
ヒトK+イオンチャンネルおよびその治療的適用
【課題】細胞増殖に対し明白な割当てが可能な細胞周期分裂に対する様々な効果を有するカリウムチャンネルの集合の中の生物学的な成分を同定すること。
【解決手段】核酸分子の望まれていない発現または過剰発現によって引き起こされる疾患の治療用薬物の設計方法であって、
(a)特異的かつ効力の高い薬物を同定する工程;
(b)部位特異的突然変異誘発およびキメラタンパク質研究による、前記薬物の結合部位を同定する工程;
(c)(ポリ)ペプチドの結合部位および前記薬物の構造の両方を分子モデリングする工程;ならびに
(d)(ポリ)ペプチドに対するその結合特異性を改良するために薬物を改変する工程、
を含み、該核酸分子が、
(A)配列番号:3または4のアミノ酸配列を有するポリペプチドをコードする核酸分子を含有してなる核酸分子;
(B)配列番号:13または14のDNA配列を有する核酸分子を含有してなる核酸分子;
(C)(A)または(B)の核酸分子の相補的な鎖にハイブリダイズする核酸分子;または
(D)(C)の核酸分子の配列に縮重した核酸分子、
である、ヒトK+ イオンeagチャンネルの機能を有する(ポリ)ペプチドをコードする核酸分子を含有してなる、治療用薬物の設計方法。
【解決手段】核酸分子の望まれていない発現または過剰発現によって引き起こされる疾患の治療用薬物の設計方法であって、
(a)特異的かつ効力の高い薬物を同定する工程;
(b)部位特異的突然変異誘発およびキメラタンパク質研究による、前記薬物の結合部位を同定する工程;
(c)(ポリ)ペプチドの結合部位および前記薬物の構造の両方を分子モデリングする工程;ならびに
(d)(ポリ)ペプチドに対するその結合特異性を改良するために薬物を改変する工程、
を含み、該核酸分子が、
(A)配列番号:3または4のアミノ酸配列を有するポリペプチドをコードする核酸分子を含有してなる核酸分子;
(B)配列番号:13または14のDNA配列を有する核酸分子を含有してなる核酸分子;
(C)(A)または(B)の核酸分子の相補的な鎖にハイブリダイズする核酸分子;または
(D)(C)の核酸分子の配列に縮重した核酸分子、
である、ヒトK+ イオンeagチャンネルの機能を有する(ポリ)ペプチドをコードする核酸分子を含有してなる、治療用薬物の設計方法。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、新規なヒトK+ イオンチャンネル、それをコードする核酸分子および該核酸分子を含有してなるベクターに関する。本発明は、さらに、新規なK+ イオンチャンネルに特異的に生じさせた抗体、ならびに上記化合物の少なくとも1つを含む医薬組成物および診断用キットに関する。さらに、本発明は、本発明のポリペプチドの機能不全または本発明の核酸分子の(過剰)発現によって引き起こされる疾患を治療する方法に関し、該方法は、前記(過剰)発現もしくはイオンチャンネル機能のインヒビター、または該機能不全を排除するインヒビターを、それの必要な患者に投与する工程を含む。前記疾患を治療または予防するための薬物を案出する方法、細胞増殖を阻害する方法および癌を予知する方法は、本発明により含まれるさらなる態様である。本発明はまた、例えば癌に関連するまたは神経変性疾患での、望まれていない細胞増殖を阻害するための本発明の核酸分子に基づいた特異的なアンチセンス治療または遺伝子治療も認識する。
【背景技術】
【0002】
カリウムチャンネルは細胞の静止電位の調節に関連する因子であり、これは興奮性および非興奮性組織でのそれらの主要な役割とみなされている。一方、それらの偏在性の存在およびそれらの特性における特徴的な変異性に対する説明は、不明瞭なままである。カリウムチャンネルは、それらがさらにいくつかの「ハウスキーピング」の役割、例えば細胞増殖1 での「ハウスキーピング」の役割を有するため、全ての細胞型で存在するということは、妥当な仮説である。細胞分裂周期の調節でのそれらの密接な関係がたびたび試験され、いくつかの実験的な証拠が存在している2,3 。しかしながら、とりわけ、細胞周期の間の膜電位の脱分極および過分極の両方が細胞型に依存すると報告されているため1,4 、細胞周期におけるカリウムチャンネルの機能を説明する一般的なモデルはない。K+ チャンネルの役割を説明するために、2つの機構が提案されている:それらは、細胞内でのCa2+濃度に影響を及ぼすか、または細胞容積を制御するかのどちらかである(17、18)。両機構は、間接的に細胞増殖に影響を及ぼすであろう。eagファミリーのメンバーはまた、癌細胞で優先的に発現されると提案されている(19)。数種のカリウムチャンネルブロッカーが癌細胞増殖をブロック(block)する能力に対して試験されており、それらのいくつかは、腫瘍化学治療に対するコアジュバントとして、特に複数の薬物に耐性のある腫瘍において使用されている。それにもかかわらず、細胞増殖の制御に直接関係する特定のカリウムチャンネルの同定の不足は、現在でも、より特異的で効果的な治療プロトコルの記述を妨げている。
【発明の概要】
【発明が解決しようとする課題】
【0003】
したがって、本発明の基礎となる技術的な問題は、ヒト細胞増殖に対する特定の観点から、細胞増殖に対し明白な割当てが可能な細胞周期分裂に対する様々な効果を有するカリウムチャンネルの集合の中の生物学的な成分を同定することであった。かかる技術問題に対する解答は、特許請求の範囲で特徴づけられる態様を提供することにより達成される。
【課題を解決するための手段】
【0004】
即ち、本発明の要旨は、
核酸分子の望まれていない発現または過剰発現によって引き起こされる疾患の治療用薬物の設計方法であって、
(a)特異的かつ効力の高い薬物を同定する工程;
(b)部位特異的突然変異誘発およびキメラタンパク質研究による、前記薬物の結合部位を同定する工程;
(c)(ポリ)ペプチドの結合部位および前記薬物の構造の両方を分子モデリングする工程;ならびに
(d)(ポリ)ペプチドに対するその結合特異性を改良するために薬物を改変する工程、
を含み、該核酸分子が、
(A)配列番号:3または4のアミノ酸配列を有するポリペプチドをコードする核酸分子を含有してなる核酸分子;
(B)配列番号:13または14のDNA配列を有する核酸分子を含有してなる核酸分子;
(C)(A)または(B)の核酸分子の相補的な鎖にハイブリダイズする核酸分子;または
(D)(C)の核酸分子の配列に縮重した核酸分子、
である、ヒトK+ イオンeagチャンネルの機能を有する(ポリ)ペプチドをコードする核酸分子を含有してなる、治療用薬物の設計方法
に関する。
【発明の効果】
【0005】
本発明により、細胞増殖に対し明白な割当てが可能な細胞周期分裂に対する様々な効果を有するカリウムチャンネルの集合の中の生物学的な成分が同定される。
【図面の簡単な説明】
【0006】
【図1】Figure 1。機能時間での野生型(円)およびreag発現CHO細胞の増殖。細胞を96穴ディッシュで培養し、示された時間にテトラゾリウム塩MTT6 (50μg/ml)をプレートに添加した。加湿雰囲気下に(37℃、5%CO2 )4時間インキュベーション後、2容量の1M HCl中10%SDSを添加することにより反応を止めた。生存細胞で産生された青色ホルマザン結晶を1晩かけて可溶化し、得られた色調を指示された波長での光学密度として測定した。細胞増殖のトランスフェクションの起こり得る非特異的影響は無視できる。なぜなら、a)異なる種由来の3つの独立した細胞株で(ラット、ハムスターおよびヒト)結果が類似した;b)異なる独立したクローンでのトランスフェクションが同一の結果を与えた;およびc)同じベクターでの異なるカリウムチャンネル(Kv1.4)でのトランスフェクション(したがって、同じ部位で再結合する傾向を有する)がWTと匹敵する結果を与え、reagトランスフェクションの効果を再現しなかったからである。
【図2】Figure 2。0.5%FCS存在下での、野生型(円)およびreag発現(三角)CHO細胞の増殖。この血清濃度では正常細胞の成長を維持できないが、トランスフェクトされた細胞ではほとんど3周期完了する。方法はFigure 1と同様。
【図3】Figure 3。通常濃度(10%)または低濃度(0.5%)のFCSの存在下での、異なるカリウムチャンネルを発現するCHO細胞でのDNA合成。対照細胞、WTまたはKv1.4でトランスフェクトされた細胞で、DNA合成レベルは低血清濃度の存在下で有意に下がり、一方、reag発現細胞は、高血清濃度と同様の複製レベルを維持した。
【図4a】Figure 4。(A)野生型、Kv1.4トランスフェクトまたはreagトランスフェクトNIH3T3細胞を有するプレートの写真。細胞を低密度で播種し、標準条件下で野生型細胞をコンフルエンスに達するまで成長させた。細胞を次いでメタノールで固定し、ギムエザブルーで染色した。これらの条件下で、野生型およびKv1.4発現細胞は両方とも単層で成長し、一方、reag発現細胞は、フォーカスを形成した。(B)rKv1.4でトランスフェクトされた細胞および野生型細胞と比較したreagトランスフェクトNIH−3T3細胞のフォーカス形成。ベクター対照(pcDNA3トランスフェクト細胞)は、野生型細胞と同様の表現型を与えた(示さず)。一過性のトランスフェクションをリン酸カルシウムを用いて行なった(33)。対照細胞がコンフルエンスに達するまで細胞をリッチ培地で維持し、次いでメタノールで固定し、ギムエザブルーで染色した。
【図4b】Figure 4。(A)野生型、Kv1.4トランスフェクトまたはreagトランスフェクトNIH3T3細胞を有するプレートの写真。細胞を低密度で播種し、標準条件下で野生型細胞をコンフルエンスに達するまで成長させた。細胞を次いでメタノールで固定し、ギムエザブルーで染色した。これらの条件下で、野生型およびKv1.4発現細胞は両方とも単層で成長し、一方、reag発現細胞は、フォーカスを形成した。(B)rKv1.4でトランスフェクトされた細胞および野生型細胞と比較したreagトランスフェクトNIH−3T3細胞のフォーカス形成。ベクター対照(pcDNA3トランスフェクト細胞)は、野生型細胞と同様の表現型を与えた(示さず)。一過性のトランスフェクションをリン酸カルシウムを用いて行なった(33)。対照細胞がコンフルエンスに達するまで細胞をリッチ培地で維持し、次いでメタノールで固定し、ギムエザブルーで染色した。
【図5】Figure 5。電圧クランプ条件下でのMCF7細胞の脱分極により導かれる電流。左のトレースは全体の細胞電流であり、右のトレースは削除されたアウトサイドアウトパッチにより得られる。全体的な電流およびI−V関係の両方が(CおよびD)reag電流を暗示する。
【図6】Figure 6。5μMアステミゾールの存在下または非存在下での、電圧クランプされたアウトサイドアウト膜パッチの0mVでのシングルチャンネル活性。ピペット溶液は、140mM KCl、10mM BAPTA、10mM HEPES pH 7.2を含む;浴溶液は、140mM NaCl、2mM CaCl2 、2mM MgCl2 、2.5mM KCl、10 HEPES pH 7.2を含む。
【図7】Figure 7。A.異なるeagブロッカー下での、MCF7細胞のDNA合成。B.アステミゾール、グリベンクルアミド(glibenclamide)およびテルフェナジンの存在下でのHEK293DNA合成レベル。
【図8】Figure 8。MCF7細胞でのDNA合成の2つのH1アンタゴニストの効果に対する投与量−応答曲線(LY 91241およびアステミゾールそれぞれに対して7mMおよび10mMのIC50)。
【図9】Figure 9。ヘキスト33342で染色された、対照(処理なし、A)およびアステミゾール処理(B)MCF7細胞の蛍光像。Bに、細胞核の同様の表面、および非常に低い細胞密度(細胞死による)を認める。
【図10−1】Figure 10。ラット配列およびウシ配列と比較したヒト脳由来のヒトeag cDNAのヌクレオチド配列。いずれかの配列で異なったヌクレオチドを示す位置に影をつけた。
【図10−2】Figure 10。ラット配列およびウシ配列と比較したヒト脳由来のヒトeag cDNAのヌクレオチド配列。いずれかの配列で異なったヌクレオチドを示す位置に影をつけた。
【図10−3】Figure 10。ラット配列およびウシ配列と比較したヒト脳由来のヒトeag cDNAのヌクレオチド配列。いずれかの配列で異なったヌクレオチドを示す位置に影をつけた。
【図10−4】Figure 10。ラット配列およびウシ配列と比較したヒト脳由来のヒトeag cDNAのヌクレオチド配列。いずれかの配列で異なったヌクレオチドを示す位置に影をつけた。
【図10−5】Figure 10。ラット配列およびウシ配列と比較したヒト脳由来のヒトeag cDNAのヌクレオチド配列。いずれかの配列で異なったヌクレオチドを示す位置に影をつけた。
【図10−6】Figure 10。ラット配列およびウシ配列と比較したヒト脳由来のヒトeag cDNAのヌクレオチド配列。いずれかの配列で異なったヌクレオチドを示す位置に影をつけた。
【図10−7】Figure 10。ラット配列およびウシ配列と比較したヒト脳由来のヒトeag cDNAのヌクレオチド配列。いずれかの配列で異なったヌクレオチドを示す位置に影をつけた。
【図10−8】Figure 10。ラット配列およびウシ配列と比較したヒト脳由来のヒトeag cDNAのヌクレオチド配列。いずれかの配列で異なったヌクレオチドを示す位置に影をつけた。
【図10−9】Figure 10。ラット配列およびウシ配列と比較したヒト脳由来のヒトeag cDNAのヌクレオチド配列。いずれかの配列で異なったヌクレオチドを示す位置に影をつけた。
【図10−10】Figure 10。ラット配列およびウシ配列と比較したヒト脳由来のヒトeag cDNAのヌクレオチド配列。いずれかの配列で異なったヌクレオチドを示す位置に影をつけた。
【図10−11】Figure 10。ラット配列およびウシ配列と比較したヒト脳由来のヒトeag cDNAのヌクレオチド配列。いずれかの配列で異なったヌクレオチドを示す位置に影をつけた。
【図11−1】Figure 11。対応するウシ、マウスおよびラット配列と比較した、ヒトeag cDNA転写から得られる両スプライス変異体のアミノ酸配列。黒いボックスは、いずれかの配列での異なる残基を示す。
【図11−2】Figure 11。対応するウシ、マウスおよびラット配列と比較した、ヒトeag cDNA転写から得られる両スプライス変異体のアミノ酸配列。黒いボックスは、いずれかの配列での異なる残基を示す。
【図11−3】Figure 11。対応するウシ、マウスおよびラット配列と比較した、ヒトeag cDNA転写から得られる両スプライス変異体のアミノ酸配列。黒いボックスは、いずれかの配列での異なる残基を示す。
【図11−4】Figure 11。対応するウシ、マウスおよびラット配列と比較した、ヒトeag cDNA転写から得られる両スプライス変異体のアミノ酸配列。黒いボックスは、いずれかの配列での異なる残基を示す。
【図11−5】Figure 11。対応するウシ、マウスおよびラット配列と比較した、ヒトeag cDNA転写から得られる両スプライス変異体のアミノ酸配列。黒いボックスは、いずれかの配列での異なる残基を示す。
【図11−6】Figure 11。対応するウシ、マウスおよびラット配列と比較した、ヒトeag cDNA転写から得られる両スプライス変異体のアミノ酸配列。黒いボックスは、いずれかの配列での異なる残基を示す。
【図11−7】Figure 11。対応するウシ、マウスおよびラット配列と比較した、ヒトeag cDNA転写から得られる両スプライス変異体のアミノ酸配列。黒いボックスは、いずれかの配列での異なる残基を示す。
【図11−8】Figure 11。対応するウシ、マウスおよびラット配列と比較した、ヒトeag cDNA転写から得られる両スプライス変異体のアミノ酸配列。黒いボックスは、いずれかの配列での異なる残基を示す。
【図11−9】Figure 11。対応するウシ、マウスおよびラット配列と比較した、ヒトeag cDNA転写から得られる両スプライス変異体のアミノ酸配列。黒いボックスは、いずれかの配列での異なる残基を示す。
【図11−10】Figure 11。対応するウシ、マウスおよびラット配列と比較した、ヒトeag cDNA転写から得られる両スプライス変異体のアミノ酸配列。黒いボックスは、いずれかの配列での異なる残基を示す。
【図12】Figure 12。ヒト脳、ヒト乳腺およびMCF−7細胞の全RNA由来のRT−PCR。増幅は、脳で予期されるサイズのheag1およびheag2に対応する2つの特異的な断片を提供し、MCF−7細胞でheag1に対応するバンドを提供し、一方、正常胸RNAで増幅は検出されなかった。
【図13】Figure 13。高い細胞外カリウムでの活性化の電圧依存、2電極電圧クランプ:コンダクタンス−電圧プロットにおいて、半活性化に対する電圧は、reagチャンネルに関してheagチャンネルで40mV右にシフトする。
【図14】Figure 14。示されたDNAでトランスフェクトされたNIH−3T3細胞の半固体培地でのコロニー形成。細胞を0.55%寒天培地の層の上に0.3%寒天を含むレギュラー培地で培養した。直径0.1mmより大きいコロニーをトランスフェクションの14日後に評価した。顕微鏡範囲で少なくとも10を数えるコロニーの平均数は、トランスフェクションに用いられるDNA1μg当たりで表した(「トランスフェクション緩衝液」および「処理なし」のレーンを除く、ここで数字は絶対値である)。reagおよびKv1.4はpcDNA3またはpTracerをCMVベクターのどちらかを使用してトランスフェクトした。
【図15】Figure 15。(A)異なるヒト組織および293細胞由来のRNAのRT−PCR産物のサザンブロット。トランスフェリンレセプター(TFR)シグナルが底に示される。(B)異なるヒト細胞株および一次培養での乳上皮細胞(Epith.cell)由来の全RNAのRT−PCR産物のサザンブロット解析。TRFシグナルが底に示される。
【図16】Figure 16。(A)アンチセンスODNでの、heag発現腫瘍細胞株の処理。(B)SHSY−5Y神経芽腫細胞でのheag電流。(C)アンチセンスODNで処理されたSHSY−5Y細胞での電流密度。(D)heagに特異的なアンチセンスODNによる、ヒト癌細胞(EFM−19、HeLaおよびSHSY−5Y)でのDNA合成の阻害。
【図17−1】Figure 17。(A)速く成長してキャリアマウスの皮膚をすぐに傷つける攻撃的な腫瘍を誘導するCHOhEAG細胞の皮下埋め込み。2×106 細胞の埋め込み後3週間で撮られた写真。(B、C)埋め込み後2週間(B;平均±S.E.M.;p=0.002)または3週間(C;平均±S.E.M.;p=0.03)の両方で、CHOhEAG腫瘍の平均の質量はCHOKv腫瘍のものより有意に大きかった。インサイチュで写真を撮られた(D)CHOhEAG腫瘍および(E)CHOKv腫瘍。主に肉眼で見える相違はより濃い色およびCHOhEAG腫瘍の皮膚への固定である。(F、G)CHOhEAG(F)およびCHOKv(G)腫瘍を切り開き、前者でより広い範囲の壊死を示した(矢頭)。(H、I)より大きな程度の壊死および皮膚への固定がまた、パラフィン包理およびヘマトキシリン−エオシン染色後、顕微鏡的に明らかである。組織構造は両方の顕微鏡写真で匹敵するが、(H)で非常に大きな壊死面積が観察され(矢頭)、皮下脂肪と腫瘍の間に境界はない。(目盛りバー、100μm)。(J)これらの像の定量的測定では、CHOKv腫瘍での生命範囲の平均の広さは、CHOhEAG腫瘍のものより有意に大きかった(平均±S.E.M.;p<0.0005)。
【図17−2】Figure 17。(A)速く成長してキャリアマウスの皮膚をすぐに傷つける攻撃的な腫瘍を誘導するCHOhEAG細胞の皮下埋め込み。2×106 細胞の埋め込み後3週間で撮られた写真。(B、C)埋め込み後2週間(B;平均±S.E.M.;p=0.002)または3週間(C;平均±S.E.M.;p=0.03)の両方で、CHOhEAG腫瘍の平均の質量はCHOKv腫瘍のものより有意に大きかった。インサイチュで写真を撮られた(D)CHOhEAG腫瘍および(E)CHOKv腫瘍。主に肉眼で見える相違はより濃い色およびCHOhEAG腫瘍の皮膚への固定である。(F、G)CHOhEAG(F)およびCHOKv(G)腫瘍を切り開き、前者でより広い範囲の壊死を示した(矢頭)。(H、I)より大きな程度の壊死および皮膚への固定がまた、パラフィン包理およびヘマトキシリン−エオシン染色後、顕微鏡的に明らかである。組織構造は両方の顕微鏡写真で匹敵するが、(H)で非常に大きな壊死面積が観察され(矢頭)、皮下脂肪と腫瘍の間に境界はない。(目盛りバー、100μm)。(J)これらの像の定量的測定では、CHOKv腫瘍での生命範囲の平均の広さは、CHOhEAG腫瘍のものより有意に大きかった(平均±S.E.M.;p<0.0005)。
【図18】Figure 18。rEAGトランスフェクトCHO細胞の増殖アッセイ(A〜C)。10%(黒塗りの記号)または0.5%(白抜きの記号)のウシ胎仔血清での、処理なしの細胞(三角)と比較したrEAGでトランスフェクトされたCHO細胞(円)の成長曲線。値は、培養12時間後測定されたものをいい(プロットで時間0)、同じプレートで8ウェルの平均±S.E.M.を表す。細胞株をpcDNA3ベクターでコードされるG−418耐性を介する選択により確立した。MTT加水分解(22)を使用して、生存能力がある細胞の代謝活性を測定した。12時間培養後、血清を注意深く希釈した。(B)S期ブロックの除去後最初の12時間の間の代謝活性の増加。細胞同調のために、12時間培養培地にチミジン2mMを添加した。追加の12時間、培地からチミジンを除去し、次いで第2停止パルスを12時間適用した。代謝活性およびDNA合成の測定のために、次いで細胞をトリプシン処理し培養した。(C)10%FCSで12時間のインキュベーション、または0.5%FCSの存在下(24時間のインキュベーション)でのS期ブロックの除去後最初の12時間の間のBrdU取り込み。BrdUの取り込みを、製造業者の指示にしたがってBoehringer−Mannheimの「BrdUラベリングおよび検出キット」を使用して、測定した。バーは、野生型CHO細胞(白抜きバー)、Kv1.4トランスフェクト(影をつけたバー)およびeag−トランスフェクト(中実バー)に対する平均±S.D.を表す。BrdUの取り込みを、抗BrdU抗体に結合するペルオキシダーゼによりABTSTM基質で生じる405nm(参照490nm)での光学密度として定量する。
【発明を実施するための形態】
【0007】
それゆえに、本発明は、
(a)配列番号:3または4のアミノ酸配列を有するポリペプチドをコードする核酸分子を含有してなる核酸分子;
(b)配列番号:13または14のDNA配列を有する核酸分子を含有してなる核酸分子;
(c)(a)または(b)の核酸分子の相補的な鎖にハイブリダイズする核酸分子;または
(d)(c)の核酸分子の配列に縮重した核酸分子、
である、ヒトK+ イオンeagチャンネルの機能を有する(ポリ)ペプチドをコードする核酸分子を含有してなる核酸分子に関する。
【0008】
本発明の核酸分子は、ラットeagチャンネルのヒトホモローグであるかまたはそれを含有する(ポリ)ペプチドをコードする。この点において、用語「ヒトK+ イオンeagチャンネルの機能を有する(ポリ)ペプチドをコードする核酸分子を含有してなる核酸分子」は、前記最初に記載された核酸分子が前記(ポリ)ペプチドを単にコードすることを意味してもよい。したがって、それは前記2番目に記載された核酸分子と同一であってもよい。他方、それらは、調節領域または他の非翻訳領域を含有してもよい。さらなる態様において、前記最初に記載された核酸は、異種構造のタンパク質材料、したがって、例えば、融合タンパク質を生じさせる材料をコードしてよい異種構造の核酸を含んでいてよい。配列番号:13および14のDNA配列は、本発明の(ポリ)ペプチドをコードする核酸配列のスプライス変異体(variants)であることを、さらに指摘する。対応するアミノ酸配列を、配列番号:3および4に記載する。
【0009】
用語「ヒトK+ イオンeagチャンネルの機能を有する」は、本発明に関連して使用され、次の意味を有する:チャンネルは、約6pSの0mVで、非対称のカリウムでのシングルチャンネルコンダクタンスを有する。この値は、約7pSの値が測定されるラットチャンネルからヒトチャンネルを明確に区別する。さらにまたは他方、上記用語は、次の意味を有していてもよい:reagに対する400μMと比較して、アフリカツメガエル卵母細胞で発現される時、キニジンに対し約1mMのIC50を有する。さらに、2電極電圧クランプを用いて高い細胞外カリウムでの活性化の電圧依存性を測定する場合、コンダクタンス−電圧プロットにおいて、半活性化に対する電圧は、reagチャンネルに関してheagチャンネルで約40mV以上右にシフトすることが見出された(Figure 13を参照のこと)。前記特徴の単一または組み合わせに基づいて、、本発明のヒトイオンチャンネルと、先行技術のチャンネル、特にラットイオンチャンネルとの間の機能に基づく区別は、当業者のさらなる苦労なしに可能である。好ましくは、チャンネルは全ての列挙された機能を有する。前記値は、本明細書で記載された実験計画で得られ得る値を指す。異なる発現系の使用等の実験パラメーターの変更は、当業者に知られているように、また前記値を変化させる。さらに、これらの態様はまた、本発明の範囲に含まれる。
【0010】
用語「ハイブリダイズする(hybridizing)」は、本発明に従って使用される場合、ストリンジェントまたは非ストリンジェントなハイブリダイゼーション条件に関する。好ましくは、それはストリンジェント条件に関する。かかるハイブリダイゼーション条件は、例えば、Sambrookによる、“Molecular Cloning,A Laboratory Manual”、Cold Spring Harbor Laboratory(1989)N.Y.、Ausubel、“Current Protocols in Molecular Biology”、Green Publishing AssociatesおよびWiley Interscience、N.Y.(1989)、またはHigginsおよびHames(編)“Nucleic acid hybridization,a practical approach”IRL Press Oxford,Washington DC,(1985)に記載された慣用のプロトコルによって確立されてよい。ハイブリダイズする分子または上記もう1つの(d)に該当する分子は、(a)または(b)で同定される分子の断片も含有する、ここでヌクレオチド配列は、配列番号:13または14でのその対照物(counterpart)と同一である必要はなく、前記断片は上記で示されるような機能を有するものである。
【0011】
1つのかかるストリンジェントなハイブリダイゼーション条件の例は、65℃での4XSSC、続く65℃での0.1XSSCでの1時間の洗浄によるハイブリダイゼーションである。他方、典型的なストリンジェントなハイブリダイゼーション条件は、42℃での50%ホルムアミド、4XSSC中である。かかる非ストリンジェントなハイブリダイゼーション条件の例は、50℃での4XSSCまたは42℃での30〜40%ホルムアミドでのハイブリダイゼーションである。分子にハイブリダイズする相補的な鎖は、本発明のポリペプチドの断片、アナログまたは誘導体をコードし、例えば、アミノ酸および/またはヌクレオチドの1つまたは複数の欠失、1つまたは複数の挿入、1つまたは複数の置換、1つまたは複数の付加および/または1つまたは複数の組換えまたは当該分野において既知のいずれかの他の1つまたは複数の修飾の単独または組み合わせを介して、前記アミノ酸配列またはそれらの基礎となる1つまたは複数のヌクレオチド配列と異なるものを含有する。PESTFINDプログラム(Rogers,Science 234(1986)、364〜348)を使用して、不安定なタンパク質に特徴的に存在するPEST配列(プロリン、グルタミン酸、セリンおよびトレオニンがリッチである)を同定できる。かかる配列は、タンパク質の安定性および任意にタンパク質の活性を増加するために、本発明のポリペプチドから除去してもよい。本発明による核酸分子におけるかかる修飾を導入する方法は、当業者によく知られている。本発明はまた、その配列が遺伝コードの縮重のために前記核酸分子のいずれのヌクレオチド配列とも異なっている核酸分子に関する。本発明のタンパク質をコードするかかるすべての断片、アナログおよび誘導体は、前記規定するような必須の特徴的な免疫学的および/または生物学的特性が本質的に影響されないままである限り、本発明の範囲内に含まれ、すなわち、本発明の新規核酸分子は、上記の核酸分子によってコードされ、生物学的活性に関して同等のまたは同一の特徴を有する前記ポリペプチドに対する抗体と反応できる1以上のエピトープに対する少なくとも一部の一次構造形態を有するタンパク質またはペプチドをコードする全ヌクレオチド配列を含む。本発明の一部はそれゆえにまた、本発明による核酸分子に含まれる核酸配列によりコードされる前記同定されたポリペプチドの少なくとも機能的部分を含有してなるポリペプチドをコードする核酸分子に関する。
【0012】
本発明者らは最近、G1−SおよびG2−M移行の両方においてサイクリン依存性キナーゼ(細胞周期調節における重要な分子)の活性化直後に強くダウンレギュレートされるカリウムチャンネル(reag)について記載している5 。サイクリン依存性キナーゼの活性化後に、電圧依存性ナトリウムブロックによりK+ 電流(current)は阻害される。それは細胞周期の全ての段階において明白であるというわけではない。本明細書に存在する実験は、eagがまた、細胞周期により制御されるのに加えて、細胞増殖および細胞成長に直接影響し得るか否かを決定することを意図する(20)。本発明およびヒト細胞での(疾患に関連する)増殖を評価するための適切なシステムの開発の観点にしたがって、細胞周期におけるチャンネルの関係が、細胞周期の進行によって制御されるだけでなく、細胞周期の進行のレギュレーターでもあるというような、両方向に進むかどうかの研究を、さらに試みた。
【0013】
このラット由来のイオンチャンネルシステムで得られた結果は、異なる種から得られた3つの異なる細胞株(チャイニーズハムスター−CHO−、ヒト−HEK293−およびマウス−NIH3T3−)において、チャンネルが、サイトメガロウイルスプロモーターの制御下のチャンネルDNAを含むプラスミドによる細胞のトランスフェクションの後に過剰発現される場合、増殖速度がより速くなることを示す。Figure 1およびFigure 18aは、通常濃度のウシ胎仔血清(10% FCS)の存在下の、CHO細胞の培養物における代謝活性の上昇を示す。これらの通常条件下で、reagトランスフェクト細胞は、非トランスフェクト細胞(WT)より数倍速く成長する。
【0014】
Figure 2は、非常に低濃度のウシ胎仔血清(0.5% FCS)での比較例を示す。これらの低い血清濃度では、野生型細胞が成長できない;数時間後、細胞は死に始める。しかしながら、reagトランスフェクト細胞は、同条件下に増殖可能である。成長因子の欠如により誘導される成長停止に打ち勝つ能力は、悪性の形質転換の1つの典型的な特性である(Figure 18参照)。
【0015】
培養における増殖の追跡のために代謝活性以外のものを使用することができる。(分裂に付される)S期に入る細胞のみがDNAを合成するために、DNA合成の測定は、細胞成長速度のより直接的な評価である。また、DNA合成は、reagトランスフェクト細胞において血清非依存的となる、すなわち、成長が、成長因子の非存在下に維持される(一方、それは、非トランスフェクト細胞のプログラムされた死を誘導する)。これはFigure 3に描かれており、5−ブロモ−2’−デオキシウリジン7-10(BrdU)の取り込みを用いて、CHO細胞において、10%FCSまたは0.5%FCSの存在下にDNA合成をモニターした。野生型またはラット脳由来の不活性化された電圧依存性カリウムチャンネルでトランスフェクトされた細胞(Kv1.4)と対照的に、reag発現細胞で通常FCS濃度または低FCS濃度の存在下に合成されるDNAの量に有意な相違はない。HEK−293細胞で上皮成長因子(EGF)をまたはCHO細胞で血小板由来成長因子(PDGF)を使用して、同様の実験を行ない、本質的に同一の結果を得た。純粋な成長因子を使用して、全血清の使用により導入される複雑さを避けた。
【0016】
細胞増殖に対するeagの効果をより直接的に試験するために、チミジン停止により細胞周期のS期で同時に発生する細胞での5−ブロモ−デオキシウリジン(BrdU)の取り込みを通してDNA合成を測定した(23)。上記知見と一致して、細胞周期のS期が進行できる場合、reag発現CHO細胞(CHOrEAG)は、より高い代謝活性(Fig 18B)およびBrdU取り込みの増加(Fig 18C)を示した。これらの結果は、CHOrEAG細胞でより速い増殖速度を示すいずれの場合においても、分裂停止期の間により多くeagトランスフェクト細胞がS期に入ること、および/またはDNA合成が上昇することを示唆する。低血清の存在下に、BrdU取り込みは、野生型細胞でよりも、CHOrEAGで有意に高かった(Fig 18C)。
【0017】
さらにもう1つの細胞株、NIH3T3は、これらの細胞が非常に強く接触阻害されるため(すなわち、培養がコンフルエンシーに達した時、それらの成長がストップする)、しばしば腫瘍形質転換アッセイに使用されている。これらは、野生型細胞で均一な単層を生じる。この株の悪性の形質転換(癌遺伝子の発現を介する)は、通常この特性の損失を誘導し、NIH3T3細胞はいくつかの層の細胞からなるコロニーを形成し始める。これは、reag DNAによるトランスフェクション後に見られ得、いくつかの非依存性クローンでかかるフォーカス(foci)の形成を誘導した(Fig.4AおよびB)。形質転換活性に対するもう1つの標準的な試験は、付着するための基質が得られない場合にコロニーで成長するNIH3T3細胞の能力である。これを試験するために、細胞を寒天を含む培地で培養する。ここで、寒天は細胞とプレートの表面との間の接触を妨げるであろう。これらの条件下に、野生型NIH3T3細胞は成長できず、一方、reagを発現する細胞は、単にプレートの視覚観察により検出可能でさえある大きなコロニーを形成した。表Iは、reag(しかし、rKv1.4ではない)トランスフェクト細胞は、トランスフェクションで使用されたベクターにかかわらず、0.3%寒天を含む半個体の培地でコロニーを形成した(Fig.14)ことを示す。全ての上記結果は、eagの形質転換能を示す。
【0018】
要するに、トランスフェクトされた細胞から得られた結果は、reagが少なくともある条件下に癌遺伝子特性を示し得ることを示す。
【0019】
一旦、reagの形質転換能力を本発明により決定した後、ヒト癌細胞でのそれぞれのチャンネルの発現を研究した。この研究のために、胸腺癌(breast adenocarcinoma)の胸膜滲出液(pleural effusion)から初めに得られた細胞株MCF−7を使用した。該株は、エストロゲンレセプター陽性ならびにエストロゲン感受性であり、かつ相対的によく分化する。次の戦略は、まずeagと同様の機能的電流の存在について電気生理学的および薬理学的に試験し、次に分子レベルで対応するチャンネルの同定を試みることであった。しかしながら、かかる同定に対する慣用のアプローチで失敗した。
【0020】
すなわち、ほとんどの細胞で電流密度は低過ぎて、細胞電流全体の信頼度の高い測定はできなかった。低い電流密度は、パッチチャンプ(champ)に全細胞形態を使用するチャンネル特性の正確な測定を妨げた。それゆえに、かかる低い電流密度に遭遇したため、もう1つのアプローチに頼った。かかる細胞当たりの低いチャンネル数のために、1つ(または少し)のチャンネルを含む膜のパッチを取り出し、かつ単分子レベルで特徴付けができる特別なパッチチャンプ法により、チャンネルの機能的な特性を特徴付けることができるのみである。このアプローチでは、シングルチャンネルコンダクタンス、薬理学特性、電圧依存性、平均開口時間等の単分子レベルでの特性を比較するためにも、シングルチャンネル測定をあてにした。実際に、ほとんどの膜パッチにおいて速度論、電圧依存性、および薬理学に関してreagと矛盾しないいくつかの特性を有するチャンネルが、したがって同定され得るであろう。Figure 5は、全細胞電流が、電流−電圧関係と共に、ナイスタチンパッチ条件およびシングルチャンネル電流下にMCF7細胞から得られたことを示す。ほとんど脱分極した電圧での速度論での相違にもかかわらず、ヒト細胞でのチャンネルの電圧依存性は、reagチャンネルの電圧依存性に対して高く暗示される。さらに、推定上のヒト−eagのシングルチャンネル特性はまた、reagのものに非常によく似ている。
【0021】
さらに、分子レベルで前記チャンネルを単離する標準アプローチはまた成功しなかった。いくつかの他のグループが、ヒトeagをコードする遺伝子を単離しようと試みてきたおよび/または未だ試みている最中であるが、成功しておらず、これはラットeagチャンネルが、既に1994年に発表されたという事実にもかかわらずである。例えば、WarmkeおよびGanetzky(Proc.Natl.Acad.Sci.USA 91(1994)、3428〜3442)は、慣用の技術を使用してヒトeag遺伝子をクローニングすることについて詳細に説明した。しかしながら、彼らは成功せず、彼らはh−ergと呼ぶ(またはHERGと呼ぶ)新規なeag関連遺伝子をクローニングした。さらに、Wymoreら、Circulation Res.80(1997)、261〜268は、全eag/ergスーパーファミリーに渡って保存される増幅のためのプライマーが使用されるという事実にもかかわらず、eag特異的クローンが、ヒト心臓由来のcDNAライブラリーで検出され得ないことを報告した。したがって、HERGはその分野の他の研究者によって系統的に検出されたが、ファミリーのメンバーの配列に基づく縮重オリゴヌクレオチドを用いる標準的なアプローチは不成功であることを明らかにした。有意に、ヒトeag遺伝子をクローニングするための、ほとんどのこれらのアプローチは、脳のライブラリーを使用してなされた。これらの先行技術データの組み合わせからの結論は、ヒトeag遺伝子は最も明白な源、すなわち脳組織を使用する慣用の技術によってクローニングできないであろうということであった。代わりにHERGクローンの繰り返された同定は、ほとんどたぶん脳ライブラリーでの比較的多数のHERG転写物のためであり、また2つのチャンネル間の高い相同性のためである。それゆえに、異なった戦略が、eagチャンネルに対しより特異的にスクリーニングを行なえるように案出されなければならなかった。最初に、本明細書の前記のように、reagに機能的に類似したチャンネルを発現する細胞株を同定した。ついでラット、ウシおよびマウスのeag間で保存されるが、HERGとは異なる配列に基づいて縮重オリゴヌクレオチドを設計した。これらのプライマーを用いて、PCRによってMCF7細胞から得られたcDNAを増幅し、予期されたサイズのバンドを適切なベクターにクローニングし、配列を決定した。増幅断片は、チャンネルタンパク質のコア領域内の約400bpに対応し、reag配列に対してDNAレベルで90%の同一性を有し、アミノ酸レベルで99%の同一性を有した。しかしながら、この段階では、こうして同定されたクローンが何に対応するのかはまだかなり不明瞭であった。例えば、eagファミリーのさらなるメンバーが同定されたということは、かなりの可能性があった。これは、脳のライブラリーに対する多くの試みにもかかわらず、誰もヒトeag遺伝子をクローニングすることができなかった、およびMCF7株は胸癌由来株である、という事実の観点から、特に真実である。
【0022】
MCF7細胞は不死の細胞であるために、遺伝子の多数が変異すると想定される。最初から、ヒトeagチャンネルは、もし少しでもこの細胞株で発現されるなら、変異されることが予測されたであろう。この仮定の下で、この細胞株は、所望の遺伝子の単離に少しでも使用され得るかどうかは、かなり不確実であった。
【0023】
脳のライブラリーからのヒトeag遺伝子のクローニングの先行技術の失敗および前記不死化した細胞株の不確実性のために、ライブラリーに対するもう1つの源が必要であった。400bpの断片がそれゆえに正常ヒト胸cDNAライブラリーをスクリーニングするために使用された。胸癌細胞でのeagの存在のため、かかるライブラリーはheagクローンを含有することが予期された。しかしながら、驚くべきことに、2×106 ファージのスクリーニングの後、ヒトeagクローンは、前記ライブラリーで同定され得なかった。これは、チャンネルが正常な組織でなく腫瘍細胞でのみ発現されるという可能性を高める。上記ライブラリーから直接PCR増幅によりheag断片について調べるための特異的なオリゴヌクレオチド、すなわち5’−CCAAACACACACACCAGC、5’−CGTGGATGTTATCTTTTTGGを設計したが、このライブラリーでいずれのeagクローンの存在に対する証拠も見出せなかった。上記先行技術の結果を考慮し、さらに驚くべきことに、同じプライマーが正常なヒト脳のcDNAライブラリーでheagを検出し、それゆえにスクリーニングされるようになった。最初に、MCF7細胞から得られたプローブを、106 ファージを調べるのに使用した。この手順により、ヒトeag由来の1.6kbp断片を単離できた。この断片を次に同じライブラリー由来の2×106 ファージのスクリーニングのプローブとして使用した。いくつかの独立したクローンを単離したが、それらのいずれも全長のクローンではなかった。さらに、1つのクローンのみが配列の5’末端を含み、一方、それらの2つが3’末端および3’非コード領域の一部を含んだ。チャンネルをコードする核酸配列の豊富な制限部位が、cDNAのこの多数の断片化を誘導しているようである。例えば、EcoRIをEcoRI部位においてλ−gt10ファージにクローニングされたライブラリーの挿入物を抽出するのに使用した場合、この慣用の研究方法では体系的に分子の5’末端を見つけることはできなかった(クローンの400位にEcoRI部位がある)。それゆえ、プールされた陽性クローンを、PCRによって再度スクリーニングし、開始コドンの増幅を試み、そしてこの手段によってのみ、このATGを含む1ファージを単離することが可能であった。heagの2つのスプライス変異体をクローニングした。両方とも脳組織で発現した。heag 1およびheag 2について得られた配列ならびにそれらの予想アミノ酸配列をFigure 10およびFigure 11に示し、ファミリーの他のメンバーと比較した。
【0024】
予想したアミノ酸配列は、Occhidoro(27)による本発明の優先日の後に公表された配列と同一であり、reagと97.7%同一である。前記のように、第2(81bpさらに長い)のスプライス変異体(heag 2)も単離し、該変異体はウシおよびマウスeagチャンネルで報告されたもの(28)と類似しており、スプライス挿入は3種全てで同一であった。heagの染色体の局在を、FISH検出で測定し(29)、染色体lq32.1〜32.3に位置づけた(参照文献26も参照のこと)。
【0025】
MCF−7癌細胞とは対照的に、heagが正常の乳腺で発現されない可能性をさらに調べるために、我々は、ヒト脳、ヒト乳腺、およびMCF−7細胞由来の全RNAを使用し、heagの2つのスプライス変異体間で区別するように設計された2つのオリゴヌクレオチドをプライマーとして使用して、シングルチューブRT−PCR実験を行なった(Fig 12)。ヒト脳において、2つのスプライス変異体が検出され、一方、短いもののみがMCF−7細胞で発現された(これは、逆転写酵素の非存在下における増幅の欠如と共に、RNA調製物のゲノムDNAによる汚染の可能性を除外する)。heagシグナルは、正常乳腺RNAで、この高感度技術を用いて検出されなかった。予備結果は発現が同じ臓器由来の腫瘍細胞に存在することを示唆したため、この結果は全く予期されなかった。さらに、RT−PCR産物のサザンブロット解析後、乳腺でheagプローブとハイブリダイズする弱いバンドを同定した。したがって、これらの矛盾した実験データを考慮すると、胸のheagメッセージの完全な欠如を強く主張することはかなり困難である。
【0026】
さらに、heagの電気生理学的特性(21,30)を、ツメガエル(Xenopus)卵母細胞で試験した。前記のように、前記例外、例えば両チャンネルが同一の条件下に測定された時の、40mVのより脱分極された電位への活性化のシフトを除いて、それらは、それらのものまたはreagに有意な相違はなかった。ツメガエル卵母細胞で発現されるheagチャンネルの電気生理学的観察は、Bijlengaらにより報告されたホース(31)によく対応する。
【0027】
本発明はまた、配列5’−GGGAGGATGACCATGGCTを含有してなる本発明の核酸分子に特異的にハイブリダイズする核酸分子に関する。
【0028】
本発明のこの態様は、本明細書の下記に(例えば実施例5において)さらに詳細に説明するように、細胞増殖を阻害するための特異的なアンチセンス治療に特に有用である。さらに本発明の核酸分子のこの態様はまた、当然、例えばノーザンブロット技術を用いることにより、組織のheag mRNAを特異的に検出するためのプローブとして使用される。ノーザンブロットによる様々な組織のheag mRNA発現の解析は、脳で約9.2kbの強いハイブリダイゼーションシグナルおよび胎盤で同様のサイズの弱いシグナルを明らかにした。心臓、肺、肝臓、骨格筋、腎臓および膵臓は、長い曝露後でさえ陰性であった。さらに、ヒト脳、心臓、気管、副腎、肝臓、腎臓、骨格筋および乳腺由来の全RNA、および脊髄ポリ(A)+ RNA、ならびにアデノウイルスで形質転換された293株由来の全RNA(ヒト非腫瘍細胞株)をシンングルチューブRT−PCRおよびサザンブロットにより解析した。これらの実験条件下に、heagは脳でのみ検出され、そこで、両スプライス変異体を同定した(Fig.15;実施例3)。
【0029】
第1のcDNAが上皮腫瘍細胞株(MCF−7)から単離され、脳組織から単離されなかった(前記参照)ため、脳におけるheagの優先的な発現は興味深かった。他の腫瘍細胞株でheagの存在を解明するために、全RNAをHeLa(子宮頸癌)、SHSY−5Y(神経芽腫)、および乳腺腫瘍由来の株:COLO−824(癌)、EFM−19(癌)、およびBT−474(管癌)から調製した。脳、MCF−7細胞、293細胞由来の全RNA、および乳腺上皮細胞の培養物(全体の乳腺の混合細胞集団を取り囲むことを含む)由来のRNAを対照として用いた。全細胞株はDSMZ(Deutsche Sammlung von Mikroorganismen und Zellkulturen)より得られ、DSMZカタログガイドラインに従って維持された。正常ヒト乳腺上皮細胞をBio Whittakerから購入した。プライマーをheag 1およびheag 2の異なるバンドを増幅するために設計し、これによりゲノムDNA汚染のために間違って陽性になるのを防ぐことができた(逆転写酵素非存在下での対照も行なった)。HeLa、SHSY−5Y、EFM−19およびMCF−7 RNAはheagバンドを示し、一方、COLO−824およびBT−474シグナルは背景と区別がつかなかった(Fig.15B)。培養された上皮細胞および293細胞は(Fig.15A)、陰性であった。前記のように、reagでトランスフェクトされた細胞は癌遺伝子特性を示し得ることが、本発明にしたがって示され得た。したがって、heagの発現がインビボで腫瘍細胞に有利であるかどうかを測定するために、雌のシッド(scid)(厳しい複合免疫不全、32)マウスの脇腹に、チャンネルを発現するCHO細胞(CHOhEAG細胞)の皮下移植を行ない、それは、CHOhEAG腫瘍が、CHOKv腫瘍よりも速く成長し、より攻撃的であるため、heagの発現がインビボで腫瘍細胞の増殖に有利であることを意味することを示し得た。したがって、本発明の核酸分子の態様は、例えば、癌(特に、乳癌、神経芽腫)、乾癬、アルツハイマー病、パーキンソン病、筋萎縮性側索硬化症または多発性硬化症等の神経変性疾患を示すような組織で検出可能な種々の疾患状態で、ヒトeagの発現レベルの定量的および定性的解析に使用され得る。
【0030】
本発明の核酸分子の好ましい態様において、前記核酸分子はゲノムDNA等のDNAである。一方、本発明はまた、ペプチド核酸等の合成または半合成DNA分子またはその誘導体を含む。最も好ましい本発明のDNA分子は、cDNAである。
【0031】
本発明のさらに好ましい態様において、前記核酸分子はRNA、好ましくは、mRNAである。
【0032】
本発明の核酸分子のもう1つの好ましい態様は、融合タンパク質をコードする。例えば、本発明の核酸分子は、FLAGまたはGFP等の検出可能なマーカーにフレーム中で融合し得る。
【0033】
本発明はさらに、本発明の核酸分子を含有するベクター、特にプラスミド、コスミド、ウイルスおよびバクテリオファージに関する。かかるベクターは、適切な宿主細胞で適切な条件下に該ベクターの選択を可能にするマーカー遺伝子等のさらなる遺伝子を含んでもよい。このように、本発明のポリヌクレオチドは、該ベクターにおいて発現制御配列に作動可能に連結され得、原核細胞および真核細胞で発現可能である。該ポリヌクレオチドの発現は、転写可能なmRNAへのポリヌクレオチドの転写を含む。真核細胞、好ましくは哺乳類細胞での発現を確実にする調節エレメントは、当業者によく知られている。それらは通常、転写開始を確実にする調節配列および転写の終結および転写物の安定化を確実にする任意のポリ−Aシグナルを含有する。さらなる調節エレメントとしては、転写エンハンサーならびに翻訳エンハンサーを含んでもよい。原核宿主細胞で発現を可能にする可能な調節エレメントとしては、例えば、大腸菌のlac、trpまたはtacプロモーターを含み、真核宿主細胞で発現を可能にする調節エレメントの例としては、酵母のAOX1もしくはGAL1プロモーターまたは哺乳類および他の動物細胞のCMV−、SV40−、RSV−プロモーター(ラウス肉腫ウイルス)、CMV−エンハンサー、SV40−エンハンサーまたはグロビンイントロンである。転写開始を担うエレメントの他に、かかる調節エレメントはまた、ポリヌクレオチドの下流に、SV40−ポリ−A部位またはtk−ポリ−A部位等の転写終結シグナルを含有してよい。これに関連して、岡山−バーグ cDNA発現ベクターpcDV1(Pharmacia)、pCDM8、pRc/CMV、pcDNA1、pcDNA3(In−vitrogene)、pSPORT1(GIBCO BRL)等の適切な発現ベクターが、当該技術分野で知られている。
【0034】
好ましくは、該ベクターは、発現ベクターおよび/または遺伝子導入ベクターもしくは遺伝子ターゲッティングベクターである。発現ベクターおよび遺伝子ターゲッティングベクターまたは遺伝子導入ベクターは、当該技術分野において既知であり、当業者により本発明の特定の目的に適合させ得る。したがって、レトロウイルス、ワクシニアウイルス、アデノ随伴ウイルス、ヘルペスウイルスまたはウシパピローマウイルス等のウイルス由来発現ベクターは、本発明のポリヌクレオチドまたはベクターの標的細胞集団への送達に使用され得る。当業者によく知られた方法が、組換えウイルスベクターを構築するために使用され得る;例えば、Sambrook、Molecular Cloning、A Laboratory Manual、Cold Spring Harbor Laboratory(1989)N.Y.およびAusubel、Current Protocols in Molecular Biology、Green Publishing Associates and Wiley Interscience、N.Y.(1989)に記載の技術を参照のこと。他方、本発明のポリヌクレオチドおよびベクターは、標的細胞への送達のために、リポソームに再構築され得る。
【0035】
本発明はさらに、本発明のベクターで形質転換された宿主に関する。該宿主は、原核細胞または真核細胞であり得る;上記参照のこと。宿主細胞に存在する本発明のポリヌクレオチドまたはベクターは、宿主細胞のゲノムに統合されていてもよくまたは染色体外で維持されていてもよい。これに関連して、本発明の組換えDNA分子は、相同組換えを介して変異遺伝子を修復するまたは変異遺伝子を作るために、「遺伝子ターゲッティング」および/または「遺伝子置換」に使用され得ることがまた理解されるべきである;例えば、Mouellic、Proc.Natl.Acad.Sci.USA、87(1990)、4712〜4716;Joyner、Gene Targeting、A Practical Approach,Oxford University Pressを参照のこと。好ましくは、宿主は、哺乳類細胞、真菌細胞、植物細胞、昆虫細胞または細菌細胞である。好ましい真菌細胞は、例えば、サッカロミセス属のもの、特にS.セレビジエ(cerevisiae)種のものである。用語「原核(prokaryotic)」は、本発明のタンパク質の発現のためのポリヌクレオチドで形質転換またはトランスフェクトされ得る全ての細菌を含むことを意図する。原核生物宿主は、例えば、大腸菌、S.チフィミュリウム(typhimurium)、セラチア マルセッセンス(Serratia marcescens)およびバチルス サブティリス(Bucillus subtilis)等のグラム陰性細菌ならびにグラム陽性細菌を含み得る。融合された、作動可能に連結された遺伝子を調製し、細菌細胞または動物細胞でそれらを発現する方法は、当該分野においてよく知られている(Maniatisら、Molecular Cloning:A Laboratory Manual、Cold Spring Harbor Laboratory,Cold Spring Harbor,NY,1989)。本明細書に記載の遺伝子構築および方法は、原核生物宿主で本発明のタンパク質を発現するのに利用できる。一般的に、挿入されたポリヌクレオチドの有効な転写を促進するプロモーター配列を含む発現ベクターが、宿主と関連して使用される。発現ベクターは典型的に、複製の起点、プロモーター、ターミネーター、ならびに形質転換された細胞の表現型選択を与えることができる特異的な遺伝子を含む。形質転換された原核生物宿主は、発酵槽で成長でき、当該分野で既知の技術により培養して最適な細胞成長を得ることができる。本発明のポリペプチドは次いで、成長培地、細胞溶解物または細胞膜フラクションから単離することができる。微生物学的または別の方法で発現させた本発明のポリペプチドの単離および精製は、例えば、モノクローナル抗体またはポリクローナル抗体の使用に関連する方法等の予備のクロマトグラフィー分離および免疫学的分離等のいくつかの慣用の手段によることができる。哺乳類細胞に関しては、HEK 293、CHO、HeLaおよびNIH 3T3が好ましい。昆虫細胞に関しては、スポドプテラ フルギペルダ(Spodoptera frugiperda)細胞が最も好ましく、一方、最も好ましい細菌細胞は大腸菌細胞である。
【0036】
本発明はまた、本発明の宿主を培養する工程、および産生された(ポリ)ペプチドを単離する工程を含む、本発明の核酸分子によってコードされた(ポリ)ペプチドの生産方法に関する。
【0037】
使用されるベクター構築に依存して、本発明の(ポリ)ペプチドは、培養培地に輸送されるか、または宿主中で維持され得る。産生される(ポリ)ペプチドを得るための適切なプロトコルは、両方法の(ポリ)ペプチド産生に対して当該分野でよく知られている。
【0038】
本発明はさらに、本発明の核酸分子によってコードされる(ポリ)ペプチド、または、本発明の方法によって生産される(ポリ)ペプチドに関する。新しいチャンネルは、短いアミノ末端領域、おそらく細胞内の5つのメンブレンスパンニングセグメント、膜に入る疎水性のヘアピン、6番目の膜貫通セグメント、サイクリック−ヌクレオチド結合コンセンサス配列を含有する長いC末端細胞質部分、細胞核局在コンセンサス配列、およびおそらくコイルドコイル(coiled−coil)構造を形成する疎水性領域を有する構造を示すと考えられる。本発明のポリペプチドはまた、ヒトK+ イオンチャンネルの機能的断片でもあり得る。「機能的断片」により、ポリペプチドは前記のいくつかのheagの活性を示すことが意味される。組換えDNA技術を用いて、本発明の(ポリ)ペプチドの断片が産生され得る。これらの断片は、例えば上記で示したように本発明で記載されたもののような種々のアッセイシステムを用いて、所望の機能に対して試験できる。好ましくは、該断片は新規イオンチャンネルのC末端部分を含有する。
【0039】
本発明はまた、本発明の(ポリ)ペプチドに対し、特異的な抗体に関する。本発明の抗体は、特に、ヒトeagチャンネルと、マウスeagおよびラットeag等の先行技術のチャンネルを区別し、好ましくは、イオンチャンネルのC末端部分でエピトープに結合する。用語「抗体」はまた、本発明により使用される時、抗体断片もしくはF(ab)2 、Fab’、FvもしくはscFv断片等の誘導体に関する;例えば、HarlowおよびLane、「Antibodies、A Laboratory Manual」、CSH Press 1988、Cold Spring Harbor,NYを参照のこと。好ましくは、本発明の抗体は、モノクローナル抗体である。
【0040】
本発明はまた、本発明の核酸分子、本発明のベクター、本発明のポリペプチドおよび/または本発明の抗体および薬学的に許容できる担体および/または希釈剤および/または賦形剤を含有してなる医薬組成物に関する。
【0041】
適切な医薬担体および希釈剤ならびに賦形剤の例が当該分野でよく知られており、リン酸緩衝生理食塩水、水、油/水エマルジョン等のエマルジョン、種々の型の湿潤剤、滅菌溶液等を含む。かかる担体を含有する組成物は、よく知られた慣用の方法により調製され得る。これらの医薬組成物は、それの必要な患者に、適切な投与量で投与され得る。適切な組成物の投与は、例えば、経口、静脈内、腹膜内、皮下、筋内、局所または皮内投与等の異なった方法により行なわれ得る。投与量摂生は、担当の医師および臨床的因子によって決定されるであろう。医学分野においてよく知られているように、いずれのある患者に対する投与量も、患者のサイズ、体表面積、年齢、投与対象の特定の化合物、性別、投与時間および投与経路、一般的な健康、ならびに同時に投与される他の薬物を含む多くの因子に依存する。一般的に、医薬組成物の規則的な投与のような摂生は、1日当たり1μg〜10mg単位の範囲であるべきである。摂生が連続注入である場合、それぞれ、体重1キログラム当たり、1分当たり、それはまた1μg〜10mg単位の範囲であるべきである。経過は、周期的な評価によりモニターされ得る。投与量は変化するであろうが、DNAの静脈内投与に対する好ましい投与量は、DNA分子の約106 〜1012コピーである。本発明の組成物は、局部にまたは全身に投与され得る。投与は一般的に、非経口的、例えば静脈内であろう;DNAはまた、例えば、内部もしくは外部ターゲット部位へバイオリスティック(biolistic)送達によりまたは動脈内のある部位にカテーテルにより、直接ターゲット部位に投与され得る。
【0042】
本発明の種々のポリヌクレオチドおよびベクターが、単独で、または標準ベクターおよび/または遺伝子送達系を使用するあらゆる組み合わせで、ならびに任意に薬学的に許容できる担体または賦形剤と一緒に投与されることが、本発明により認識される。投与に続き、前記ポリヌクレオチドまたはベクターは、対象物のゲノムへ安定に統合されてもよい。一方、ある細胞または組織に特異的であり、該細胞または組織で存続するウイルスベクターが使用され得る。適切な医薬担体および賦形剤は、前記で述べたように、当該分野でよく知られている。本発明により調製される医薬組成物は、本発明の前記同定された核酸分子の望まない(過剰)発現に関連する異なる種類の疾患の予防または治療または遅延に使用され得る。好ましい態様において、医薬組成物は、例えば、実施例5に記載のように、調節、好ましくは多量の発現を低減できるアンチセンスオリゴデオキシヌクレオチドを含有する。
【0043】
さらに、本発明のポリヌクレオチドまたはベクターを含有する本発明の医薬組成物を、遺伝子治療で使用することができる。適切な遺伝子送達系は、リポソーム、レセプター媒介送達系、ネイキッド(naked)DNAおよびヘルペスウイルス、レトロウイルス、アデノウイルス、アデノ随伴ウイルス等の特にウイルスベクターを含み得る。例えば、イクスビボまたはインビボ技術による細胞へのワクチン接種のために治療的遺伝子を導入することに基づく遺伝子治療は、遺伝子導入の最も重要な適用の1つである。インビトロまたはインビボ遺伝子治療のための適切なベクター、方法または遺伝子送達系は、文献に記載されており、当業者に知られている;例えば、Giordano、Nature Medicine 2(1996)、534〜539;Schaper、Circ.Res.79 81996)、911〜919;Anderson、Science 256(1992)、808〜813;Isner、Lancet 348(1996)、370〜374;Muhlhauser、Circ.Res.77(1995)、1077〜1086;Onodera、Blood 91(1998)、30〜36;Verzeletti、Hum.Gene Ther.9(1998)、2243〜2251;Verma、Nature 389(1997)、239〜242;Anderson、Nature 392(Supp.1998)、25〜30;Wang、Gene Therapy 4(1997)、343〜400;Wang、Nature Medicine 2(1996)、714〜716;WO94/29469;WO97/00957;US5,580,859;US5,589,466;US4,394,448またはSchaper、Current Opinion in Biotechnology 7(1996),635〜640および本明細書に引用された参照文献を参照のこと。本発明の核酸分子およびベクターは、細胞への直接の導入またはリポソームまたはウイルスベクター(例えば、アデノウイルス、レトロウイルス)を介する導入に対して設計され得る。さらに、バキュロウイルス系が、本発明の核酸分子に対する真核生物発現系として使用できる。遺伝子治療のための体の特異的部位への核酸の送達はまた、Williams(Proc.Natl.Acad.Sci.USA88(1991)、2726〜2729)により記載されたもの等のバイオリスティック送達系を使用して達成され得る。
【0044】
組換えDNAで細胞をトランスフェクトする標準的な方法は、分子生物学の分野の当業者によく知られている。例えば、WO94/29469を参照のこと。遺伝子治療は、本発明の組換えDNA分子またはベクターを患者に直接投与することにより、または本発明のポリヌクレオチドまたはベクターで細胞をイクスビボでトランスフェクトし、トランスフェクトされた細胞を患者に注入することにより、行ない得る。さらに、生殖系の細胞への遺伝子導入に関する研究は、生殖生物学で最も速い成長分野の1つである。イクスビボまたはインビボ技術による細胞への治療的遺伝子の導入に基づく遺伝子治療は、遺伝子導入の最も重要な適用の1つである。インビトロまたはインビボでの遺伝子治療のための適切なベクターおよび方法は、文献に記載されており、当業者に知られている;例えば、WO94/29469、WO97/00957またはSchaper(Current Opinion in Biotechnology 7(1996)、635〜640)および上記に引用された参照文献を参照のこと。本発明の医薬組成物に含有されるポリヌクレオチドおよびベクターは、細胞への直接の導入、またはリポソームもしくは該組換えDNA分子を含むウイルスベクター(例えば、アデノウイルス、レトロウイルス)を介する導入のために設計され得る。好ましくは、該細胞は、生殖系細胞、胚性細胞、幹細胞または卵細胞もしくはそれら由来の細胞である。胚性細胞は、例えば、Nagy、Proc.Natl.Acad.Sci.USA90(1993)8424〜8428等に記載のような胚性幹細胞であり得る。
【0045】
本発明の導入されたポリヌクレオチドおよびベクターは、該細胞への導入後本発明の(ポリ)ペプチドを発現し、好ましくは該細胞のライフタイムの間この状態のままであることが理解されるべきである。例えば、適切な調節配列の制御下に安定にポリヌクレオチドを発現する細胞株は、当業者によく知られた方法により作製されてもよい。ウイルスの複製起点を含む発現ベクターを使用するよりむしろ、宿主細胞は、本発明のポリヌクレオチドまたはベクターおよび同一のベクターもしくは異なるベクターのどちらかにある選択可能なマーカーで形質転換され得る。外来のDNAの導入に続き、作製した細胞を、強化培地で1〜2日間、成長させてよく、次いで選択培地に変える。組換えプラスミドの選択可能なマーカーは、選択に耐性を与え、プラスミドを安定にそれらの染色体中に統合化しており、成長して順番に細胞株にクローニングされ、拡張され得るフォーカスを形成する細胞の選択を可能にする。かかる作製した細胞株の、スクリーニング方法、または、下記のような本発明のポリペプチドのインヒビターの同定方法に特に有用である。
【0046】
限定されないが、それぞれtk細胞、hgprt細胞またはaprt細胞のヘルペスシンプレックスウイルスチミジンキナーゼ(Wigler、Cell 11(1977)、223)、ヒポキサンチン−グアニンホスホリボシルトランスフェラーゼ(Szybalska、Proc.Natl.Acad.Sci.USA 48(1962)、2026)、アデニンホスホリボシルトランスフェラーゼ(Lowy、Cell 22(1980)、817)を含む多数の選択系が使用され得る。代謝拮抗物質耐性がまた、メトトレキサートに耐性を与えるdhfr(Wigler、Proc.Natl.Acad.Sci.USA 77(1980)、3567;O’Hare、Proc.Natl.Acad.Sci.USA 78(1981)、1527)、ミコフェノール酸に耐性を与えるgpt(Mulligan、Proc.Natl.Acad.Sci.USA 78(1981)、2072)、アミノグリコシドG−418に耐性を与えるneo(Colberre−Garapin、J.Mol.Biol.150(1981)、1)、ハイグロマイシンに耐性を与えるhygro(Santerre、Gene 30(1984)、147)、ゼオシン〔Zeocin(登録商標)〕に耐性を与えるShble(Mulsant、Somat.Cell.Mol.Genet.14(1988)、243〜252またはピューロマイシン(pat、ピューロマイシンN−アセチルトランスフェラーゼ)に対する選択の基礎として使用され得る。さらなる選択可能な遺伝子が記載されており、例えば、細胞がトリプトファンの代わりにインドールを利用できるようにするtrpB;細胞がヒスチジンの代わりにヒスチノールを利用できるようにするhisD(Hartman、Proc.Natl.Acad.Sci.USA 85(1988)、8047);およびオルニチンデカルボキシラーゼインヒビター、2−(ジフルオロメチル)−DL−オルニチン、DFMO(McConlogue、1987、In: Current Communications in Molecular Biology、Cold Spring Harbor Laboratory 編)に耐性を与えるODC(オルニチンデカルボキシラーゼ)がある。イクスビボ遺伝子治療の使用対象の細胞は、当業者によく知られている。例えば、かかる細胞は、例えば血液にもしくは組織に存在する癌細胞または好ましくは対応する幹細胞を含む。
【0047】
さらに、本発明は、本発明の核酸分子、本発明のベクター、本発明のポリペプチドおよび/または本発明の抗体を含有してなる診断用組成物に関する。
【0048】
本発明の診断用組成物は、本発明の核酸分子の望まない発現または過剰発現に関連する疾患の開始または進行を検出するのに有用である。本明細書において前記に指摘したように、かかる疾患は、細胞増殖の増加または進行と相互関係があるか、またはそれらにより引き起こされる。したがって、本発明の診断用組成物は、癌の開始または疾患状態を評価するために使用され得る。このように腫瘍活性に対する初期のクリチリウム(criterium)を有しており、適切なカウンター測定を直ちに適用し得る。かかる即時行動は、もちろん、患者の予後を有意に改良するであろう。これらの考慮は、転移腫瘍および反回性腫瘍の診断に等しく適用される。
【0049】
一方、腫瘍の全ての型が本発明の核酸分子の望まない発現および過剰発現によって特徴づけられ得るというわけではない。他方、該(過剰)発現は、腫瘍発達の初期の段階のようなある段階のみで生じるかもしれない。それゆえに、本発明の診断用組成物はまた、すなわち他方で腫瘍の分類もしくは腫瘍の発達状態の分類のための手段として使用され得る。当然、腫瘍に対する本明細書に記載された本発明の組成物の適用またはほとんどの適用もまた、本発明の核酸分子の望まない(過剰)発現それらと相互関係があるまたはそれにより生じる他の疾患に応用する。
【0050】
さらに、本明細書を通して引用されるような疾患はまた、本発明のポリペプチドの機能不全により引き起こされ得るであろう。該疾患は、例えば、本発明のポリペプチドの遺伝子量(dosis)増加または低減、細胞もしくは組織の対応する野生型ポリペプチドと比較した一次アミノ酸配列での修飾または該ポリペプチドの活性の調節の損失等による該ポリペプチドの活性の増加または低減と相互関係があり得、またはそれらにより生じ得るであろう。該疾患はさらに、細胞周期進行または細胞発達の間のポリペプチドの間違った発現によって引き起こされるかもしれない。例えば、イオンまたは二次メッセンジャーまたは調節タンパク質等の細胞内または細胞外化合物への変異した結合部位は、それが該ポリペプチドの活性を調節する該化合物に対する結合特性を変える場合、本発明のポリペプチドの機能不全を生じさせ得るかもしれない。機能不全はまた、不完全な修飾部位、例えば、リン酸化部位またはグリコシル化部位により引き起こされ得るであろう。それはまた、例えば、heag1がheagh2の代わりに発現する場合、または逆の場合、間違ったスプライシング事象、それゆえ切形もしくは伸長ポリペプチドの発現により、引き起こされるかもしれない。
【0051】
したがって、さらなる態様において、前記診断用組成物はまた、本発明のポリペプチドの機能不全を検出するのに使用され得る。
【0052】
さらなる態様において、本発明は、本発明の核酸分子の発現のインヒビターまたは本発明の(ポリ)ペプチドの機能不全のインヒビターもしくは改変剤またはheagをコードする核酸分子またはheag活性を有するポリペプチドを、該疾患の影響を受けた哺乳類または該疾患の影響を受けやすいと推測される哺乳類に導入する工程を含む、本発明のポリペプチドの機能不全により引き起こされる疾患を予防または治療する方法に関する。heagをコードする本発明の核酸分子の細胞または対象物への導入方法、すなわち遺伝子治療は、本発明の核酸分子の発現のインヒビターの同定方法と同様に本明細書に記載されている。さらに本発明のポリペプチドの機能不全のインヒビターまたは改変剤は、当業者に既知の本発明のポリペプチドのインヒビターインヒビターの同定方法により同定され得る(下記を参照のこと)。例えば、本発明のポリペプチドの機能不全を生じるある遺伝子変化は、タンパク質高次構造状態に変化をもたらす。変異タンパク質は、イオン輸送の促進をほとんど不可能にする三次構造を有し得る。変異タンパク質の正常なまたは調節された高次構造を回復することは、これらの分子欠陥を正すために最も明快で特有の意味を持つ。薬理学操作は、このようにタンパク質の野生型構造の復活を助けるかもしれない。このように、本発明のポリヌクレオチドおよびコードされたタンパク質はまた、該機能不全を示す本発明のポリペプチドの誘導体の野性型機能を活性化できる分子を設計および/または同定するために使用され得る。
【0053】
それの必要な患者の治療のための投与量および投与経路は、本発明の医薬組成物に関連して本発明の前記ですでに記載した。本発明の方法を使用して治療され得る疾患は、細胞増殖に関係するいずれの疾患をも含む。このカテゴリーに分類される好ましい疾患は、癌(乳癌、神経芽腫等)等の腫瘍性疾患、乾癬、および変性疾患、とりわけアルツハイマー病、多発性硬化症、筋萎縮性側索硬化症、パーキンソン病等の神経系の疾患である。
【0054】
好ましくは、該核酸分子の発現または過剰発現の該インヒビターは、本発明のイオンチャンネルをコードする核酸分子またはその断片にハイブリダイズする本発明の核酸分子である。例えば、この核酸分子は、アンチセンスオリゴデソキシヌクレオチド(oligodesoxynucleotido)(ODN)であり得る。発明者は、アンチセンスODN治療が、いくつかの腫瘍細胞、例えばEFM細胞、SHSY−5Y細胞およびHeLa細胞のDNA合成を有意に低減することを示し得た(実施例5)。したがって、好ましい態様において、核酸分子はアンチセンスODNを含有する。
【0055】
さらに好ましい態様において、該ポリペプチド機能のインヒビターは、本発明の抗体または薬物である。該薬物は、ヒスタミンレセプターH1インヒビターであり得る。好ましくは、該薬物は、活性heagを阻害し、例えば使用依存的におそらくオープンチャンネルブロッカーとして作用し、好ましくは該薬物はアステミゾール(astemizole)またはテルフェナジン(terfenazine)である。さらに、適切な薬物が、本発明の教示に基づいて当業者により同定または設計され得る。好ましくは、薬物は、heagチャンネルにmM範囲、さらに好ましくはnM範囲またはそれより低い範囲で親和性を有するであろう。好ましくは、該薬物は、他のチャンネル、例えば心臓チャンネルに影響しない。
【0056】
本発明のさらに好ましい態様において、該方法は、導入工程の前に、
(a)前記疾患により冒された哺乳動物から細胞を得る工程ならびに前記導入工程の後に前記疾患により冒された哺乳動物から細胞を得る工程、ここで、前記導入は前記細胞に行われる、
(b)前記細胞を前記哺乳動物または同種の哺乳動物に再導入する工程、
をさらに含む。
【0057】
本発明の本態様は、治療継続時間を大きく低減し、効力を増加し、副作用を低減する(消去さえする)遺伝子治療目的のために特に有用である。さらに、本発明の方法の本態様はまた、これに関連してまたは通常の医学治療と組み合わせて使用し得る。該哺乳類からの除去および該哺乳類への再導入は標準手順によって行なうことができる。
【0058】
好ましくは、前記参照細胞は、生殖細胞、胚性細胞もしくは卵細胞またはこれらの細胞いずれか由来の細胞である。
【0059】
本発明は、さらに、本発明の核酸分子の望まれていない発現または過剰発現によって引き起こされる疾患の治療用薬物の設計方法であって、
(a)特異的かつ効力の高い薬物を同定する工程;
(b)部位特異的突然変異誘発およびキメラタンパク質研究による、前記薬物の結合部位を同定する工程;
(c)(ポリ)ペプチドの結合部位および前記薬物の構造の両方を分子モデリングする工程;ならびに
(d)(ポリ)ペプチドに対するその結合特異性を改良するために薬物を改変する工程、
を含む、治療用薬物の設計方法に関する。
【0060】
本明細書で使用される用語「特異的かつ効力の高い薬物」は、heag機能を高い効力でかつ特異的にブロックする薬物に関する。
【0061】
本発明の方法の様々な段階で使用される全技術は通常のものであるかまたは、さらなる努力なしの通常の技術により当業者によりもたらされ得る。したがって、本発明のイオンチャンネルの特徴を同定する本明細書に基づく生物学的アッセイが、薬物の特異性または効能を評価するために使用され、そこで、イオンチャンネルの1以上の本発明の活性の減少が該特異性または効能をモニターするのに使用され得る。工程(b)および(d)は、例えば、K.L.Choi、C.Mossman、J.Aube & G.Yellen.The International Quaternary Ammonium Receptor Site of Shaker Potassium Channels.Neuron 10、533〜541(1993)、C.−C.Shieh & G.E.Kirsch:Mutational Analysis of Ion Conduction and Drug Binding Sites in the Inner Mouth of Voltage−Gated K+ −Channels.Biophys.J.67、2316〜2325(1994)、またはC.Miller:The Charybdotoxin Family of K+ −Channel−Blocking Peptide.Neuron 15、5〜10(1995)に記載される通常のプロトコルにしたがって行なうことができる。
【0062】
例えば、部位特異的突然変異誘発およびキメラタンパク質研究による該薬物の結合部位の同定は、薬物親和性に影響を及ぼす(ポリ)ペプチド一次配列での修飾により達成され得る;これは、通常、薬物に対する結合ポケットを正確にマッピングすることを可能にする。
【0063】
工程(c)に関しては、次のプロトコルが認識され得る;一度、薬物に対するエフェクター部位がマップされると、薬物の正確な三次元構造が既知である場合には、薬物の異なる部分と相互作用する正確な残基が、突然変異誘発研究〔工程(b)〕から得られる情報と結合部位の構造のコンピューターシミュレーション(カリウムチャンネルが最近当該分野において結晶化されているので、これは現在さらなる努力なしに当業者によってなされ得る)との組み合わせによって同定され得る(そうでないなら、それはコンピューターシミュレーションによって予測され得る)。該薬物それ自身がペプチドである場合、それはまた、どの残基がheag分子の他の残基と相互作用するか決定するために変異され得る。
【0064】
最終的に工程(d)で薬物は修飾され、その結合親和性またはその効能および特異性を改良し得る。例えば、もしheagの特定の残基と薬物分子のある領域の間に静電的相互作用があるなら、その領域の全体的な電荷を修飾し、その特定の相互作用を増加できる;さらに、もしそれらの相互作用が他のチャンネルタンパク質で変換されないheagの領域で生じるなら、この相互作用の改良は、他の結合因子が弱められる一方で、薬物の特異性を改良すると考えられている。
【0065】
結合部位の同定は、コンピュータープログラムによって補佐され得る。したがって、適切なコンピュータープログラムが、相補的な構造モチーフに対するコンピューター補佐探究によって、本発明の推定上のインヒビターおよびポリペプチドの相互作用部位の同定に使用され得る(Fassina、Immunomethods 5(1994)、114〜120)。さらに、タンパク質およびペプチドのコンピューター補佐設計のための適切なコンピューターシステムが、例えば、Berry、Biochem.Soc.Trans.22(1994)、1033〜1036;Wodak、Ann.N.Y.Acad.Sci.501(1987)、1〜13;Pabo、Biochemistry 25(1986)、5987〜5991の先行技術で記載されている。薬物の修飾は、例えば、ぺプチド擬似体により製造され、他のインヒビターはまた、次の化学修飾および得られた化合物を試験することを介するぺプチド擬似体コンビナトリアルライブラリーの合成により同定し得る。ぺプチド擬似体コンビナトリアルライブラリーの生成および使用方法は、例えば、Ostresh,Methods in Enzymology 267(1996)、220〜234およびDorner、Bioorg.Med.Chem.4(1996)、709〜715の先行技術で記載されている。さらに、本発明のポリペプチドのインヒビターの三次元構造および/または結晶構造は、例えば、本発明の(ポリ)ペプチドと組み合わせて(Rose、Biochemistry 35(1996)、12933〜12944;Rutenber、Bioorg.Med.Chem.4(1996)、1545〜1558)、ペプチド擬似体インヒビターの設計に使用され得る。
【0066】
本発明にしたがって使用され得る特定のインヒビターを同定するための具体的な戦略が、添付した実施例で提供される。
【0067】
本発明はまた、
(a)アンチセンスオリゴヌクレオチドおよびリボザイムから選ばれる化合物を、翻訳の阻害または低減に対して試験する工程;または
(b)本発明の(ポリ)ペプチドをコードし、かつ好ましくはその転写因子応答エレメントを有する遺伝子のプロモーター領域に結合する化合物を、転写の阻害に対して試験する工程;または
(c)本発明の(ポリ)ペプチドの増殖活性をブロックすると推測されるペプチドまたは抗体を、前記ブロック活性に対して試験する工程、
を含む、本発明の核酸分子の発現のインヒビターまたは本発明の(ポリ)ペプチドの機能のインヒビターを同定する方法に関する。
【0068】
前記に関するもう1つの(b)に関しては、まずプロモーター領域を特徴づけ、その中に転写因子応答配列があることが有利であり得る。次いで、プロモーターを遺伝子的に操作して、それをリプレッサーに対する反応性を高くし、エンハンサーに対する反応性を低くすることが可能であろう。今度はもう1つの(c)を見ると、それはまず増殖障害の発生に影響を与える本発明のイオンチャンネルの1以上の部分にあることが有利である。1つの試験系で陽性である化合物は、インヒビターとして有用であることが明白である。
【0069】
ペプチド擬似体、ファージディスプレイおよびコンビナトリアルライブラリー技術は当該分野でよく知られており、それは当業者によりさらなる努力なしに前記本明細書に関連する基本的方法によって同定される薬物またはインヒビターの改良に適用できる。
【0070】
さらなる態様において、本発明は、本発明の核酸の発現または本発明の(ポリ)ペプチドにインヒビターを適用する工程を含む、細胞増殖を阻害する方法に関する。本発明の方法は、インビトロ、イクスビボ、または適用が対象物に対する場合にはインビボで行ない得る。
【0071】
本発明はまた、本発明の核酸分子の発現を評価する工程、または本発明の(ポリ)ペプチドの定量的な存在を評価する工程を含む、癌および/または神経変性疾患および/または乾癬を予知する方法に関する。好ましい態様において、該癌は、乳癌または神経芽腫、さらに好ましい態様において、該癌は、胸腺癌、胸癌管型または子宮頸癌である。さらなる態様において、該神経変性疾患は、アルツハイマー病、パーキンソン病、筋萎縮性側索硬化症または多発性硬化症である。
【0072】
本発明の方法は、インビトロ、インビボまたはイクスビボで行ない得る。本発明の方法を行なうための適切なプロトコルは当該分野でよく知られており、インビトロ技術に関しては、mRNAのレベルの評価のためのノーザンブロット、または、例えば本発明の(ポリ)ペプチドを特異的に認識する抗体を用いる顕微鏡技術による組織の解析を含む。1以上のこれらの技術は、同様にまたはさらなる(通常の)技術を組み合わせて前記引用された評価に使用される技術に基づいたPCRと組み合わされ得る。
【0073】
本発明の前記方法のさらなる好ましい態様において、該哺乳類は、ヒト、ラットまたはマウスである。
【0074】
本発明はさらに、遺伝子治療における本発明の核酸分子の使用に関する。本明細書前記で指摘したように、遺伝子治療は細胞増殖を阻害するように設計され、それにより癌または乾癬等のそれにより影響されるいずれの疾患をも特定の方法で治療し得る。本発明は特に、かかる遺伝子治療プロトコルを行なう2つの非依存系を認識する:
(a)ヒトeagに特異的な薬物を得るための、H1アンタゴニスト(好ましくはアステミゾール)の化学工学を共に有するチャンネルの変異誘発;
(b)診断および/または予知方法を設計するための、癌細胞でのeagの発現レベルの定量的および定性的解析。これはまた、特定の腫瘍に対する遺伝子治療の設計を可能にするであろう。
【0075】
例えば、核酸分子は、レトロウイルスベクター(Naldiniら、Science 272(1996)、263〜267;Mulligan、Science 260(1993)、926〜932)または適切なベクターを用いて細胞にインビボで導入され得る。同様に本発明により、患者由来の細胞が単離され、標準組織培養技術を用いてインビトロで修飾され、そして患者に再導入され得る。かかる方法は、本明細書前記に関連する遺伝子治療または遺伝子転写方法を含む。
【0076】
最後に、本発明は、本発明の(ポリ)ペプチドをコードする核酸分子に特異的にハイブリダイズする核酸分子、本発明のベクター、本発明のポリペプチドおよび/または本発明の抗体を含有してなるキットに関する。
【0077】
本発明のキットは特に、前記に関連する多数の診断方法に使用され得る。本発明のキットは、形質転換された宿主細胞およびトランスジェニック植物細胞、植物組織または植物の発生に適切な選択培地に対する選択マーカーおよび成分等の材料をさらに含む。さらにまた、キットは、本発明の組換え遺伝子またはベクターに存在し得るレポーター遺伝子のための緩衝液および基質を含み得る。本発明のキットは、本発明の方法を行なうために有利に使用することができ、特に、本明細書に言及する様々な適用、例えば診断分野または研究手段の際に使用し得る。本発明のキットの部分は、バイアルに個々に、または容器もしくは多容器単位に組み合わせてパッケージされ得る。キットの製造は、好ましくは、当業者に既知の標準手段に従う。
【0078】
いくつかの文書が、本明細書の本文を通して引用されている。本明細書で引用されている各文書は(いずれの製造業者の仕様書、指示等をも含む)、これによって本明細書に参照により取り込まれる;しかしながら、引用されたいずれの文書も実際に本発明に関して先行技術であると認めているというわけではない。
【実施例】
【0079】
実施例により本発明を説明する。
【0080】
実施例1:K+ イオンチャンネルのクローニング
標準手法に従ってMCF−7細胞から得た全RNAよりmRNAを精製した。次いで、スーパースクリプト(Superscript)II逆転写酵素を用い逆転写によりcDNAを調製した;このcDNAを、高度に保存されたeag配列に適合するように設計された縮重オリゴヌクレオチドを使用するPCR増幅のための鋳型として用いた。増幅後、ラットeag由来のSacII/SacII断片を、結果のサザンブロット解析のためのプローブとして用いた。陽性ハイブリダイゼーションを示すそれらのバンドを、次いで、pGEM−Tベクター(Promega社)にクローニングし、配列決定した。それらの全てがHERGに対応する配列を与えた。
【0081】
次いで、ラット、マウスおよびウシのeagを考慮し、HERG cDNA増幅を避けるように作製される特異的オリゴヌクレオチドを設計した。我々は、種々のeagクローン間で高い相同性を有するが、HERG配列に対し最も効果的に相違している配列を探した。
【0082】
オリゴヌクレオチドの配列は以下であった:
5'-CAGAA(T,C)AA(T,C)GTGGC(A,C,T,G,)TGGCT
5'-TCACT(G,A)AAGATCTATA(A,G)TC
【0083】
PCR増幅後、期待されたサイズのバンドをpGEMTにクローニングし、配列決定した。得られた配列はラットeag(ヌクレオチド942〜1108)に高度に相同性を示した。
【0084】
このバンドを標識し、乳腺cDNAライブラリーをスクリーニングするためのプローブとして用いた。2×106 ファージのスクリーニング後、陽性クローンを全く見出せなかった。
【0085】
次いで、我々は、特異的オリゴヌクレオチドを用い、(Clontech社から購入した全RNAより得た)ヒト心臓およびヒト脳由来のcDNAをPCRを使用して解析した。脳由来の2つのPCR産物を配列決定し、該配列はeagの2つの選択的にスプライシングされた変異体(variants)に対応した。ヒト脳由来の全長分子のクローニングの可能性をさらに試験するために、我々は、ヒトcDNAライブラリーのPCR解析を行い、この結果をヒト乳腺ライブラリーにおける同じ実験と比較した(両方ともClontech社由来)。脳ライブラリーのみが陽性の結果を与えた。
【0086】
続いて、増幅された断片を用いてヒト脳ライブラリーのスクリーニングを行い(2回、106 ファージ)、pBSKベクターにクローニングされたいくつかのクローンを見出し、配列決定した。それらの全ては分子の中央部分に相当したが、5’および3’末端が欠失していた。これらの陽性クローンの内で最も長鎖のものを用いてプローブを調製し、再度ライブラリーのスクリーニングを行った(再び2回、2×106 クローン)。
【0087】
この場合に得られた配列は、コード配列の一部(約400bpの5’が開始コドンまで欠失していた)と長鎖の3’非翻訳配列に相当した。分子の5’末端に近い断片は全ての場合にEcoRI部位で始まったので、かかる部位はheag配列に実際に存在し、配列決定のためのベクターへの断片のサブクローニングにおいて消失したということが推測された。
【0088】
全長配列を得るため、我々は、5’末端に近い断片を保持するそれらのファージをプールし、前記EcoRI部位に対する配列3’とラムダgt10由来の配列をPCRのためのプライマーとして用い、PCR増幅によりそれらを解析した。プールの連続した分画の後、コード配列の5’末端を保持する2つのファージを得、それらの1つは、5’非翻訳領域の一部を含んでいた。
【0089】
我々は完全配列を知得した後、それらの1つは3’UTRとほとんどのコード配列を含んでおり、他方は5’末端を含んでいる、2つのファージから始めて、全クローンを組み立てた。最初の断片をSphI/HindIII消化によりファージから抽出し、pBKS−にサブクローニングしてpBKSheag1を作製した。このワズ(was)に、1.2kbpのSphI−SphI断片もクローンから除去し、その後、それを再度導入することが必要であった。5’末端を含む断片をHindIII/MunI消化により抽出した。この断片をpBKSheag1のHindIII/MunI消化物とライゲートさせた。この手法を用いてのみ、我々は単一プラスミドにおいて全長クローンを得ることができた。次いで、我々はSphI−SphI断片を再度導入する必要があった。というのは、我々はSphI部位の1つを欠失させたからであった。続いて、EagI/NotI断片をpCDNA3ベクターのNotI部位にサブクローニングし、汚染したファージ配列を除去し、チャンネルの機能的発現に好適なベクターを得た。最終的に得られた配列を配列番号:1および配列番号:2として配列表に記載する。
【0090】
実施例2:ヒトeagの作用を特異的にブロック(bloc)するインヒビターの同定
eagファミリーの他の1つのメンバーであるHERG11-16 は、長鎖QT症候群(LQT)のファミリー型に関連付けられている。これにより、それらのLQT型不整脈を誘導する能力に基づいてHERGのいくつかのブロッカーの同定が可能となっている。このように、アステミゾール(astemizole)およびテルフェナジン(terfenadine)等の特定のヒスタミンH1レセプターブロッカー、ならびにIII型不整脈薬物〔ドフェチリド(dofetilide)、E−4031)〕は、HERG15,17 の強力で特異的なブロッカーである。しかしながら、eagチャンネルについての特異的なブロッカーは未だ記載されていない。HERGとeagチャンネル間の配列類似性のため、reagに対する両群の薬物を本発明に従って試験した。H1ブロッカーもreagに影響し、一方、チャンネルはIII型抗不整脈剤(ドフェチリド)に対しむしろ無感受性である。このことは、選択的にeag型チャンネルをブロックし、(MCF7細胞にも存在する)HERGチャンネルの可能な影響を排除するための有効な道具を提供する。単一の推定ヒトeagチャンネルに対する、これらの薬物の1つ(アステミゾール 5μM)の影響をFig.6に示す。
【0091】
いくつかのreagおよび他のカリウムチャンネルブロッカーがMCF7細胞の成長を阻害することができるか否かをさらに試験した。「陽性」対象として、ATP感受性カリウムチャンネルのブロッカーであるグリベンクルアミド(glibenclamide)も含めた。というのは、この細胞株の増殖を阻害することが記載されているためである2 。DNA合成の速度を測定するために、細胞を、約105 細胞/mlの密度で、かつ成長因子の非存在下で、96穴マイクロタイタープレートにて培養した。24時間の飢餓の後、BrdUの存在下に10%FCSの添加により細胞を刺激した。新たに合成されたDNAに取り込まれたBrdUの量を市販の抗体(Boehringer Mannheim)を用いて測定した。試験された薬物は、刺激と同時に、または刺激の12時間前に添加した。異なるヒト細胞株であるHEK293において、10μM アステミゾールまたは100μM グリベンクルアミドの添加は有意にはDNA合成を低下させなかったが、テルフェナジン(10μM)は強力な阻害を示した。この理由について、アステミゾール(およびその密接に関連するアナログLY91241)の影響のみが考慮され、テルフェナジンにより生成されるもの(MCF7細胞は、対照細胞よりもテルフェナジンによる成長阻害に対し有意により感受性であるが)は排除された。MCF7細胞において、5μM アステミゾールは40%だけDNA合成を低下させ、一方、同濃度のHERG特異的ブロッカーであるドフェチリドは有意な影響を示さなかった。10倍高い濃度(50μM)の他のカリウムチャンネルブロッカー〔キニジン(quinidine)またはグリベンクルアミド(glibenclamide)〕が類似の影響を誘導するのに必要とされる。MCF7細胞におけるDNA合成に対するアステミゾール効果についてのドーズレスポンスカーブをFigure 8に示す。最大効果の半分は10μM アステミゾールについて得られた。
【0092】
MCF7細胞における増殖阻害の基礎となる機構を明らかにする試みにおいて、超生体核染料ヘキスト(Hoechst)33342を用い、5μM アステミゾールで処理された細胞の核形態を調べた。24時間の処理後、ほとんどの細胞が、アポトーシス細胞死の典型的な特徴である、核濃縮および核断片化を示した(Fig.9)。
【0093】
結論として、reagチャンネルのヒト対応物はヒト癌細胞に存在しており、それらは、いくつかの異なる細胞型において悪性変態を誘導する能力を有する。
【0094】
実施例3:異なるヒト組織におけるheagの発現
異なるヒト組織由来の全RNA500ng(または脊髄のポリA+ RNA5ng)を逆転写し、配列 5'-CGCATGAACTACCTGAAGACG(フォワード)および5'-TCTGTGGATGGGGCGATGTTC(リバース)の1組のオリゴヌクレオチドを用いて増幅した。増幅したDNAを特異的なヒトeagプローブ(チャンネルのコア由来の1.5Kb EcoRI断片)を用いてサザンブロットにより解析した。試験したRNAの内、脳全RNAのみが陽性シグナルを与えた。脊髄、副腎、骨格筋、心臓、気管、肝臓、腎臓および乳腺由来のRNAは陰性であった。RNAの正確性をトランスフェリン増幅を用いて調べた。同じアプローチを用いて、いくつかの腫瘍性ヒト細胞株におけるheagの発現を調べた:MCF−7(胸腺癌)BT−474〔胸管癌(breast ductal carcinoma)、充実性腫瘍由来〕、EFM−19〔胸癌、管型胸膜液(pleural fluid)由来〕、COLO−824(胸癌、胸膜液由来)、SHSY5Y(神経芽腫)。正常組織と比較して、試験した全ての癌細胞株はheag発現について陽性であることが見出された。さらに、異なるヒト組織および293細胞由来のRNAのRT−PCR産物のサザンブロットは、脳由来RNAにおいてのみ、heagAおよびBに相当する2つのバンドが増幅され得、同定され得るということを示す。トランスフェリンレセプター(TFR)シグナルを下部に示す(Fig.15A)。さらに、異なるヒト細胞株および乳房上皮細胞の一次培養物(Epith.cells)由来の全RNAのRT−PCR産物のサザンブロット解析。TRFシグナルを下部に示す。異なる細胞株由来のRNA(34)およびヒト組織由来の市販のRNA(Clontech社)をシングル−チューブRT−PCR(35)に供した。脊髄を除き、全RNAを用いた。その際、ポリ(A)+ RNAを用いた(プライマー配列は以下:フォワード:5'-CGCATGAACTACTGAAGACG、およびリバース:5'-TCTGTGGATGGGGCGATGTTC。5'-TCAGCCCAGCAGAAGCATTATおよびリバース:5'-CTGGCAGCGTGTGAGAGC をRNAおよびPCR性能を制御するために用いた。)。TFRの特異的プライマーをRNAおよびPCR性能を制御するために用いた。これらのODNは、エクソン11に始まり、エクソン19まで広がっている(37)公表されたTFR配列(36)に従って設計した。これは、2つのheagスプライス断片および逆転写酵素の非存在下での対照の増幅と共に、ゲノムDNA汚染による偽陽性を排除する。50μl(heag)または15μl(TFR)のPCR反応物を2%アガロースゲルで解析した。DNAをメンブレンに転写し、高度ストリンジェンシーにて、980bp heag断片および脳RNAから増幅されたTFR断片からなる〔32P〕−dCTP標識したランダムにプライミングするプローブと連続的にハイブリダイズさせた。
【0095】
実施例4:インビボでのheagの発現
heagの発現はインビボで腫瘍細胞にとって有利であるか否かを決定するため、本発明者らは、雌性シッド(scid)(極度の複合免疫不全、33)マウスの脇腹へのチャンネルを発現するCHO細胞(CHOhEAG細胞)の皮下移植を行った。CHOKv細胞を対照として用いた。従って、100μlのPBS中に懸濁した2×106 CHOhEAG細胞またはCHO−Kv1.4細胞を、Bomholtgard,Ry,Denmarkから得た6〜8週齢の雌性Fox Chase scidマウス(C.B−17/Icr sicd/scid)の脇腹上で皮下に移植した。腫瘍の存在を、全てのマウスの触診により2日ごとに調べた。2または3週間後、動物を頸の脱臼により屠殺し、腫瘍を切開し、続くパラフィン包埋および染色のためにパラホルムアルデヒドにて固定した。CHOhEAG細胞の同定は、pTracerベクター(Invitrogen)にコードされるグリーン蛍光タンパク質由来の蛍光を生起する腫瘍のUVイルミネーションにより確立した。移植の1週間後、CHOhEAGを注入した全てのマウスが触診により検出可能な腫瘍を有したが、対照においては1mmを超える容量のものは観察されなかった。移植後2週間の間には、heag発現腫瘍は直径で5mmを超過するに至り、ほとんどの場合、皮膚を通して目に見える程に現れた(Fig.17A);マウスを2週間後(N=6)または3週間後(N=7)に屠殺した。使用した11匹の対照動物の内1匹のみが目に見える腫瘍を有さなかった;全13匹のCHOhEAGを注入した動物は腫瘍を示した。heag発現腫瘍の平均容量(Fig.17B,C)は、対照のそれより有意に大きく、特に移植後2週目が大きかった(Fig.17B)。肉眼観察から、腫瘍はもろく、出血性であるようであった;CHOhEAG腫瘍は対照よりも黒ずんでおり、2週目に全てのCHOhEAGを注入したマウスにおいて皮膚に癒着した(Fig.17D,E)。7匹のマウスの内6匹が3週目に同様な特徴を示した。対照的に、腫瘍は、2週間後、全ての対照マウスにおいて皮膚から容易に切開でき、3週目に6匹のマウスの内5匹においてであった。腫瘍の下の組織は全ての場合に影響を受けていないようであった。組織学により確認し(Fig.17H,I,矢頭)、黒ずんだ色は多量の腫瘍内の壊死によるためであった(Fig.17F,G,矢印)。このことは、CHOhEAG腫瘍のより早い成長を示す。EAG発現腫瘍における生命維持範囲の厚さは、有意に対照におけるより小さかった(Fig.17J)。腫瘍の急速な成長により、CHOhEAG群における多量の腫瘍内壊死を説明することができる。このことはまた、移植2週間後における腫瘍の容積に見出される差異の増大を説明する。というのは、CHOhEAG腫瘍は、大量の壊死のために成長を止めるであろうからである。これらのデータは、heag腫瘍の発現はより早く成長し、CHOKv腫瘍よりもより攻撃的であることを強力に示唆する。
【0096】
実施例5:heagの阻害
ある腫瘍細胞におけるheagの発現は、それらの異常な成長の結果ではないが、このK+ チャンネルが、それらの増殖に必要であるということが想定される。それゆえ、アンチセンスオリゴデオキシヌクレオチド(ODN)によるheag発現の阻害は、これらの腫瘍細胞における増殖速度を減少させるはずである。そこで、heagの推定開始コドンに広がる19merのアンチセンスホスホロチオエート ODN(5'-CAGCCATGGTCATCCTCCC)を用いて増殖の阻害を試験した。センスODNおよびスクランブル(scrambled)配列(gtcggtaccagtaggaggg )を対照として用いた。Figure 16Aに示すデータは、EFM細胞におけるheag mRNA含量を減少させるアンチセンスODN処理の有効性を明確にする。アンチセンスODNでの処理によるSHSY−5Y細胞におけるheag媒介K+ 電流の減少をFig.16BおよびCに示す。
【0097】
アンチセンスODNでのheag発現腫瘍細胞株の処理は有意に増幅PCR産物の生成量を減少させた。EFM−19細胞を一夜、10μg/ml DAC30(レーン「C」)または10μg/ml DAC30(Eurogentec)プラス1μM アンチセンスODN(レーン「AS」)で処理し、全RNAを抽出し、heagまたはトランスフェリンレセプターを増幅するように設計されたODNを用いて、実施例3に記載の同条件下でアッセイした。Fig.16Aの矢印は増幅断片の期待されるサイズを示す。さらに、SHSY−5Y神経芽腫細胞におけるheag電流を詳細に解析するために、本発明者らは、細胞外Mg2+の存在下にeag(30)の活性化の電圧依存性を利用した。電流は−120mVから+60mVへ脱分極後に測定した(Fig.16B、灰色の線)。差し引きトレースの最初の部分(Fig.16B、黒線)はeag電流に相当する。該電流は、保持電位が非常に弱い場合(−120mV)、未だ活性化されていないが、保持電位が−60mVである場合、明確になる。19〜21msの平均電流を選択し、電流密度を決定した。アンチセンスODNにより処理したSHSY−5Y細胞における電流密度は、対照細胞と比較して有意に減少した〔電気生理学的測定は、(mM単位)140 NaCl、2.5 KCl、2 CaCl2、2 MgCl2、10 Hepes/NaOH pH7.2、10 グルコースを含む細胞外液を用いてパッチクランプ法のホールセルコンフィギュレーションにて標準プロトコルを使用して行った(Hamill,O.P.,Marty,A.,Neher,E.,Sakmann,B.,Sigworth,F.J.Pfluegers Arch- Eur.J.Physiol 391,85(1981))。ピペット液は、(mM単位)140 KCl、10 BAPTA、10 Hepes/KOH pH7.2であった。〕。細胞を一夜、1μMの蛍光標識ODNを含むアンチセンスODNで処理した。電流は、それらの核において蛍光を示す細胞において1〜3日後に測定した。Fig.16Cのバーは9個の細胞(対照)または25個の細胞(アンチセンス)についての平均±S.E.M.を示す。外部電流のみを解析において評価した。さらに、heagに対し向けられたアンチセンスODNによるヒト癌細胞(EFM−19、HeKaおよびSHSY−5Y)におけるDNA合成の阻害を調査した。DNA合成をODNの非存在下におけるBrdU取り込みと比較して表わす。蛍光標識アンチセンスODNを用いた細胞への取り込み条件を至適化した。細胞を105細胞/mlの密度にて96穴プレートに播いた。播いた1日後、細胞を培養培地で洗浄し、ODNを添加した(終濃度10μM)。ODNを予め無血清培地にて20μg/mlのトランスフェクションラゲネント(ragenent)DAC−30(Eurogentec)と共に混合しておき、20〜30分間、室温にてインキュベートした。次いで、混合物を培養培地に1:1希釈で添加し、一夜、細胞との接触を維持した。このインキュベーション後、細胞を洗浄し、2時間、BrdU(100μM)で標識した。取り込みはBoehringer Mannheim 社のキットを用いて検出し、非特異的バックグラウンド取り込みを差し引いた後、405nm(参照490nm)にてOD単位で測定した(Fig.16D)。バーは、典型的な実験における条件当たり8つのウェルについての平均±S.D.を示す。
【0098】
【表1】

【0099】
参考文献:
1. Moody,W.J.(1995)。イオンチャンネル特性の統合調節によって創られる初期発達の臨界期。Perspect Dev Neurobiol 2、309〜315。
2. Woodfork,K.A.、Wonderlin,W.F.、Peterson,V.A.、およびStrobl,J.S.(1995)。ATP感度の高いカリウムチャンネルの阻害は、組織培養でヒト乳癌細胞の可逆的細胞停止を生じる。J Cell Physiol 162、163〜71。
3. Lepple Wienhues,A.、Berweck,S.、Bohmig,M.、Leo,C.P.、Meyling,B.、Garbe,C.、およびWiederholt,M.(1996)。ヒトメラノーマ細胞増殖での、K+ チャンネルおよび細胞内カルシウムシグナル。J Membr Biol 151、149〜57。
4. Wonderlin,W.F.、Woodfork,K.A.、およびStrobl,J.S.(1995)。細胞周期を通じたMCF−7ヒト乳腫瘍細胞の進行の間の膜電位での変化。J Cell Physiol 165、177〜85。
5. Brueggemann,A.、Stuehmer,W.、およびPardo,L.A.(1997)。アフリカツメガエル卵母細胞で発現されるクローニングさせた遅延レクティファイヤーカリウムチャンネルの有糸分裂促進因子が媒介する抑制。Proc Nat Acad Sci USA 94、537〜542。
6. Mosmann,T.(1983)。細胞増殖および生存のための速い比色アッセイ:増殖および細胞毒アッセイへの適用。J.Immunol.Meth.65、55〜63。
7. Muir,D.、Varon,S.、およびManthorpe,M.(1990)。固定化したミクロ培養を使用するブロモデオキシウリジン取り込みに対する酵素結合免疫吸着剤アッセイ。Anal.Biochem.185、377〜82。
8. Magaud,J.P.、Sargent,I.、およびMason,D.Y.(1998)。ブロモデオキシウリジン取り込みの免疫酵素的測定によるヒト白血球増殖応答の検出。J.Immunol.Meth.106、95〜100。
9. Huong,P.L.、Kolk,A.H.、Eggelte,T.A.、Verstijnen,C.P.、Gilis,H.、およびHendriks,J.T.(1991)。5−ブロモ−デオキシウリジン取り込みを使用した抗原特異的リンパ球増殖の測定。放射性チミジン取り込みに代わる容易でかつ低コスト。J.Immunol.Meth.140、243〜8。
10. Ellwart,J.、およびDormer,P.(1985)。モノクローナルBrdUrd抗体でBrdUrd/ヘキスト消光効果により測定されたDNAへの5−ブロモ−2’−デオキシウリジン(BrdUrd)取り込みでの5−フルオロ−2’−デオキシウリジン(FdUrd)の効果。Cytometry 6、513〜20。
11. Benson,D.W.、MacRae,C.A.、Vesely,M.R.、Walsh,E.P.、Seidman,J.G.、Seidman,C.E.、およびSatler,C.A.(1996)。HERGの穴領域でのミスセンス変異は、家族性の長鎖QT症候群を引き起こす。Circulation 93、1791〜5。
12. Curran,M.E.、Splawski,I.、Timothy,K.W.、Vincent,G.M.、Green,E.D.、およびKeating,M.T.(1995)。心臓性不整脈に対する分子基礎:HERG変異は、長鎖QT症候群を引き起こす。Cell 80、795〜803。
13. Sanguinetti,M.C.、Jiang,C.、Curran,M.E.、およびKeating,M.T.(1995)。遺伝された心臓性不整脈と後天性の心臓性不整脈との機械的結合:HERGはIKrカリウムチャンネルをコードする。Cell 81、299〜307。
14. Spector,P.S.、Curran,M.E.、Keating,M.T.、およびSanguinetti,M.C.(1996)。クラスIII 抗不整脈薬物はHERG、ヒト心臓遅延レクティファイヤーK+ チャンネルをブロックする。メタンスルホンアニリドによるオープンチャンネルブロック。Circ Res 78、499〜503。
15. Trudeau,M.C.、Warmke,J.W.、Ganetzky,B.、およびRobertson,G.A.(1995)。HERG、電圧ゲートカリウムチャンネルファミリーでのヒト内部レクティファイヤー。Science 269、92〜5。
16. Suessbrich,H.、Waldegger,S.、Lang,F.、およびBusch,A.E.(1996)。ヒスタミンレセプターアンタゴニストであるテルフェナジンおよびアステミゾールによるツメガエル卵母細胞で発現されたHERGチャンネルの遮断。FEBS Lett 385、77〜80。
17. 例えば、DeCoursey,T.E.、Chandy,K.G.、Gupta,S.、Cahalan,M.D. Nature 307、465(1984);Mauro,T.、Dixon,D.B.、Komuves,L.、Hanley,K.、Pappone,P.A. J.Invest.Dermat.108、864(1997)を参照のこと。
18. Rouzaire−Dubois,B.およびDubois,J.M. J.Physiol.510、93(1998)。
19. Arcangeli,A.、L.Bianchiら、J.Physiol.489、455〜471(1995)。Bianchi,L.ら、Cancer Res.58、815〜822(1998)。
20. Brueggemann,A.,Stuehmer,W.、Pardo,L.A. Proc.Natl.Acad.Sci.USA.94,537(1997)。Pardo,L.A.、Brueggemann,A.、Camacho,J.、Stuehmer,W. J.Cell.Biol.143、767(1998)。
21. Ludwig,J.ら、EMBO J.13、4451(1994)。
22. Mosmann,T. J.Immunol.Meth.65、55(1983)。細胞をMTT(チアゾリルブルー)50μg/mlで4時間インキュベートし、ホルマザン結晶を10%SDS、10mM HClで12〜14時間で可溶化した。ELISAリーダーを使用して、650nmの参照波長で、570nmでの光学密度を測定した。
23. Stein,G.S.ら、細胞増殖およびアポトーシス。実用的な研究方法、G.P.Studzinski編(IRL Press,Oxford,1995)pp.193〜203。
24. Jainchill,J.L.、Aaronson,S.A.、Todaro,G.J. J.Viol.4、549(1969)。
25. Hermouet,S.、Merendino,J.J.,Jr.、Gutkind,S.、Spiegel,A.M. Proc.Natl.Acad.Sci.USA 88、10455(1991)。
26. 本論文で報告された配列に対するGenBank受入れ番号は、AF078741(heag A)およびAF078742(heag B)である。
27. Occhiodoro,T.ら、FEBS Lett.434,177(1998)。
28. Warmke,J.W.およびGanetzki,B. Proc.Natl.Acad.Sci,USA.91,3438(1994)。Frings,S.ら、J.Gen.Physiol.111、583(1998)。
29. Heng,H.H.Q.、Squire,J.、Tsui,L.−C. Proc.Natl.Acad.Sci,USA、89、9509(1992);Heng,H.H.Q.、およびTsui,L.−C. Chromosoma、102、325(1993)。我々は、FISH解析に対して、Dr.Henry Heng(DNA Biotech,Downsview,Ontario,Canadaを参照のこと)に謝意を表明する。
30. Terlau,H.ら、Pfluegers Arch.−Eur.J.Physiol.、432、301(1996)。Terlau,H.、Heinemann,S.H.、Stuehmer,W.、Pongs,O.、Ludwig,J. J.Physiol.、502、537〜543(1997)。Meyer,R.およびHeinemann,S.H. J.Physiol.508、49(1998)。
31. Bijlenga,P.ら、J.Physiol.512、317(1998)。
32. Bosma,G.C.,ら、Nature 301、527〜530(1987)。
33. Chen,C.およびOkayama,H. Mol.Cell Biol.7、2745(1987)。
34. Chomczynski,P.およびSacchi,N. Anal.Biochem.162、156(1987)。
35. Soto,F.ら、Proc.Natl.Acad.Sci.USA.93、3684(1996)。
36. McClelland,A.、Kuehn,L.C.、Ruddle,F.H. Cell 39、267(1984)。Schneider,C.、Owen,M.J.、Banville,D.、Williams,J.G. Nature 311、675(1984)。
37. Evans,P.およびKemp,J. Gene 199、123(1997)。
【0100】
本発明の態様として、以下のものが挙げられる。
[1](a)配列番号:3または4のアミノ酸配列を有するポリペプチドをコードする核酸分子を含有してなる核酸分子;
(b)配列番号:13または14のDNA配列を有する核酸分子を含有してなる核酸分子;
(c)(a)または(b)の核酸分子の相補的な鎖にハイブリダイズする核酸分子;または
(d)(c)の核酸分子の配列に縮重した核酸分子、
である、ヒトK+ イオンeagチャンネルの機能を有する(ポリ)ペプチドをコードする核酸分子を含有してなる核酸分子。
[2]配列5’−GGGAGGATGACCATGGCTを含有してなる[1]記載の核酸分子に特異的にハイブリダイズする核酸分子。
[3]DNAである[1]または[2]記載の核酸分子。
[4]RNAである[1]または[2]記載の核酸分子。
[5]融合タンパク質をコードする[1]〜[4]いずれか1つに記載の核酸分子。
[6][1]〜[5]いずれか1つに記載の核酸分子を含有してなるベクター。
[7]発現ベクターおよび/または遺伝子ターゲッティングまたは遺伝子導入ベクターである[6]記載のベクター。
[8][6]または[7]記載のベクターで形質転換された宿主。
[9]哺乳類細胞、真菌細胞、植物細胞、昆虫細胞または細菌細胞である[8]記載の宿主。
[10][8]または[9]記載の宿主を培養する工程、および産生された(ポリ)ペプチドを単離する工程を含む、[1]または[3]〜[5]いずれか1つに記載の核酸分子によってコードされた(ポリ)ペプチドの生産方法。
[11][1]または[3]〜[5]いずれか1つに記載の核酸分子によってコードされた、または[10]記載の方法によって生産された(ポリ)ペプチド。
[12][11]記載の(ポリ)ペプチドに対し特異的な抗体。
[13]モノクローナル抗体である[12]記載の抗体。
[14][2]記載の核酸分子、[6]記載のベクター、[11]記載のポリペプチドおよび/または[12]または[13]記載の抗体および薬学的に許容できる担体および/または希釈剤および/または賦形剤を含有してなる医薬組成物。
[15][1]〜[5]いずれか1つに記載の核酸分子、[6]記載のベクター、[11]記載のポリペプチドおよび/または[12]または[13]記載の抗体を含有してなる診断用組成物。
[16][1]または[3]〜[5]いずれか1つに記載の核酸分子の望まれていない発現または過剰発現によって引き起こされる疾患の予防または治療方法であって、[1]または[3]〜[5]いずれか1つに記載の核酸分子の発現のインヒビター、または[11]記載の(ポリ)ペプチドの機能のインヒビターを、前記疾患により影響を受けた哺乳動物または前記疾患の影響を受けやすいと推測される哺乳動物に導入する工程を含む、前記疾患の予防または治療方法。
[17][11]記載の(ポリ)ペプチドの機能不全により引き起こされる疾患の予防または治療方法であって、[1]または[3]〜[5]いずれか1つに記載の核酸分子の発現のインヒビター、[11]記載の(ポリ)ペプチドの機能不全のインヒビターもしくは改変剤、heagをコードする[1]または[3]〜[5]いずれか1つに記載の核酸分子、またはheag活性を有する[11]記載のポリペプチドを、前記疾患の影響を受けた哺乳動物または前記疾患の影響を受けやすいと推測される哺乳動物に導入する工程を含む、前記疾患の予防または治療方法。
[18]前記核酸分子の発現または過剰発現の前記インヒビターが、[2]記載の核酸分子である、[16]記載の方法。
[19]ポリペプチド機能の前記インヒビターが、[12]または[13]記載の抗体、または薬物、好ましくはアステミゾールまたはテルフェナジンである、[16]記載の方法。
[20]導入工程の前に、
(a)前記疾患により冒された哺乳動物から細胞を得る工程ならびに前記導入工程の後に前記疾患により冒された哺乳動物から細胞を得る工程、ここで、前記導入は前記細胞に行われる、
(b)前記細胞を前記哺乳動物または同種の哺乳動物に再導入する工程、
をさらに含む、[16]〜[19]いずれか1つに記載の方法。
[21]前記細胞が、生殖細胞、胚性細胞もしくは卵細胞またはそれらに由来する細胞である、[16]〜[20]いずれか1つに記載の方法。
[22][1]および[3]〜[5]いずれか1つに記載の核酸分子の望まれていない発現または過剰発現によって引き起こされる疾患の治療用薬物の設計方法であって、
(a)特異的かつ効力の高い薬物を同定する工程;
(b)部位特異的突然変異誘発およびキメラタンパク質研究による、前記薬物の結合部位を同定する工程;
(c)(ポリ)ペプチドの結合部位および前記薬物の構造の両方を分子モデリングする工程;ならびに
(c)(ポリ)ペプチドに対するその結合特異性を改良するために薬物を改変する工程、
を含む、治療用薬物の設計方法。
[23](a)アンチセンスオリゴヌクレオチドおよび/またはリボザイムから選ばれた化合物を、翻訳の阻害または低減に対して試験する工程;または
(b)[11]記載の(ポリ)ペプチドをコードし、かつ好ましくはその転写因子応答エレメントを有する遺伝子のプロモーター領域に結合する化合物を、転写の阻害に対して試験する工程;または
(c)[11]記載の(ポリ)ペプチドの増殖活性をブロックすると推測されるペプチドまたは抗体を、前記ブロック活性に対して試験する工程、
を含む、[1]または[3]〜[5]いずれか1つに記載の核酸分子の発現のインヒビターまたは[11]記載の(ポリ)ペプチドの機能のインヒビターの同定方法。
[24]前記薬物またはインヒビターが、ペプチド疑似体またはファージディスプレイもしくはコンビナトリアルライブラリー技術を適用することによってさらに改良された、[22]または[23]記載の方法。
[25][1]または[3]〜[5]いずれか1つに記載の核酸分子または[11]記載の(ポリ)ペプチドの発現に対し、インヒビターを適用する工程を含む、細胞増殖の阻害方法。
[26][1]および[3]〜[5]いずれか1つに記載の核酸分子の発現を評価する工程、または哺乳動物の細胞において[11]記載のポリペプチドの定量的な存在を評価する工程を含む、癌および/または神経変性疾患(neurodegenerative diseases)および/または乾癬の予知方法。
[27]前記癌が、乳癌または神経芽腫または子宮頸癌(cervix carcinoma)である、[26]記載の方法。
[28]前記乳癌が、胸腺癌(breast adenocarcinoma)、胸癌管型(breast carcinoma ductal type)である、[27]記載の方法。
[29]前記神経変性疾患が、アルツハイマー病、パーキンソン病、筋萎縮性側索硬化症または多発性硬化症である、[26]記載の方法。
[30]前記哺乳動物が、ヒト、ラットまたはマウスである、[16]〜[21]および[26]〜[29]いずれか1つに記載の方法。
[31]遺伝子治療における[1]〜[5]いずれか1つに記載の核酸分子の使用。
[32][2]記載の核酸分子、[6]記載のベクター、[11]記載のポリペプチドおよび/または[12]または[13]記載の抗体を含有してなるキット。
【技術分野】
【0001】
本発明は、新規なヒトK+ イオンチャンネル、それをコードする核酸分子および該核酸分子を含有してなるベクターに関する。本発明は、さらに、新規なK+ イオンチャンネルに特異的に生じさせた抗体、ならびに上記化合物の少なくとも1つを含む医薬組成物および診断用キットに関する。さらに、本発明は、本発明のポリペプチドの機能不全または本発明の核酸分子の(過剰)発現によって引き起こされる疾患を治療する方法に関し、該方法は、前記(過剰)発現もしくはイオンチャンネル機能のインヒビター、または該機能不全を排除するインヒビターを、それの必要な患者に投与する工程を含む。前記疾患を治療または予防するための薬物を案出する方法、細胞増殖を阻害する方法および癌を予知する方法は、本発明により含まれるさらなる態様である。本発明はまた、例えば癌に関連するまたは神経変性疾患での、望まれていない細胞増殖を阻害するための本発明の核酸分子に基づいた特異的なアンチセンス治療または遺伝子治療も認識する。
【背景技術】
【0002】
カリウムチャンネルは細胞の静止電位の調節に関連する因子であり、これは興奮性および非興奮性組織でのそれらの主要な役割とみなされている。一方、それらの偏在性の存在およびそれらの特性における特徴的な変異性に対する説明は、不明瞭なままである。カリウムチャンネルは、それらがさらにいくつかの「ハウスキーピング」の役割、例えば細胞増殖1 での「ハウスキーピング」の役割を有するため、全ての細胞型で存在するということは、妥当な仮説である。細胞分裂周期の調節でのそれらの密接な関係がたびたび試験され、いくつかの実験的な証拠が存在している2,3 。しかしながら、とりわけ、細胞周期の間の膜電位の脱分極および過分極の両方が細胞型に依存すると報告されているため1,4 、細胞周期におけるカリウムチャンネルの機能を説明する一般的なモデルはない。K+ チャンネルの役割を説明するために、2つの機構が提案されている:それらは、細胞内でのCa2+濃度に影響を及ぼすか、または細胞容積を制御するかのどちらかである(17、18)。両機構は、間接的に細胞増殖に影響を及ぼすであろう。eagファミリーのメンバーはまた、癌細胞で優先的に発現されると提案されている(19)。数種のカリウムチャンネルブロッカーが癌細胞増殖をブロック(block)する能力に対して試験されており、それらのいくつかは、腫瘍化学治療に対するコアジュバントとして、特に複数の薬物に耐性のある腫瘍において使用されている。それにもかかわらず、細胞増殖の制御に直接関係する特定のカリウムチャンネルの同定の不足は、現在でも、より特異的で効果的な治療プロトコルの記述を妨げている。
【発明の概要】
【発明が解決しようとする課題】
【0003】
したがって、本発明の基礎となる技術的な問題は、ヒト細胞増殖に対する特定の観点から、細胞増殖に対し明白な割当てが可能な細胞周期分裂に対する様々な効果を有するカリウムチャンネルの集合の中の生物学的な成分を同定することであった。かかる技術問題に対する解答は、特許請求の範囲で特徴づけられる態様を提供することにより達成される。
【課題を解決するための手段】
【0004】
即ち、本発明の要旨は、
核酸分子の望まれていない発現または過剰発現によって引き起こされる疾患の治療用薬物の設計方法であって、
(a)特異的かつ効力の高い薬物を同定する工程;
(b)部位特異的突然変異誘発およびキメラタンパク質研究による、前記薬物の結合部位を同定する工程;
(c)(ポリ)ペプチドの結合部位および前記薬物の構造の両方を分子モデリングする工程;ならびに
(d)(ポリ)ペプチドに対するその結合特異性を改良するために薬物を改変する工程、
を含み、該核酸分子が、
(A)配列番号:3または4のアミノ酸配列を有するポリペプチドをコードする核酸分子を含有してなる核酸分子;
(B)配列番号:13または14のDNA配列を有する核酸分子を含有してなる核酸分子;
(C)(A)または(B)の核酸分子の相補的な鎖にハイブリダイズする核酸分子;または
(D)(C)の核酸分子の配列に縮重した核酸分子、
である、ヒトK+ イオンeagチャンネルの機能を有する(ポリ)ペプチドをコードする核酸分子を含有してなる、治療用薬物の設計方法
に関する。
【発明の効果】
【0005】
本発明により、細胞増殖に対し明白な割当てが可能な細胞周期分裂に対する様々な効果を有するカリウムチャンネルの集合の中の生物学的な成分が同定される。
【図面の簡単な説明】
【0006】
【図1】Figure 1。機能時間での野生型(円)およびreag発現CHO細胞の増殖。細胞を96穴ディッシュで培養し、示された時間にテトラゾリウム塩MTT6 (50μg/ml)をプレートに添加した。加湿雰囲気下に(37℃、5%CO2 )4時間インキュベーション後、2容量の1M HCl中10%SDSを添加することにより反応を止めた。生存細胞で産生された青色ホルマザン結晶を1晩かけて可溶化し、得られた色調を指示された波長での光学密度として測定した。細胞増殖のトランスフェクションの起こり得る非特異的影響は無視できる。なぜなら、a)異なる種由来の3つの独立した細胞株で(ラット、ハムスターおよびヒト)結果が類似した;b)異なる独立したクローンでのトランスフェクションが同一の結果を与えた;およびc)同じベクターでの異なるカリウムチャンネル(Kv1.4)でのトランスフェクション(したがって、同じ部位で再結合する傾向を有する)がWTと匹敵する結果を与え、reagトランスフェクションの効果を再現しなかったからである。
【図2】Figure 2。0.5%FCS存在下での、野生型(円)およびreag発現(三角)CHO細胞の増殖。この血清濃度では正常細胞の成長を維持できないが、トランスフェクトされた細胞ではほとんど3周期完了する。方法はFigure 1と同様。
【図3】Figure 3。通常濃度(10%)または低濃度(0.5%)のFCSの存在下での、異なるカリウムチャンネルを発現するCHO細胞でのDNA合成。対照細胞、WTまたはKv1.4でトランスフェクトされた細胞で、DNA合成レベルは低血清濃度の存在下で有意に下がり、一方、reag発現細胞は、高血清濃度と同様の複製レベルを維持した。
【図4a】Figure 4。(A)野生型、Kv1.4トランスフェクトまたはreagトランスフェクトNIH3T3細胞を有するプレートの写真。細胞を低密度で播種し、標準条件下で野生型細胞をコンフルエンスに達するまで成長させた。細胞を次いでメタノールで固定し、ギムエザブルーで染色した。これらの条件下で、野生型およびKv1.4発現細胞は両方とも単層で成長し、一方、reag発現細胞は、フォーカスを形成した。(B)rKv1.4でトランスフェクトされた細胞および野生型細胞と比較したreagトランスフェクトNIH−3T3細胞のフォーカス形成。ベクター対照(pcDNA3トランスフェクト細胞)は、野生型細胞と同様の表現型を与えた(示さず)。一過性のトランスフェクションをリン酸カルシウムを用いて行なった(33)。対照細胞がコンフルエンスに達するまで細胞をリッチ培地で維持し、次いでメタノールで固定し、ギムエザブルーで染色した。
【図4b】Figure 4。(A)野生型、Kv1.4トランスフェクトまたはreagトランスフェクトNIH3T3細胞を有するプレートの写真。細胞を低密度で播種し、標準条件下で野生型細胞をコンフルエンスに達するまで成長させた。細胞を次いでメタノールで固定し、ギムエザブルーで染色した。これらの条件下で、野生型およびKv1.4発現細胞は両方とも単層で成長し、一方、reag発現細胞は、フォーカスを形成した。(B)rKv1.4でトランスフェクトされた細胞および野生型細胞と比較したreagトランスフェクトNIH−3T3細胞のフォーカス形成。ベクター対照(pcDNA3トランスフェクト細胞)は、野生型細胞と同様の表現型を与えた(示さず)。一過性のトランスフェクションをリン酸カルシウムを用いて行なった(33)。対照細胞がコンフルエンスに達するまで細胞をリッチ培地で維持し、次いでメタノールで固定し、ギムエザブルーで染色した。
【図5】Figure 5。電圧クランプ条件下でのMCF7細胞の脱分極により導かれる電流。左のトレースは全体の細胞電流であり、右のトレースは削除されたアウトサイドアウトパッチにより得られる。全体的な電流およびI−V関係の両方が(CおよびD)reag電流を暗示する。
【図6】Figure 6。5μMアステミゾールの存在下または非存在下での、電圧クランプされたアウトサイドアウト膜パッチの0mVでのシングルチャンネル活性。ピペット溶液は、140mM KCl、10mM BAPTA、10mM HEPES pH 7.2を含む;浴溶液は、140mM NaCl、2mM CaCl2 、2mM MgCl2 、2.5mM KCl、10 HEPES pH 7.2を含む。
【図7】Figure 7。A.異なるeagブロッカー下での、MCF7細胞のDNA合成。B.アステミゾール、グリベンクルアミド(glibenclamide)およびテルフェナジンの存在下でのHEK293DNA合成レベル。
【図8】Figure 8。MCF7細胞でのDNA合成の2つのH1アンタゴニストの効果に対する投与量−応答曲線(LY 91241およびアステミゾールそれぞれに対して7mMおよび10mMのIC50)。
【図9】Figure 9。ヘキスト33342で染色された、対照(処理なし、A)およびアステミゾール処理(B)MCF7細胞の蛍光像。Bに、細胞核の同様の表面、および非常に低い細胞密度(細胞死による)を認める。
【図10−1】Figure 10。ラット配列およびウシ配列と比較したヒト脳由来のヒトeag cDNAのヌクレオチド配列。いずれかの配列で異なったヌクレオチドを示す位置に影をつけた。
【図10−2】Figure 10。ラット配列およびウシ配列と比較したヒト脳由来のヒトeag cDNAのヌクレオチド配列。いずれかの配列で異なったヌクレオチドを示す位置に影をつけた。
【図10−3】Figure 10。ラット配列およびウシ配列と比較したヒト脳由来のヒトeag cDNAのヌクレオチド配列。いずれかの配列で異なったヌクレオチドを示す位置に影をつけた。
【図10−4】Figure 10。ラット配列およびウシ配列と比較したヒト脳由来のヒトeag cDNAのヌクレオチド配列。いずれかの配列で異なったヌクレオチドを示す位置に影をつけた。
【図10−5】Figure 10。ラット配列およびウシ配列と比較したヒト脳由来のヒトeag cDNAのヌクレオチド配列。いずれかの配列で異なったヌクレオチドを示す位置に影をつけた。
【図10−6】Figure 10。ラット配列およびウシ配列と比較したヒト脳由来のヒトeag cDNAのヌクレオチド配列。いずれかの配列で異なったヌクレオチドを示す位置に影をつけた。
【図10−7】Figure 10。ラット配列およびウシ配列と比較したヒト脳由来のヒトeag cDNAのヌクレオチド配列。いずれかの配列で異なったヌクレオチドを示す位置に影をつけた。
【図10−8】Figure 10。ラット配列およびウシ配列と比較したヒト脳由来のヒトeag cDNAのヌクレオチド配列。いずれかの配列で異なったヌクレオチドを示す位置に影をつけた。
【図10−9】Figure 10。ラット配列およびウシ配列と比較したヒト脳由来のヒトeag cDNAのヌクレオチド配列。いずれかの配列で異なったヌクレオチドを示す位置に影をつけた。
【図10−10】Figure 10。ラット配列およびウシ配列と比較したヒト脳由来のヒトeag cDNAのヌクレオチド配列。いずれかの配列で異なったヌクレオチドを示す位置に影をつけた。
【図10−11】Figure 10。ラット配列およびウシ配列と比較したヒト脳由来のヒトeag cDNAのヌクレオチド配列。いずれかの配列で異なったヌクレオチドを示す位置に影をつけた。
【図11−1】Figure 11。対応するウシ、マウスおよびラット配列と比較した、ヒトeag cDNA転写から得られる両スプライス変異体のアミノ酸配列。黒いボックスは、いずれかの配列での異なる残基を示す。
【図11−2】Figure 11。対応するウシ、マウスおよびラット配列と比較した、ヒトeag cDNA転写から得られる両スプライス変異体のアミノ酸配列。黒いボックスは、いずれかの配列での異なる残基を示す。
【図11−3】Figure 11。対応するウシ、マウスおよびラット配列と比較した、ヒトeag cDNA転写から得られる両スプライス変異体のアミノ酸配列。黒いボックスは、いずれかの配列での異なる残基を示す。
【図11−4】Figure 11。対応するウシ、マウスおよびラット配列と比較した、ヒトeag cDNA転写から得られる両スプライス変異体のアミノ酸配列。黒いボックスは、いずれかの配列での異なる残基を示す。
【図11−5】Figure 11。対応するウシ、マウスおよびラット配列と比較した、ヒトeag cDNA転写から得られる両スプライス変異体のアミノ酸配列。黒いボックスは、いずれかの配列での異なる残基を示す。
【図11−6】Figure 11。対応するウシ、マウスおよびラット配列と比較した、ヒトeag cDNA転写から得られる両スプライス変異体のアミノ酸配列。黒いボックスは、いずれかの配列での異なる残基を示す。
【図11−7】Figure 11。対応するウシ、マウスおよびラット配列と比較した、ヒトeag cDNA転写から得られる両スプライス変異体のアミノ酸配列。黒いボックスは、いずれかの配列での異なる残基を示す。
【図11−8】Figure 11。対応するウシ、マウスおよびラット配列と比較した、ヒトeag cDNA転写から得られる両スプライス変異体のアミノ酸配列。黒いボックスは、いずれかの配列での異なる残基を示す。
【図11−9】Figure 11。対応するウシ、マウスおよびラット配列と比較した、ヒトeag cDNA転写から得られる両スプライス変異体のアミノ酸配列。黒いボックスは、いずれかの配列での異なる残基を示す。
【図11−10】Figure 11。対応するウシ、マウスおよびラット配列と比較した、ヒトeag cDNA転写から得られる両スプライス変異体のアミノ酸配列。黒いボックスは、いずれかの配列での異なる残基を示す。
【図12】Figure 12。ヒト脳、ヒト乳腺およびMCF−7細胞の全RNA由来のRT−PCR。増幅は、脳で予期されるサイズのheag1およびheag2に対応する2つの特異的な断片を提供し、MCF−7細胞でheag1に対応するバンドを提供し、一方、正常胸RNAで増幅は検出されなかった。
【図13】Figure 13。高い細胞外カリウムでの活性化の電圧依存、2電極電圧クランプ:コンダクタンス−電圧プロットにおいて、半活性化に対する電圧は、reagチャンネルに関してheagチャンネルで40mV右にシフトする。
【図14】Figure 14。示されたDNAでトランスフェクトされたNIH−3T3細胞の半固体培地でのコロニー形成。細胞を0.55%寒天培地の層の上に0.3%寒天を含むレギュラー培地で培養した。直径0.1mmより大きいコロニーをトランスフェクションの14日後に評価した。顕微鏡範囲で少なくとも10を数えるコロニーの平均数は、トランスフェクションに用いられるDNA1μg当たりで表した(「トランスフェクション緩衝液」および「処理なし」のレーンを除く、ここで数字は絶対値である)。reagおよびKv1.4はpcDNA3またはpTracerをCMVベクターのどちらかを使用してトランスフェクトした。
【図15】Figure 15。(A)異なるヒト組織および293細胞由来のRNAのRT−PCR産物のサザンブロット。トランスフェリンレセプター(TFR)シグナルが底に示される。(B)異なるヒト細胞株および一次培養での乳上皮細胞(Epith.cell)由来の全RNAのRT−PCR産物のサザンブロット解析。TRFシグナルが底に示される。
【図16】Figure 16。(A)アンチセンスODNでの、heag発現腫瘍細胞株の処理。(B)SHSY−5Y神経芽腫細胞でのheag電流。(C)アンチセンスODNで処理されたSHSY−5Y細胞での電流密度。(D)heagに特異的なアンチセンスODNによる、ヒト癌細胞(EFM−19、HeLaおよびSHSY−5Y)でのDNA合成の阻害。
【図17−1】Figure 17。(A)速く成長してキャリアマウスの皮膚をすぐに傷つける攻撃的な腫瘍を誘導するCHOhEAG細胞の皮下埋め込み。2×106 細胞の埋め込み後3週間で撮られた写真。(B、C)埋め込み後2週間(B;平均±S.E.M.;p=0.002)または3週間(C;平均±S.E.M.;p=0.03)の両方で、CHOhEAG腫瘍の平均の質量はCHOKv腫瘍のものより有意に大きかった。インサイチュで写真を撮られた(D)CHOhEAG腫瘍および(E)CHOKv腫瘍。主に肉眼で見える相違はより濃い色およびCHOhEAG腫瘍の皮膚への固定である。(F、G)CHOhEAG(F)およびCHOKv(G)腫瘍を切り開き、前者でより広い範囲の壊死を示した(矢頭)。(H、I)より大きな程度の壊死および皮膚への固定がまた、パラフィン包理およびヘマトキシリン−エオシン染色後、顕微鏡的に明らかである。組織構造は両方の顕微鏡写真で匹敵するが、(H)で非常に大きな壊死面積が観察され(矢頭)、皮下脂肪と腫瘍の間に境界はない。(目盛りバー、100μm)。(J)これらの像の定量的測定では、CHOKv腫瘍での生命範囲の平均の広さは、CHOhEAG腫瘍のものより有意に大きかった(平均±S.E.M.;p<0.0005)。
【図17−2】Figure 17。(A)速く成長してキャリアマウスの皮膚をすぐに傷つける攻撃的な腫瘍を誘導するCHOhEAG細胞の皮下埋め込み。2×106 細胞の埋め込み後3週間で撮られた写真。(B、C)埋め込み後2週間(B;平均±S.E.M.;p=0.002)または3週間(C;平均±S.E.M.;p=0.03)の両方で、CHOhEAG腫瘍の平均の質量はCHOKv腫瘍のものより有意に大きかった。インサイチュで写真を撮られた(D)CHOhEAG腫瘍および(E)CHOKv腫瘍。主に肉眼で見える相違はより濃い色およびCHOhEAG腫瘍の皮膚への固定である。(F、G)CHOhEAG(F)およびCHOKv(G)腫瘍を切り開き、前者でより広い範囲の壊死を示した(矢頭)。(H、I)より大きな程度の壊死および皮膚への固定がまた、パラフィン包理およびヘマトキシリン−エオシン染色後、顕微鏡的に明らかである。組織構造は両方の顕微鏡写真で匹敵するが、(H)で非常に大きな壊死面積が観察され(矢頭)、皮下脂肪と腫瘍の間に境界はない。(目盛りバー、100μm)。(J)これらの像の定量的測定では、CHOKv腫瘍での生命範囲の平均の広さは、CHOhEAG腫瘍のものより有意に大きかった(平均±S.E.M.;p<0.0005)。
【図18】Figure 18。rEAGトランスフェクトCHO細胞の増殖アッセイ(A〜C)。10%(黒塗りの記号)または0.5%(白抜きの記号)のウシ胎仔血清での、処理なしの細胞(三角)と比較したrEAGでトランスフェクトされたCHO細胞(円)の成長曲線。値は、培養12時間後測定されたものをいい(プロットで時間0)、同じプレートで8ウェルの平均±S.E.M.を表す。細胞株をpcDNA3ベクターでコードされるG−418耐性を介する選択により確立した。MTT加水分解(22)を使用して、生存能力がある細胞の代謝活性を測定した。12時間培養後、血清を注意深く希釈した。(B)S期ブロックの除去後最初の12時間の間の代謝活性の増加。細胞同調のために、12時間培養培地にチミジン2mMを添加した。追加の12時間、培地からチミジンを除去し、次いで第2停止パルスを12時間適用した。代謝活性およびDNA合成の測定のために、次いで細胞をトリプシン処理し培養した。(C)10%FCSで12時間のインキュベーション、または0.5%FCSの存在下(24時間のインキュベーション)でのS期ブロックの除去後最初の12時間の間のBrdU取り込み。BrdUの取り込みを、製造業者の指示にしたがってBoehringer−Mannheimの「BrdUラベリングおよび検出キット」を使用して、測定した。バーは、野生型CHO細胞(白抜きバー)、Kv1.4トランスフェクト(影をつけたバー)およびeag−トランスフェクト(中実バー)に対する平均±S.D.を表す。BrdUの取り込みを、抗BrdU抗体に結合するペルオキシダーゼによりABTSTM基質で生じる405nm(参照490nm)での光学密度として定量する。
【発明を実施するための形態】
【0007】
それゆえに、本発明は、
(a)配列番号:3または4のアミノ酸配列を有するポリペプチドをコードする核酸分子を含有してなる核酸分子;
(b)配列番号:13または14のDNA配列を有する核酸分子を含有してなる核酸分子;
(c)(a)または(b)の核酸分子の相補的な鎖にハイブリダイズする核酸分子;または
(d)(c)の核酸分子の配列に縮重した核酸分子、
である、ヒトK+ イオンeagチャンネルの機能を有する(ポリ)ペプチドをコードする核酸分子を含有してなる核酸分子に関する。
【0008】
本発明の核酸分子は、ラットeagチャンネルのヒトホモローグであるかまたはそれを含有する(ポリ)ペプチドをコードする。この点において、用語「ヒトK+ イオンeagチャンネルの機能を有する(ポリ)ペプチドをコードする核酸分子を含有してなる核酸分子」は、前記最初に記載された核酸分子が前記(ポリ)ペプチドを単にコードすることを意味してもよい。したがって、それは前記2番目に記載された核酸分子と同一であってもよい。他方、それらは、調節領域または他の非翻訳領域を含有してもよい。さらなる態様において、前記最初に記載された核酸は、異種構造のタンパク質材料、したがって、例えば、融合タンパク質を生じさせる材料をコードしてよい異種構造の核酸を含んでいてよい。配列番号:13および14のDNA配列は、本発明の(ポリ)ペプチドをコードする核酸配列のスプライス変異体(variants)であることを、さらに指摘する。対応するアミノ酸配列を、配列番号:3および4に記載する。
【0009】
用語「ヒトK+ イオンeagチャンネルの機能を有する」は、本発明に関連して使用され、次の意味を有する:チャンネルは、約6pSの0mVで、非対称のカリウムでのシングルチャンネルコンダクタンスを有する。この値は、約7pSの値が測定されるラットチャンネルからヒトチャンネルを明確に区別する。さらにまたは他方、上記用語は、次の意味を有していてもよい:reagに対する400μMと比較して、アフリカツメガエル卵母細胞で発現される時、キニジンに対し約1mMのIC50を有する。さらに、2電極電圧クランプを用いて高い細胞外カリウムでの活性化の電圧依存性を測定する場合、コンダクタンス−電圧プロットにおいて、半活性化に対する電圧は、reagチャンネルに関してheagチャンネルで約40mV以上右にシフトすることが見出された(Figure 13を参照のこと)。前記特徴の単一または組み合わせに基づいて、、本発明のヒトイオンチャンネルと、先行技術のチャンネル、特にラットイオンチャンネルとの間の機能に基づく区別は、当業者のさらなる苦労なしに可能である。好ましくは、チャンネルは全ての列挙された機能を有する。前記値は、本明細書で記載された実験計画で得られ得る値を指す。異なる発現系の使用等の実験パラメーターの変更は、当業者に知られているように、また前記値を変化させる。さらに、これらの態様はまた、本発明の範囲に含まれる。
【0010】
用語「ハイブリダイズする(hybridizing)」は、本発明に従って使用される場合、ストリンジェントまたは非ストリンジェントなハイブリダイゼーション条件に関する。好ましくは、それはストリンジェント条件に関する。かかるハイブリダイゼーション条件は、例えば、Sambrookによる、“Molecular Cloning,A Laboratory Manual”、Cold Spring Harbor Laboratory(1989)N.Y.、Ausubel、“Current Protocols in Molecular Biology”、Green Publishing AssociatesおよびWiley Interscience、N.Y.(1989)、またはHigginsおよびHames(編)“Nucleic acid hybridization,a practical approach”IRL Press Oxford,Washington DC,(1985)に記載された慣用のプロトコルによって確立されてよい。ハイブリダイズする分子または上記もう1つの(d)に該当する分子は、(a)または(b)で同定される分子の断片も含有する、ここでヌクレオチド配列は、配列番号:13または14でのその対照物(counterpart)と同一である必要はなく、前記断片は上記で示されるような機能を有するものである。
【0011】
1つのかかるストリンジェントなハイブリダイゼーション条件の例は、65℃での4XSSC、続く65℃での0.1XSSCでの1時間の洗浄によるハイブリダイゼーションである。他方、典型的なストリンジェントなハイブリダイゼーション条件は、42℃での50%ホルムアミド、4XSSC中である。かかる非ストリンジェントなハイブリダイゼーション条件の例は、50℃での4XSSCまたは42℃での30〜40%ホルムアミドでのハイブリダイゼーションである。分子にハイブリダイズする相補的な鎖は、本発明のポリペプチドの断片、アナログまたは誘導体をコードし、例えば、アミノ酸および/またはヌクレオチドの1つまたは複数の欠失、1つまたは複数の挿入、1つまたは複数の置換、1つまたは複数の付加および/または1つまたは複数の組換えまたは当該分野において既知のいずれかの他の1つまたは複数の修飾の単独または組み合わせを介して、前記アミノ酸配列またはそれらの基礎となる1つまたは複数のヌクレオチド配列と異なるものを含有する。PESTFINDプログラム(Rogers,Science 234(1986)、364〜348)を使用して、不安定なタンパク質に特徴的に存在するPEST配列(プロリン、グルタミン酸、セリンおよびトレオニンがリッチである)を同定できる。かかる配列は、タンパク質の安定性および任意にタンパク質の活性を増加するために、本発明のポリペプチドから除去してもよい。本発明による核酸分子におけるかかる修飾を導入する方法は、当業者によく知られている。本発明はまた、その配列が遺伝コードの縮重のために前記核酸分子のいずれのヌクレオチド配列とも異なっている核酸分子に関する。本発明のタンパク質をコードするかかるすべての断片、アナログおよび誘導体は、前記規定するような必須の特徴的な免疫学的および/または生物学的特性が本質的に影響されないままである限り、本発明の範囲内に含まれ、すなわち、本発明の新規核酸分子は、上記の核酸分子によってコードされ、生物学的活性に関して同等のまたは同一の特徴を有する前記ポリペプチドに対する抗体と反応できる1以上のエピトープに対する少なくとも一部の一次構造形態を有するタンパク質またはペプチドをコードする全ヌクレオチド配列を含む。本発明の一部はそれゆえにまた、本発明による核酸分子に含まれる核酸配列によりコードされる前記同定されたポリペプチドの少なくとも機能的部分を含有してなるポリペプチドをコードする核酸分子に関する。
【0012】
本発明者らは最近、G1−SおよびG2−M移行の両方においてサイクリン依存性キナーゼ(細胞周期調節における重要な分子)の活性化直後に強くダウンレギュレートされるカリウムチャンネル(reag)について記載している5 。サイクリン依存性キナーゼの活性化後に、電圧依存性ナトリウムブロックによりK+ 電流(current)は阻害される。それは細胞周期の全ての段階において明白であるというわけではない。本明細書に存在する実験は、eagがまた、細胞周期により制御されるのに加えて、細胞増殖および細胞成長に直接影響し得るか否かを決定することを意図する(20)。本発明およびヒト細胞での(疾患に関連する)増殖を評価するための適切なシステムの開発の観点にしたがって、細胞周期におけるチャンネルの関係が、細胞周期の進行によって制御されるだけでなく、細胞周期の進行のレギュレーターでもあるというような、両方向に進むかどうかの研究を、さらに試みた。
【0013】
このラット由来のイオンチャンネルシステムで得られた結果は、異なる種から得られた3つの異なる細胞株(チャイニーズハムスター−CHO−、ヒト−HEK293−およびマウス−NIH3T3−)において、チャンネルが、サイトメガロウイルスプロモーターの制御下のチャンネルDNAを含むプラスミドによる細胞のトランスフェクションの後に過剰発現される場合、増殖速度がより速くなることを示す。Figure 1およびFigure 18aは、通常濃度のウシ胎仔血清(10% FCS)の存在下の、CHO細胞の培養物における代謝活性の上昇を示す。これらの通常条件下で、reagトランスフェクト細胞は、非トランスフェクト細胞(WT)より数倍速く成長する。
【0014】
Figure 2は、非常に低濃度のウシ胎仔血清(0.5% FCS)での比較例を示す。これらの低い血清濃度では、野生型細胞が成長できない;数時間後、細胞は死に始める。しかしながら、reagトランスフェクト細胞は、同条件下に増殖可能である。成長因子の欠如により誘導される成長停止に打ち勝つ能力は、悪性の形質転換の1つの典型的な特性である(Figure 18参照)。
【0015】
培養における増殖の追跡のために代謝活性以外のものを使用することができる。(分裂に付される)S期に入る細胞のみがDNAを合成するために、DNA合成の測定は、細胞成長速度のより直接的な評価である。また、DNA合成は、reagトランスフェクト細胞において血清非依存的となる、すなわち、成長が、成長因子の非存在下に維持される(一方、それは、非トランスフェクト細胞のプログラムされた死を誘導する)。これはFigure 3に描かれており、5−ブロモ−2’−デオキシウリジン7-10(BrdU)の取り込みを用いて、CHO細胞において、10%FCSまたは0.5%FCSの存在下にDNA合成をモニターした。野生型またはラット脳由来の不活性化された電圧依存性カリウムチャンネルでトランスフェクトされた細胞(Kv1.4)と対照的に、reag発現細胞で通常FCS濃度または低FCS濃度の存在下に合成されるDNAの量に有意な相違はない。HEK−293細胞で上皮成長因子(EGF)をまたはCHO細胞で血小板由来成長因子(PDGF)を使用して、同様の実験を行ない、本質的に同一の結果を得た。純粋な成長因子を使用して、全血清の使用により導入される複雑さを避けた。
【0016】
細胞増殖に対するeagの効果をより直接的に試験するために、チミジン停止により細胞周期のS期で同時に発生する細胞での5−ブロモ−デオキシウリジン(BrdU)の取り込みを通してDNA合成を測定した(23)。上記知見と一致して、細胞周期のS期が進行できる場合、reag発現CHO細胞(CHOrEAG)は、より高い代謝活性(Fig 18B)およびBrdU取り込みの増加(Fig 18C)を示した。これらの結果は、CHOrEAG細胞でより速い増殖速度を示すいずれの場合においても、分裂停止期の間により多くeagトランスフェクト細胞がS期に入ること、および/またはDNA合成が上昇することを示唆する。低血清の存在下に、BrdU取り込みは、野生型細胞でよりも、CHOrEAGで有意に高かった(Fig 18C)。
【0017】
さらにもう1つの細胞株、NIH3T3は、これらの細胞が非常に強く接触阻害されるため(すなわち、培養がコンフルエンシーに達した時、それらの成長がストップする)、しばしば腫瘍形質転換アッセイに使用されている。これらは、野生型細胞で均一な単層を生じる。この株の悪性の形質転換(癌遺伝子の発現を介する)は、通常この特性の損失を誘導し、NIH3T3細胞はいくつかの層の細胞からなるコロニーを形成し始める。これは、reag DNAによるトランスフェクション後に見られ得、いくつかの非依存性クローンでかかるフォーカス(foci)の形成を誘導した(Fig.4AおよびB)。形質転換活性に対するもう1つの標準的な試験は、付着するための基質が得られない場合にコロニーで成長するNIH3T3細胞の能力である。これを試験するために、細胞を寒天を含む培地で培養する。ここで、寒天は細胞とプレートの表面との間の接触を妨げるであろう。これらの条件下に、野生型NIH3T3細胞は成長できず、一方、reagを発現する細胞は、単にプレートの視覚観察により検出可能でさえある大きなコロニーを形成した。表Iは、reag(しかし、rKv1.4ではない)トランスフェクト細胞は、トランスフェクションで使用されたベクターにかかわらず、0.3%寒天を含む半個体の培地でコロニーを形成した(Fig.14)ことを示す。全ての上記結果は、eagの形質転換能を示す。
【0018】
要するに、トランスフェクトされた細胞から得られた結果は、reagが少なくともある条件下に癌遺伝子特性を示し得ることを示す。
【0019】
一旦、reagの形質転換能力を本発明により決定した後、ヒト癌細胞でのそれぞれのチャンネルの発現を研究した。この研究のために、胸腺癌(breast adenocarcinoma)の胸膜滲出液(pleural effusion)から初めに得られた細胞株MCF−7を使用した。該株は、エストロゲンレセプター陽性ならびにエストロゲン感受性であり、かつ相対的によく分化する。次の戦略は、まずeagと同様の機能的電流の存在について電気生理学的および薬理学的に試験し、次に分子レベルで対応するチャンネルの同定を試みることであった。しかしながら、かかる同定に対する慣用のアプローチで失敗した。
【0020】
すなわち、ほとんどの細胞で電流密度は低過ぎて、細胞電流全体の信頼度の高い測定はできなかった。低い電流密度は、パッチチャンプ(champ)に全細胞形態を使用するチャンネル特性の正確な測定を妨げた。それゆえに、かかる低い電流密度に遭遇したため、もう1つのアプローチに頼った。かかる細胞当たりの低いチャンネル数のために、1つ(または少し)のチャンネルを含む膜のパッチを取り出し、かつ単分子レベルで特徴付けができる特別なパッチチャンプ法により、チャンネルの機能的な特性を特徴付けることができるのみである。このアプローチでは、シングルチャンネルコンダクタンス、薬理学特性、電圧依存性、平均開口時間等の単分子レベルでの特性を比較するためにも、シングルチャンネル測定をあてにした。実際に、ほとんどの膜パッチにおいて速度論、電圧依存性、および薬理学に関してreagと矛盾しないいくつかの特性を有するチャンネルが、したがって同定され得るであろう。Figure 5は、全細胞電流が、電流−電圧関係と共に、ナイスタチンパッチ条件およびシングルチャンネル電流下にMCF7細胞から得られたことを示す。ほとんど脱分極した電圧での速度論での相違にもかかわらず、ヒト細胞でのチャンネルの電圧依存性は、reagチャンネルの電圧依存性に対して高く暗示される。さらに、推定上のヒト−eagのシングルチャンネル特性はまた、reagのものに非常によく似ている。
【0021】
さらに、分子レベルで前記チャンネルを単離する標準アプローチはまた成功しなかった。いくつかの他のグループが、ヒトeagをコードする遺伝子を単離しようと試みてきたおよび/または未だ試みている最中であるが、成功しておらず、これはラットeagチャンネルが、既に1994年に発表されたという事実にもかかわらずである。例えば、WarmkeおよびGanetzky(Proc.Natl.Acad.Sci.USA 91(1994)、3428〜3442)は、慣用の技術を使用してヒトeag遺伝子をクローニングすることについて詳細に説明した。しかしながら、彼らは成功せず、彼らはh−ergと呼ぶ(またはHERGと呼ぶ)新規なeag関連遺伝子をクローニングした。さらに、Wymoreら、Circulation Res.80(1997)、261〜268は、全eag/ergスーパーファミリーに渡って保存される増幅のためのプライマーが使用されるという事実にもかかわらず、eag特異的クローンが、ヒト心臓由来のcDNAライブラリーで検出され得ないことを報告した。したがって、HERGはその分野の他の研究者によって系統的に検出されたが、ファミリーのメンバーの配列に基づく縮重オリゴヌクレオチドを用いる標準的なアプローチは不成功であることを明らかにした。有意に、ヒトeag遺伝子をクローニングするための、ほとんどのこれらのアプローチは、脳のライブラリーを使用してなされた。これらの先行技術データの組み合わせからの結論は、ヒトeag遺伝子は最も明白な源、すなわち脳組織を使用する慣用の技術によってクローニングできないであろうということであった。代わりにHERGクローンの繰り返された同定は、ほとんどたぶん脳ライブラリーでの比較的多数のHERG転写物のためであり、また2つのチャンネル間の高い相同性のためである。それゆえに、異なった戦略が、eagチャンネルに対しより特異的にスクリーニングを行なえるように案出されなければならなかった。最初に、本明細書の前記のように、reagに機能的に類似したチャンネルを発現する細胞株を同定した。ついでラット、ウシおよびマウスのeag間で保存されるが、HERGとは異なる配列に基づいて縮重オリゴヌクレオチドを設計した。これらのプライマーを用いて、PCRによってMCF7細胞から得られたcDNAを増幅し、予期されたサイズのバンドを適切なベクターにクローニングし、配列を決定した。増幅断片は、チャンネルタンパク質のコア領域内の約400bpに対応し、reag配列に対してDNAレベルで90%の同一性を有し、アミノ酸レベルで99%の同一性を有した。しかしながら、この段階では、こうして同定されたクローンが何に対応するのかはまだかなり不明瞭であった。例えば、eagファミリーのさらなるメンバーが同定されたということは、かなりの可能性があった。これは、脳のライブラリーに対する多くの試みにもかかわらず、誰もヒトeag遺伝子をクローニングすることができなかった、およびMCF7株は胸癌由来株である、という事実の観点から、特に真実である。
【0022】
MCF7細胞は不死の細胞であるために、遺伝子の多数が変異すると想定される。最初から、ヒトeagチャンネルは、もし少しでもこの細胞株で発現されるなら、変異されることが予測されたであろう。この仮定の下で、この細胞株は、所望の遺伝子の単離に少しでも使用され得るかどうかは、かなり不確実であった。
【0023】
脳のライブラリーからのヒトeag遺伝子のクローニングの先行技術の失敗および前記不死化した細胞株の不確実性のために、ライブラリーに対するもう1つの源が必要であった。400bpの断片がそれゆえに正常ヒト胸cDNAライブラリーをスクリーニングするために使用された。胸癌細胞でのeagの存在のため、かかるライブラリーはheagクローンを含有することが予期された。しかしながら、驚くべきことに、2×106 ファージのスクリーニングの後、ヒトeagクローンは、前記ライブラリーで同定され得なかった。これは、チャンネルが正常な組織でなく腫瘍細胞でのみ発現されるという可能性を高める。上記ライブラリーから直接PCR増幅によりheag断片について調べるための特異的なオリゴヌクレオチド、すなわち5’−CCAAACACACACACCAGC、5’−CGTGGATGTTATCTTTTTGGを設計したが、このライブラリーでいずれのeagクローンの存在に対する証拠も見出せなかった。上記先行技術の結果を考慮し、さらに驚くべきことに、同じプライマーが正常なヒト脳のcDNAライブラリーでheagを検出し、それゆえにスクリーニングされるようになった。最初に、MCF7細胞から得られたプローブを、106 ファージを調べるのに使用した。この手順により、ヒトeag由来の1.6kbp断片を単離できた。この断片を次に同じライブラリー由来の2×106 ファージのスクリーニングのプローブとして使用した。いくつかの独立したクローンを単離したが、それらのいずれも全長のクローンではなかった。さらに、1つのクローンのみが配列の5’末端を含み、一方、それらの2つが3’末端および3’非コード領域の一部を含んだ。チャンネルをコードする核酸配列の豊富な制限部位が、cDNAのこの多数の断片化を誘導しているようである。例えば、EcoRIをEcoRI部位においてλ−gt10ファージにクローニングされたライブラリーの挿入物を抽出するのに使用した場合、この慣用の研究方法では体系的に分子の5’末端を見つけることはできなかった(クローンの400位にEcoRI部位がある)。それゆえ、プールされた陽性クローンを、PCRによって再度スクリーニングし、開始コドンの増幅を試み、そしてこの手段によってのみ、このATGを含む1ファージを単離することが可能であった。heagの2つのスプライス変異体をクローニングした。両方とも脳組織で発現した。heag 1およびheag 2について得られた配列ならびにそれらの予想アミノ酸配列をFigure 10およびFigure 11に示し、ファミリーの他のメンバーと比較した。
【0024】
予想したアミノ酸配列は、Occhidoro(27)による本発明の優先日の後に公表された配列と同一であり、reagと97.7%同一である。前記のように、第2(81bpさらに長い)のスプライス変異体(heag 2)も単離し、該変異体はウシおよびマウスeagチャンネルで報告されたもの(28)と類似しており、スプライス挿入は3種全てで同一であった。heagの染色体の局在を、FISH検出で測定し(29)、染色体lq32.1〜32.3に位置づけた(参照文献26も参照のこと)。
【0025】
MCF−7癌細胞とは対照的に、heagが正常の乳腺で発現されない可能性をさらに調べるために、我々は、ヒト脳、ヒト乳腺、およびMCF−7細胞由来の全RNAを使用し、heagの2つのスプライス変異体間で区別するように設計された2つのオリゴヌクレオチドをプライマーとして使用して、シングルチューブRT−PCR実験を行なった(Fig 12)。ヒト脳において、2つのスプライス変異体が検出され、一方、短いもののみがMCF−7細胞で発現された(これは、逆転写酵素の非存在下における増幅の欠如と共に、RNA調製物のゲノムDNAによる汚染の可能性を除外する)。heagシグナルは、正常乳腺RNAで、この高感度技術を用いて検出されなかった。予備結果は発現が同じ臓器由来の腫瘍細胞に存在することを示唆したため、この結果は全く予期されなかった。さらに、RT−PCR産物のサザンブロット解析後、乳腺でheagプローブとハイブリダイズする弱いバンドを同定した。したがって、これらの矛盾した実験データを考慮すると、胸のheagメッセージの完全な欠如を強く主張することはかなり困難である。
【0026】
さらに、heagの電気生理学的特性(21,30)を、ツメガエル(Xenopus)卵母細胞で試験した。前記のように、前記例外、例えば両チャンネルが同一の条件下に測定された時の、40mVのより脱分極された電位への活性化のシフトを除いて、それらは、それらのものまたはreagに有意な相違はなかった。ツメガエル卵母細胞で発現されるheagチャンネルの電気生理学的観察は、Bijlengaらにより報告されたホース(31)によく対応する。
【0027】
本発明はまた、配列5’−GGGAGGATGACCATGGCTを含有してなる本発明の核酸分子に特異的にハイブリダイズする核酸分子に関する。
【0028】
本発明のこの態様は、本明細書の下記に(例えば実施例5において)さらに詳細に説明するように、細胞増殖を阻害するための特異的なアンチセンス治療に特に有用である。さらに本発明の核酸分子のこの態様はまた、当然、例えばノーザンブロット技術を用いることにより、組織のheag mRNAを特異的に検出するためのプローブとして使用される。ノーザンブロットによる様々な組織のheag mRNA発現の解析は、脳で約9.2kbの強いハイブリダイゼーションシグナルおよび胎盤で同様のサイズの弱いシグナルを明らかにした。心臓、肺、肝臓、骨格筋、腎臓および膵臓は、長い曝露後でさえ陰性であった。さらに、ヒト脳、心臓、気管、副腎、肝臓、腎臓、骨格筋および乳腺由来の全RNA、および脊髄ポリ(A)+ RNA、ならびにアデノウイルスで形質転換された293株由来の全RNA(ヒト非腫瘍細胞株)をシンングルチューブRT−PCRおよびサザンブロットにより解析した。これらの実験条件下に、heagは脳でのみ検出され、そこで、両スプライス変異体を同定した(Fig.15;実施例3)。
【0029】
第1のcDNAが上皮腫瘍細胞株(MCF−7)から単離され、脳組織から単離されなかった(前記参照)ため、脳におけるheagの優先的な発現は興味深かった。他の腫瘍細胞株でheagの存在を解明するために、全RNAをHeLa(子宮頸癌)、SHSY−5Y(神経芽腫)、および乳腺腫瘍由来の株:COLO−824(癌)、EFM−19(癌)、およびBT−474(管癌)から調製した。脳、MCF−7細胞、293細胞由来の全RNA、および乳腺上皮細胞の培養物(全体の乳腺の混合細胞集団を取り囲むことを含む)由来のRNAを対照として用いた。全細胞株はDSMZ(Deutsche Sammlung von Mikroorganismen und Zellkulturen)より得られ、DSMZカタログガイドラインに従って維持された。正常ヒト乳腺上皮細胞をBio Whittakerから購入した。プライマーをheag 1およびheag 2の異なるバンドを増幅するために設計し、これによりゲノムDNA汚染のために間違って陽性になるのを防ぐことができた(逆転写酵素非存在下での対照も行なった)。HeLa、SHSY−5Y、EFM−19およびMCF−7 RNAはheagバンドを示し、一方、COLO−824およびBT−474シグナルは背景と区別がつかなかった(Fig.15B)。培養された上皮細胞および293細胞は(Fig.15A)、陰性であった。前記のように、reagでトランスフェクトされた細胞は癌遺伝子特性を示し得ることが、本発明にしたがって示され得た。したがって、heagの発現がインビボで腫瘍細胞に有利であるかどうかを測定するために、雌のシッド(scid)(厳しい複合免疫不全、32)マウスの脇腹に、チャンネルを発現するCHO細胞(CHOhEAG細胞)の皮下移植を行ない、それは、CHOhEAG腫瘍が、CHOKv腫瘍よりも速く成長し、より攻撃的であるため、heagの発現がインビボで腫瘍細胞の増殖に有利であることを意味することを示し得た。したがって、本発明の核酸分子の態様は、例えば、癌(特に、乳癌、神経芽腫)、乾癬、アルツハイマー病、パーキンソン病、筋萎縮性側索硬化症または多発性硬化症等の神経変性疾患を示すような組織で検出可能な種々の疾患状態で、ヒトeagの発現レベルの定量的および定性的解析に使用され得る。
【0030】
本発明の核酸分子の好ましい態様において、前記核酸分子はゲノムDNA等のDNAである。一方、本発明はまた、ペプチド核酸等の合成または半合成DNA分子またはその誘導体を含む。最も好ましい本発明のDNA分子は、cDNAである。
【0031】
本発明のさらに好ましい態様において、前記核酸分子はRNA、好ましくは、mRNAである。
【0032】
本発明の核酸分子のもう1つの好ましい態様は、融合タンパク質をコードする。例えば、本発明の核酸分子は、FLAGまたはGFP等の検出可能なマーカーにフレーム中で融合し得る。
【0033】
本発明はさらに、本発明の核酸分子を含有するベクター、特にプラスミド、コスミド、ウイルスおよびバクテリオファージに関する。かかるベクターは、適切な宿主細胞で適切な条件下に該ベクターの選択を可能にするマーカー遺伝子等のさらなる遺伝子を含んでもよい。このように、本発明のポリヌクレオチドは、該ベクターにおいて発現制御配列に作動可能に連結され得、原核細胞および真核細胞で発現可能である。該ポリヌクレオチドの発現は、転写可能なmRNAへのポリヌクレオチドの転写を含む。真核細胞、好ましくは哺乳類細胞での発現を確実にする調節エレメントは、当業者によく知られている。それらは通常、転写開始を確実にする調節配列および転写の終結および転写物の安定化を確実にする任意のポリ−Aシグナルを含有する。さらなる調節エレメントとしては、転写エンハンサーならびに翻訳エンハンサーを含んでもよい。原核宿主細胞で発現を可能にする可能な調節エレメントとしては、例えば、大腸菌のlac、trpまたはtacプロモーターを含み、真核宿主細胞で発現を可能にする調節エレメントの例としては、酵母のAOX1もしくはGAL1プロモーターまたは哺乳類および他の動物細胞のCMV−、SV40−、RSV−プロモーター(ラウス肉腫ウイルス)、CMV−エンハンサー、SV40−エンハンサーまたはグロビンイントロンである。転写開始を担うエレメントの他に、かかる調節エレメントはまた、ポリヌクレオチドの下流に、SV40−ポリ−A部位またはtk−ポリ−A部位等の転写終結シグナルを含有してよい。これに関連して、岡山−バーグ cDNA発現ベクターpcDV1(Pharmacia)、pCDM8、pRc/CMV、pcDNA1、pcDNA3(In−vitrogene)、pSPORT1(GIBCO BRL)等の適切な発現ベクターが、当該技術分野で知られている。
【0034】
好ましくは、該ベクターは、発現ベクターおよび/または遺伝子導入ベクターもしくは遺伝子ターゲッティングベクターである。発現ベクターおよび遺伝子ターゲッティングベクターまたは遺伝子導入ベクターは、当該技術分野において既知であり、当業者により本発明の特定の目的に適合させ得る。したがって、レトロウイルス、ワクシニアウイルス、アデノ随伴ウイルス、ヘルペスウイルスまたはウシパピローマウイルス等のウイルス由来発現ベクターは、本発明のポリヌクレオチドまたはベクターの標的細胞集団への送達に使用され得る。当業者によく知られた方法が、組換えウイルスベクターを構築するために使用され得る;例えば、Sambrook、Molecular Cloning、A Laboratory Manual、Cold Spring Harbor Laboratory(1989)N.Y.およびAusubel、Current Protocols in Molecular Biology、Green Publishing Associates and Wiley Interscience、N.Y.(1989)に記載の技術を参照のこと。他方、本発明のポリヌクレオチドおよびベクターは、標的細胞への送達のために、リポソームに再構築され得る。
【0035】
本発明はさらに、本発明のベクターで形質転換された宿主に関する。該宿主は、原核細胞または真核細胞であり得る;上記参照のこと。宿主細胞に存在する本発明のポリヌクレオチドまたはベクターは、宿主細胞のゲノムに統合されていてもよくまたは染色体外で維持されていてもよい。これに関連して、本発明の組換えDNA分子は、相同組換えを介して変異遺伝子を修復するまたは変異遺伝子を作るために、「遺伝子ターゲッティング」および/または「遺伝子置換」に使用され得ることがまた理解されるべきである;例えば、Mouellic、Proc.Natl.Acad.Sci.USA、87(1990)、4712〜4716;Joyner、Gene Targeting、A Practical Approach,Oxford University Pressを参照のこと。好ましくは、宿主は、哺乳類細胞、真菌細胞、植物細胞、昆虫細胞または細菌細胞である。好ましい真菌細胞は、例えば、サッカロミセス属のもの、特にS.セレビジエ(cerevisiae)種のものである。用語「原核(prokaryotic)」は、本発明のタンパク質の発現のためのポリヌクレオチドで形質転換またはトランスフェクトされ得る全ての細菌を含むことを意図する。原核生物宿主は、例えば、大腸菌、S.チフィミュリウム(typhimurium)、セラチア マルセッセンス(Serratia marcescens)およびバチルス サブティリス(Bucillus subtilis)等のグラム陰性細菌ならびにグラム陽性細菌を含み得る。融合された、作動可能に連結された遺伝子を調製し、細菌細胞または動物細胞でそれらを発現する方法は、当該分野においてよく知られている(Maniatisら、Molecular Cloning:A Laboratory Manual、Cold Spring Harbor Laboratory,Cold Spring Harbor,NY,1989)。本明細書に記載の遺伝子構築および方法は、原核生物宿主で本発明のタンパク質を発現するのに利用できる。一般的に、挿入されたポリヌクレオチドの有効な転写を促進するプロモーター配列を含む発現ベクターが、宿主と関連して使用される。発現ベクターは典型的に、複製の起点、プロモーター、ターミネーター、ならびに形質転換された細胞の表現型選択を与えることができる特異的な遺伝子を含む。形質転換された原核生物宿主は、発酵槽で成長でき、当該分野で既知の技術により培養して最適な細胞成長を得ることができる。本発明のポリペプチドは次いで、成長培地、細胞溶解物または細胞膜フラクションから単離することができる。微生物学的または別の方法で発現させた本発明のポリペプチドの単離および精製は、例えば、モノクローナル抗体またはポリクローナル抗体の使用に関連する方法等の予備のクロマトグラフィー分離および免疫学的分離等のいくつかの慣用の手段によることができる。哺乳類細胞に関しては、HEK 293、CHO、HeLaおよびNIH 3T3が好ましい。昆虫細胞に関しては、スポドプテラ フルギペルダ(Spodoptera frugiperda)細胞が最も好ましく、一方、最も好ましい細菌細胞は大腸菌細胞である。
【0036】
本発明はまた、本発明の宿主を培養する工程、および産生された(ポリ)ペプチドを単離する工程を含む、本発明の核酸分子によってコードされた(ポリ)ペプチドの生産方法に関する。
【0037】
使用されるベクター構築に依存して、本発明の(ポリ)ペプチドは、培養培地に輸送されるか、または宿主中で維持され得る。産生される(ポリ)ペプチドを得るための適切なプロトコルは、両方法の(ポリ)ペプチド産生に対して当該分野でよく知られている。
【0038】
本発明はさらに、本発明の核酸分子によってコードされる(ポリ)ペプチド、または、本発明の方法によって生産される(ポリ)ペプチドに関する。新しいチャンネルは、短いアミノ末端領域、おそらく細胞内の5つのメンブレンスパンニングセグメント、膜に入る疎水性のヘアピン、6番目の膜貫通セグメント、サイクリック−ヌクレオチド結合コンセンサス配列を含有する長いC末端細胞質部分、細胞核局在コンセンサス配列、およびおそらくコイルドコイル(coiled−coil)構造を形成する疎水性領域を有する構造を示すと考えられる。本発明のポリペプチドはまた、ヒトK+ イオンチャンネルの機能的断片でもあり得る。「機能的断片」により、ポリペプチドは前記のいくつかのheagの活性を示すことが意味される。組換えDNA技術を用いて、本発明の(ポリ)ペプチドの断片が産生され得る。これらの断片は、例えば上記で示したように本発明で記載されたもののような種々のアッセイシステムを用いて、所望の機能に対して試験できる。好ましくは、該断片は新規イオンチャンネルのC末端部分を含有する。
【0039】
本発明はまた、本発明の(ポリ)ペプチドに対し、特異的な抗体に関する。本発明の抗体は、特に、ヒトeagチャンネルと、マウスeagおよびラットeag等の先行技術のチャンネルを区別し、好ましくは、イオンチャンネルのC末端部分でエピトープに結合する。用語「抗体」はまた、本発明により使用される時、抗体断片もしくはF(ab)2 、Fab’、FvもしくはscFv断片等の誘導体に関する;例えば、HarlowおよびLane、「Antibodies、A Laboratory Manual」、CSH Press 1988、Cold Spring Harbor,NYを参照のこと。好ましくは、本発明の抗体は、モノクローナル抗体である。
【0040】
本発明はまた、本発明の核酸分子、本発明のベクター、本発明のポリペプチドおよび/または本発明の抗体および薬学的に許容できる担体および/または希釈剤および/または賦形剤を含有してなる医薬組成物に関する。
【0041】
適切な医薬担体および希釈剤ならびに賦形剤の例が当該分野でよく知られており、リン酸緩衝生理食塩水、水、油/水エマルジョン等のエマルジョン、種々の型の湿潤剤、滅菌溶液等を含む。かかる担体を含有する組成物は、よく知られた慣用の方法により調製され得る。これらの医薬組成物は、それの必要な患者に、適切な投与量で投与され得る。適切な組成物の投与は、例えば、経口、静脈内、腹膜内、皮下、筋内、局所または皮内投与等の異なった方法により行なわれ得る。投与量摂生は、担当の医師および臨床的因子によって決定されるであろう。医学分野においてよく知られているように、いずれのある患者に対する投与量も、患者のサイズ、体表面積、年齢、投与対象の特定の化合物、性別、投与時間および投与経路、一般的な健康、ならびに同時に投与される他の薬物を含む多くの因子に依存する。一般的に、医薬組成物の規則的な投与のような摂生は、1日当たり1μg〜10mg単位の範囲であるべきである。摂生が連続注入である場合、それぞれ、体重1キログラム当たり、1分当たり、それはまた1μg〜10mg単位の範囲であるべきである。経過は、周期的な評価によりモニターされ得る。投与量は変化するであろうが、DNAの静脈内投与に対する好ましい投与量は、DNA分子の約106 〜1012コピーである。本発明の組成物は、局部にまたは全身に投与され得る。投与は一般的に、非経口的、例えば静脈内であろう;DNAはまた、例えば、内部もしくは外部ターゲット部位へバイオリスティック(biolistic)送達によりまたは動脈内のある部位にカテーテルにより、直接ターゲット部位に投与され得る。
【0042】
本発明の種々のポリヌクレオチドおよびベクターが、単独で、または標準ベクターおよび/または遺伝子送達系を使用するあらゆる組み合わせで、ならびに任意に薬学的に許容できる担体または賦形剤と一緒に投与されることが、本発明により認識される。投与に続き、前記ポリヌクレオチドまたはベクターは、対象物のゲノムへ安定に統合されてもよい。一方、ある細胞または組織に特異的であり、該細胞または組織で存続するウイルスベクターが使用され得る。適切な医薬担体および賦形剤は、前記で述べたように、当該分野でよく知られている。本発明により調製される医薬組成物は、本発明の前記同定された核酸分子の望まない(過剰)発現に関連する異なる種類の疾患の予防または治療または遅延に使用され得る。好ましい態様において、医薬組成物は、例えば、実施例5に記載のように、調節、好ましくは多量の発現を低減できるアンチセンスオリゴデオキシヌクレオチドを含有する。
【0043】
さらに、本発明のポリヌクレオチドまたはベクターを含有する本発明の医薬組成物を、遺伝子治療で使用することができる。適切な遺伝子送達系は、リポソーム、レセプター媒介送達系、ネイキッド(naked)DNAおよびヘルペスウイルス、レトロウイルス、アデノウイルス、アデノ随伴ウイルス等の特にウイルスベクターを含み得る。例えば、イクスビボまたはインビボ技術による細胞へのワクチン接種のために治療的遺伝子を導入することに基づく遺伝子治療は、遺伝子導入の最も重要な適用の1つである。インビトロまたはインビボ遺伝子治療のための適切なベクター、方法または遺伝子送達系は、文献に記載されており、当業者に知られている;例えば、Giordano、Nature Medicine 2(1996)、534〜539;Schaper、Circ.Res.79 81996)、911〜919;Anderson、Science 256(1992)、808〜813;Isner、Lancet 348(1996)、370〜374;Muhlhauser、Circ.Res.77(1995)、1077〜1086;Onodera、Blood 91(1998)、30〜36;Verzeletti、Hum.Gene Ther.9(1998)、2243〜2251;Verma、Nature 389(1997)、239〜242;Anderson、Nature 392(Supp.1998)、25〜30;Wang、Gene Therapy 4(1997)、343〜400;Wang、Nature Medicine 2(1996)、714〜716;WO94/29469;WO97/00957;US5,580,859;US5,589,466;US4,394,448またはSchaper、Current Opinion in Biotechnology 7(1996),635〜640および本明細書に引用された参照文献を参照のこと。本発明の核酸分子およびベクターは、細胞への直接の導入またはリポソームまたはウイルスベクター(例えば、アデノウイルス、レトロウイルス)を介する導入に対して設計され得る。さらに、バキュロウイルス系が、本発明の核酸分子に対する真核生物発現系として使用できる。遺伝子治療のための体の特異的部位への核酸の送達はまた、Williams(Proc.Natl.Acad.Sci.USA88(1991)、2726〜2729)により記載されたもの等のバイオリスティック送達系を使用して達成され得る。
【0044】
組換えDNAで細胞をトランスフェクトする標準的な方法は、分子生物学の分野の当業者によく知られている。例えば、WO94/29469を参照のこと。遺伝子治療は、本発明の組換えDNA分子またはベクターを患者に直接投与することにより、または本発明のポリヌクレオチドまたはベクターで細胞をイクスビボでトランスフェクトし、トランスフェクトされた細胞を患者に注入することにより、行ない得る。さらに、生殖系の細胞への遺伝子導入に関する研究は、生殖生物学で最も速い成長分野の1つである。イクスビボまたはインビボ技術による細胞への治療的遺伝子の導入に基づく遺伝子治療は、遺伝子導入の最も重要な適用の1つである。インビトロまたはインビボでの遺伝子治療のための適切なベクターおよび方法は、文献に記載されており、当業者に知られている;例えば、WO94/29469、WO97/00957またはSchaper(Current Opinion in Biotechnology 7(1996)、635〜640)および上記に引用された参照文献を参照のこと。本発明の医薬組成物に含有されるポリヌクレオチドおよびベクターは、細胞への直接の導入、またはリポソームもしくは該組換えDNA分子を含むウイルスベクター(例えば、アデノウイルス、レトロウイルス)を介する導入のために設計され得る。好ましくは、該細胞は、生殖系細胞、胚性細胞、幹細胞または卵細胞もしくはそれら由来の細胞である。胚性細胞は、例えば、Nagy、Proc.Natl.Acad.Sci.USA90(1993)8424〜8428等に記載のような胚性幹細胞であり得る。
【0045】
本発明の導入されたポリヌクレオチドおよびベクターは、該細胞への導入後本発明の(ポリ)ペプチドを発現し、好ましくは該細胞のライフタイムの間この状態のままであることが理解されるべきである。例えば、適切な調節配列の制御下に安定にポリヌクレオチドを発現する細胞株は、当業者によく知られた方法により作製されてもよい。ウイルスの複製起点を含む発現ベクターを使用するよりむしろ、宿主細胞は、本発明のポリヌクレオチドまたはベクターおよび同一のベクターもしくは異なるベクターのどちらかにある選択可能なマーカーで形質転換され得る。外来のDNAの導入に続き、作製した細胞を、強化培地で1〜2日間、成長させてよく、次いで選択培地に変える。組換えプラスミドの選択可能なマーカーは、選択に耐性を与え、プラスミドを安定にそれらの染色体中に統合化しており、成長して順番に細胞株にクローニングされ、拡張され得るフォーカスを形成する細胞の選択を可能にする。かかる作製した細胞株の、スクリーニング方法、または、下記のような本発明のポリペプチドのインヒビターの同定方法に特に有用である。
【0046】
限定されないが、それぞれtk細胞、hgprt細胞またはaprt細胞のヘルペスシンプレックスウイルスチミジンキナーゼ(Wigler、Cell 11(1977)、223)、ヒポキサンチン−グアニンホスホリボシルトランスフェラーゼ(Szybalska、Proc.Natl.Acad.Sci.USA 48(1962)、2026)、アデニンホスホリボシルトランスフェラーゼ(Lowy、Cell 22(1980)、817)を含む多数の選択系が使用され得る。代謝拮抗物質耐性がまた、メトトレキサートに耐性を与えるdhfr(Wigler、Proc.Natl.Acad.Sci.USA 77(1980)、3567;O’Hare、Proc.Natl.Acad.Sci.USA 78(1981)、1527)、ミコフェノール酸に耐性を与えるgpt(Mulligan、Proc.Natl.Acad.Sci.USA 78(1981)、2072)、アミノグリコシドG−418に耐性を与えるneo(Colberre−Garapin、J.Mol.Biol.150(1981)、1)、ハイグロマイシンに耐性を与えるhygro(Santerre、Gene 30(1984)、147)、ゼオシン〔Zeocin(登録商標)〕に耐性を与えるShble(Mulsant、Somat.Cell.Mol.Genet.14(1988)、243〜252またはピューロマイシン(pat、ピューロマイシンN−アセチルトランスフェラーゼ)に対する選択の基礎として使用され得る。さらなる選択可能な遺伝子が記載されており、例えば、細胞がトリプトファンの代わりにインドールを利用できるようにするtrpB;細胞がヒスチジンの代わりにヒスチノールを利用できるようにするhisD(Hartman、Proc.Natl.Acad.Sci.USA 85(1988)、8047);およびオルニチンデカルボキシラーゼインヒビター、2−(ジフルオロメチル)−DL−オルニチン、DFMO(McConlogue、1987、In: Current Communications in Molecular Biology、Cold Spring Harbor Laboratory 編)に耐性を与えるODC(オルニチンデカルボキシラーゼ)がある。イクスビボ遺伝子治療の使用対象の細胞は、当業者によく知られている。例えば、かかる細胞は、例えば血液にもしくは組織に存在する癌細胞または好ましくは対応する幹細胞を含む。
【0047】
さらに、本発明は、本発明の核酸分子、本発明のベクター、本発明のポリペプチドおよび/または本発明の抗体を含有してなる診断用組成物に関する。
【0048】
本発明の診断用組成物は、本発明の核酸分子の望まない発現または過剰発現に関連する疾患の開始または進行を検出するのに有用である。本明細書において前記に指摘したように、かかる疾患は、細胞増殖の増加または進行と相互関係があるか、またはそれらにより引き起こされる。したがって、本発明の診断用組成物は、癌の開始または疾患状態を評価するために使用され得る。このように腫瘍活性に対する初期のクリチリウム(criterium)を有しており、適切なカウンター測定を直ちに適用し得る。かかる即時行動は、もちろん、患者の予後を有意に改良するであろう。これらの考慮は、転移腫瘍および反回性腫瘍の診断に等しく適用される。
【0049】
一方、腫瘍の全ての型が本発明の核酸分子の望まない発現および過剰発現によって特徴づけられ得るというわけではない。他方、該(過剰)発現は、腫瘍発達の初期の段階のようなある段階のみで生じるかもしれない。それゆえに、本発明の診断用組成物はまた、すなわち他方で腫瘍の分類もしくは腫瘍の発達状態の分類のための手段として使用され得る。当然、腫瘍に対する本明細書に記載された本発明の組成物の適用またはほとんどの適用もまた、本発明の核酸分子の望まない(過剰)発現それらと相互関係があるまたはそれにより生じる他の疾患に応用する。
【0050】
さらに、本明細書を通して引用されるような疾患はまた、本発明のポリペプチドの機能不全により引き起こされ得るであろう。該疾患は、例えば、本発明のポリペプチドの遺伝子量(dosis)増加または低減、細胞もしくは組織の対応する野生型ポリペプチドと比較した一次アミノ酸配列での修飾または該ポリペプチドの活性の調節の損失等による該ポリペプチドの活性の増加または低減と相互関係があり得、またはそれらにより生じ得るであろう。該疾患はさらに、細胞周期進行または細胞発達の間のポリペプチドの間違った発現によって引き起こされるかもしれない。例えば、イオンまたは二次メッセンジャーまたは調節タンパク質等の細胞内または細胞外化合物への変異した結合部位は、それが該ポリペプチドの活性を調節する該化合物に対する結合特性を変える場合、本発明のポリペプチドの機能不全を生じさせ得るかもしれない。機能不全はまた、不完全な修飾部位、例えば、リン酸化部位またはグリコシル化部位により引き起こされ得るであろう。それはまた、例えば、heag1がheagh2の代わりに発現する場合、または逆の場合、間違ったスプライシング事象、それゆえ切形もしくは伸長ポリペプチドの発現により、引き起こされるかもしれない。
【0051】
したがって、さらなる態様において、前記診断用組成物はまた、本発明のポリペプチドの機能不全を検出するのに使用され得る。
【0052】
さらなる態様において、本発明は、本発明の核酸分子の発現のインヒビターまたは本発明の(ポリ)ペプチドの機能不全のインヒビターもしくは改変剤またはheagをコードする核酸分子またはheag活性を有するポリペプチドを、該疾患の影響を受けた哺乳類または該疾患の影響を受けやすいと推測される哺乳類に導入する工程を含む、本発明のポリペプチドの機能不全により引き起こされる疾患を予防または治療する方法に関する。heagをコードする本発明の核酸分子の細胞または対象物への導入方法、すなわち遺伝子治療は、本発明の核酸分子の発現のインヒビターの同定方法と同様に本明細書に記載されている。さらに本発明のポリペプチドの機能不全のインヒビターまたは改変剤は、当業者に既知の本発明のポリペプチドのインヒビターインヒビターの同定方法により同定され得る(下記を参照のこと)。例えば、本発明のポリペプチドの機能不全を生じるある遺伝子変化は、タンパク質高次構造状態に変化をもたらす。変異タンパク質は、イオン輸送の促進をほとんど不可能にする三次構造を有し得る。変異タンパク質の正常なまたは調節された高次構造を回復することは、これらの分子欠陥を正すために最も明快で特有の意味を持つ。薬理学操作は、このようにタンパク質の野生型構造の復活を助けるかもしれない。このように、本発明のポリヌクレオチドおよびコードされたタンパク質はまた、該機能不全を示す本発明のポリペプチドの誘導体の野性型機能を活性化できる分子を設計および/または同定するために使用され得る。
【0053】
それの必要な患者の治療のための投与量および投与経路は、本発明の医薬組成物に関連して本発明の前記ですでに記載した。本発明の方法を使用して治療され得る疾患は、細胞増殖に関係するいずれの疾患をも含む。このカテゴリーに分類される好ましい疾患は、癌(乳癌、神経芽腫等)等の腫瘍性疾患、乾癬、および変性疾患、とりわけアルツハイマー病、多発性硬化症、筋萎縮性側索硬化症、パーキンソン病等の神経系の疾患である。
【0054】
好ましくは、該核酸分子の発現または過剰発現の該インヒビターは、本発明のイオンチャンネルをコードする核酸分子またはその断片にハイブリダイズする本発明の核酸分子である。例えば、この核酸分子は、アンチセンスオリゴデソキシヌクレオチド(oligodesoxynucleotido)(ODN)であり得る。発明者は、アンチセンスODN治療が、いくつかの腫瘍細胞、例えばEFM細胞、SHSY−5Y細胞およびHeLa細胞のDNA合成を有意に低減することを示し得た(実施例5)。したがって、好ましい態様において、核酸分子はアンチセンスODNを含有する。
【0055】
さらに好ましい態様において、該ポリペプチド機能のインヒビターは、本発明の抗体または薬物である。該薬物は、ヒスタミンレセプターH1インヒビターであり得る。好ましくは、該薬物は、活性heagを阻害し、例えば使用依存的におそらくオープンチャンネルブロッカーとして作用し、好ましくは該薬物はアステミゾール(astemizole)またはテルフェナジン(terfenazine)である。さらに、適切な薬物が、本発明の教示に基づいて当業者により同定または設計され得る。好ましくは、薬物は、heagチャンネルにmM範囲、さらに好ましくはnM範囲またはそれより低い範囲で親和性を有するであろう。好ましくは、該薬物は、他のチャンネル、例えば心臓チャンネルに影響しない。
【0056】
本発明のさらに好ましい態様において、該方法は、導入工程の前に、
(a)前記疾患により冒された哺乳動物から細胞を得る工程ならびに前記導入工程の後に前記疾患により冒された哺乳動物から細胞を得る工程、ここで、前記導入は前記細胞に行われる、
(b)前記細胞を前記哺乳動物または同種の哺乳動物に再導入する工程、
をさらに含む。
【0057】
本発明の本態様は、治療継続時間を大きく低減し、効力を増加し、副作用を低減する(消去さえする)遺伝子治療目的のために特に有用である。さらに、本発明の方法の本態様はまた、これに関連してまたは通常の医学治療と組み合わせて使用し得る。該哺乳類からの除去および該哺乳類への再導入は標準手順によって行なうことができる。
【0058】
好ましくは、前記参照細胞は、生殖細胞、胚性細胞もしくは卵細胞またはこれらの細胞いずれか由来の細胞である。
【0059】
本発明は、さらに、本発明の核酸分子の望まれていない発現または過剰発現によって引き起こされる疾患の治療用薬物の設計方法であって、
(a)特異的かつ効力の高い薬物を同定する工程;
(b)部位特異的突然変異誘発およびキメラタンパク質研究による、前記薬物の結合部位を同定する工程;
(c)(ポリ)ペプチドの結合部位および前記薬物の構造の両方を分子モデリングする工程;ならびに
(d)(ポリ)ペプチドに対するその結合特異性を改良するために薬物を改変する工程、
を含む、治療用薬物の設計方法に関する。
【0060】
本明細書で使用される用語「特異的かつ効力の高い薬物」は、heag機能を高い効力でかつ特異的にブロックする薬物に関する。
【0061】
本発明の方法の様々な段階で使用される全技術は通常のものであるかまたは、さらなる努力なしの通常の技術により当業者によりもたらされ得る。したがって、本発明のイオンチャンネルの特徴を同定する本明細書に基づく生物学的アッセイが、薬物の特異性または効能を評価するために使用され、そこで、イオンチャンネルの1以上の本発明の活性の減少が該特異性または効能をモニターするのに使用され得る。工程(b)および(d)は、例えば、K.L.Choi、C.Mossman、J.Aube & G.Yellen.The International Quaternary Ammonium Receptor Site of Shaker Potassium Channels.Neuron 10、533〜541(1993)、C.−C.Shieh & G.E.Kirsch:Mutational Analysis of Ion Conduction and Drug Binding Sites in the Inner Mouth of Voltage−Gated K+ −Channels.Biophys.J.67、2316〜2325(1994)、またはC.Miller:The Charybdotoxin Family of K+ −Channel−Blocking Peptide.Neuron 15、5〜10(1995)に記載される通常のプロトコルにしたがって行なうことができる。
【0062】
例えば、部位特異的突然変異誘発およびキメラタンパク質研究による該薬物の結合部位の同定は、薬物親和性に影響を及ぼす(ポリ)ペプチド一次配列での修飾により達成され得る;これは、通常、薬物に対する結合ポケットを正確にマッピングすることを可能にする。
【0063】
工程(c)に関しては、次のプロトコルが認識され得る;一度、薬物に対するエフェクター部位がマップされると、薬物の正確な三次元構造が既知である場合には、薬物の異なる部分と相互作用する正確な残基が、突然変異誘発研究〔工程(b)〕から得られる情報と結合部位の構造のコンピューターシミュレーション(カリウムチャンネルが最近当該分野において結晶化されているので、これは現在さらなる努力なしに当業者によってなされ得る)との組み合わせによって同定され得る(そうでないなら、それはコンピューターシミュレーションによって予測され得る)。該薬物それ自身がペプチドである場合、それはまた、どの残基がheag分子の他の残基と相互作用するか決定するために変異され得る。
【0064】
最終的に工程(d)で薬物は修飾され、その結合親和性またはその効能および特異性を改良し得る。例えば、もしheagの特定の残基と薬物分子のある領域の間に静電的相互作用があるなら、その領域の全体的な電荷を修飾し、その特定の相互作用を増加できる;さらに、もしそれらの相互作用が他のチャンネルタンパク質で変換されないheagの領域で生じるなら、この相互作用の改良は、他の結合因子が弱められる一方で、薬物の特異性を改良すると考えられている。
【0065】
結合部位の同定は、コンピュータープログラムによって補佐され得る。したがって、適切なコンピュータープログラムが、相補的な構造モチーフに対するコンピューター補佐探究によって、本発明の推定上のインヒビターおよびポリペプチドの相互作用部位の同定に使用され得る(Fassina、Immunomethods 5(1994)、114〜120)。さらに、タンパク質およびペプチドのコンピューター補佐設計のための適切なコンピューターシステムが、例えば、Berry、Biochem.Soc.Trans.22(1994)、1033〜1036;Wodak、Ann.N.Y.Acad.Sci.501(1987)、1〜13;Pabo、Biochemistry 25(1986)、5987〜5991の先行技術で記載されている。薬物の修飾は、例えば、ぺプチド擬似体により製造され、他のインヒビターはまた、次の化学修飾および得られた化合物を試験することを介するぺプチド擬似体コンビナトリアルライブラリーの合成により同定し得る。ぺプチド擬似体コンビナトリアルライブラリーの生成および使用方法は、例えば、Ostresh,Methods in Enzymology 267(1996)、220〜234およびDorner、Bioorg.Med.Chem.4(1996)、709〜715の先行技術で記載されている。さらに、本発明のポリペプチドのインヒビターの三次元構造および/または結晶構造は、例えば、本発明の(ポリ)ペプチドと組み合わせて(Rose、Biochemistry 35(1996)、12933〜12944;Rutenber、Bioorg.Med.Chem.4(1996)、1545〜1558)、ペプチド擬似体インヒビターの設計に使用され得る。
【0066】
本発明にしたがって使用され得る特定のインヒビターを同定するための具体的な戦略が、添付した実施例で提供される。
【0067】
本発明はまた、
(a)アンチセンスオリゴヌクレオチドおよびリボザイムから選ばれる化合物を、翻訳の阻害または低減に対して試験する工程;または
(b)本発明の(ポリ)ペプチドをコードし、かつ好ましくはその転写因子応答エレメントを有する遺伝子のプロモーター領域に結合する化合物を、転写の阻害に対して試験する工程;または
(c)本発明の(ポリ)ペプチドの増殖活性をブロックすると推測されるペプチドまたは抗体を、前記ブロック活性に対して試験する工程、
を含む、本発明の核酸分子の発現のインヒビターまたは本発明の(ポリ)ペプチドの機能のインヒビターを同定する方法に関する。
【0068】
前記に関するもう1つの(b)に関しては、まずプロモーター領域を特徴づけ、その中に転写因子応答配列があることが有利であり得る。次いで、プロモーターを遺伝子的に操作して、それをリプレッサーに対する反応性を高くし、エンハンサーに対する反応性を低くすることが可能であろう。今度はもう1つの(c)を見ると、それはまず増殖障害の発生に影響を与える本発明のイオンチャンネルの1以上の部分にあることが有利である。1つの試験系で陽性である化合物は、インヒビターとして有用であることが明白である。
【0069】
ペプチド擬似体、ファージディスプレイおよびコンビナトリアルライブラリー技術は当該分野でよく知られており、それは当業者によりさらなる努力なしに前記本明細書に関連する基本的方法によって同定される薬物またはインヒビターの改良に適用できる。
【0070】
さらなる態様において、本発明は、本発明の核酸の発現または本発明の(ポリ)ペプチドにインヒビターを適用する工程を含む、細胞増殖を阻害する方法に関する。本発明の方法は、インビトロ、イクスビボ、または適用が対象物に対する場合にはインビボで行ない得る。
【0071】
本発明はまた、本発明の核酸分子の発現を評価する工程、または本発明の(ポリ)ペプチドの定量的な存在を評価する工程を含む、癌および/または神経変性疾患および/または乾癬を予知する方法に関する。好ましい態様において、該癌は、乳癌または神経芽腫、さらに好ましい態様において、該癌は、胸腺癌、胸癌管型または子宮頸癌である。さらなる態様において、該神経変性疾患は、アルツハイマー病、パーキンソン病、筋萎縮性側索硬化症または多発性硬化症である。
【0072】
本発明の方法は、インビトロ、インビボまたはイクスビボで行ない得る。本発明の方法を行なうための適切なプロトコルは当該分野でよく知られており、インビトロ技術に関しては、mRNAのレベルの評価のためのノーザンブロット、または、例えば本発明の(ポリ)ペプチドを特異的に認識する抗体を用いる顕微鏡技術による組織の解析を含む。1以上のこれらの技術は、同様にまたはさらなる(通常の)技術を組み合わせて前記引用された評価に使用される技術に基づいたPCRと組み合わされ得る。
【0073】
本発明の前記方法のさらなる好ましい態様において、該哺乳類は、ヒト、ラットまたはマウスである。
【0074】
本発明はさらに、遺伝子治療における本発明の核酸分子の使用に関する。本明細書前記で指摘したように、遺伝子治療は細胞増殖を阻害するように設計され、それにより癌または乾癬等のそれにより影響されるいずれの疾患をも特定の方法で治療し得る。本発明は特に、かかる遺伝子治療プロトコルを行なう2つの非依存系を認識する:
(a)ヒトeagに特異的な薬物を得るための、H1アンタゴニスト(好ましくはアステミゾール)の化学工学を共に有するチャンネルの変異誘発;
(b)診断および/または予知方法を設計するための、癌細胞でのeagの発現レベルの定量的および定性的解析。これはまた、特定の腫瘍に対する遺伝子治療の設計を可能にするであろう。
【0075】
例えば、核酸分子は、レトロウイルスベクター(Naldiniら、Science 272(1996)、263〜267;Mulligan、Science 260(1993)、926〜932)または適切なベクターを用いて細胞にインビボで導入され得る。同様に本発明により、患者由来の細胞が単離され、標準組織培養技術を用いてインビトロで修飾され、そして患者に再導入され得る。かかる方法は、本明細書前記に関連する遺伝子治療または遺伝子転写方法を含む。
【0076】
最後に、本発明は、本発明の(ポリ)ペプチドをコードする核酸分子に特異的にハイブリダイズする核酸分子、本発明のベクター、本発明のポリペプチドおよび/または本発明の抗体を含有してなるキットに関する。
【0077】
本発明のキットは特に、前記に関連する多数の診断方法に使用され得る。本発明のキットは、形質転換された宿主細胞およびトランスジェニック植物細胞、植物組織または植物の発生に適切な選択培地に対する選択マーカーおよび成分等の材料をさらに含む。さらにまた、キットは、本発明の組換え遺伝子またはベクターに存在し得るレポーター遺伝子のための緩衝液および基質を含み得る。本発明のキットは、本発明の方法を行なうために有利に使用することができ、特に、本明細書に言及する様々な適用、例えば診断分野または研究手段の際に使用し得る。本発明のキットの部分は、バイアルに個々に、または容器もしくは多容器単位に組み合わせてパッケージされ得る。キットの製造は、好ましくは、当業者に既知の標準手段に従う。
【0078】
いくつかの文書が、本明細書の本文を通して引用されている。本明細書で引用されている各文書は(いずれの製造業者の仕様書、指示等をも含む)、これによって本明細書に参照により取り込まれる;しかしながら、引用されたいずれの文書も実際に本発明に関して先行技術であると認めているというわけではない。
【実施例】
【0079】
実施例により本発明を説明する。
【0080】
実施例1:K+ イオンチャンネルのクローニング
標準手法に従ってMCF−7細胞から得た全RNAよりmRNAを精製した。次いで、スーパースクリプト(Superscript)II逆転写酵素を用い逆転写によりcDNAを調製した;このcDNAを、高度に保存されたeag配列に適合するように設計された縮重オリゴヌクレオチドを使用するPCR増幅のための鋳型として用いた。増幅後、ラットeag由来のSacII/SacII断片を、結果のサザンブロット解析のためのプローブとして用いた。陽性ハイブリダイゼーションを示すそれらのバンドを、次いで、pGEM−Tベクター(Promega社)にクローニングし、配列決定した。それらの全てがHERGに対応する配列を与えた。
【0081】
次いで、ラット、マウスおよびウシのeagを考慮し、HERG cDNA増幅を避けるように作製される特異的オリゴヌクレオチドを設計した。我々は、種々のeagクローン間で高い相同性を有するが、HERG配列に対し最も効果的に相違している配列を探した。
【0082】
オリゴヌクレオチドの配列は以下であった:
5'-CAGAA(T,C)AA(T,C)GTGGC(A,C,T,G,)TGGCT
5'-TCACT(G,A)AAGATCTATA(A,G)TC
【0083】
PCR増幅後、期待されたサイズのバンドをpGEMTにクローニングし、配列決定した。得られた配列はラットeag(ヌクレオチド942〜1108)に高度に相同性を示した。
【0084】
このバンドを標識し、乳腺cDNAライブラリーをスクリーニングするためのプローブとして用いた。2×106 ファージのスクリーニング後、陽性クローンを全く見出せなかった。
【0085】
次いで、我々は、特異的オリゴヌクレオチドを用い、(Clontech社から購入した全RNAより得た)ヒト心臓およびヒト脳由来のcDNAをPCRを使用して解析した。脳由来の2つのPCR産物を配列決定し、該配列はeagの2つの選択的にスプライシングされた変異体(variants)に対応した。ヒト脳由来の全長分子のクローニングの可能性をさらに試験するために、我々は、ヒトcDNAライブラリーのPCR解析を行い、この結果をヒト乳腺ライブラリーにおける同じ実験と比較した(両方ともClontech社由来)。脳ライブラリーのみが陽性の結果を与えた。
【0086】
続いて、増幅された断片を用いてヒト脳ライブラリーのスクリーニングを行い(2回、106 ファージ)、pBSKベクターにクローニングされたいくつかのクローンを見出し、配列決定した。それらの全ては分子の中央部分に相当したが、5’および3’末端が欠失していた。これらの陽性クローンの内で最も長鎖のものを用いてプローブを調製し、再度ライブラリーのスクリーニングを行った(再び2回、2×106 クローン)。
【0087】
この場合に得られた配列は、コード配列の一部(約400bpの5’が開始コドンまで欠失していた)と長鎖の3’非翻訳配列に相当した。分子の5’末端に近い断片は全ての場合にEcoRI部位で始まったので、かかる部位はheag配列に実際に存在し、配列決定のためのベクターへの断片のサブクローニングにおいて消失したということが推測された。
【0088】
全長配列を得るため、我々は、5’末端に近い断片を保持するそれらのファージをプールし、前記EcoRI部位に対する配列3’とラムダgt10由来の配列をPCRのためのプライマーとして用い、PCR増幅によりそれらを解析した。プールの連続した分画の後、コード配列の5’末端を保持する2つのファージを得、それらの1つは、5’非翻訳領域の一部を含んでいた。
【0089】
我々は完全配列を知得した後、それらの1つは3’UTRとほとんどのコード配列を含んでおり、他方は5’末端を含んでいる、2つのファージから始めて、全クローンを組み立てた。最初の断片をSphI/HindIII消化によりファージから抽出し、pBKS−にサブクローニングしてpBKSheag1を作製した。このワズ(was)に、1.2kbpのSphI−SphI断片もクローンから除去し、その後、それを再度導入することが必要であった。5’末端を含む断片をHindIII/MunI消化により抽出した。この断片をpBKSheag1のHindIII/MunI消化物とライゲートさせた。この手法を用いてのみ、我々は単一プラスミドにおいて全長クローンを得ることができた。次いで、我々はSphI−SphI断片を再度導入する必要があった。というのは、我々はSphI部位の1つを欠失させたからであった。続いて、EagI/NotI断片をpCDNA3ベクターのNotI部位にサブクローニングし、汚染したファージ配列を除去し、チャンネルの機能的発現に好適なベクターを得た。最終的に得られた配列を配列番号:1および配列番号:2として配列表に記載する。
【0090】
実施例2:ヒトeagの作用を特異的にブロック(bloc)するインヒビターの同定
eagファミリーの他の1つのメンバーであるHERG11-16 は、長鎖QT症候群(LQT)のファミリー型に関連付けられている。これにより、それらのLQT型不整脈を誘導する能力に基づいてHERGのいくつかのブロッカーの同定が可能となっている。このように、アステミゾール(astemizole)およびテルフェナジン(terfenadine)等の特定のヒスタミンH1レセプターブロッカー、ならびにIII型不整脈薬物〔ドフェチリド(dofetilide)、E−4031)〕は、HERG15,17 の強力で特異的なブロッカーである。しかしながら、eagチャンネルについての特異的なブロッカーは未だ記載されていない。HERGとeagチャンネル間の配列類似性のため、reagに対する両群の薬物を本発明に従って試験した。H1ブロッカーもreagに影響し、一方、チャンネルはIII型抗不整脈剤(ドフェチリド)に対しむしろ無感受性である。このことは、選択的にeag型チャンネルをブロックし、(MCF7細胞にも存在する)HERGチャンネルの可能な影響を排除するための有効な道具を提供する。単一の推定ヒトeagチャンネルに対する、これらの薬物の1つ(アステミゾール 5μM)の影響をFig.6に示す。
【0091】
いくつかのreagおよび他のカリウムチャンネルブロッカーがMCF7細胞の成長を阻害することができるか否かをさらに試験した。「陽性」対象として、ATP感受性カリウムチャンネルのブロッカーであるグリベンクルアミド(glibenclamide)も含めた。というのは、この細胞株の増殖を阻害することが記載されているためである2 。DNA合成の速度を測定するために、細胞を、約105 細胞/mlの密度で、かつ成長因子の非存在下で、96穴マイクロタイタープレートにて培養した。24時間の飢餓の後、BrdUの存在下に10%FCSの添加により細胞を刺激した。新たに合成されたDNAに取り込まれたBrdUの量を市販の抗体(Boehringer Mannheim)を用いて測定した。試験された薬物は、刺激と同時に、または刺激の12時間前に添加した。異なるヒト細胞株であるHEK293において、10μM アステミゾールまたは100μM グリベンクルアミドの添加は有意にはDNA合成を低下させなかったが、テルフェナジン(10μM)は強力な阻害を示した。この理由について、アステミゾール(およびその密接に関連するアナログLY91241)の影響のみが考慮され、テルフェナジンにより生成されるもの(MCF7細胞は、対照細胞よりもテルフェナジンによる成長阻害に対し有意により感受性であるが)は排除された。MCF7細胞において、5μM アステミゾールは40%だけDNA合成を低下させ、一方、同濃度のHERG特異的ブロッカーであるドフェチリドは有意な影響を示さなかった。10倍高い濃度(50μM)の他のカリウムチャンネルブロッカー〔キニジン(quinidine)またはグリベンクルアミド(glibenclamide)〕が類似の影響を誘導するのに必要とされる。MCF7細胞におけるDNA合成に対するアステミゾール効果についてのドーズレスポンスカーブをFigure 8に示す。最大効果の半分は10μM アステミゾールについて得られた。
【0092】
MCF7細胞における増殖阻害の基礎となる機構を明らかにする試みにおいて、超生体核染料ヘキスト(Hoechst)33342を用い、5μM アステミゾールで処理された細胞の核形態を調べた。24時間の処理後、ほとんどの細胞が、アポトーシス細胞死の典型的な特徴である、核濃縮および核断片化を示した(Fig.9)。
【0093】
結論として、reagチャンネルのヒト対応物はヒト癌細胞に存在しており、それらは、いくつかの異なる細胞型において悪性変態を誘導する能力を有する。
【0094】
実施例3:異なるヒト組織におけるheagの発現
異なるヒト組織由来の全RNA500ng(または脊髄のポリA+ RNA5ng)を逆転写し、配列 5'-CGCATGAACTACCTGAAGACG(フォワード)および5'-TCTGTGGATGGGGCGATGTTC(リバース)の1組のオリゴヌクレオチドを用いて増幅した。増幅したDNAを特異的なヒトeagプローブ(チャンネルのコア由来の1.5Kb EcoRI断片)を用いてサザンブロットにより解析した。試験したRNAの内、脳全RNAのみが陽性シグナルを与えた。脊髄、副腎、骨格筋、心臓、気管、肝臓、腎臓および乳腺由来のRNAは陰性であった。RNAの正確性をトランスフェリン増幅を用いて調べた。同じアプローチを用いて、いくつかの腫瘍性ヒト細胞株におけるheagの発現を調べた:MCF−7(胸腺癌)BT−474〔胸管癌(breast ductal carcinoma)、充実性腫瘍由来〕、EFM−19〔胸癌、管型胸膜液(pleural fluid)由来〕、COLO−824(胸癌、胸膜液由来)、SHSY5Y(神経芽腫)。正常組織と比較して、試験した全ての癌細胞株はheag発現について陽性であることが見出された。さらに、異なるヒト組織および293細胞由来のRNAのRT−PCR産物のサザンブロットは、脳由来RNAにおいてのみ、heagAおよびBに相当する2つのバンドが増幅され得、同定され得るということを示す。トランスフェリンレセプター(TFR)シグナルを下部に示す(Fig.15A)。さらに、異なるヒト細胞株および乳房上皮細胞の一次培養物(Epith.cells)由来の全RNAのRT−PCR産物のサザンブロット解析。TRFシグナルを下部に示す。異なる細胞株由来のRNA(34)およびヒト組織由来の市販のRNA(Clontech社)をシングル−チューブRT−PCR(35)に供した。脊髄を除き、全RNAを用いた。その際、ポリ(A)+ RNAを用いた(プライマー配列は以下:フォワード:5'-CGCATGAACTACTGAAGACG、およびリバース:5'-TCTGTGGATGGGGCGATGTTC。5'-TCAGCCCAGCAGAAGCATTATおよびリバース:5'-CTGGCAGCGTGTGAGAGC をRNAおよびPCR性能を制御するために用いた。)。TFRの特異的プライマーをRNAおよびPCR性能を制御するために用いた。これらのODNは、エクソン11に始まり、エクソン19まで広がっている(37)公表されたTFR配列(36)に従って設計した。これは、2つのheagスプライス断片および逆転写酵素の非存在下での対照の増幅と共に、ゲノムDNA汚染による偽陽性を排除する。50μl(heag)または15μl(TFR)のPCR反応物を2%アガロースゲルで解析した。DNAをメンブレンに転写し、高度ストリンジェンシーにて、980bp heag断片および脳RNAから増幅されたTFR断片からなる〔32P〕−dCTP標識したランダムにプライミングするプローブと連続的にハイブリダイズさせた。
【0095】
実施例4:インビボでのheagの発現
heagの発現はインビボで腫瘍細胞にとって有利であるか否かを決定するため、本発明者らは、雌性シッド(scid)(極度の複合免疫不全、33)マウスの脇腹へのチャンネルを発現するCHO細胞(CHOhEAG細胞)の皮下移植を行った。CHOKv細胞を対照として用いた。従って、100μlのPBS中に懸濁した2×106 CHOhEAG細胞またはCHO−Kv1.4細胞を、Bomholtgard,Ry,Denmarkから得た6〜8週齢の雌性Fox Chase scidマウス(C.B−17/Icr sicd/scid)の脇腹上で皮下に移植した。腫瘍の存在を、全てのマウスの触診により2日ごとに調べた。2または3週間後、動物を頸の脱臼により屠殺し、腫瘍を切開し、続くパラフィン包埋および染色のためにパラホルムアルデヒドにて固定した。CHOhEAG細胞の同定は、pTracerベクター(Invitrogen)にコードされるグリーン蛍光タンパク質由来の蛍光を生起する腫瘍のUVイルミネーションにより確立した。移植の1週間後、CHOhEAGを注入した全てのマウスが触診により検出可能な腫瘍を有したが、対照においては1mmを超える容量のものは観察されなかった。移植後2週間の間には、heag発現腫瘍は直径で5mmを超過するに至り、ほとんどの場合、皮膚を通して目に見える程に現れた(Fig.17A);マウスを2週間後(N=6)または3週間後(N=7)に屠殺した。使用した11匹の対照動物の内1匹のみが目に見える腫瘍を有さなかった;全13匹のCHOhEAGを注入した動物は腫瘍を示した。heag発現腫瘍の平均容量(Fig.17B,C)は、対照のそれより有意に大きく、特に移植後2週目が大きかった(Fig.17B)。肉眼観察から、腫瘍はもろく、出血性であるようであった;CHOhEAG腫瘍は対照よりも黒ずんでおり、2週目に全てのCHOhEAGを注入したマウスにおいて皮膚に癒着した(Fig.17D,E)。7匹のマウスの内6匹が3週目に同様な特徴を示した。対照的に、腫瘍は、2週間後、全ての対照マウスにおいて皮膚から容易に切開でき、3週目に6匹のマウスの内5匹においてであった。腫瘍の下の組織は全ての場合に影響を受けていないようであった。組織学により確認し(Fig.17H,I,矢頭)、黒ずんだ色は多量の腫瘍内の壊死によるためであった(Fig.17F,G,矢印)。このことは、CHOhEAG腫瘍のより早い成長を示す。EAG発現腫瘍における生命維持範囲の厚さは、有意に対照におけるより小さかった(Fig.17J)。腫瘍の急速な成長により、CHOhEAG群における多量の腫瘍内壊死を説明することができる。このことはまた、移植2週間後における腫瘍の容積に見出される差異の増大を説明する。というのは、CHOhEAG腫瘍は、大量の壊死のために成長を止めるであろうからである。これらのデータは、heag腫瘍の発現はより早く成長し、CHOKv腫瘍よりもより攻撃的であることを強力に示唆する。
【0096】
実施例5:heagの阻害
ある腫瘍細胞におけるheagの発現は、それらの異常な成長の結果ではないが、このK+ チャンネルが、それらの増殖に必要であるということが想定される。それゆえ、アンチセンスオリゴデオキシヌクレオチド(ODN)によるheag発現の阻害は、これらの腫瘍細胞における増殖速度を減少させるはずである。そこで、heagの推定開始コドンに広がる19merのアンチセンスホスホロチオエート ODN(5'-CAGCCATGGTCATCCTCCC)を用いて増殖の阻害を試験した。センスODNおよびスクランブル(scrambled)配列(gtcggtaccagtaggaggg )を対照として用いた。Figure 16Aに示すデータは、EFM細胞におけるheag mRNA含量を減少させるアンチセンスODN処理の有効性を明確にする。アンチセンスODNでの処理によるSHSY−5Y細胞におけるheag媒介K+ 電流の減少をFig.16BおよびCに示す。
【0097】
アンチセンスODNでのheag発現腫瘍細胞株の処理は有意に増幅PCR産物の生成量を減少させた。EFM−19細胞を一夜、10μg/ml DAC30(レーン「C」)または10μg/ml DAC30(Eurogentec)プラス1μM アンチセンスODN(レーン「AS」)で処理し、全RNAを抽出し、heagまたはトランスフェリンレセプターを増幅するように設計されたODNを用いて、実施例3に記載の同条件下でアッセイした。Fig.16Aの矢印は増幅断片の期待されるサイズを示す。さらに、SHSY−5Y神経芽腫細胞におけるheag電流を詳細に解析するために、本発明者らは、細胞外Mg2+の存在下にeag(30)の活性化の電圧依存性を利用した。電流は−120mVから+60mVへ脱分極後に測定した(Fig.16B、灰色の線)。差し引きトレースの最初の部分(Fig.16B、黒線)はeag電流に相当する。該電流は、保持電位が非常に弱い場合(−120mV)、未だ活性化されていないが、保持電位が−60mVである場合、明確になる。19〜21msの平均電流を選択し、電流密度を決定した。アンチセンスODNにより処理したSHSY−5Y細胞における電流密度は、対照細胞と比較して有意に減少した〔電気生理学的測定は、(mM単位)140 NaCl、2.5 KCl、2 CaCl2、2 MgCl2、10 Hepes/NaOH pH7.2、10 グルコースを含む細胞外液を用いてパッチクランプ法のホールセルコンフィギュレーションにて標準プロトコルを使用して行った(Hamill,O.P.,Marty,A.,Neher,E.,Sakmann,B.,Sigworth,F.J.Pfluegers Arch- Eur.J.Physiol 391,85(1981))。ピペット液は、(mM単位)140 KCl、10 BAPTA、10 Hepes/KOH pH7.2であった。〕。細胞を一夜、1μMの蛍光標識ODNを含むアンチセンスODNで処理した。電流は、それらの核において蛍光を示す細胞において1〜3日後に測定した。Fig.16Cのバーは9個の細胞(対照)または25個の細胞(アンチセンス)についての平均±S.E.M.を示す。外部電流のみを解析において評価した。さらに、heagに対し向けられたアンチセンスODNによるヒト癌細胞(EFM−19、HeKaおよびSHSY−5Y)におけるDNA合成の阻害を調査した。DNA合成をODNの非存在下におけるBrdU取り込みと比較して表わす。蛍光標識アンチセンスODNを用いた細胞への取り込み条件を至適化した。細胞を105細胞/mlの密度にて96穴プレートに播いた。播いた1日後、細胞を培養培地で洗浄し、ODNを添加した(終濃度10μM)。ODNを予め無血清培地にて20μg/mlのトランスフェクションラゲネント(ragenent)DAC−30(Eurogentec)と共に混合しておき、20〜30分間、室温にてインキュベートした。次いで、混合物を培養培地に1:1希釈で添加し、一夜、細胞との接触を維持した。このインキュベーション後、細胞を洗浄し、2時間、BrdU(100μM)で標識した。取り込みはBoehringer Mannheim 社のキットを用いて検出し、非特異的バックグラウンド取り込みを差し引いた後、405nm(参照490nm)にてOD単位で測定した(Fig.16D)。バーは、典型的な実験における条件当たり8つのウェルについての平均±S.D.を示す。
【0098】
【表1】
【0099】
参考文献:
1. Moody,W.J.(1995)。イオンチャンネル特性の統合調節によって創られる初期発達の臨界期。Perspect Dev Neurobiol 2、309〜315。
2. Woodfork,K.A.、Wonderlin,W.F.、Peterson,V.A.、およびStrobl,J.S.(1995)。ATP感度の高いカリウムチャンネルの阻害は、組織培養でヒト乳癌細胞の可逆的細胞停止を生じる。J Cell Physiol 162、163〜71。
3. Lepple Wienhues,A.、Berweck,S.、Bohmig,M.、Leo,C.P.、Meyling,B.、Garbe,C.、およびWiederholt,M.(1996)。ヒトメラノーマ細胞増殖での、K+ チャンネルおよび細胞内カルシウムシグナル。J Membr Biol 151、149〜57。
4. Wonderlin,W.F.、Woodfork,K.A.、およびStrobl,J.S.(1995)。細胞周期を通じたMCF−7ヒト乳腫瘍細胞の進行の間の膜電位での変化。J Cell Physiol 165、177〜85。
5. Brueggemann,A.、Stuehmer,W.、およびPardo,L.A.(1997)。アフリカツメガエル卵母細胞で発現されるクローニングさせた遅延レクティファイヤーカリウムチャンネルの有糸分裂促進因子が媒介する抑制。Proc Nat Acad Sci USA 94、537〜542。
6. Mosmann,T.(1983)。細胞増殖および生存のための速い比色アッセイ:増殖および細胞毒アッセイへの適用。J.Immunol.Meth.65、55〜63。
7. Muir,D.、Varon,S.、およびManthorpe,M.(1990)。固定化したミクロ培養を使用するブロモデオキシウリジン取り込みに対する酵素結合免疫吸着剤アッセイ。Anal.Biochem.185、377〜82。
8. Magaud,J.P.、Sargent,I.、およびMason,D.Y.(1998)。ブロモデオキシウリジン取り込みの免疫酵素的測定によるヒト白血球増殖応答の検出。J.Immunol.Meth.106、95〜100。
9. Huong,P.L.、Kolk,A.H.、Eggelte,T.A.、Verstijnen,C.P.、Gilis,H.、およびHendriks,J.T.(1991)。5−ブロモ−デオキシウリジン取り込みを使用した抗原特異的リンパ球増殖の測定。放射性チミジン取り込みに代わる容易でかつ低コスト。J.Immunol.Meth.140、243〜8。
10. Ellwart,J.、およびDormer,P.(1985)。モノクローナルBrdUrd抗体でBrdUrd/ヘキスト消光効果により測定されたDNAへの5−ブロモ−2’−デオキシウリジン(BrdUrd)取り込みでの5−フルオロ−2’−デオキシウリジン(FdUrd)の効果。Cytometry 6、513〜20。
11. Benson,D.W.、MacRae,C.A.、Vesely,M.R.、Walsh,E.P.、Seidman,J.G.、Seidman,C.E.、およびSatler,C.A.(1996)。HERGの穴領域でのミスセンス変異は、家族性の長鎖QT症候群を引き起こす。Circulation 93、1791〜5。
12. Curran,M.E.、Splawski,I.、Timothy,K.W.、Vincent,G.M.、Green,E.D.、およびKeating,M.T.(1995)。心臓性不整脈に対する分子基礎:HERG変異は、長鎖QT症候群を引き起こす。Cell 80、795〜803。
13. Sanguinetti,M.C.、Jiang,C.、Curran,M.E.、およびKeating,M.T.(1995)。遺伝された心臓性不整脈と後天性の心臓性不整脈との機械的結合:HERGはIKrカリウムチャンネルをコードする。Cell 81、299〜307。
14. Spector,P.S.、Curran,M.E.、Keating,M.T.、およびSanguinetti,M.C.(1996)。クラスIII 抗不整脈薬物はHERG、ヒト心臓遅延レクティファイヤーK+ チャンネルをブロックする。メタンスルホンアニリドによるオープンチャンネルブロック。Circ Res 78、499〜503。
15. Trudeau,M.C.、Warmke,J.W.、Ganetzky,B.、およびRobertson,G.A.(1995)。HERG、電圧ゲートカリウムチャンネルファミリーでのヒト内部レクティファイヤー。Science 269、92〜5。
16. Suessbrich,H.、Waldegger,S.、Lang,F.、およびBusch,A.E.(1996)。ヒスタミンレセプターアンタゴニストであるテルフェナジンおよびアステミゾールによるツメガエル卵母細胞で発現されたHERGチャンネルの遮断。FEBS Lett 385、77〜80。
17. 例えば、DeCoursey,T.E.、Chandy,K.G.、Gupta,S.、Cahalan,M.D. Nature 307、465(1984);Mauro,T.、Dixon,D.B.、Komuves,L.、Hanley,K.、Pappone,P.A. J.Invest.Dermat.108、864(1997)を参照のこと。
18. Rouzaire−Dubois,B.およびDubois,J.M. J.Physiol.510、93(1998)。
19. Arcangeli,A.、L.Bianchiら、J.Physiol.489、455〜471(1995)。Bianchi,L.ら、Cancer Res.58、815〜822(1998)。
20. Brueggemann,A.,Stuehmer,W.、Pardo,L.A. Proc.Natl.Acad.Sci.USA.94,537(1997)。Pardo,L.A.、Brueggemann,A.、Camacho,J.、Stuehmer,W. J.Cell.Biol.143、767(1998)。
21. Ludwig,J.ら、EMBO J.13、4451(1994)。
22. Mosmann,T. J.Immunol.Meth.65、55(1983)。細胞をMTT(チアゾリルブルー)50μg/mlで4時間インキュベートし、ホルマザン結晶を10%SDS、10mM HClで12〜14時間で可溶化した。ELISAリーダーを使用して、650nmの参照波長で、570nmでの光学密度を測定した。
23. Stein,G.S.ら、細胞増殖およびアポトーシス。実用的な研究方法、G.P.Studzinski編(IRL Press,Oxford,1995)pp.193〜203。
24. Jainchill,J.L.、Aaronson,S.A.、Todaro,G.J. J.Viol.4、549(1969)。
25. Hermouet,S.、Merendino,J.J.,Jr.、Gutkind,S.、Spiegel,A.M. Proc.Natl.Acad.Sci.USA 88、10455(1991)。
26. 本論文で報告された配列に対するGenBank受入れ番号は、AF078741(heag A)およびAF078742(heag B)である。
27. Occhiodoro,T.ら、FEBS Lett.434,177(1998)。
28. Warmke,J.W.およびGanetzki,B. Proc.Natl.Acad.Sci,USA.91,3438(1994)。Frings,S.ら、J.Gen.Physiol.111、583(1998)。
29. Heng,H.H.Q.、Squire,J.、Tsui,L.−C. Proc.Natl.Acad.Sci,USA、89、9509(1992);Heng,H.H.Q.、およびTsui,L.−C. Chromosoma、102、325(1993)。我々は、FISH解析に対して、Dr.Henry Heng(DNA Biotech,Downsview,Ontario,Canadaを参照のこと)に謝意を表明する。
30. Terlau,H.ら、Pfluegers Arch.−Eur.J.Physiol.、432、301(1996)。Terlau,H.、Heinemann,S.H.、Stuehmer,W.、Pongs,O.、Ludwig,J. J.Physiol.、502、537〜543(1997)。Meyer,R.およびHeinemann,S.H. J.Physiol.508、49(1998)。
31. Bijlenga,P.ら、J.Physiol.512、317(1998)。
32. Bosma,G.C.,ら、Nature 301、527〜530(1987)。
33. Chen,C.およびOkayama,H. Mol.Cell Biol.7、2745(1987)。
34. Chomczynski,P.およびSacchi,N. Anal.Biochem.162、156(1987)。
35. Soto,F.ら、Proc.Natl.Acad.Sci.USA.93、3684(1996)。
36. McClelland,A.、Kuehn,L.C.、Ruddle,F.H. Cell 39、267(1984)。Schneider,C.、Owen,M.J.、Banville,D.、Williams,J.G. Nature 311、675(1984)。
37. Evans,P.およびKemp,J. Gene 199、123(1997)。
【0100】
本発明の態様として、以下のものが挙げられる。
[1](a)配列番号:3または4のアミノ酸配列を有するポリペプチドをコードする核酸分子を含有してなる核酸分子;
(b)配列番号:13または14のDNA配列を有する核酸分子を含有してなる核酸分子;
(c)(a)または(b)の核酸分子の相補的な鎖にハイブリダイズする核酸分子;または
(d)(c)の核酸分子の配列に縮重した核酸分子、
である、ヒトK+ イオンeagチャンネルの機能を有する(ポリ)ペプチドをコードする核酸分子を含有してなる核酸分子。
[2]配列5’−GGGAGGATGACCATGGCTを含有してなる[1]記載の核酸分子に特異的にハイブリダイズする核酸分子。
[3]DNAである[1]または[2]記載の核酸分子。
[4]RNAである[1]または[2]記載の核酸分子。
[5]融合タンパク質をコードする[1]〜[4]いずれか1つに記載の核酸分子。
[6][1]〜[5]いずれか1つに記載の核酸分子を含有してなるベクター。
[7]発現ベクターおよび/または遺伝子ターゲッティングまたは遺伝子導入ベクターである[6]記載のベクター。
[8][6]または[7]記載のベクターで形質転換された宿主。
[9]哺乳類細胞、真菌細胞、植物細胞、昆虫細胞または細菌細胞である[8]記載の宿主。
[10][8]または[9]記載の宿主を培養する工程、および産生された(ポリ)ペプチドを単離する工程を含む、[1]または[3]〜[5]いずれか1つに記載の核酸分子によってコードされた(ポリ)ペプチドの生産方法。
[11][1]または[3]〜[5]いずれか1つに記載の核酸分子によってコードされた、または[10]記載の方法によって生産された(ポリ)ペプチド。
[12][11]記載の(ポリ)ペプチドに対し特異的な抗体。
[13]モノクローナル抗体である[12]記載の抗体。
[14][2]記載の核酸分子、[6]記載のベクター、[11]記載のポリペプチドおよび/または[12]または[13]記載の抗体および薬学的に許容できる担体および/または希釈剤および/または賦形剤を含有してなる医薬組成物。
[15][1]〜[5]いずれか1つに記載の核酸分子、[6]記載のベクター、[11]記載のポリペプチドおよび/または[12]または[13]記載の抗体を含有してなる診断用組成物。
[16][1]または[3]〜[5]いずれか1つに記載の核酸分子の望まれていない発現または過剰発現によって引き起こされる疾患の予防または治療方法であって、[1]または[3]〜[5]いずれか1つに記載の核酸分子の発現のインヒビター、または[11]記載の(ポリ)ペプチドの機能のインヒビターを、前記疾患により影響を受けた哺乳動物または前記疾患の影響を受けやすいと推測される哺乳動物に導入する工程を含む、前記疾患の予防または治療方法。
[17][11]記載の(ポリ)ペプチドの機能不全により引き起こされる疾患の予防または治療方法であって、[1]または[3]〜[5]いずれか1つに記載の核酸分子の発現のインヒビター、[11]記載の(ポリ)ペプチドの機能不全のインヒビターもしくは改変剤、heagをコードする[1]または[3]〜[5]いずれか1つに記載の核酸分子、またはheag活性を有する[11]記載のポリペプチドを、前記疾患の影響を受けた哺乳動物または前記疾患の影響を受けやすいと推測される哺乳動物に導入する工程を含む、前記疾患の予防または治療方法。
[18]前記核酸分子の発現または過剰発現の前記インヒビターが、[2]記載の核酸分子である、[16]記載の方法。
[19]ポリペプチド機能の前記インヒビターが、[12]または[13]記載の抗体、または薬物、好ましくはアステミゾールまたはテルフェナジンである、[16]記載の方法。
[20]導入工程の前に、
(a)前記疾患により冒された哺乳動物から細胞を得る工程ならびに前記導入工程の後に前記疾患により冒された哺乳動物から細胞を得る工程、ここで、前記導入は前記細胞に行われる、
(b)前記細胞を前記哺乳動物または同種の哺乳動物に再導入する工程、
をさらに含む、[16]〜[19]いずれか1つに記載の方法。
[21]前記細胞が、生殖細胞、胚性細胞もしくは卵細胞またはそれらに由来する細胞である、[16]〜[20]いずれか1つに記載の方法。
[22][1]および[3]〜[5]いずれか1つに記載の核酸分子の望まれていない発現または過剰発現によって引き起こされる疾患の治療用薬物の設計方法であって、
(a)特異的かつ効力の高い薬物を同定する工程;
(b)部位特異的突然変異誘発およびキメラタンパク質研究による、前記薬物の結合部位を同定する工程;
(c)(ポリ)ペプチドの結合部位および前記薬物の構造の両方を分子モデリングする工程;ならびに
(c)(ポリ)ペプチドに対するその結合特異性を改良するために薬物を改変する工程、
を含む、治療用薬物の設計方法。
[23](a)アンチセンスオリゴヌクレオチドおよび/またはリボザイムから選ばれた化合物を、翻訳の阻害または低減に対して試験する工程;または
(b)[11]記載の(ポリ)ペプチドをコードし、かつ好ましくはその転写因子応答エレメントを有する遺伝子のプロモーター領域に結合する化合物を、転写の阻害に対して試験する工程;または
(c)[11]記載の(ポリ)ペプチドの増殖活性をブロックすると推測されるペプチドまたは抗体を、前記ブロック活性に対して試験する工程、
を含む、[1]または[3]〜[5]いずれか1つに記載の核酸分子の発現のインヒビターまたは[11]記載の(ポリ)ペプチドの機能のインヒビターの同定方法。
[24]前記薬物またはインヒビターが、ペプチド疑似体またはファージディスプレイもしくはコンビナトリアルライブラリー技術を適用することによってさらに改良された、[22]または[23]記載の方法。
[25][1]または[3]〜[5]いずれか1つに記載の核酸分子または[11]記載の(ポリ)ペプチドの発現に対し、インヒビターを適用する工程を含む、細胞増殖の阻害方法。
[26][1]および[3]〜[5]いずれか1つに記載の核酸分子の発現を評価する工程、または哺乳動物の細胞において[11]記載のポリペプチドの定量的な存在を評価する工程を含む、癌および/または神経変性疾患(neurodegenerative diseases)および/または乾癬の予知方法。
[27]前記癌が、乳癌または神経芽腫または子宮頸癌(cervix carcinoma)である、[26]記載の方法。
[28]前記乳癌が、胸腺癌(breast adenocarcinoma)、胸癌管型(breast carcinoma ductal type)である、[27]記載の方法。
[29]前記神経変性疾患が、アルツハイマー病、パーキンソン病、筋萎縮性側索硬化症または多発性硬化症である、[26]記載の方法。
[30]前記哺乳動物が、ヒト、ラットまたはマウスである、[16]〜[21]および[26]〜[29]いずれか1つに記載の方法。
[31]遺伝子治療における[1]〜[5]いずれか1つに記載の核酸分子の使用。
[32][2]記載の核酸分子、[6]記載のベクター、[11]記載のポリペプチドおよび/または[12]または[13]記載の抗体を含有してなるキット。
【特許請求の範囲】
【請求項1】
核酸分子の望まれていない発現または過剰発現によって引き起こされる疾患の治療用薬物の設計方法であって、
(a)特異的かつ効力の高い薬物を同定する工程;
(b)部位特異的突然変異誘発およびキメラタンパク質研究による、前記薬物の結合部位を同定する工程;
(c)(ポリ)ペプチドの結合部位および前記薬物の構造の両方を分子モデリングする工程;ならびに
(d)(ポリ)ペプチドに対するその結合特異性を改良するために薬物を改変する工程、
を含み、該核酸分子が、
(A)配列番号:3または4のアミノ酸配列を有するポリペプチドをコードする核酸分子を含有してなる核酸分子;
(B)配列番号:13または14のDNA配列を有する核酸分子を含有してなる核酸分子;
(C)(A)または(B)の核酸分子の相補的な鎖にハイブリダイズする核酸分子;または
(D)(C)の核酸分子の配列に縮重した核酸分子、
である、ヒトK+ イオンeagチャンネルの機能を有する(ポリ)ペプチドをコードする核酸分子を含有してなる、治療用薬物の設計方法。
【請求項1】
核酸分子の望まれていない発現または過剰発現によって引き起こされる疾患の治療用薬物の設計方法であって、
(a)特異的かつ効力の高い薬物を同定する工程;
(b)部位特異的突然変異誘発およびキメラタンパク質研究による、前記薬物の結合部位を同定する工程;
(c)(ポリ)ペプチドの結合部位および前記薬物の構造の両方を分子モデリングする工程;ならびに
(d)(ポリ)ペプチドに対するその結合特異性を改良するために薬物を改変する工程、
を含み、該核酸分子が、
(A)配列番号:3または4のアミノ酸配列を有するポリペプチドをコードする核酸分子を含有してなる核酸分子;
(B)配列番号:13または14のDNA配列を有する核酸分子を含有してなる核酸分子;
(C)(A)または(B)の核酸分子の相補的な鎖にハイブリダイズする核酸分子;または
(D)(C)の核酸分子の配列に縮重した核酸分子、
である、ヒトK+ イオンeagチャンネルの機能を有する(ポリ)ペプチドをコードする核酸分子を含有してなる、治療用薬物の設計方法。
【図1】


【図2】
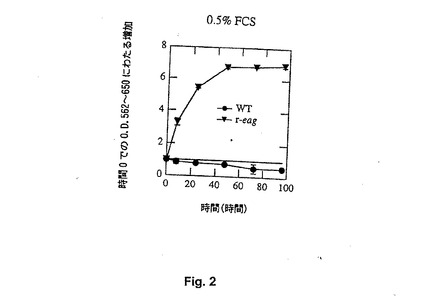
【図3】
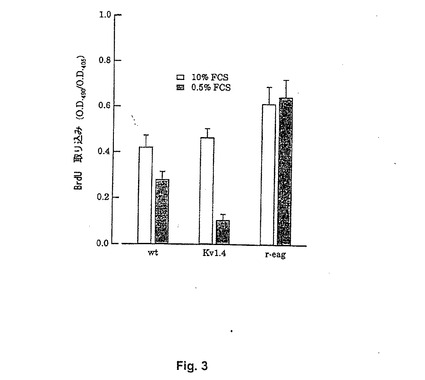
【図4a】
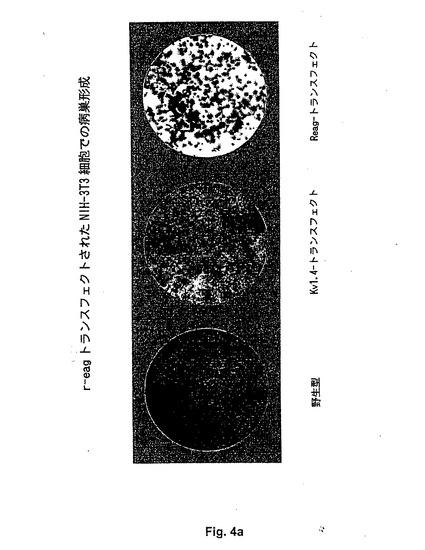
【図4b】
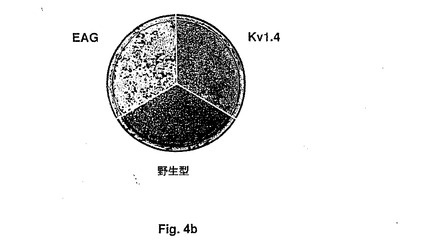
【図5】
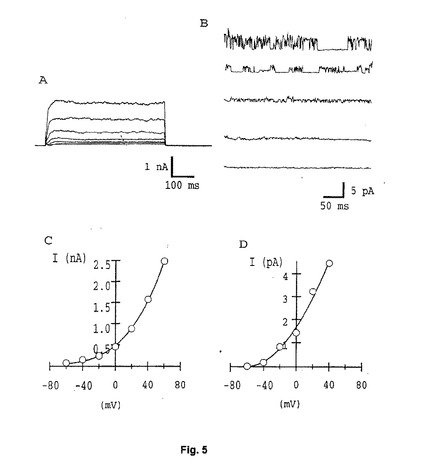
【図6】

【図7】

【図8】
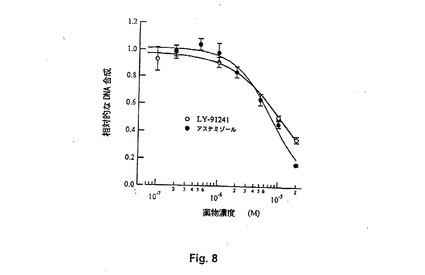
【図9】

【図10−1】

【図10−2】

【図10−3】

【図10−4】

【図10−5】

【図10−6】

【図10−7】

【図10−8】

【図10−9】

【図10−10】

【図10−11】

【図11−1】

【図11−2】

【図11−3】

【図11−4】

【図11−5】

【図11−6】

【図11−7】

【図11−8】

【図11−9】

【図11−10】

【図12】
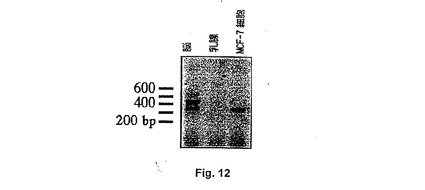
【図13】

【図14】

【図15】
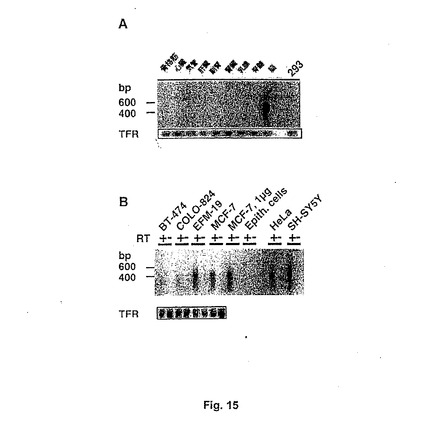
【図16】

【図17−1】
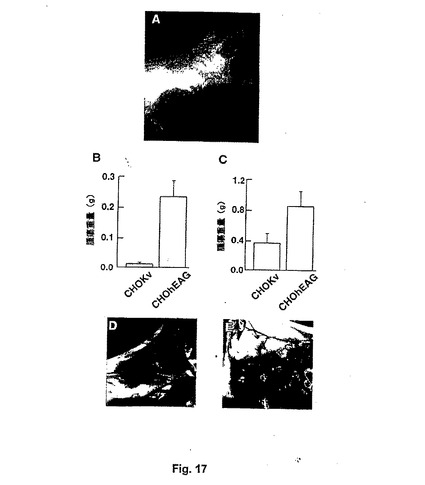
【図17−2】

【図18】

【図2】
【図3】
【図4a】
【図4b】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10−1】
【図10−2】
【図10−3】
【図10−4】
【図10−5】
【図10−6】
【図10−7】
【図10−8】
【図10−9】
【図10−10】
【図10−11】
【図11−1】
【図11−2】
【図11−3】
【図11−4】
【図11−5】
【図11−6】
【図11−7】
【図11−8】
【図11−9】
【図11−10】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17−1】
【図17−2】
【図18】
【公開番号】特開2009−235091(P2009−235091A)
【公開日】平成21年10月15日(2009.10.15)
【国際特許分類】
【出願番号】特願2009−160236(P2009−160236)
【出願日】平成21年7月6日(2009.7.6)
【分割の表示】特願2000−544795(P2000−544795)の分割
【原出願日】平成11年4月21日(1999.4.21)
【出願人】(500059047)マックス−プランク−ゲゼルシャフト ツール フォルデルング デル ヴィッセンシャフテン エー.ファウ. (1)
【Fターム(参考)】
【公開日】平成21年10月15日(2009.10.15)
【国際特許分類】
【出願日】平成21年7月6日(2009.7.6)
【分割の表示】特願2000−544795(P2000−544795)の分割
【原出願日】平成11年4月21日(1999.4.21)
【出願人】(500059047)マックス−プランク−ゲゼルシャフト ツール フォルデルング デル ヴィッセンシャフテン エー.ファウ. (1)
【Fターム(参考)】
[ Back to top ]