発現されるgp130/OSMRβ受容体複合体が完全に機能的であり、観察されたシグナル全体がII型受容体を介して媒介されることを示した。
【0053】
実施例4:OSMは、ケラチノサイト移動の強力な誘発物質である
OSMに対するNEHKの機能的応答を強調するために、本発明者らは、創傷治癒を模倣しかつケラチノサイト移動に基づくインビトロモデルに対するOSMの潜在的影響を分析した(Kira, Sanoら 2002)。培養開始から48時間後に、ウェルの中位(middle)に存在する細胞をかきとることにより回収し、残りのケラチノサイトを、ケラチノサイト移動を引き起こすことが知られている EGF、又はOSMのいずれかで刺激した。さらに36時間培養した後に、細胞移動を誘導するサイトカインの潜在性を、視覚的に又は細胞計数により決定した(図3)。得られた結果は、OSMが、EGFの存在下で観察されたことと同様に、NHEKの著しい移動を導いたことを示す。
【0054】
実施例5:ヒトケラチノサイトにおけるOSM誘発遺伝子発現の同定
NHEK機能的応答をよりよく理解するために、本発明者らは、cDNAアレイを用いて、OSMにより誘発されるケラチノサイト遺伝子発現プロフィールの改変を分析した。用いたアレイは、ケラチノサイト研究のために特別に設計され、二重にスポットされた586の異なるcDNAからなった。これらは、ケラチノサイトの細胞構造、代謝、細胞外マトリクス、接着、分化、シグナル伝達、シグナル変換、アポトーシス及びストレスに関係する遺伝子であった(Bernardら, 2002)。コントロール又はOSMで刺激したNHEKから抽出したRNAは、逆転写により、標識cDNAプローブを作製するのに用いた。これらのcDNAプローブでAtlas cDNAアレイメンブレンをプローブすることにより、OSMが36遺伝子の発現を増加させ、38遺伝子の発現を減少させたことが明らかになった。OSMは、ケラチノサイト分化に関連する多くの一連の遺伝子、例えばサイトケラチン(CK)1、CK10、フィラグリン及びロリクリン遺伝子をダウンレギュレートする。アップレギュレートされる遺伝子のうち、本発明者らは、カルシウム結合性タンパク質であるソリアシン(S100A7)、カルグラヌリン(S100A8、S100A9)及びS100好中性タンパク質について著しい増加を見出した(図4)。興味深いことに、これらのタンパク質の発現は、炎症性組織においてアップレギュレートされることが知られている(Madsen, Rasmussenら 1991; Nagase及びWoessner 1999; Roth, Voglら 2003)。OSMはまた、組織防御に関わる、Gタンパク質結合受容体であるHM74、スーパーオキシドジスムターゼ2及びβデフェンシン遺伝子を誘導した。マトリクスメタロプロテイナーゼ1及びテナシン(tenascin)のような組織リモデリングに関わる遺伝子も、OSMにより誘導された。さらに、OSMは、ケモカインCXCL1 (MIP-2α)、CXCL5 (上皮由来好中球活性化ペプチド(ENA 78))及びCXCL8 (IL-8)、並びに血小板由来成長因子A (PDGF-A)遺伝子の発現を増
加させた。
【0055】
得られた結果は、ヒトケラチノサイトにおいて、OSMが、炎症性プロセス及び先天免疫応答に関わるいくつかの遺伝子を補充できたことを示す。
【0056】
実施例6:OSMは、ケラチノサイトを誘発してソリアシン、カルグラヌリン、βデフェンシン及びケモカインを産生させた
設計されたアレイを用いて得られた結果をさらに強化するために、mRNA及びタンパク質レベルでの定量的分析を、選択された数の同定された遺伝子について行った。OSMに応答してのソリアシン/S100A7 mRNA発現の定量的分析を、動態及び用量応答の研究とともにRT-リアルタイムPCRにより行った。本発明者らは、ソリアシン/S100A7 mRNAが、48時間の処理の後に1.6〜6.3 ng/mlの範囲のOSMに応答して用量依存的な様式でアップレギュレートされ、6.3 ng/ml OSMについて、コントロールを超えてシグナルが50倍増加して、プラトーに達したことを示す(図5A)。動態研究は、10 ng/mlのOSMでの刺激の12時間後に、ソリアシン/S100A7 mRNA発現の増加が開始したことを明らかにした(図5B)。これは96時間まで増加し続け、コントロールの値を超える約290倍の強い誘導であった。このことは、ソリアシン/S100A7並びに2つの関連するカルシウム結合性タンパク質であるS100A8及びS100A 9カルグラヌリンのウェスタンブロット分析により、タンパク質レベルで確かめられた(図5D)。結果は、10 ng/mlのOSMにNHEKを曝すことにより、研究したタンパク質の発現が増加し、これは2日目よりも4日目のほうが強かったことを示す(図5D)。図5Cは、皮膚活性化の2つの重要なマーカーであるフィラグリン及びβデフェンシン-2のRNA定量的発現を分析することにより得られた結果を描写する。
【0057】
48時間のNHEKでのケモカインCXCL5及びCXCL8の産生も、OSM刺激の下で明確に増強される(図6B)。
【0058】
実施例7:OSMは、再構成されたヒト表皮の過形成を引き起こし、分化関連抗原(differentiation related antigens)の発現を変調する
表皮分化の原動力にさらに取り組むために、本発明者らは、基底細胞層増殖及び段階的表皮分化プロセスを評価するためにインビトロRHEに対するOSMの生物学的影響を試験した。コントロールRHEの組織学的分析は、インビボでの表皮に類似し、無損傷の基底棘状顆粒角化細胞層を含み、かつ上方顆粒層に多数のケラトヒアリン顆粒を含む角化多重層上皮を示した(図7)。OSMは、ケラチノサイト層の過形成を引き起こし、RHEの全体的な厚みの増加を導いた。さらに、顆粒層におけるケラトヒアリン顆粒の損失及び凝縮核(picnotic nuclei)の存在が観察された。RHEのcDNAアレイプロフィール分析は、NHEKについて以前に記載されたことと同様に、OSMがS100A7、S100A8、S100A9及びS100好中性タンパク質遺伝子を強くアップレギュレートしたことを確かにした(図4)。免疫組織化学により、OSMで処理したRHEにおけるS100A7タンパク質のアップレギュレーションを確認した(図7)。NHEKについてのデータに合致して、RHEに対するOSM処理も、CXCL5、CXCL8ケモカイン遺伝子、並びにPDGF-A及びカドヘリン3遺伝子の転写をアップレギュレートした。RHEに特異的でありNHEKでは検出されないが、OSMは、CK6A、6B、6D、7、13、14、16、皮膚由来抗ロイコプロテイナーゼ及びTGFβ-誘導性早期タンパク質をアップレギュレートした。
【0059】
一方、OSMはケラチノサイト分化に関連する遺伝子、例えばインボルクリン(involucrin)、フィラグリン及びカルモジュリン様皮膚タンパク質をダウンレギュレートする(Mehul, Bernardら 2000; Rogers, Kobayashiら 2001; Jonak, Klosnerら 2002; Wagener, van Beurdenら 2003)。RHE切片に対して行った免疫組織化学的分析は、OSMで処理したRHEにおけるフィラグリン及びケラチン10の発現の減少により示されるように(図7)、ケラチノサイトの分化の阻害を確かにした。
【0060】
実施例8:考察
cDNAアレイアプローチ、特にヒトの皮膚における遺伝子発現の分析のために設計されたcDNAアレイアプローチの使用により、ヒトケラチノサイトにおけるOSM標的遺伝子の同定、並びに移動及び分化を含む種々のプロセスにおけるOSMの関与の証明が可能になった。特に、NHEK及びRHEにおけるS100A7、S100A8及びS100A9タンパク質の発現の強く、用量応答性で、OSMにより媒介される誘導は、サイトカインの炎症誘発性及び走化性の効果を証明する。皮膚の炎症におけるIL-10及びOSMの対抗する効果は、S100A8のIL-10により誘発されるダウンレギュレーションと、単球によるS100A9放出とにより強調される(Lugering, Kucharzikら 1997)。S100A7、S100A8及びS100A9は、カルシウム結合タンパク質の多形質発現性S100ファミリーに属する(Roth, Voglら 2003)。それらの主要な機能は未だ明らかではないが、これらは顕著な炎症性機能を演じ (Watson, Leygueら 1998; Donato 1999; Roth, Voglら 2003)、かつ多数の細胞内及び細胞外の活動、例えば細胞骨格成分の運動性又は走化性の原動力の厳密な調節に関わるようである(Ryckman, Vandalら 2003)。興味深いことに、S100A7、S100A8及びS100A9の3つ全てのタンパク質が正常な皮膚表皮及び未分化の培養ケラチノサイトにおいて低いか又は検出不可能なレベルで発現されると報告されているが、これらは、異常に分化した乾癬ケラチノサイトにおいて(Broome, Ryanら 2003)、創傷修復の間(Thorey, Rothら 2001)、及び表皮皮膚腫瘍において(Watson, Leygueら 1998; Gebhardt, Breitenbachら 2002; Alowami, Qingら 2003)多く発現される。炎症性細胞、特に好中球及びCD4+ Tリンパ球に対するS100A7の走化性効果により、S100A7は乾癬外傷の形成に関与し得ることが示唆される(Watson, Leygueら 1998)。S100A7はこれらの機構の上流で作用するので、本発明者らは、OSMが病的な状態でのS100A7の誘導のための鍵となる分子であり、病的段階に関与することを証明している。OSMによるさらなる遺伝子の変調は、OSMの炎症誘発性及び走化性の役割に好ましいものでもある。実際に、皮膚における接着性相互作用及び細胞浸潤を減じることにより炎症を弱める好中性誘引物質ケモカインCXCL5/ENA-78の誘導及びヘムオキシゲナーゼ1のダウンレギュレーションは、皮膚における好中球の流入に貢献し得る(Koch, Kunkelら 1994; Wagener, van Beurdenら 2003)。
【0061】
OSMにより誘導されるMMP-3発現は、炎症性皮膚疾患及び創傷修復の関係においても興味深い。MMP-3は正常な皮膚においては検出不可能であるが、損傷の後に基底層の増殖性ケラチノサイトにより発現される(Pilcher, Wangら 1999)。多くの疾患の進行の間に、MMP-3は、組織再吸収の間の細胞外マトリクスの除去による表皮リモデリングに関わり(Nagase及びWoessner 1999; Pilcher, Wangら 1999)、MMP-3遺伝子を欠損するマウスは、表皮の創傷修復に欠陥がある(Bullard, Lundら 1999)。インビトロ創傷アッセイを用いて、本発明者らは、ケラチノサイト移動がOSM刺激により強く増大することを証明した。これらのデータは、ケラチノサイトにおけるSTAT3欠陥が移動を弱めることに導くことの証明と合致する(Sano, Itamiら 1999)。炎症性条件の下で、OSMは、MMP-3又はS100A8-S100A9依存性の機構を介してケラチノサイト移動及び創傷治癒を増強する必須のメディエイタの1つであると考えられる。創傷治癒におけるOSMの関与を確立するさらなる証拠は、皮膚修復プロセスの開始の間の結合組織についての主要な増殖性及び移動性の刺激物質であるPDGFのRHEにおける強い誘導である(Rollman, Jensenら 2003)。
【0062】
本発明者らは、OSMがRHEのケラチノサイト層の全体的な厚みを増加させることも示した。このプロセスは、基底細胞の過剰増殖とは関係しないようである。なぜなら、フィラグリン、ロリクリン又はインボルクリンの産生の減少により示されるように、Ki67発現がOSMに応答しては誘導されないが、終末のケラチノサイト分化の阻害に起因する可能性がより高いからである。OSMは、ケラチノサイト分化に直接関連する、カルモジュリンファミリーの2つのメンバーであるカルモジュリン様皮膚タンパク質(CLSP)及びカルモジュリン関連タンパク質NB-1の発現をダウンレギュレートする(Mehul, Bernardら 2001; Rogers, Kobayashiら 2001)。CLSPは、ロリクリン又はインボルクリンのようなタンパク質の形成及び組み立てに関係する鍵となる酵素であるトランスグルタミナーゼ-3に結合して、表皮の角化細胞エンベロープを形成する(Mehul, Bernardら 2000)。ケラチン発現プロフィール、すなわちケラチン6の過剰発現及びケラチン10の阻害に対するOSMの変調効果も、表皮分化の阻害の概念を支持する。ケラチン6は、創傷治癒、乾癬、発癌を含む過剰増殖及び/又は炎症性の状況の下で、或いは表皮過形成を引き起こすレチノイン酸のような作用物質により、誘導されることが知られている(Navarro, Casatorresら 1995; Komine, Raoら 2000)。対照的に、終末分化している表皮ケラチノサイトにおいて通常は発現しているケラチン10は、創傷治癒の間に減少される(Paramio, Casanovaら 1999)。
【0063】
実施例9:IL-31を用いて得られる同様の結果
実施例3、4及び5で示す研究を、OSMの代わりにIL-31を用いて行い、類似の結果を得た。IL-31は、NHEKにおいてSTAT3シグナル伝達経路を補充し(図8)、ソリアシン(S100A)及びカルグラヌリンA及びB (S100A8-9)の発現を誘発する(図9)。IL-31も、ケラチノサイト移動を誘発できる(図10)。
【0064】
実施例10:ガンマインターフェロンは、シグナル変換に対するIL-31の作用を強化する
NHEKを、IL-31刺激(50 ng/ml)の前に50 g/mlのガンマインターフェロン(INFγ)の存在下で24時間プレインキュベーションしたとき、P-STAT3のレベルは、培地のみの中でプレインキュベートしたNHEKに比べて3〜4倍増加した(図8について行ったものと同じ研究)。このことは、INFγがシグナル変換に対するIL-31の作用の修飾物質であることを証明する。
【0065】
実施例11:IL-17及びTNFαは、いくつかの炎症性マーカーの発現に対するOSMの作用を強化する
ケラチノサイトに対するいくつかのサイトカインの併用効果を、ケラチノサイト炎症マーカーであるソリアシン(S100A7)、デフェンシンβ-2/β-4 (hBD2/4) mRNAの発現を、種々のサイトカインカクテルの存在下で測定することにより試験した。これらのカクテルの影響を、ケラチン10 (KRT10) mRNAに対しても試験した。なぜなら、KRT10は組織治癒に関連する分化マーカーだからである。
【0066】
この目的のために、集密な正常ヒトケラチノサイト(NHEK)を、記載するサイトカイン混合物と24時間処理した(各サイトカインは1 ng/mlの最終濃度)。トータルRNAを抽出し、逆転写し、選択された遺伝子の発現を、記載するようにしてリアルタイムPCRにより分析した。
これらの結果を、以下の表1及び図11に示す。
【0067】
【表1】

【0068】
6つの選択されたサイトカインの混合物(M3)は、ケラチノサイト炎症マーカーであるソリアシン(S100A7)及びデフェンシンβ-2/β-4 (hBD2/4)の発現に対して強い相乗効果を示した。予想されたように、KRT10発現は、これらのサイトカインの存在下で低下する。IFNγ、IL-22又はIL-1αのいずれかを混合物から除去しても、サイトカインの完全なカクテルの活性を著しくは減少させなかった。よって、これらのサイトカインはおそらく、観察された相乗作用に直接は関与していない。逆に:
- 5種サイトカイン参照混合物(不活性であるのでIFNγを除外した)からOSMを除外すると、混合物の活性は、S100A7について3倍、及びhBD2/4について2.5倍減少された;
- TNFαを除外すると、混合物の活性は、S100A7について3倍、及びhBD2/4について5.6倍減少された;かつ
- IL-17を除外すると、混合物の活性は、S100A7について4.5倍、及びhBD2/4について36倍減少された。
【0069】
これらの結果は、ケラチノサイトにおける最大限の応答について、OSM、TNFα及びIL-17の間の強い相乗作用を示す。
よって、OSMは、IL-17及び/又はTNFαとともに相乗作用を示すことができると考えられる。さらに、OSMは、低いサイトカイン濃度であっても、創傷再上皮形成促進のためにIL-17及び/又はTNFαと相乗的に作用できることが示唆される。
【0070】
実施例12:OSMは、皮膚先天性免疫応答を改善する
乾癬患者の皮膚におけるケラチノサイトは、高レベルの抗菌性ペプチドを産生する(Nomura, Golevaら 2003)。正常な皮膚又はアトピー性皮膚炎の患者の皮膚にはごくわずかなレベルでしか存在しないこれらのペプチドは、感染の動物モデルにおいて示されるように、侵入微生物に対する皮膚先天性免疫応答において必須である(Nizet, Ohtakeら 2001)。S100A7-ソリアシンは、大腸菌(Escherichia coli)による皮膚の感染に対する耐性を与えることが示されているので(Glaser, Harderら 2005)、OSMを用いた刺激の後のケラチノサイトにおけるS100A7-ソリアシン遺伝子発現の増強は、皮膚の先天性免疫応答におけるOSMについての役割を指摘する。同様に、抗菌性活性を有する別のタンパク質であるβ-デフェンシン2 (Harder, Bartelsら 1997)は、ヒトケラチノサイトにおいてOSMにより誘導される。β-デフェンシン2は、乾癬外傷においては高いレベルで発現されるが、アトピー性皮膚炎において観察されるものにおいては低いレベルでしか発現されない(Ong, Ohtakeら 2002)。この観察と調和して、乾癬の患者は、アトピー性皮膚炎を有する患者に比べると、感染に対して感受性がより低いと報告されている(Nomura, Golevaら 2003)。
よって、上記の実施例6及び11で報告する結果は、OSM、IL-17及びTNFαがケラチノサイトの有効な活性化物質であり、これらのサイトカインの単独又は組み合わせでの局所投与が、抗菌性ペプチドの発現を増強し、かつ皮膚先天免疫を改善できることを示す。
【0071】
参考文献
Alowami, S., G. Qing,ら (2003). "Psoriasin (S100A7) expression is altered during
skin tumorigenesis." BMC Dermatol 3(1): 1.
Benigni, F., G. Fantuzzi,ら (1996). "Six different cytokines that share GP130 as
a receptor subunit, induce serum amyloid A and potentiate the induction of interleukin-6 and the activation of the hypothalamus-pituitary-adrenal axis by interleukin-1." Blood 87(5): 1851-4.
Bernard, F. X., N. Pedretti,ら (2002). "Comparison of gene expression profiles in human keratinocyte mono-layer cultures, reconstituted epidermis and normal human skin; transcriptional effects of retinoid treatments in reconstituted human epidermis." Exp Dermatol 11(1): 59-74.
Bonifati, C., A. Mussi,ら (1998). "Spontaneous release of leukemia inhibitory factor and oncostatin-M is increased in supernatants of short-term organ cultures
from lesional psoriatic skin." Arch Dermatol Res 290(1-2): 9-13.
Boulton, T. G., N. Stahlら (1994). "Ciliary neurotrophic factor/leukemia inhibitory factor/interleukin 6/oncostatin M family of cytokines induces tyrosine phosphorylation of a common set of proteins overlapping those induced by other cytokines and growth factors." J Biol Chem 269(15): 11648-55.
Broome, A. M., D. Ryan,ら (2003). "S100 protein subcellular localization during epidermal differentiation and psoriasis." J Histochem Cytochem 51(5): 675-85.
Bullard, K. M., L. Lund,ら. (1999). "Impaired wound contraction in stromelysin-1-deficient mice." Ann Surg 230(2): 260-5.
【0072】
Dillon, S. R., C. Sprecher,ら (2004). "Interleukin 31, a cytokine produced by activated T cells, induces dermatitis in mice." Nat Immunol 5(7): 752-60.
Donato, R. (1999). "Functional roles of S100 proteins, calcium-binding proteins of the EF-hand type." Biochim Biophys Acta 1450(3): 191-231.
Gallucci, R. M., P. P. Simeonova,ら (2000). "Impaired cutaneous wound healing in
interleukin-6-deficient and immunosuppressed mice." Faseb J 14(15): 2525-31.
Gallucci, R. M., D. K. Sloan,ら (2004). "Interleukin 6 indirectly induces keratinocyte migration." J Invest Dermatol 122(3): 764-72.
Gebhardt, C., U. Breitenbach,ら (2002). "Calgranulins S100A8 and S100A9 are negatively regulated by glucocorticoids in a c-Fos-dependent manner and overexpressed throughout skin carcinogenesis." Oncogene 21(27): 4266-76.
Glaser, R., J. Harder,ら (2005). "Antimicrobial psoriasin (S100A7) protects human skin from Escherichia coli infection." Nat Immunol 6(1): 57-64.
Harder, J., J. Bartels,ら (1997). "A peptide antibiotic from human skin." Nature
387(6636): 861.
【0073】
Ihn, H.及びK. Tamaki (2000). "Oncostatin M stimulates the growth of dermal fibroblasts via a mitogen-activated protein kinase-dependent pathway." J Immunol 165(4): 2149-55.
Jonak, C., G. Klosner,ら (2002). "Subcorneal colocalization of the small heat shock protein, hsp27, with keratins and proteins of the cornified cell envelope." Br J Dermatol 147(1): 13-9.
Kira, M., S. Sano,ら (2002). "STAT3 deficiency in keratinocytes leads to compromised cell migration through hyperphosphorylation of p130(cas)." J Biol Chem 277(15): 12931-6.
Koch, A. E., S. L. Kunkel,ら (1994). "Epithelial neutrophil activating peptide-78: a novel chemotactic cytokine for neutrophils in arthritis." J Clin Invest 94(3): 1012-8.
Komine, M., L. S. Rao,ら (2000). "Inflammatory versus proliferative processes in
epidermis. Tumor necrosis factor alpha induces K6b keratin synthesis through a transcriptional complex containing NFkappa B and C/EBPbeta." J Biol Chem 275(41): 32077-88.
【0074】
Lugering, N., T. Kucharzik,ら (1997). "Importance of combined treatment with IL-10 and IL-4, but not IL-13, for inhibition of monocyte release of the Ca(2+)-binding protein MRP8/14." Immunology 91(1): 130-4.
Madsen, P., H. H. Rasmussen,ら (1991). "Molecular cloning, occurrence, and expression of a novel partially secreted protein "psoriasin" that is highly up-regulated in psoriatic skin." J Invest Dermatol 97(4): 701-12.
Mehul, B., D. Bernard,ら (2001). "Calmodulin-like skin protein: a new marker of
keratinocyte differentiation." J Invest Dermatol 116(6): 905-9.
Mehul, B., D. Bernard,ら (2000). "Identification and cloning of a new calmodulin-like protein from human epidermis." J Biol Chem 275(17): 12841-7.
Nagase, H.及びJ. F. Woessner, Jr. (1999). "Matrix metalloproteinases." J Biol Chem 274(31): 21491-4.
【0075】
Navarro, J. M., J. Casatorres,ら (1995). "Elements controlling the expression and induction of the skin hyperproliferation-associated keratin K6." J Biol Chem 270(36): 21362-7.
Nizet, V., T. Ohtake,ら (2001). "Innate antimicrobial peptide protects the skin from invasive bacterial infection." Nature 414(6862): 454-7.
Nomura, I., E. Goleva,ら (2003). "Cytokine milieu of atopic dermatitis, as compared to psoriasis, skin prevents induction of innate immune response genes." J Immunol 171(6): 3262-9.
Ong, P. Y., T. Ohtake,ら (2002). "Endogenous antimicrobial peptides and skin infections in atopic dermatitis." N Engl J Med 347(15): 1151-60.
Paglia, D., S. Kondo,ら (1996). "Leukaemia inhibitory factor is expressed by normal human keratinocytes in vitro and in vivo." Br J Dermatol 134(5): 817-23.
【0076】
Paramio, J. M., M. L. Casanova,ら (1999). "Modulation of cell proliferation by cytokeratins K10 and K16." Mol Cell Biol 19(4): 3086-94.
Pattyn, F., F. Speleman,ら (2003). "RTPrimerDB: the real-time PCR primer and probe database." Nucleic Acids Res 31(1): 122-3.
Pfaffl, M. W. (2001). "A new mathematical model for relative quantification in real-time RT-PCR." Nucleic Acids Res 29(9): e45.
Pilcher, B. K., M. Wang,ら (1999). "Role of matrix metalloproteinases and their inhibition in cutaneous wound healing and allergic contact hypersensitivity." Ann N Y Acad Sci 878: 12-24.
Rogers, M. S., T. Kobayashi,ら (2001). "Human calmodulin-like protein is an epithelial-specific protein regulated during keratinocyte differentiation." Exp Cell
Res 267(2): 216-24.
Rollman, O., U. B. Jensen,ら (2003). "Platelet derived growth factor (PDGF) responsive epidermis formed from human keratinocytes transduced with the PDGF beta receptor gene." J Invest Dermatol 120(5): 742-9.
【0077】
Rosdy, M., B. Bertino,ら (1997). "Retinoic acid inhibits epidermal differentiation when applied topically on the stratum corneum of epidermis formed in vitro by
human keratinocytes grown on defined medium." In Vitro Toxicology 10(1): 39-47.Roth, J., T. Vogl,ら (2003). "Phagocyte-specific S100 proteins: a novel group of
proinflammatory molecules." Trends Immunol 24(4): 155-8.
Ryckman, C., K. Vandal,ら (2003). "Proinflammatory activities of S100: proteins S100A8, S100A9, and S100A8/A9 induce neutrophil chemotaxis and adhesion." J Immunol 170(6): 3233-42.
Sano, S., S. Itami,ら (1999). "Keratinocyte-specific ablation of Stat3 exhibits impaired skin remodeling, but does not affect skin morphogenesis." Embo J 18(17): 4657-68.
Sugawara, T., R. M. Gallucci,ら (2001). "Regulation and role of interleukin 6 in
wounded human epithelial keratinocytes." Cytokine 15(6): 328-36.
【0078】
Taga, T.及びT. Kishimoto (1997). "Gp130 and the interleukin-6 family of cytokine
s." Annu Rev Immunol 15: 797-819.
Thorey, I. S., J. Roth,ら (2001). "The Ca2+-binding proteins S100A8 and S100A9 are encoded by novel injury-regulated genes." J Biol Chem 276(38): 35818-25.
Vandesompele, J., K. De Preter,ら (2002). "Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes." Genome Biol 3(7): RESEARCH0034.
Wagener, F. A., H. E. van Beurden,ら (2003). "The heme-heme oxygenase system: a molecular switch in wound healing." Blood 102(2): 521-8.
Wahl, A. F.及びP. M. Wallace (2001). "Oncostatin M in the anti-inflammatory response." Ann Rheum Dis 60 Suppl 3: iii75-80.
Watson, P. H., E. R. Leygue,ら (1998). "Psoriasin (S100A7)." Int J Biochem Cell Biol 30(5): 567-71.
【図面の簡単な説明】
【0079】
【図1】NHEKによるOSM受容体の発現を示す。
【図2】NHEKでのOSMによるSTAT3及びMAPキナーゼのリン酸化の誘導を示す。
【図3】ケラチノサイト移動に対するOSMの影響を示す。
【図4】OSMで刺激したNHEK及びOSMで処理したRHEから得られる発現プロフィールを示す。
【図5】NHEKによるS100A7〜9合成に対するOSMの影響を示す。
【図6】NHEKによるサイトカイン及びケモカインの産生を示す。
【図7】10 ng/mlのOSMを用いて4日間刺激したか又はしなかったRHEの組織学的及び免疫組織化学的分析を示す。
【図8】NHEKにおけるIL-31及びOSMによるSTAT3及びMAPキナーゼリン酸化の誘導を示す。
【図9】S100A7 mRNA発現に対するIL-31及びOSMの影響を示す。
【図10】ケラチノサイト移動に対するIL-31及びOSMの影響を示す。
【図11】S100A7、hBD4/2及びKRT10 mRNA発現に対するいくつかのサイトカインのカクテルの影響を示す。
【技術分野】
【0001】
本発明は、表皮修復(repair)の分野に関する。より具体的には、本発明は、インビトロ及び/又はインビボのケラチノサイト移動(migration)を増強するための組成物の製造のための、サブユニットとしてOSMRβを含むヘテロマー受容体を活性化できる分子の使用に関する。
【背景技術】
【0002】
皮膚は、生体とその環境との間の保護的界面を提供する大きく複雑な組織である。表皮は、その外表面を形成し、ケラチノサイトと名付けられた特殊化された上皮細胞の複数の層から主に構成されている。皮膚は、微生物、化学物質、動作、物理的損傷、加齢、U.V.照射、癌、自己免疫疾患又は炎症性疾患を含む多くの異なる原因により損傷され得る。
【0003】
表皮の恒常性は、皮膚の基底層から角化層に分化するケラチノサイトの分化と増殖とのバランスにより調節される。表皮ストレスに応答して、又はある皮膚疾患において、この平衡が破壊される。ケラチノサイトは、表皮成長因子(Epidermal Growth Factor (EGF))ファミリーのメンバーのような可溶性のメディエイタ、並びにさらなる成長因子及びサイトカイン(FGFs、IGF-1、PDGF、HGF、TGFβファミリーのメンバー、GM-CSF、TSLP、IL-1、TNF-α)に対して異なる応答ができるようになる。これらの修飾物質は、ケラチノサイト自体、皮膚繊維芽細胞、ランゲルハンス細胞により産生されるか、又はTリンパ球のような免疫浸潤細胞により産生される。これに応答して、ケラチノサイトはさらなるシグナル伝達分子を放出し、細胞表面受容体の発現レベルを変調し、それらの細胞骨格形態を修飾して、それらの移動、分化及び増殖の能力を変調する。これらの変化は炎症反応に関連し、創傷治癒又は慢性疾患のいずれかに導く。
【0004】
IL-6ファミリーのサイトカインは、多数の生体のシステム、例えば免疫、造血、神経発達、生殖、骨モデリング及び炎症プロセスにおいて細胞の成長及び分化を調節する多機能タンパク質である。このサイトカインファミリーは、9つの異なるメンバーを含む:IL-6、IL-11、IL-27、白血病抑制因子(LIF)、カルジオトロフィン(cardiotrophin)-1、カルジオトロフィン様因子、繊毛様神経栄養因子、ニューロポエチン(neuropoietin)、及びオンコスタチンM (OSM)である。これらのサイトカインの活性は、ダイマー又はトリマーの受容体複合体のリガンド誘発性オリゴマー化を介して媒介される。サイトカインのIL-6ファミリーは、それらの各ヘテロマー受容体の形成においてgp130受容体サブユニットを共有している(Taga及びKishimoto 1997)。IL-31と名付けられた最近記載されたサイトカインは、Dillonらによってgp130-IL6ファミリーの新しいメンバーとして分類された。なぜなら、その受容体は、gp130様I型サイトカイン受容体(GPL)及びOSMRサブユニットを含むヘテロダイマーであるからである(Dillon, Sprecherら 2004)。
【0005】
いくつかの文献は、IL-6ファミリーのいくつかのメンバーがある皮膚疾患及び創傷治癒プロセスに加わっているであろうことを報告している。IL-6、IL-11、LIF及びOSMが、乾癬外傷において増加することが見出されており(Bonifati, Mussiら 1998)、IL-6及びLIFは、精製ケラチノサイトから産生される(Paglia, Kondoら 1996; Sugawara, Gallucciら 2001)。創傷治癒プロセスの欠陥が、IL-6及びSTAT3欠損マウスにおいて報告されている(Sano, Itamiら 1999; Gallucci, Simeonovaら 2000)。しかし、IL-6欠損マウスから単離された培養ケラチノサイトでのさらなる研究は、ケラチノサイト移動に対するIL-6の作用が、皮膚繊維芽細胞により媒介されることを示した。実際に、IL-6単独では、該IL-6欠損ケラチノサイトの増殖又は移動を著しく変調はしなかったが、IL-6は、皮膚繊維芽細胞と同
時培養したときに、その移動を著しく誘発した(Gallucci, Sloanら 2004)。
【0006】
OSMは、活性化されたT細胞、サイトカインにより刺激された単球及び樹状細胞から分泌される。OSMは、炎症誘発性メディエイタであり、肝細胞においてタンパク質合成を強く引き起こす(Benigni, Fantuzziら 1996)。ヒトにおいては、OSM及びLIFは、成長調節の増加、分化、遺伝子発現、細胞生存により、いくつかの組織において重複した生物学的機能を示す。OSMは、LIFと共有していないいくつかの独特の生物学的機能、例えばいくつかの腫瘍細胞系統の成長阻害又はAIDS関連カポジ肉腫細胞の刺激を惹起することも知られている。OSMのこれらの共通及び独特の機能は、OSM受容体複合体が2種類存在することにより説明される。gp130/LIFRβサブユニットで形成される通常のLIF/OSM受容体複合体の他に、OSMは、gp130をOSMRβ(「OSMR」又は「OSM-R」ともよばれる)と結合させるII型受容体を特異的に認識でき、これは内皮細胞、肝細胞、肺細胞、繊維芽細胞、造血細胞及びいくつかの腫瘍細胞系統により発現される。その後のシグナル伝達カスケードは、ヤヌスキナーゼ(JAK 1、JAK 2、Tyk 2)の活性化を含み、その後、シグナル伝達性転写因子(STAT1、STAT3)及びMapキナーゼ経路の活性化が続く。
【0007】
その抗腫瘍性活性及び炎症誘発性反応におけるその役割に加えて(Wahl及びWallace 2001; Shoyabら, US 5,451,506; Richardsら, US 5,744,442)、OSMは、MAPキナーゼ依存性経路により皮膚繊維芽細胞の成長を刺激して、皮膚の創傷治癒を促進すると記載されている(Ihn及びTamaki 2000)。
【0008】
他のサイトカインも、皮膚に対して影響があることが知られている。例えば、Dillonら(上記)は、IL-31の過剰発現が、アレルギー性及び非アレルギー性の疾患を特徴付ける皮膚炎及び上皮反応の促進に関与し得ることを示唆する。これらの著者らは、IL-31を皮膚修復促進のために用いることを示唆していない。
【0009】
皮膚が損傷したときに、その完全な修復は、真皮及び表皮の両方が修復されることを包含する。異なる細胞の種類を含む表皮及び真皮の治癒(healing)は、異なる機構を伴う。
【0010】
現在、皮膚の治癒を改善するための治療は、主に真皮を標的としている。しかし、表皮の再構成が、完全な回復のために必要である。例えば大きい熱傷、潰瘍又は褥瘡の場合は、生理的表皮治癒プロセスは、皮膚の防御機能を回復するために充分に効果的ではない場合がある。このような場合には、損傷を受けた領域を迅速に覆い、感染及び乾燥の可能性を回避することが必要である。表皮の再生を刺激することも必要である。体表面の半分未満の重篤な熱傷の場合には、焼けた皮膚の切除の後に自己移植が行われる。その目的のために、患者から健全な皮膚が取られ、その表面積を増やすために機械的に処理される。この「ウィック(wick)皮膚」は、次いで、損傷の上に移植される。熱傷の表面積が大きすぎる場合(体表面の半分を超える)、このプロセスは実行不可能である。そこで、乾燥及び感染を回避するために創傷を一時的に覆うことが必要である。このことは、現在、死体からの皮膚又は創傷治癒を改善するために成長因子を組み込んでもよい無細胞ドレッシング、例えばツルグラのような皮膚代替物のいずれかを用いて行われている。このような皮膚代替物の例は、US 6,132,759又はWO 01/41820に記載される。並行して、患者からの皮膚細胞を、上皮層を得てこれを次いで移植するためにインビトロで増殖させる。数cm2の健全な皮膚から、1〜2 m2が3週間で得られる。しかし、これらの技術は、長期間で費用がかかり、そして成功して行うためには重大なインフラストラクチャーが必要である。
【発明の開示】
【発明が解決しようとする課題】
【0011】
よって、表皮の修復を改善するための新規な皮膚科学的アプローチが真に必要とされていることが明らかである。ケラチノサイトの求心的移動(centripetal migration)を増強することは、治癒及び再上皮形成を明らかに促進/可能にするであろう。ケラチノサイトの分化及び移動に対して作用することは、表皮水疱症のような特定の疾患の治療のためにも必要である。「表皮水疱症(bullous emolysis)」の用語は、表皮の剥離及び真皮と表皮の間の液体の蓄積に導く、起源が異なる(例えば出血、熱傷、自己免疫疾患など)いくつかの皮膚炎のことをいう。表皮水疱症の具体例は、水疱性類天疱瘡である。
【0012】
最近の小さい創傷又は古い切傷、斑、伸展裂創(stretch marks)などのいずれかに由来する瘢痕の外観(aspect)を改善するための効果的な組成物が存在しない化粧分野においても、表皮修復を改善することは重要である。
【0013】
この関係において、本発明者らは、いくつかのサイトカイン、特にOSM及びIL-31が、ケラチノサイトの移動を増強できることを見出した。興味深いことに、これらの2種のサイトカインは、ともにサブユニットとしてOSMRβを含む異なるヘテロマー受容体に結合する。本発明者らは、通常のヒト表皮ケラチノサイトが、gp130、GPL及びOSMRβを発現することを示している。
【0014】
以下の実験実施例において開示されるように、OSMは、ヒト表皮ケラチノサイトにおいて、STAT3シグナル伝達経路及びMAPキナーゼ経路を補充する(recruit)。OSMは、これらの細胞において、ケモカイン、デフェンシン及びソリアシン(psoriasin)を含む炎症誘発性遺伝子の発現をアップレギュレートする。OSMは、また、再構成されたヒト表皮の厚みを増加させ、一連の分化抗原をダウンレギュレートする。興味深いことに、他のサイトカイン、特にIL-17及びTNFαは、OSMと相乗的に作用し、その効果を強化する。
【0015】
本発明者らが行った実験はまた、IL-31がケラチノサイト移動を媒介することも明らかにした。しかし、本発明者らは、グリア芽腫及び黒色腫の腫瘍細胞において、IL-31の作用が、ヘテロマー受容体の形成におけるOSMRβに含まれるGPLサブユニットの種類に依存することを観察した。特に、本発明者らは、GPL受容体の短い形がIL-31のシグナル伝達に対して著しい阻害効果を発揮し、優性の負の受容体としてふるまうことに注目した。
【0016】
これらの結果を合わせると、OSM及びIL-31が、創傷治癒及びケラチノサイト移動を伴う他のプロセスにおいて重要な役割を演じることが示される。これらの効果は、ヘテロマー受容体のOSMRβサブユニットが参加するシグナル変換を必要とする。
【課題を解決するための手段】
【0017】
本発明の第一の目的は、よって、サブユニットとしてOSMRβを有するヘテロマー受容体の少なくとも1つの活性化物質の、表皮修復を改善するための組成物の製造のための使用である。
【発明を実施するための最良の形態】
【0018】
このような活性化物質の例は、OSMRβサブユニットを含むヘテロマー受容体のリガンド、及びそのアゴニストである。本発明による好ましい活性化物質は、サイトカイン、特にオンコスタチンM (OSM)及びインターロイキン31 (IL-31)、並びにそれらのアゴニストである。以下において、「OSM」及び「IL-31」は、サイトカイン自体、並びにそれらの各アゴニストのことをいうのに用いる。本発明による活性化物質のその他の例は、OSMRβとともにヘテロマー受容体を形成できる受容体サブユニットの発現をアップレギュレートする分子である。
【0019】
OSMの作用を強化するIL-17及びTNFα、並びにIL-31の効果を強化するガンマインターフェロン(IFN-γ)も、本発明による活性化物質と考えることができ、それぞれOSM又はIL-31と組み合わせて、それらの効果を増大し、それにより本発明による組成物中でより低い濃度で用いることを可能にするために、有利に用いることができる。もちろん、これらのサイトカインは、それらのアゴニストで置き換えることができる。変異サイトカイン、サイトカインではない天然又は変異のペプチド、及び非ペプチドの合成又は天然の分子は、本発明による活性化物質として用いることができる。
【0020】
本発明の好ましい実施形態によると、OSM、IL-17及びTNFαは、表皮修復を改善するための組成物の製造のために用いられる。
本発明により得られる組成物は、ケラチノサイト移動の促進、又は表皮治癒の促進のために用いられる。これらの組成物は、大きい創傷の場合に、ケラチノサイトの求心的移動の刺激に有利に用いられる。これらは、表皮水疱症の予防、減弱又は治療のために用いることもできる。実際に、その起源及び段階に応じて、表皮水疱症は、ケラチノサイト移動を増強する分子、又は逆に抗炎症性分子のいずれかを投与することにより治療できる。医師は、状況に応じて、表皮水疱症を罹患した患者にとって、OSM、IL-31及び/又はそれらのアゴニスト並びに強化物質を用いる処置がいつ効果的であり得るか、理解するであろう。
【0021】
本発明による組成物は、例えば局所投与によりインビボで、又はインビトロでのいずれかで表皮の厚みを増加させるために用いて、(ヒトの)再構成される表皮の質及び/又は量を増加させ、例えば移植の必要がある患者のために上皮層の生成を促進することもできる。
【0022】
本発明の特定の実施形態によると、サブユニットとしてOSMRβを有するヘテロマー受容体の活性化物質、又は該受容体を活性化する分子の組み合わせは、手、口唇、顔若しくは体のひび割れを予防するか及び/又は減じるため、或いは伸展裂創を予防するか及び/又は減じるための組成物の製造のために用いられる。本発明により得られる組成物の別の適用は、瘢痕の外観及び快適さ(comfort)の改善、及び/又は表皮創傷の治癒の間の表皮創傷の外観及び快適さの改善である。本発明のこの態様によると、創傷はいずれの起源でもあり得る。創傷は、切傷、熱傷、擦過傷などのような外傷によるもの、外科切開及び皮膚移植のような外科的手法によるもの、並びににきび、アトピー性皮膚炎、湿疹、職業上の皮膚炎、脂漏性皮膚炎、酒さ、紅斑、焼痂、糖尿病(足)、角化上皮症、鱗状化(squama)、潰瘍、魚鱗癬(ichtyosis)、表皮水疱症、足穿孔、疣贅、らい病感染(leprae infection)などのような障害及び疾患によるものを含む。もちろん、炎症性疾患又はケラチノサイト過剰増殖に関連する疾患による創傷の場合、医師は、本発明による組成物を投与する前に該疾患が治療されるように制御するであろう。
【0023】
本発明により製造される組成物は、局所投与用に製剤化されることが好ましい。これらは、例えばクリーム、ローション、軟膏又はドレッシングの形であり得る。
【0024】
本発明による具体的な化粧用及び/又は皮膚用組成物は、OSM又はIL-31を含む。
【0025】
本発明は、また、皮膚用及び/又は化粧用組成物の製造のための、上記の少なくとも2種のサイトカインの使用にも関する。これらの少なくとも2種のサイトカインは、同じ組成物中に混合されるか、又は部分品(parts)のキットで提供されるかのいずれかであり得る。本発明のこの態様によると、好ましい部分品のキット又は組成物は、OSMと、IL-17、TNFα及びこれらのサイトカインのアゴニストから選択される少なくとも1種のサイトカインとを含む。本発明による好ましい化粧用及び/又は皮膚用組成物は、OSM、IL-17及びTNFαを含む。本発明による組成物の別の具体的な実施形態において、上記の組成物は、IL-31及びIFN-γ、及び/又はOSMを含む。
【0026】
OSMを含む本発明による組成物において、OSMは、好ましくは0.1〜100 ng/ml、より好ま
しくは1〜20 ng/mlの濃度である。この濃度は、IL-17及び/又はTNFαの存在下で0.01 ng/mlまで減じることができる。このような複合組成物において、OSM及び/又はIL-17及び/又はTNFαの濃度は、好ましくは0.01 ng/ml〜10 ng/ml、より好ましくは0.1〜2 ng/mlである。IL-31を含む組成物において、後者は、好ましくは1〜500 ng/ml、より好ましくは10〜100 ng/mlの濃度である。
【0027】
上記のように、培養したケラチノサイトに、サブユニットとしてOSMRβを有するヘテロマー受容体の少なくとも1種の活性化物質を加えることにより、(ヒトの)再構成される表皮の質及び/又は量を増加させることができる。よって、本発明は、上記の少なくとも1種の活性化物質を、培養にあるケラチノサイトに加える工程を含む、再構成されたヒト表皮を製造する方法にも関する。例えば、この方法は、OSM及び/又はIL-17及び/又はTNFαを、該ケラチノサイトの培養培地に、各サイトカインについて好ましくは0.01〜10 ng/mlの範囲の濃度で加えることにより行うことができる。
【0028】
本発明は、以下の図面及び実施例によりさらに説明される。
図の説明
図1は、NHEKによるOSM受容体の発現を示す。
(A) トータルRNAを、4人の別個のドナーのNHEKから抽出した。RT-PCRを、OSMR、gp130、LIFR及びGAPDH遺伝子についての特異的プライマーを用いて行った。cDNAの連続希釈を増幅して、転写産物の発現レベルの半定量的分析を行った。PCR産物を、アガロースゲル電気泳動により分析した。(B) NHEK細胞表面の免疫標識及びフラックスサイトメトリー分析。Gp130及びOSMRは、細胞上で明確に検出されるが、LIFRは検出されない。(C) NHEK及びグリア芽腫細胞系統GO-G-UVMからの細胞溶解物20μgを、SDS-PAGE (10%)により分離し、ニトロセルロースメンブレンに移した。ポンソーレッド染色を用いて、ロード量(loading)の均一性を制御した。gp130、OSMR、LIFR及びチューブリンのバンドの検出は、ウェスタンブロットにより評価した。結果は、3つの独立した実験の代表である。
【0029】
図2は、NHEKでのOSMによるSTAT3及びMAPキナーゼのリン酸化の誘導を示す。
(A) NHEKは、LIF若しくはOSM (50 ng/ml)で刺激したか、又はしなかった。(B) NHEKは、15分間、50 ng/mlのIL-5 (ネガティブコントロール)又は100、50、25、12.5、6.25若しくは3.1 ng/mlのOSMで刺激したか、又は刺激せず、ホスホ-STAT3 (P-STAT3)及びSTAT3タンパク質のレベルをウェスタンブロットにより評価した。サイトカインを用いた刺激の前に、細胞を、中和抗体、抗-gp130 (AN-HH1)又は抗-OSMR (XR-M70)モノクローナル抗体の存在下で、又はアイソタイプコントロールmAb MC192 (最終抗体濃度15μg/ml)とともに、2時間インキュベートした(C) 。OSMに応答してのホスホ-MAPK (P-MAPK)及びMAPKタンパク質レベルは、ウェスタンブロットにより評価した(D)。
【0030】
図3は、ケラチノサイト移動に対するOSMの影響を示す。
インビトロ創傷をマイトマイシンで処理した集密なNHEK培養物に導入し、ケラチノサイトを10 ng/mlのEGF若しくはOSMとともに、又は用いずに48時間培養した。無細胞領域への細胞移動を、材料及び方法に記載されるようにして評価した。各バーは、4つの非重複領域で計数された移動ケラチノサイトの平均±SEMを表す。1つの実験は2の代表である。
* スチューデントのt検定に基いて、サイトカインを用いない各コントロールと比較してp<0.001。
【0031】
図4は、OSMで刺激したNHEK及びOSMで処理したRHEから得られる発現プロフィールを示す。
NHEK (A)又はRHE (B)は、10 ng/mlのOSMとともに、又は用いずに24時間培養した。トータルRNAを単離し、Dnase Iで処理し、33P-標識cDNAプローブを作製するのに用い、これをcDNAアレイにハイブリダイズさせた。Molecular Dynamics Storm保存スクリーンに5日間
露光し、さらにスキャンした後に、コンピュータ画像を得た。局所のバックグラウンドを引いた後に、二重のスポットからの平均シグナル強度を、ハウスキーピング遺伝子について得られた値を用いて、プローブ標識における差について標準化した。(C) OSM-誘発変調は、刺激していない細胞についてのシグナル強度に対する、各サイトカインで処理した細胞についてのシグナル強度のパーセンテージ比率として表す。
【0032】
図5は、NHEKによるS100A7〜9合成に対するOSMの影響を示す。
NHEKは、0.4、0.8、1.6、3.1、6.3、12.5若しくは25 ng/mlのOSMとともに、又は用いずに48時間(A)、或いは10ng/mlのOSMとともに、又は用いずに6、12、24、48、72、96時間(B)培養した。トータルRNAを抽出し、逆転写し、S100A7及びHMBS mRNAの相対的発現を、リアルタイムPCRにより定量した。HMBSをハウスキーピング遺伝子として用いて、材料及び方法に説明するようにして遺伝子発現を標準化した。コントロール細胞に対する刺激された細胞の相対的発現として表される結果は、2つの独立した実験の代表である。(C) NHEKは、10 ng/mlのOSMとともに、又は用いずに48時間培養した。S100A8-カルグラヌリンA、S100A9-カルグラヌリンB、β-デフェンシン2及びフィラグリンmRNAの相対的発現を、定量RT-PCRにより定量した。結果は、コントロール細胞に対する刺激された細胞の相対的発現として表す。(D) NHEKを、10 ng/mlのOSMとともに、又は用いずに48及び96時間培養した。細胞溶解物20μgをSDS-PAGE (16%)により分離し、ニトロセルロースメンブレンに移した。ポンソーレッド染色を用いて、ロード量の均一性を制御した。S100A7、S100A8及びS100A9タンパク質レベルは、ウェスタンブロットにより決定した。結果は、3つの独立した実験の代表である。
【0033】
図6は、NHEKによるサイトカイン及びケモカインの産生を示す。IL-1β、IL-6、IL-8、IL-10、IL-12p70、TNFα、ENA-78、MIP 3βは、48時間のNHEK培養上清中で特異的ELISAにより測定した。細胞は、OSMの存在下又は非存在下で培養した。ENA78産生の用量応答(0.4〜25 ng/ml)、及び動態(6〜96h)の研究も行った。
【0034】
図7は、10 ng/mlのOSMを用いて4日間刺激したか又はしなかったRHEの組織学的及び免疫組織化学的分析を示す。RHEは固定し、パラフィンに包埋した。4ミクロンの垂直切片を、ヘマトキシリン/エオシンで染色したか、又は抗-K10ケラチンmAb、抗-フィラグリンmAb又は抗-S100A7 mAbと反応させ、次いで顕微鏡の下で写真撮影した(倍率×200)。
【0035】
図8は、NHEKにおけるIL-31及びOSMによるSTAT3及びMAPキナーゼリン酸化の誘導を示す。
NHEKは、10 ng/mlのOSM若しくは100 ng/mlのIL-31(A)、又は種々の濃度のこれらのサイトカイン(B)で15分間刺激したか又はせずに、ホスホ-STAT3 (P-STAT3)及びSTAT3タンパク質レベルを、ウェスタンブロットにより評価した。サイトカインでの刺激の前に、細胞を、中和抗体である抗-gp130 (AN-HH1)若しくは抗-OSMR (XR-M70)モノクローナル抗体の存在下で、又はアイソタイプコントロールであるmAb MC192 (最終抗体濃度15μg/ml) (C)と2時間インキュベートした。IL-31及びOSMに応答するホスホ-MAPK (P-MAPK)及びMAPKタンパク質レベルは、ウェスタンブロットにより評価した(D)。
【0036】
図9は、S100A7 mRNA発現に対するIL-31及びOSMの影響を示す。
NHEKは、3.1、6.3、12.5、25、50又は100 ng/mlのIL-31とともに、若しくは用いずに(A)、又は0.4、0.8、1.6、3.1、6.3、12.5又は25 ng/mlのOSMとともに、若しくは用いずに(B) 48時間培養した。100 ng/mlのIL-31 (C)又は10 ng/mlのOSM (D)の存在下又は非存在下でのS100A7 mRNA発現の動態の研究。トータルRNAを抽出し、逆転写し、S100A7及びHMBS mRNAの相対的発現を、リアルタイムPCRにより定量した。HMBSは、材料及び方法に説明するようにして、ハウスキーピング遺伝子として用いて遺伝子発現を標準化した。コントロール細胞に対する刺激された細胞の相対的発現として表される結果は、2つの独立した実験の代表である。
【0037】
図10は、ケラチノサイト移動に対するIL-31及びOSMの影響を示す。
インビトロ創傷は、マイトマイシンで処理した集密なNHEK培養物に導入し、ケラチノサイトは、100 ng/mlのIL-31又は10 ng/mlのEGF若しくはOSMとともに、又は用いずに48時間培養した。無細胞領域への細胞移動は、材料及び方法に記載するようにして評価した。各バーは、4つの非重複領域で計数された移動ケラチノサイトの平均±SEMを表す。1つの実験は、2の代表である。* スチューデントのt検定に基づいて、サイトカインを用いない各コントロールと比較してp<0.001。
【0038】
図11は、S100A7、hBD4/2及びKRT10 mRNA発現に対するいくつかのサイトカインのカクテルの影響を示す。
集密な通常の正常ヒトケラチノサイト(NHEK)を、記載する混合サイトカイン(各サイトカインは1 ng/mlの最終濃度)と24時間処理した。トータルRNAを抽出し、逆転写し、選択した遺伝子の発現を、リアルタイムPCRにより分析した。
【0039】
実施例1:材料及び方法
細胞培養、サイトカイン及び試薬
NHEKは、健康な胸部の皮膚の外科的サンプルから得た。研究実験のためのこれらのサンプルの使用は、ポワチエ病院の倫理委員会(the Ethical Committee of the Poitiers Hospital)により承認された。皮膚サンプルを、ディスパーゼ溶液(25 U/ml;Invitrogen Life Technologies, Cergy Pontoise, France)中で4℃にて一晩インキュベートした。表皮のシートを真皮からはずし、NHEKを、37℃にて15分間のトリプシン消化(トリプシン-EDTA, Invitrogen)により分離した。細胞を、ウシ脳下垂体抽出物(25μg/ml)及び組換え表皮成長因子(0.25 ng/ml)を補った無血清ケラチノサイト培地(Keratinocyte SFM;全てInvitrogenから購入)で培養した。刺激の前に、NHEKは、成長因子を加えないKeratinocyte SFM中で48時間枯渇させた。
【0040】
ヒト組換えOSM、IL-5、IL-6、可溶性IL-6Rは、R&D Systems (Oxon, UK)から購入した。IgG1アイソタイプコントロール(MC192)、抗-gp130 (AN-HH1)、抗-OSMR抗体(AN-A2)及び抗-LIFR (AN-E1)は、実験室で作製した。ホスホ-STAT3、ホスホ-MAPK、MAPKに指向させた抗体は、Upstate Biotechnology (Lake Placid, NY)から購入した。抗-STAT3、抗-S100A8及び抗-S100A9抗体は、Santa Cruz Biotechnology (Santa Cruz, CA, USA)からであった。抗-S100A7抗体は、Imgenex (San Diego, CA, USA)から購入した。ヤギ抗マウス及び抗ウサギペルオキシダーゼ標識免疫グロブリンは、Clinisciences (Montrouge, France)からであり、ウサギ抗ヤギペルオキシダーゼ結合抗体は、Sigma (Amersham Biosciences)からであった。
【0041】
RT-PCR及びRT-リアルタイムPCR分析
トータル細胞RNAは、Trizol試薬(Invitrogen)を用いて単離し、DNase I (0.05 U/μl;Clontech, Palo Alto, CA, USA)で処理した。cDNAを、2μgのトータルRNAから、MMLV逆転写酵素(Promega, Madison, WI)を用いるランダムヘキサマープライマーにより合成した。逆転写産物は、その後、OSMR (フォワード5'-CCTGCCTACCTGAAAACCAG-3' (配列番号1)とリバース5'-ACATTGGTGCCTTCTTCCAC-3' (配列番号2))、gp130 (フォワード5'-GGGCAATATGACTCTTTGAAGG-3' (配列番号3)とリバース5'-TTCCTGTTGATGTTCAGAATGG-3' (配列番号4))、LIFR (フォワード5'-CAGTACAAGAGCAGCGGAAT-3' (配列番号5)とリバース5'-CCAGTCCATAAGGCATGGTT-3' (配列番号6))及びGAPDH (フォワード5'-ACCACAGTCCATGCCATCAC-3' (配列番号7)とリバースTCCACCACCCTGTTGCTGTA (配列番号8))についてのプライマーを用いる25サイクルのPCRにより増幅した。増幅産物は、2%アガロースゲル電気泳動により分析した。
【0042】
定量リアルタイムPCRは、LightCycler-FastStart DNA MasterPLUS SYBR Green Iキット
(Roche, Mannheim, Germany)を用いて行った。反応成分は、1×FastStart DNA MasterPLUS SYBR Green I、並びに0.5μMのS100A7 (フォワード5'-GCATGATCGACATGTTTCACAAATACAC-3' (配列番号9)とリバース5'- TGGTAGTCTGTGGCTATGTCTCCC-3' (配列番号10))、S100A8 (Pattyn, Spelemanら 2003)、S100A9 (フォワード5'-GCTCCTCGGCTTTGACAGAGTGCAAG-3' (配列番号11)とリバース5'-GCATTTGTGTCCAGGTCCTCCATGATGTGT-3' (配列番号12))、hBD2/4 (フォワード5'-GCCATCAGCCATGAGGGTCTTG-3' (配列番号13)とリバース5'-AATCCGCATCAGCCACAGCAG- 3' (配列番号14))、KRT10 (フォワード5'-GCCCGACGGTAGAGTTCTTT-3' (配列番号15)とリバース5'-CAGAAACCACAAAACACCTTG -3' (配列番号16))、ハウスキーピング遺伝子としてのヒドロキシメチル-ビラン合成酵素(HMBS) (Vandesompele, De Preterら 2002)についてのフォワード及びリバースプライマーであった。ヨウ化プロピジウムを用いるcDNAの蛍光定量の後に、250 ng、25 ng及び2.5 ngのcDNAを、LightCyclerのガラスキャピラリにPCR鋳型として加えた。サイクル条件は、95℃にて10分間のポリメラーゼ活性化と、95℃で10秒、64℃で5秒及び72℃で18秒の50サイクルと、単独の蛍光測定とを含んでいた。1秒当たり0.1℃の加熱速度で60℃から95℃まで温度を増加させることにより得られた融解曲線分析、及び連続蛍光測定により、融合温度と思われる狭い単独ピークが明らかになった。数学的モデルを用いて、HMBS参照遺伝子と比較した標的遺伝子の相対的定量を決定した(Pfaffl 2001)。
【0043】
cDNAマクロアレイを用いる遺伝子発現のプロファイリング
トータルRNAを、PCR実験について記載したようにして単離した。DNase処理、ポリA+ RNA富化、33P-標識cDNAプローブ合成、精製及びカスタムAtlasアレイメンブレンへのハイブリダイゼーション(Bernard, Pedrettiら 2002)は、Clontech (Clontech, Palo Alto, USA)の推奨に従って行った。メンブレンは、Molecular Dynamics Storm保存スクリーン(storage screen)に5日間露光し、ホスホリイメージャースキャナ(Molecular Dynamics Stormアナライザ, Amersham Biosciences, Uppsala, Sweden)を用いてスキャンした。局所的バックグラウンドを差し引いた後に、二重のスポットからの平均シグナル強度を、ハウスキーピング遺伝子について得られた値を用いてプローブ標識における差について標準化した(Bernardら, 2002)。各遺伝子について、OSMにより誘発された変調は、刺激したサンプル対コントロールサンプルについての相対的発現値として表した。任意に、2を超える変調は、RT-リアルタイムPCRアッセイを用いる確認について著しいとみなした。
【0044】
ウェスタンブロッティング分析
STAT3及びMAPKのリン酸化のために、NHEKを、記載するサイトカインの存在下で15分間刺激した。細胞を、SDSサンプルバッファー(62.5 mM Tris-HCl pH 6.8、2% SDS、10%グリセロール、50mM DTT、0.1%ブロモフェノールブルー)中で溶解し、超音波破砕し、次いでSDS-PAGEに供してImmobilonメンブレンに移した。メンブレンは、その後、一次抗体と一晩インキュベートした後に適切なペルオキシダーゼ標識二次抗体と60分間インキュベートした。反応は、製造業者の使用説明に従う化学発光により視覚化した。メンブレンを、0.1 Mグリシン、pH 2.8中で抗体をはがし、1 M Tris-HCl、pH 7.6中で中和した後に再ブロットした。中和実験のために、NHEKは、適切な抗体と2時間インキュベートした後に刺激した。
【0045】
gp130、LIFR及びOSMRの発現を決定するために、細胞を10 mM Tris HCl pH 7.6、5 mM EDTA、50 mM NaCl、30 mMピロリン酸ナトリウム、50 mMフッ化ナトリウム、1 mMオルトバナジン酸ナトリウム、プロテイナーゼ阻害剤及び1% Brij 96中で溶解した。溶解及び遠心分離により細胞破片を除去した後に、上清を上記のようにして処理した。
【0046】
S100タンパク質発現のために、NHEKをOSM (10 ng/ml)の存在下に2日間刺激した。細胞溶解は、50 mM Tris HCl pH 7.4、150 mM NaCl、1 mM EDTA、1% Triton、1%デオキシコール酸ナトリウム、0.1 % SDS、1 mM PMSF、1 mMオルトバナジン酸ナトリウム、1%プロテア
ーゼ阻害剤を用いて行った。S100A7、S100A8及びS100A9は、上記のようにして免疫化学により検出した。ポンソーレッド染色を用いて、ロード量の均一性を制御した。
【0047】
インビトロケラチノサイト移動アッセイ
ケラチノサイトを、I型コラーゲン(200μg/ml, Institut Jacques Boy, Reims, France)で予めコートしたウェル中で、80 %の集密性に到達するまで培養した。細胞を、Keratinocyte SFM中で48時間枯渇させ、10μg/mlのマイトマイシンC (Sigma)で2時間処理して細胞増殖を妨げた。無細胞領域は、ケラチノサイトの単層をプラスチックのピペットチップでかきとることにより作製した。無細胞領域へのケラチノサイトの移動は、EGF又はOSMの存在下若しくは非存在下での48時間の培養の後に評価した。倒立位相差顕微鏡を用いる。移動ケラチノサイトの数は、4つの非重複領域において計数した。値は、インビトロ損傷の境界(frontiers)を超えるmm2当たりの細胞の平均±SEMを表す。スチューデントのt検定を、統計学的分析に用いた。
【0048】
再構成されたヒト表皮モデル
組織学的及び免疫組織化学的な研究のために、空気‐培地の界面で12日間増殖させたRHEを、SkinEthic Laboratories (Nice, France)から購入した。これらは、多層表皮からなり、これはヒトの皮膚に類似した形態学的及び成長の特徴を示す(Rosdy, Bertinoら 1997)。推奨されるようにして、RHEをSkinEthic増殖培地中で1日間増殖させ、その後、OSMの存在下又は非存在下に4日間刺激した。次いで、これらを平衡10%ホルマリン溶液中で固定し、パラフィンに包埋した。4ミクロンの垂直切片を、ヘマトキシリン/エオシンで染色するか、又は特異的Abとペルオキシダーゼ結合抗体とで染色し、かつ標準的な方法に従ってヘマトキシリンで対比染色した(Rosdy, Bertinoら 1997)。抗-K10ケラチン及び抗フィラグリンモノクローナル抗体は、Lab Vision Corporation (Fremont, CA, USA)からであった。
【0049】
cDNAマクロアレイを用いる遺伝子発現プロファイリングのために、17日齢のRHEを、SkinEthic維持培地で1日増殖させた後に、OSMの存在下又は非存在下に24時間刺激した。トータルRNAを単離し、上記のようにしてcDNAアレイを行った。
【0050】
実施例2:ヒトケラチノサイトはその表面上にある型のOSMを発現した
正常なヒトケラチノサイトにおけるOSMの潜在的な機能を示すために、本発明者らは、まず、その受容体鎖発現の分析を行った。発現されたI型又はII型の受容体の性質を決定するために、gp130、LIFRβ及びOSMRβについてのRT-PCRを、ケラチノサイトの初代培養から開始して行った。CO-G-UVM及びグリア芽腫細胞は、LIFRについてのコントロールとして用いた。得られた結果は、NHEKはOSMR及びgp130についての転写産物を主に発現するが、LIFR鎖は低いレベルでしか証明できなかった(図1A)。RNA分析を、フローサイトメトリによる対応するタンパク質の発現レベルの測定によりさらに補強した。蛍光分析は、NHEK細胞表面でのgp130及びOSMRβの明確な発現を明らかにした(図1B)。対照的に、膜のLIFRβ発現は検出されなかったが、抗-LIFRβ抗体は、ポジティブコントロールとして用いた細胞系とインキュベートしたときに、予想される結果を示した。このことは、gp130及びOSMRβ鎖の検出、並びにNHEKにおけるLIFRβ発現の不在を示すウェスタンブロット分析によってさらに支持された(図1C)。4人の異なるドナーからのサンプルについて行われた類似の実験は、同様の結果に導き、ドナー間のI型又はII型のOSM受容体発現の多様性の可能性を除外した。これらの第一の結果は、ヒトケラチノサイトが特定のII型のOSM受容体を優先的に発現したことを示す。
【0051】
実施例3:ヒトケラチノサイトにおいて、STAT-3及びMAPキナーゼ経路はOSMにより補充される
本発明者らは、NHEKにおけるOSM誘発シグナル伝達を示す。STAT3は、通常、OSM II型受
容体経路により補充されるので、サイトカインの漸増濃度に応答するシグナル伝達分子のチロシンリン酸化を分析した。チロシンリン酸化の強い誘導がSTAT3について観察され、OSMが3 ng/mlまで下がってもプラトーのレベルの値がまだ存在していた(図2B)。gp130及びOSMRβサブユニットの関与が、OSM接触の前に受容体中和mAbをNHEK培養に加えたときにSTAT3リン酸化がブロックされることによりさらに証明された(図2C)。重要なことには、抗-OSMRβmAbの存在下で観察されたSTAT3リン酸化の完全な中和は、NHEKにおける共有(share) LIF/OSM I型受容体のOSMによる補充が存在しないことをさらに証明した。この観察に合致して、LIF接触後に、STAT3活性化の証明はNHEKにおいて観察できなかった。
【0052】
さらに、II型OSM受容体複合体は、共有LIF/OSM受容体に比べて、Mapキナーゼ経路の補充についてより強力な活性化物質であることも知られている。LIFRbではなくOSMRβを介して媒介されるERK1/ERK2とShcアダプタとの間の協力的な影響は、OSMに応答してのMAPキナーゼ経路のこの強い活性化を説明する(Boulton, Stahlら 1994)。NHEK におけるサイトカインに応答してのERK1/2シグナル伝達を、よって、それらのチロシンリン酸化レベルを決定することにより分析した。予想されたように、OSMを用いるNHEKの刺激は、MAPキナーゼリン酸化を迅速に増加させた(図2D)。まとめると、これらの結果は、ヒトケラチノサイトにおいて発現されるgp130/OSMRβ受容体複合体が完全に機能的であり、観察されたシグナル全体がII型受容体を介して媒介されることを示した。
【0053】
実施例4:OSMは、ケラチノサイト移動の強力な誘発物質である
OSMに対するNEHKの機能的応答を強調するために、本発明者らは、創傷治癒を模倣しかつケラチノサイト移動に基づくインビトロモデルに対するOSMの潜在的影響を分析した(Kira, Sanoら 2002)。培養開始から48時間後に、ウェルの中位(middle)に存在する細胞をかきとることにより回収し、残りのケラチノサイトを、ケラチノサイト移動を引き起こすことが知られている EGF、又はOSMのいずれかで刺激した。さらに36時間培養した後に、細胞移動を誘導するサイトカインの潜在性を、視覚的に又は細胞計数により決定した(図3)。得られた結果は、OSMが、EGFの存在下で観察されたことと同様に、NHEKの著しい移動を導いたことを示す。
【0054】
実施例5:ヒトケラチノサイトにおけるOSM誘発遺伝子発現の同定
NHEK機能的応答をよりよく理解するために、本発明者らは、cDNAアレイを用いて、OSMにより誘発されるケラチノサイト遺伝子発現プロフィールの改変を分析した。用いたアレイは、ケラチノサイト研究のために特別に設計され、二重にスポットされた586の異なるcDNAからなった。これらは、ケラチノサイトの細胞構造、代謝、細胞外マトリクス、接着、分化、シグナル伝達、シグナル変換、アポトーシス及びストレスに関係する遺伝子であった(Bernardら, 2002)。コントロール又はOSMで刺激したNHEKから抽出したRNAは、逆転写により、標識cDNAプローブを作製するのに用いた。これらのcDNAプローブでAtlas cDNAアレイメンブレンをプローブすることにより、OSMが36遺伝子の発現を増加させ、38遺伝子の発現を減少させたことが明らかになった。OSMは、ケラチノサイト分化に関連する多くの一連の遺伝子、例えばサイトケラチン(CK)1、CK10、フィラグリン及びロリクリン遺伝子をダウンレギュレートする。アップレギュレートされる遺伝子のうち、本発明者らは、カルシウム結合性タンパク質であるソリアシン(S100A7)、カルグラヌリン(S100A8、S100A9)及びS100好中性タンパク質について著しい増加を見出した(図4)。興味深いことに、これらのタンパク質の発現は、炎症性組織においてアップレギュレートされることが知られている(Madsen, Rasmussenら 1991; Nagase及びWoessner 1999; Roth, Voglら 2003)。OSMはまた、組織防御に関わる、Gタンパク質結合受容体であるHM74、スーパーオキシドジスムターゼ2及びβデフェンシン遺伝子を誘導した。マトリクスメタロプロテイナーゼ1及びテナシン(tenascin)のような組織リモデリングに関わる遺伝子も、OSMにより誘導された。さらに、OSMは、ケモカインCXCL1 (MIP-2α)、CXCL5 (上皮由来好中球活性化ペプチド(ENA 78))及びCXCL8 (IL-8)、並びに血小板由来成長因子A (PDGF-A)遺伝子の発現を増
加させた。
【0055】
得られた結果は、ヒトケラチノサイトにおいて、OSMが、炎症性プロセス及び先天免疫応答に関わるいくつかの遺伝子を補充できたことを示す。
【0056】
実施例6:OSMは、ケラチノサイトを誘発してソリアシン、カルグラヌリン、βデフェンシン及びケモカインを産生させた
設計されたアレイを用いて得られた結果をさらに強化するために、mRNA及びタンパク質レベルでの定量的分析を、選択された数の同定された遺伝子について行った。OSMに応答してのソリアシン/S100A7 mRNA発現の定量的分析を、動態及び用量応答の研究とともにRT-リアルタイムPCRにより行った。本発明者らは、ソリアシン/S100A7 mRNAが、48時間の処理の後に1.6〜6.3 ng/mlの範囲のOSMに応答して用量依存的な様式でアップレギュレートされ、6.3 ng/ml OSMについて、コントロールを超えてシグナルが50倍増加して、プラトーに達したことを示す(図5A)。動態研究は、10 ng/mlのOSMでの刺激の12時間後に、ソリアシン/S100A7 mRNA発現の増加が開始したことを明らかにした(図5B)。これは96時間まで増加し続け、コントロールの値を超える約290倍の強い誘導であった。このことは、ソリアシン/S100A7並びに2つの関連するカルシウム結合性タンパク質であるS100A8及びS100A 9カルグラヌリンのウェスタンブロット分析により、タンパク質レベルで確かめられた(図5D)。結果は、10 ng/mlのOSMにNHEKを曝すことにより、研究したタンパク質の発現が増加し、これは2日目よりも4日目のほうが強かったことを示す(図5D)。図5Cは、皮膚活性化の2つの重要なマーカーであるフィラグリン及びβデフェンシン-2のRNA定量的発現を分析することにより得られた結果を描写する。
【0057】
48時間のNHEKでのケモカインCXCL5及びCXCL8の産生も、OSM刺激の下で明確に増強される(図6B)。
【0058】
実施例7:OSMは、再構成されたヒト表皮の過形成を引き起こし、分化関連抗原(differentiation related antigens)の発現を変調する
表皮分化の原動力にさらに取り組むために、本発明者らは、基底細胞層増殖及び段階的表皮分化プロセスを評価するためにインビトロRHEに対するOSMの生物学的影響を試験した。コントロールRHEの組織学的分析は、インビボでの表皮に類似し、無損傷の基底棘状顆粒角化細胞層を含み、かつ上方顆粒層に多数のケラトヒアリン顆粒を含む角化多重層上皮を示した(図7)。OSMは、ケラチノサイト層の過形成を引き起こし、RHEの全体的な厚みの増加を導いた。さらに、顆粒層におけるケラトヒアリン顆粒の損失及び凝縮核(picnotic nuclei)の存在が観察された。RHEのcDNAアレイプロフィール分析は、NHEKについて以前に記載されたことと同様に、OSMがS100A7、S100A8、S100A9及びS100好中性タンパク質遺伝子を強くアップレギュレートしたことを確かにした(図4)。免疫組織化学により、OSMで処理したRHEにおけるS100A7タンパク質のアップレギュレーションを確認した(図7)。NHEKについてのデータに合致して、RHEに対するOSM処理も、CXCL5、CXCL8ケモカイン遺伝子、並びにPDGF-A及びカドヘリン3遺伝子の転写をアップレギュレートした。RHEに特異的でありNHEKでは検出されないが、OSMは、CK6A、6B、6D、7、13、14、16、皮膚由来抗ロイコプロテイナーゼ及びTGFβ-誘導性早期タンパク質をアップレギュレートした。
【0059】
一方、OSMはケラチノサイト分化に関連する遺伝子、例えばインボルクリン(involucrin)、フィラグリン及びカルモジュリン様皮膚タンパク質をダウンレギュレートする(Mehul, Bernardら 2000; Rogers, Kobayashiら 2001; Jonak, Klosnerら 2002; Wagener, van Beurdenら 2003)。RHE切片に対して行った免疫組織化学的分析は、OSMで処理したRHEにおけるフィラグリン及びケラチン10の発現の減少により示されるように(図7)、ケラチノサイトの分化の阻害を確かにした。
【0060】
実施例8:考察
cDNAアレイアプローチ、特にヒトの皮膚における遺伝子発現の分析のために設計されたcDNAアレイアプローチの使用により、ヒトケラチノサイトにおけるOSM標的遺伝子の同定、並びに移動及び分化を含む種々のプロセスにおけるOSMの関与の証明が可能になった。特に、NHEK及びRHEにおけるS100A7、S100A8及びS100A9タンパク質の発現の強く、用量応答性で、OSMにより媒介される誘導は、サイトカインの炎症誘発性及び走化性の効果を証明する。皮膚の炎症におけるIL-10及びOSMの対抗する効果は、S100A8のIL-10により誘発されるダウンレギュレーションと、単球によるS100A9放出とにより強調される(Lugering, Kucharzikら 1997)。S100A7、S100A8及びS100A9は、カルシウム結合タンパク質の多形質発現性S100ファミリーに属する(Roth, Voglら 2003)。それらの主要な機能は未だ明らかではないが、これらは顕著な炎症性機能を演じ (Watson, Leygueら 1998; Donato 1999; Roth, Voglら 2003)、かつ多数の細胞内及び細胞外の活動、例えば細胞骨格成分の運動性又は走化性の原動力の厳密な調節に関わるようである(Ryckman, Vandalら 2003)。興味深いことに、S100A7、S100A8及びS100A9の3つ全てのタンパク質が正常な皮膚表皮及び未分化の培養ケラチノサイトにおいて低いか又は検出不可能なレベルで発現されると報告されているが、これらは、異常に分化した乾癬ケラチノサイトにおいて(Broome, Ryanら 2003)、創傷修復の間(Thorey, Rothら 2001)、及び表皮皮膚腫瘍において(Watson, Leygueら 1998; Gebhardt, Breitenbachら 2002; Alowami, Qingら 2003)多く発現される。炎症性細胞、特に好中球及びCD4+ Tリンパ球に対するS100A7の走化性効果により、S100A7は乾癬外傷の形成に関与し得ることが示唆される(Watson, Leygueら 1998)。S100A7はこれらの機構の上流で作用するので、本発明者らは、OSMが病的な状態でのS100A7の誘導のための鍵となる分子であり、病的段階に関与することを証明している。OSMによるさらなる遺伝子の変調は、OSMの炎症誘発性及び走化性の役割に好ましいものでもある。実際に、皮膚における接着性相互作用及び細胞浸潤を減じることにより炎症を弱める好中性誘引物質ケモカインCXCL5/ENA-78の誘導及びヘムオキシゲナーゼ1のダウンレギュレーションは、皮膚における好中球の流入に貢献し得る(Koch, Kunkelら 1994; Wagener, van Beurdenら 2003)。
【0061】
OSMにより誘導されるMMP-3発現は、炎症性皮膚疾患及び創傷修復の関係においても興味深い。MMP-3は正常な皮膚においては検出不可能であるが、損傷の後に基底層の増殖性ケラチノサイトにより発現される(Pilcher, Wangら 1999)。多くの疾患の進行の間に、MMP-3は、組織再吸収の間の細胞外マトリクスの除去による表皮リモデリングに関わり(Nagase及びWoessner 1999; Pilcher, Wangら 1999)、MMP-3遺伝子を欠損するマウスは、表皮の創傷修復に欠陥がある(Bullard, Lundら 1999)。インビトロ創傷アッセイを用いて、本発明者らは、ケラチノサイト移動がOSM刺激により強く増大することを証明した。これらのデータは、ケラチノサイトにおけるSTAT3欠陥が移動を弱めることに導くことの証明と合致する(Sano, Itamiら 1999)。炎症性条件の下で、OSMは、MMP-3又はS100A8-S100A9依存性の機構を介してケラチノサイト移動及び創傷治癒を増強する必須のメディエイタの1つであると考えられる。創傷治癒におけるOSMの関与を確立するさらなる証拠は、皮膚修復プロセスの開始の間の結合組織についての主要な増殖性及び移動性の刺激物質であるPDGFのRHEにおける強い誘導である(Rollman, Jensenら 2003)。
【0062】
本発明者らは、OSMがRHEのケラチノサイト層の全体的な厚みを増加させることも示した。このプロセスは、基底細胞の過剰増殖とは関係しないようである。なぜなら、フィラグリン、ロリクリン又はインボルクリンの産生の減少により示されるように、Ki67発現がOSMに応答しては誘導されないが、終末のケラチノサイト分化の阻害に起因する可能性がより高いからである。OSMは、ケラチノサイト分化に直接関連する、カルモジュリンファミリーの2つのメンバーであるカルモジュリン様皮膚タンパク質(CLSP)及びカルモジュリン関連タンパク質NB-1の発現をダウンレギュレートする(Mehul, Bernardら 2001; Rogers, Kobayashiら 2001)。CLSPは、ロリクリン又はインボルクリンのようなタンパク質の形成及び組み立てに関係する鍵となる酵素であるトランスグルタミナーゼ-3に結合して、表皮の角化細胞エンベロープを形成する(Mehul, Bernardら 2000)。ケラチン発現プロフィール、すなわちケラチン6の過剰発現及びケラチン10の阻害に対するOSMの変調効果も、表皮分化の阻害の概念を支持する。ケラチン6は、創傷治癒、乾癬、発癌を含む過剰増殖及び/又は炎症性の状況の下で、或いは表皮過形成を引き起こすレチノイン酸のような作用物質により、誘導されることが知られている(Navarro, Casatorresら 1995; Komine, Raoら 2000)。対照的に、終末分化している表皮ケラチノサイトにおいて通常は発現しているケラチン10は、創傷治癒の間に減少される(Paramio, Casanovaら 1999)。
【0063】
実施例9:IL-31を用いて得られる同様の結果
実施例3、4及び5で示す研究を、OSMの代わりにIL-31を用いて行い、類似の結果を得た。IL-31は、NHEKにおいてSTAT3シグナル伝達経路を補充し(図8)、ソリアシン(S100A)及びカルグラヌリンA及びB (S100A8-9)の発現を誘発する(図9)。IL-31も、ケラチノサイト移動を誘発できる(図10)。
【0064】
実施例10:ガンマインターフェロンは、シグナル変換に対するIL-31の作用を強化する
NHEKを、IL-31刺激(50 ng/ml)の前に50 g/mlのガンマインターフェロン(INFγ)の存在下で24時間プレインキュベーションしたとき、P-STAT3のレベルは、培地のみの中でプレインキュベートしたNHEKに比べて3〜4倍増加した(図8について行ったものと同じ研究)。このことは、INFγがシグナル変換に対するIL-31の作用の修飾物質であることを証明する。
【0065】
実施例11:IL-17及びTNFαは、いくつかの炎症性マーカーの発現に対するOSMの作用を強化する
ケラチノサイトに対するいくつかのサイトカインの併用効果を、ケラチノサイト炎症マーカーであるソリアシン(S100A7)、デフェンシンβ-2/β-4 (hBD2/4) mRNAの発現を、種々のサイトカインカクテルの存在下で測定することにより試験した。これらのカクテルの影響を、ケラチン10 (KRT10) mRNAに対しても試験した。なぜなら、KRT10は組織治癒に関連する分化マーカーだからである。
【0066】
この目的のために、集密な正常ヒトケラチノサイト(NHEK)を、記載するサイトカイン混合物と24時間処理した(各サイトカインは1 ng/mlの最終濃度)。トータルRNAを抽出し、逆転写し、選択された遺伝子の発現を、記載するようにしてリアルタイムPCRにより分析した。
これらの結果を、以下の表1及び図11に示す。
【0067】
【表1】
【0068】
6つの選択されたサイトカインの混合物(M3)は、ケラチノサイト炎症マーカーであるソリアシン(S100A7)及びデフェンシンβ-2/β-4 (hBD2/4)の発現に対して強い相乗効果を示した。予想されたように、KRT10発現は、これらのサイトカインの存在下で低下する。IFNγ、IL-22又はIL-1αのいずれかを混合物から除去しても、サイトカインの完全なカクテルの活性を著しくは減少させなかった。よって、これらのサイトカインはおそらく、観察された相乗作用に直接は関与していない。逆に:
- 5種サイトカイン参照混合物(不活性であるのでIFNγを除外した)からOSMを除外すると、混合物の活性は、S100A7について3倍、及びhBD2/4について2.5倍減少された;
- TNFαを除外すると、混合物の活性は、S100A7について3倍、及びhBD2/4について5.6倍減少された;かつ
- IL-17を除外すると、混合物の活性は、S100A7について4.5倍、及びhBD2/4について36倍減少された。
【0069】
これらの結果は、ケラチノサイトにおける最大限の応答について、OSM、TNFα及びIL-17の間の強い相乗作用を示す。
よって、OSMは、IL-17及び/又はTNFαとともに相乗作用を示すことができると考えられる。さらに、OSMは、低いサイトカイン濃度であっても、創傷再上皮形成促進のためにIL-17及び/又はTNFαと相乗的に作用できることが示唆される。
【0070】
実施例12:OSMは、皮膚先天性免疫応答を改善する
乾癬患者の皮膚におけるケラチノサイトは、高レベルの抗菌性ペプチドを産生する(Nomura, Golevaら 2003)。正常な皮膚又はアトピー性皮膚炎の患者の皮膚にはごくわずかなレベルでしか存在しないこれらのペプチドは、感染の動物モデルにおいて示されるように、侵入微生物に対する皮膚先天性免疫応答において必須である(Nizet, Ohtakeら 2001)。S100A7-ソリアシンは、大腸菌(Escherichia coli)による皮膚の感染に対する耐性を与えることが示されているので(Glaser, Harderら 2005)、OSMを用いた刺激の後のケラチノサイトにおけるS100A7-ソリアシン遺伝子発現の増強は、皮膚の先天性免疫応答におけるOSMについての役割を指摘する。同様に、抗菌性活性を有する別のタンパク質であるβ-デフェンシン2 (Harder, Bartelsら 1997)は、ヒトケラチノサイトにおいてOSMにより誘導される。β-デフェンシン2は、乾癬外傷においては高いレベルで発現されるが、アトピー性皮膚炎において観察されるものにおいては低いレベルでしか発現されない(Ong, Ohtakeら 2002)。この観察と調和して、乾癬の患者は、アトピー性皮膚炎を有する患者に比べると、感染に対して感受性がより低いと報告されている(Nomura, Golevaら 2003)。
よって、上記の実施例6及び11で報告する結果は、OSM、IL-17及びTNFαがケラチノサイトの有効な活性化物質であり、これらのサイトカインの単独又は組み合わせでの局所投与が、抗菌性ペプチドの発現を増強し、かつ皮膚先天免疫を改善できることを示す。
【0071】
参考文献
Alowami, S., G. Qing,ら (2003). "Psoriasin (S100A7) expression is altered during
skin tumorigenesis." BMC Dermatol 3(1): 1.
Benigni, F., G. Fantuzzi,ら (1996). "Six different cytokines that share GP130 as
a receptor subunit, induce serum amyloid A and potentiate the induction of interleukin-6 and the activation of the hypothalamus-pituitary-adrenal axis by interleukin-1." Blood 87(5): 1851-4.
Bernard, F. X., N. Pedretti,ら (2002). "Comparison of gene expression profiles in human keratinocyte mono-layer cultures, reconstituted epidermis and normal human skin; transcriptional effects of retinoid treatments in reconstituted human epidermis." Exp Dermatol 11(1): 59-74.
Bonifati, C., A. Mussi,ら (1998). "Spontaneous release of leukemia inhibitory factor and oncostatin-M is increased in supernatants of short-term organ cultures
from lesional psoriatic skin." Arch Dermatol Res 290(1-2): 9-13.
Boulton, T. G., N. Stahlら (1994). "Ciliary neurotrophic factor/leukemia inhibitory factor/interleukin 6/oncostatin M family of cytokines induces tyrosine phosphorylation of a common set of proteins overlapping those induced by other cytokines and growth factors." J Biol Chem 269(15): 11648-55.
Broome, A. M., D. Ryan,ら (2003). "S100 protein subcellular localization during epidermal differentiation and psoriasis." J Histochem Cytochem 51(5): 675-85.
Bullard, K. M., L. Lund,ら. (1999). "Impaired wound contraction in stromelysin-1-deficient mice." Ann Surg 230(2): 260-5.
【0072】
Dillon, S. R., C. Sprecher,ら (2004). "Interleukin 31, a cytokine produced by activated T cells, induces dermatitis in mice." Nat Immunol 5(7): 752-60.
Donato, R. (1999). "Functional roles of S100 proteins, calcium-binding proteins of the EF-hand type." Biochim Biophys Acta 1450(3): 191-231.
Gallucci, R. M., P. P. Simeonova,ら (2000). "Impaired cutaneous wound healing in
interleukin-6-deficient and immunosuppressed mice." Faseb J 14(15): 2525-31.
Gallucci, R. M., D. K. Sloan,ら (2004). "Interleukin 6 indirectly induces keratinocyte migration." J Invest Dermatol 122(3): 764-72.
Gebhardt, C., U. Breitenbach,ら (2002). "Calgranulins S100A8 and S100A9 are negatively regulated by glucocorticoids in a c-Fos-dependent manner and overexpressed throughout skin carcinogenesis." Oncogene 21(27): 4266-76.
Glaser, R., J. Harder,ら (2005). "Antimicrobial psoriasin (S100A7) protects human skin from Escherichia coli infection." Nat Immunol 6(1): 57-64.
Harder, J., J. Bartels,ら (1997). "A peptide antibiotic from human skin." Nature
387(6636): 861.
【0073】
Ihn, H.及びK. Tamaki (2000). "Oncostatin M stimulates the growth of dermal fibroblasts via a mitogen-activated protein kinase-dependent pathway." J Immunol 165(4): 2149-55.
Jonak, C., G. Klosner,ら (2002). "Subcorneal colocalization of the small heat shock protein, hsp27, with keratins and proteins of the cornified cell envelope." Br J Dermatol 147(1): 13-9.
Kira, M., S. Sano,ら (2002). "STAT3 deficiency in keratinocytes leads to compromised cell migration through hyperphosphorylation of p130(cas)." J Biol Chem 277(15): 12931-6.
Koch, A. E., S. L. Kunkel,ら (1994). "Epithelial neutrophil activating peptide-78: a novel chemotactic cytokine for neutrophils in arthritis." J Clin Invest 94(3): 1012-8.
Komine, M., L. S. Rao,ら (2000). "Inflammatory versus proliferative processes in
epidermis. Tumor necrosis factor alpha induces K6b keratin synthesis through a transcriptional complex containing NFkappa B and C/EBPbeta." J Biol Chem 275(41): 32077-88.
【0074】
Lugering, N., T. Kucharzik,ら (1997). "Importance of combined treatment with IL-10 and IL-4, but not IL-13, for inhibition of monocyte release of the Ca(2+)-binding protein MRP8/14." Immunology 91(1): 130-4.
Madsen, P., H. H. Rasmussen,ら (1991). "Molecular cloning, occurrence, and expression of a novel partially secreted protein "psoriasin" that is highly up-regulated in psoriatic skin." J Invest Dermatol 97(4): 701-12.
Mehul, B., D. Bernard,ら (2001). "Calmodulin-like skin protein: a new marker of
keratinocyte differentiation." J Invest Dermatol 116(6): 905-9.
Mehul, B., D. Bernard,ら (2000). "Identification and cloning of a new calmodulin-like protein from human epidermis." J Biol Chem 275(17): 12841-7.
Nagase, H.及びJ. F. Woessner, Jr. (1999). "Matrix metalloproteinases." J Biol Chem 274(31): 21491-4.
【0075】
Navarro, J. M., J. Casatorres,ら (1995). "Elements controlling the expression and induction of the skin hyperproliferation-associated keratin K6." J Biol Chem 270(36): 21362-7.
Nizet, V., T. Ohtake,ら (2001). "Innate antimicrobial peptide protects the skin from invasive bacterial infection." Nature 414(6862): 454-7.
Nomura, I., E. Goleva,ら (2003). "Cytokine milieu of atopic dermatitis, as compared to psoriasis, skin prevents induction of innate immune response genes." J Immunol 171(6): 3262-9.
Ong, P. Y., T. Ohtake,ら (2002). "Endogenous antimicrobial peptides and skin infections in atopic dermatitis." N Engl J Med 347(15): 1151-60.
Paglia, D., S. Kondo,ら (1996). "Leukaemia inhibitory factor is expressed by normal human keratinocytes in vitro and in vivo." Br J Dermatol 134(5): 817-23.
【0076】
Paramio, J. M., M. L. Casanova,ら (1999). "Modulation of cell proliferation by cytokeratins K10 and K16." Mol Cell Biol 19(4): 3086-94.
Pattyn, F., F. Speleman,ら (2003). "RTPrimerDB: the real-time PCR primer and probe database." Nucleic Acids Res 31(1): 122-3.
Pfaffl, M. W. (2001). "A new mathematical model for relative quantification in real-time RT-PCR." Nucleic Acids Res 29(9): e45.
Pilcher, B. K., M. Wang,ら (1999). "Role of matrix metalloproteinases and their inhibition in cutaneous wound healing and allergic contact hypersensitivity." Ann N Y Acad Sci 878: 12-24.
Rogers, M. S., T. Kobayashi,ら (2001). "Human calmodulin-like protein is an epithelial-specific protein regulated during keratinocyte differentiation." Exp Cell
Res 267(2): 216-24.
Rollman, O., U. B. Jensen,ら (2003). "Platelet derived growth factor (PDGF) responsive epidermis formed from human keratinocytes transduced with the PDGF beta receptor gene." J Invest Dermatol 120(5): 742-9.
【0077】
Rosdy, M., B. Bertino,ら (1997). "Retinoic acid inhibits epidermal differentiation when applied topically on the stratum corneum of epidermis formed in vitro by
human keratinocytes grown on defined medium." In Vitro Toxicology 10(1): 39-47.Roth, J., T. Vogl,ら (2003). "Phagocyte-specific S100 proteins: a novel group of
proinflammatory molecules." Trends Immunol 24(4): 155-8.
Ryckman, C., K. Vandal,ら (2003). "Proinflammatory activities of S100: proteins S100A8, S100A9, and S100A8/A9 induce neutrophil chemotaxis and adhesion." J Immunol 170(6): 3233-42.
Sano, S., S. Itami,ら (1999). "Keratinocyte-specific ablation of Stat3 exhibits impaired skin remodeling, but does not affect skin morphogenesis." Embo J 18(17): 4657-68.
Sugawara, T., R. M. Gallucci,ら (2001). "Regulation and role of interleukin 6 in
wounded human epithelial keratinocytes." Cytokine 15(6): 328-36.
【0078】
Taga, T.及びT. Kishimoto (1997). "Gp130 and the interleukin-6 family of cytokine
s." Annu Rev Immunol 15: 797-819.
Thorey, I. S., J. Roth,ら (2001). "The Ca2+-binding proteins S100A8 and S100A9 are encoded by novel injury-regulated genes." J Biol Chem 276(38): 35818-25.
Vandesompele, J., K. De Preter,ら (2002). "Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes." Genome Biol 3(7): RESEARCH0034.
Wagener, F. A., H. E. van Beurden,ら (2003). "The heme-heme oxygenase system: a molecular switch in wound healing." Blood 102(2): 521-8.
Wahl, A. F.及びP. M. Wallace (2001). "Oncostatin M in the anti-inflammatory response." Ann Rheum Dis 60 Suppl 3: iii75-80.
Watson, P. H., E. R. Leygue,ら (1998). "Psoriasin (S100A7)." Int J Biochem Cell Biol 30(5): 567-71.
【図面の簡単な説明】
【0079】
【図1】NHEKによるOSM受容体の発現を示す。
【図2】NHEKでのOSMによるSTAT3及びMAPキナーゼのリン酸化の誘導を示す。
【図3】ケラチノサイト移動に対するOSMの影響を示す。
【図4】OSMで刺激したNHEK及びOSMで処理したRHEから得られる発現プロフィールを示す。
【図5】NHEKによるS100A7〜9合成に対するOSMの影響を示す。
【図6】NHEKによるサイトカイン及びケモカインの産生を示す。
【図7】10 ng/mlのOSMを用いて4日間刺激したか又はしなかったRHEの組織学的及び免疫組織化学的分析を示す。
【図8】NHEKにおけるIL-31及びOSMによるSTAT3及びMAPキナーゼリン酸化の誘導を示す。
【図9】S100A7 mRNA発現に対するIL-31及びOSMの影響を示す。
【図10】ケラチノサイト移動に対するIL-31及びOSMの影響を示す。
【図11】S100A7、hBD4/2及びKRT10 mRNA発現に対するいくつかのサイトカインのカクテルの影響を示す。
【特許請求の範囲】
【請求項1】
表皮修復及び/又は皮膚の先天免疫の改善用の組成物の製造のための、サブユニットとしてOSMRβを有するヘテロマー受容体の少なくとも1つの活性化物質の使用。
【請求項2】
前記活性化物質がサイトカインである請求項1に記載の使用。
【請求項3】
前記活性化物質がオンコスタチンM (OSM)又はそのアゴニストである請求項1又は2に記載の使用。
【請求項4】
前記組成物が、IL-17及び/又はTNFα及び/又はそれらのアゴニストをさらに含む請求項3に記載の使用。
【請求項5】
表皮修復及び/又は皮膚の先天免疫の改善用の組成物の製造のための、OSMとIL-17とTNFαとが組み合わせで用いられる請求項1〜4のいずれか1項に記載の使用。
【請求項6】
前記活性化物質がIL-31又はそのアゴニストである請求項1又は2に記載の使用。
【請求項7】
前記組成物が、IFN-γ又はそのアゴニストをさらに含む請求項6に記載の使用。
【請求項8】
前記組成物が、ケラチノサイト移動促進のために用いられる請求項1〜7のいずれか1項に記載の使用。
【請求項9】
前記組成物が、表皮の厚みの増加のために用いられる請求項1〜8のいずれか1項に記載の使用。
【請求項10】
前記組成物が、表皮の治癒促進のために用いられる請求項1〜9のいずれか1項に記載の使用。
【請求項11】
前記組成物が、手、口唇、顔又は体のひび割れを予防するか及び/又は減じるために用いられる請求項1〜10のいずれか1項に記載の使用。
【請求項12】
前記組成物が、伸展裂創を予防するか及び/又は減じるために用いられる請求項1〜11のいずれか1項に記載の使用。
【請求項13】
前記組成物が、瘢痕の外観及び快適さを改善するために用いられる請求項1〜12のいずれか1項に記載の使用。
【請求項14】
前記組成物が、表皮創傷の瘢痕形成の間の表皮創傷の外観及び快適さを改善するために用いられる請求項1〜13のいずれか1項に記載の使用。
【請求項15】
局所投与用の組成物を製造するための請求項1〜14のいずれか1項に記載の使用。
【請求項16】
前記組成物が、再構成されたヒト表皮の厚みをインビトロで増加させるために用いられる請求項1〜7のいずれか1項に記載の使用。
【請求項17】
OSM又はそのアゴニストを含むことを特徴とする化粧用組成物。
【請求項18】
OSM又はそのアゴニストと、IL-17、TNFα及びそれらのアゴニストからなる群より選択される少なくとも1種のサイトカインとを含むことを特徴とする化粧用及び/又は皮膚用
組成物。
【請求項19】
OSM、IL-17及びTNFαを含むことを特徴とする請求項18に記載の化粧用及び/又は皮膚用組成物。
【請求項20】
IL-31を含むことを特徴とする化粧用及び/又は皮膚用組成物。
【請求項21】
OSM及び/又はガンマインターフェロンをさらに含むことを特徴とする請求項20に記載の化粧用及び/又は皮膚用組成物。
【請求項22】
OSMが0.1〜100 ng/mlの濃度であり、及び/又はIL-31が1〜500 ng/mlの濃度である請求項17〜21のいずれか1項に記載の組成物。
【請求項23】
OSMとIL-17及び/又はTNFαとが、0.01〜10 ng/ml、好ましくは0.1〜2 ng/mlの濃度である請求項18又は19に記載の組成物。
【請求項24】
請求項1〜3及び6のいずれか1項で定義される少なくとも1つの活性化物質を培養ケラチノサイトに加える工程を含むことを特徴とする、再構成されたヒト表皮を作製する方法。
【請求項25】
OSM、IL-17及びTNFαが培養ケラチノサイトに加えられる請求項24に記載の方法。
【請求項26】
OSM及び/又はIL-17及び/又はTNFαが0.01〜10 ng/mlの濃度で加えられる請求項24又は25に記載の方法。
【請求項1】
表皮修復及び/又は皮膚の先天免疫の改善用の組成物の製造のための、サブユニットとしてOSMRβを有するヘテロマー受容体の少なくとも1つの活性化物質の使用。
【請求項2】
前記活性化物質がサイトカインである請求項1に記載の使用。
【請求項3】
前記活性化物質がオンコスタチンM (OSM)又はそのアゴニストである請求項1又は2に記載の使用。
【請求項4】
前記組成物が、IL-17及び/又はTNFα及び/又はそれらのアゴニストをさらに含む請求項3に記載の使用。
【請求項5】
表皮修復及び/又は皮膚の先天免疫の改善用の組成物の製造のための、OSMとIL-17とTNFαとが組み合わせで用いられる請求項1〜4のいずれか1項に記載の使用。
【請求項6】
前記活性化物質がIL-31又はそのアゴニストである請求項1又は2に記載の使用。
【請求項7】
前記組成物が、IFN-γ又はそのアゴニストをさらに含む請求項6に記載の使用。
【請求項8】
前記組成物が、ケラチノサイト移動促進のために用いられる請求項1〜7のいずれか1項に記載の使用。
【請求項9】
前記組成物が、表皮の厚みの増加のために用いられる請求項1〜8のいずれか1項に記載の使用。
【請求項10】
前記組成物が、表皮の治癒促進のために用いられる請求項1〜9のいずれか1項に記載の使用。
【請求項11】
前記組成物が、手、口唇、顔又は体のひび割れを予防するか及び/又は減じるために用いられる請求項1〜10のいずれか1項に記載の使用。
【請求項12】
前記組成物が、伸展裂創を予防するか及び/又は減じるために用いられる請求項1〜11のいずれか1項に記載の使用。
【請求項13】
前記組成物が、瘢痕の外観及び快適さを改善するために用いられる請求項1〜12のいずれか1項に記載の使用。
【請求項14】
前記組成物が、表皮創傷の瘢痕形成の間の表皮創傷の外観及び快適さを改善するために用いられる請求項1〜13のいずれか1項に記載の使用。
【請求項15】
局所投与用の組成物を製造するための請求項1〜14のいずれか1項に記載の使用。
【請求項16】
前記組成物が、再構成されたヒト表皮の厚みをインビトロで増加させるために用いられる請求項1〜7のいずれか1項に記載の使用。
【請求項17】
OSM又はそのアゴニストを含むことを特徴とする化粧用組成物。
【請求項18】
OSM又はそのアゴニストと、IL-17、TNFα及びそれらのアゴニストからなる群より選択される少なくとも1種のサイトカインとを含むことを特徴とする化粧用及び/又は皮膚用
組成物。
【請求項19】
OSM、IL-17及びTNFαを含むことを特徴とする請求項18に記載の化粧用及び/又は皮膚用組成物。
【請求項20】
IL-31を含むことを特徴とする化粧用及び/又は皮膚用組成物。
【請求項21】
OSM及び/又はガンマインターフェロンをさらに含むことを特徴とする請求項20に記載の化粧用及び/又は皮膚用組成物。
【請求項22】
OSMが0.1〜100 ng/mlの濃度であり、及び/又はIL-31が1〜500 ng/mlの濃度である請求項17〜21のいずれか1項に記載の組成物。
【請求項23】
OSMとIL-17及び/又はTNFαとが、0.01〜10 ng/ml、好ましくは0.1〜2 ng/mlの濃度である請求項18又は19に記載の組成物。
【請求項24】
請求項1〜3及び6のいずれか1項で定義される少なくとも1つの活性化物質を培養ケラチノサイトに加える工程を含むことを特徴とする、再構成されたヒト表皮を作製する方法。
【請求項25】
OSM、IL-17及びTNFαが培養ケラチノサイトに加えられる請求項24に記載の方法。
【請求項26】
OSM及び/又はIL-17及び/又はTNFαが0.01〜10 ng/mlの濃度で加えられる請求項24又は25に記載の方法。
【図1】


【図2】

【図3】

【図4】

【図5】

【図6】

【図7】

【図8】
【図9】

【図10】
【図11】

【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【公表番号】特表2008−524155(P2008−524155A)
【公表日】平成20年7月10日(2008.7.10)
【国際特許分類】
【出願番号】特願2007−545987(P2007−545987)
【出願日】平成17年12月15日(2005.12.15)
【国際出願番号】PCT/EP2005/014199
【国際公開番号】WO2006/063865
【国際公開日】平成18年6月22日(2006.6.22)
【出願人】(507195140)ユニヴェルシテ ダンジェー (1)
【氏名又は名称原語表記】UNIVERSITE D’ANGERS
【住所又は居所原語表記】40 Rue De Rennes,F−49100 Angers,FRANCE
【出願人】(507195151)ユニヴェルシテ デ ポワチエ (1)
【氏名又は名称原語表記】UNIVERSITE DE POITIERS
【住所又は居所原語表記】Hotel Pinet,15,Rue De L’hotel Dieu,F−86034 Poitiers Cedex,FRANCE
【出願人】(507195162)バイオアルターネイティブス エスエーエス (1)
【氏名又は名称原語表記】BIOALTERNATIVES SAS
【住所又は居所原語表記】1bis,Rue Des Plantes,F−86160 Gencay,FRANCE
【Fターム(参考)】
【公表日】平成20年7月10日(2008.7.10)
【国際特許分類】
【出願日】平成17年12月15日(2005.12.15)
【国際出願番号】PCT/EP2005/014199
【国際公開番号】WO2006/063865
【国際公開日】平成18年6月22日(2006.6.22)
【出願人】(507195140)ユニヴェルシテ ダンジェー (1)
【氏名又は名称原語表記】UNIVERSITE D’ANGERS
【住所又は居所原語表記】40 Rue De Rennes,F−49100 Angers,FRANCE
【出願人】(507195151)ユニヴェルシテ デ ポワチエ (1)
【氏名又は名称原語表記】UNIVERSITE DE POITIERS
【住所又は居所原語表記】Hotel Pinet,15,Rue De L’hotel Dieu,F−86034 Poitiers Cedex,FRANCE
【出願人】(507195162)バイオアルターネイティブス エスエーエス (1)
【氏名又は名称原語表記】BIOALTERNATIVES SAS
【住所又は居所原語表記】1bis,Rue Des Plantes,F−86160 Gencay,FRANCE
【Fターム(参考)】
[ Back to top ]