
心筋収縮性をinvivoで調節するための方法
うっ血性心不全の動物の心筋中のカルシウム輸送をin vivoで調節するための方法を開示する。本方法によれば、(うっ血性心不全の発症と同時に低減する)カルシウムATPアーゼ活性、及び心筋収縮性が、この酵素を作動的にコードしている遺伝子を心臓に送達することによって増大される。それだけには限らないが、アデノ随伴ウイルスベクターの使用を含めて、送達系が提供される。この遺伝子産物の発現及び心筋の性能に対するその効果をモニターする方法も提供される。
【発明の詳細な説明】
【技術分野】
【0001】
関連特許出願へのクロスリファレンス
本発明は、1995年11月4日出願の現在も係属中の米国特許出願第08/420,306号の一部継続出願である。
【0002】
1.発明の分野
本発明は、心筋収縮性を調節するための方法に関する。より詳細には、本発明は、SERCA2蛋白質を作動的にコードする遺伝子をin vivoで送達することによって、心臓の筋小胞体(SR)カルシウム2++ATPアーゼ(SERCA2)のin vivoのレベルを上昇させる方法に関する。
【背景技術】
【0003】
2.先行技術の変遷
うっ血性心不全は、米国において成人の主な死亡原因のうちの1つである。心虚血(心臓への血液供給が妨害され又は失われることにより生じる急性の事象)と比べて、うっ血性心不全は、心筋の収縮性及びストレスに対する心臓の適応性の漸進的な消失を伴う比較的潜行性の事象である。結局、有効な治療法はなく、CHF心臓は、身体の代謝要求を満たすのに十分な速度で血液をポンプ輸送する能力を失う。
【0004】
うっ血性心不全(CHF)に付随する心機能の異常は様々であるが、収縮性蛋白質の活性化に必要とされるCa2++イオンのSRからの放出の低減は、CHF症候群の共通の特徴である。この消失の意味は、カルシウム輸送が心臓の正常な機能活動において果たしている役割という文脈で最も良く理解することができる。
【0005】
簡単に言うと、SRは心筋の各筋原線維を取り囲む膜性構造物である。SERCA2は、SRの膜内部に含まれており、心筋の拡張期弛緩の間にSR内部空間に遊離カルシウムイオンの70〜80%を能動輸送する働きをする。輸送に利用可能な残存カルシウムイオンの多くは、SRのナトリウム/カルシウム交換輸送系、並びに程度ははるかに低いが、筋細胞膜カルシウムイオンATPアーゼが触媒するATP加水分解によって駆動される輸送、及びミトコンドリアのカルシウム取り込みによって、細胞質から取り除かれる(Bassani他、J.Physiol.、453:591〜608、1992年、及びCarafoli,E.、Ann.Rev.Biochem.、56:395〜433、1987年)。
【0006】
SERCA2のATP加水分解活性とSERCA2 mRNAの絶対レベルの双方がCHF心臓において低下することから(Hasenfuss他、Circ.Res.、75:434〜442、1994年、及びStuder他、Circ.Res.、75:443〜453、1994年)、CHF心臓の低圧力で血液を受け取る能力の損傷は、SERCA2媒介による、収縮を活性化するカルシウムイオンのSRへの輸送の遅れに直接関連づけられ、その結果心臓の拡張期弛緩が遅れると広く想定されている(例えば、Grossman,W.、N.Engl.J.Med.、325:1557〜1564、1991年;Lorell,BH、Ann.Rev.Med.、42:411〜436、1991年;及びArai他、Circ.Res.、74:555〜564、1994年を参照のこと)。これらの知見は、特にmRNAのSERCA2コードレベルの低下に関して、ヒト並びに他の哺乳動物種において確かめられている。(ヒトSERCA2 mRNAレベルについては、Arai他の上記論文、及びMercadier他、J.Clin.Invest.、85:305〜309、1990年を参照のこと。また、他の哺乳動物種の肥大心臓組織におけるSERCA2 mRNAレベルの低減については、Wang他、Am.J.Physiol.、267:H918〜H924、1994年(フェレット);Afzal及びDhella、Am.J.Physiol.、262:H868〜H874、1992年(げっ歯類);及びFeldman他、Circ.Res.、73:184〜192、1993年(げっ歯類)を参照のこと)。
【0007】
CHFにおけるカルシウム輸送の役割に関して近年関心が持たれているにもかかわらず、SERCA2カルシウム輸送活性の損傷の分子的原理は少ししか理解されておらず、CHFの治療レジメンにまだ利用されていない。そうではなくて、CHF症候群のための現在の治療様式は、CHF心臓の不全につながる機能異常を引き起こさないとしてもそれに付随すると考えられている生化学的事象及び分子的事象を直接標的としていないという意味で、ほとんど非特異的である。例えば、アドレナリン様薬物を投与することによるCHFの薬剤治療は、心筋収縮を刺激するが、筋肉の収縮性の減退を引き起こした根底にある状態を是正しない。
【0008】
従って、in vivoの心臓蛋白質レベルを補充及び/又は上昇させることは、ヒトのCHFの進行を治療及び制御するための魅力的な代替方法である。しかし、心臓組織に心臓蛋白質又は心臓ペプチドを導入することによってこの目標を実現することは、成功する可能性が低い。特に蛋白質に対する生体反応を起こすのに十分な用量で、潜在的な毒性を生じる危険性が主要な懸念である。実用的な観点からは、これらの蛋白質を単離し精製すること、又は合成することに伴うコストの問題もある。さらに、標的組織中に存在するプロテアーゼによって分解されるため、in vivoでの半減期が比較的短いことにより、これらの蛋白質の臨床上の効果も限られるはずである。
【0009】
これらの理由により、この蛋白質を発現する遺伝子の送達により、患者に蛋白質を導入することは、蛋白質それ自体の投与に代わる魅力的な代替方法である。この目的で、標的細胞に外来遺伝子を導入するための様々な戦略が開発されている。ヒトで使用するために今日までに提案されている遺伝子治療プロトコールの大半は、ex vivoの遺伝子移入に焦点を合わせている。例えば、標的組織に移植するための細胞のレトロウイルスによるトランスフェクションなどがある(例えば、Anderson,WF、Science,256:808〜813、1992年、及びMiller,AD、Nature、357:455〜460、1992年(アデノシン脱アミノ酵素欠損の治療)を参照のこと)。しかし、これらのプロトコールの有用性は、蛋白質発現が比較的非効率的であること並びに標的器官及び組織への到達性が限られていることにより、限定的であることが判明した。
【0010】
従って、in vivoの遺伝子送達方法は、当技術分野で関心の高いテーマである。この目的で、「裸の(naked)」ポリヌクレオチド(プラスミド)、ウイルスに連結したプラスミド、ウイルスに取り込ませた(cointernalized)プラスミドの導入、並びにリポソームによる遺伝子構築体のカプセル化及び送達を含めて、この目標を達成するためにいくつかの系が開発されている。
【0011】
例えば、リス肝炎の裸のクローン化プラスミドDNAをリスに肝臓内注入したところ、ウイルス感染及び抗ウイルス性抗体の形成が共にリスで生じたというNIH(米国国立衛生研究所)の研究が1984年に報告された(Seeger他、Proc.Nat’l.Acad.Sci USA、81:5849〜5852、1984年)。数年後、Felgner他は、骨格筋組織に注入した「裸の」ポリヌクレオチド(すなわち、リポソーム又はウイルス発現ベクターを伴わないDNA又はRNA)から蛋白質発現を得たことを報告した(Felgner他、Science、247:1465、1990年。また、PCT国際特許出願公開第90/11092号も参照のこと)。Felgner他は、筋組織が、多核細胞、筋小胞体、及び筋細胞の深部まで伸びる横行の管状系からなる独特な構造を有するため、筋細胞がポリヌクレオチドを効率的に取込み発現すると推測した。
【0012】
標的組織に直接遺伝子を送達するための同様の系が、Stribling他、Proc.Natl.Acad.Sci.USA、89:11277〜11281、1992年(リポゾームにカプセル化した遺伝子のエアロゾル送達後に検出された蛋白質発現)及びTang他、Nature、356:152〜154、1992年(ワクチン「銃」を用いてコロイド性の金粒に結合させたhGHプラスミドをマウスの皮膚に注入した結果、注入された遺伝子に対する免疫応答を誘発せずにhGH蛋白質を発現した)によって報告されている。筋細胞内で高レベルの蛋白質発現を起こすのに一般に有効ではあるが、長期治療のための筋組織へのDNA又はRNAの直接注入では、遺伝子の分解による発現の損失を補うために注入を繰り返し利用する必要がある。この手法は、時間がかかり高価であるだけでなく、注入部位及びその周辺で炎症が引き起こされるため、長期治療には非実用的であるかもしれない。従って、短期治療又は救急治療では有用であるが、標的組織に発現可能な遺伝子を導入するための侵襲性のより低い手段が、標的組織への直接注入よりも一般に好ましい。
【0013】
さらに、組換え発現ベクターを使用しないin vivoの遺伝子送達方法の大半は、標的細胞のトランスフェクションが非効率的で蛋白質発現が比較的弱いという欠点がある。従って、組換え発現ベクター(特に複製不能型のベクター)が現在もなおin vivoの遺伝子送達のための好ましい運搬体(vehicle)である。
【0014】
心筋細胞は、in vivo遺伝子送達に適した標的であることが示されている。例えば、CHFの治療に対する近年の1つの提案を用いれば、CHF心臓の筋細胞の活性β2−アドレナリン受容体の数が補充及び/又は増加されるはずである。マウスでの研究により、直接移植技術を使用してこれらの受容体をコードする遺伝子を導入したところ、外因性のアドレナリン供給源の不在下でさえ心筋の収縮性をin vivoで増大させたことが示されている(Lefkowitz他、Science、264:582〜586、1994年)。特に、アデノ随伴ウイルスベクターが、心筋細胞への遺伝子送達のための成功裡の運搬体として示されている(例えば、Guzman他、Circ.Res.、73:1202〜1207、1993年、及びMuhlhauser他、Circulation、88(Part2):1〜475、1993年を参照のこと)。
【発明の開示】
【発明が解決しようとする課題】
【0015】
本発明の好ましい実施形態の詳細を、添付図及び以下の説明に示す。本発明の詳細が知られれば、非常に多くのさらなる革新及び変更が当業者には明らかになるであろう。
【0016】
本発明は、SERCA2を作動的にコードするポリヌクレオチドを心筋などの組織に導入することにより触媒活性のあるSERCA2の発現をin vivoで高めることによって、心筋のSRへのカルシウム輸送を増大させる方法である。標的細胞のトランスフェクション効率の点で好ましい実施形態では、SERCA2をコードしているポリヌクレオチドを、複製不能型のウイルス組換え発現ベクター、最も好ましくはアデノ随伴ウイルスベクター(AAV)構築体を用いて心臓組織に送達する。
【0017】
或いは、SERCA2コード遺伝子を、リポソームによるカプセル化又は「裸の」ヌクレオチドの投与によって提供してもよい。SERCA2遺伝子の送達は、外科的に(即ち、ベクター又はトランスフェクトした細胞の標的組織への直接導入によって)又は冠動脈内輸注法によって行ってよい。
【0018】
SR中のSERCA2活性上昇によってもたらされるSRへのカルシウムイオン輸送の増大により、心臓の筋原線維収縮の活性化が緩和されることになる。従って、本発明の方法の特に有利な点は、特に心筋の拡張期弛緩の初期段階を短くすることにより、CHFに関連する異常に対して心臓が調整するのを助けることである。
【0019】
本発明の方法の一使用は、インターベンション治療(例えば移植)を待機している、且つ/又は既にCHF治療を受けている患者のCHFの短期改善を目的としている。本発明の方法の使用はまた、CHFに関連する心機能の異常を生じるリスクがある、或いは既に異常を生じ始めている患者に恩恵をもたらすと予想される。
【0020】
本発明の他の態様では、本発明の方法で使用するためのベクターを開発する際及び本発明の方法の適用を試験する際に使用するためのトランスジェニック動物が提供される。CHFの治療に使用したときの本発明の方法の有効性をモニターする方法も説明する。
【発明を実施するための最良の形態】
【0021】
この説明を通じて、ここに示す好ましい実施形態及び実施例は例とみなすべきで、本発明を限定するものとみなすべきではない。
【0022】
A.定義
以下の定義は、本発明の考察を簡単にするために提供される。しかし、これらの定義が本発明の正当な範囲又は趣旨から逸脱しない等価物を含むように拡張できることを、当業者なら理解するであろう。この理由から、これらの定義は本発明を限定するものと解釈されるべきではない。
1.「SERCA2ポリヌクレオチド」とは、DNA又はRNAを意味し、本発明に従って行う治療の目的に適したセンス鎖及びアンチセンス鎖を含むことができる。本明細書では、「ポリヌクレオチド」とは、分離した断片の形又は大きな構築体の成分としての、デオキシリボヌクレオチド又はリボヌクレオチドの重合体を意味する。ポリヌクレオチドの配列は、遺伝コードから推測できるが、コードの縮重を考慮しなければならない。本発明のポリヌクレオチドは、遺伝コードの結果として縮重した配列を含み、その配列は当業者によって容易に決定することができる。
2.「作動的にコードする」とは、所望の翻訳産物、例えばペプチド又は蛋白質の発現及び望むならその分泌に必要なプロモーター及び他の配列を含むように改変されたポリヌクレオチドを意味する。本発明の全ての実施形態は、公知の組換え発現ベクターを使用して実施することができる。これらのベクターは、所望の翻訳産物をコードするcDNAを含むことが好ましい。従って、文脈上別の意味で解釈する必要がない限り、「ポリヌクレオチド」とは、適切な組換え発現ベクター中に含まれ作動的にコードする配列を意味すると想定され、その例が本明細書で提供される。
3.「合成」とは、ポリヌクレオチド配列及びポリペプチド配列を合成する公知の手段を意味し、天然のポリヌクレオチド及び蛋白質の単離及び精製を含むことができる。
4.「ペプチド」とは、in vivoで所望の生体効果を有する小型ペプチド、ポリペプチド、オリゴペプチド、及び蛋白質を意味する。
5.「送達」とは、作動的にコードするポリヌクレオチドを宿主に導入する既知の方法を意味する。当業者なら、これらの送達手段を熟知しており、或いは容易に特定することができる。しかし、特に有用な送達手段に関しては、「新規の薬物送達系(Novel Drug Delivery Systems)」(Marcel Dekker、1992年)を参照することができ、その関連する開示内容を、薬物送達技術に関する当技術分野の知識状態を例示するために参照により本明細書に組み込む。
6.「宿主」とは、本発明に従って行われる治療の受容者を意味する。宿主はどんな脊椎動物でもよいが、好ましくは哺乳動物である。哺乳動物の場合は、宿主はヒトが好ましいが、飼育された家畜でもペット動物でもよい。
7.「標的組織」とは、作動的にコードするポリヌクレオチドの発現を得ようとする、宿主の組織を意味する。
8.「抗体」とは、任意のクラスの免疫グロブリン全体、キメラ抗体、2つ又は複数の抗原特異性を有するハイブリッド抗体、及びハイブリッド断片を含む断片を意味する。また、このような断片の結合体、例えば米国特許第4,704,692号に記載されているいわゆる抗原結合性蛋白質(単鎖抗体)、及び、例えば米国特許第4,699,880号に記載されている抗イディオタイプ抗体(他の抗体に結合する抗体)も、「抗体」の意味の範囲内に含まれる。
9.「組換え発現ベクター」は、真核生物又は原核生物で発現可能なポリペプチドを作動的にコードするポリヌクレオチドの系を意味する。真核生物又はウイルスの配列を有するDNA配列を原核生物で発現する方法は当技術分野で公知である。宿主中で発現及び複製できる、生物的に機能的なウイルス及びプラスミドDNAベクターも当技術分野で公知である。宿主としては、微生物、酵母、昆虫、及び哺乳類生物を挙げることができる。
【0023】
B.本発明の方法で使用するポリヌクレオチド構築体
ラットSERCA2のゲノムクローンのヌクレオチド配列(Rohrer他、J.Biol.Chem.、263:6941〜6944、1988年(ラットSERCA2 mRNA)も参照のこと。この論文で報告された配列を、参照のために本明細書に組み込む)。このクローンは、Rohrer他、J.Biol.Chem.、266:8638〜8646、1991年(その開示内容を参照のために本明細書に組み込む)に記載されているような従来のハイブリダイゼーション技術によって得られた(実施例1を参照のこと)。この論文は、クローンを転写するための開始コドン及び終止コドンも記載している。
【0024】
カルシウムATPアーゼのSERCA2アイソフォームのヌクレオチド配列(すなわち、横紋筋の「速筋型」アイソフォームに対する骨格筋の「遅筋型」アイソフォーム)は、哺乳動物種間で90%以上保存されている。従って、SERCA2は、ヒト(GENBANK #J4025、又、GENBANK M23114〜23116、M23277〜23279、及びLyttonとMacLennan、J.Biol.Chem.、263:15024〜15031、1988年も参照のこと)、及びウサギ(MacLennan,DH、Nature、316:697〜700、1985年、及びGENBANK M33834)を含めて、げっ歯類以外の哺乳動物の骨格筋組織からかなり容易に同定され、配列決定されている。当業者なら、ヒトSERCA2ポリヌクレオチドの使用がヒトの治療では大いに好ましいことを理解するであろうが、ラットSERCA2ポリヌクレオチドが、実施例で記載した実験のためには使用しやすく好ましい。
【0025】
SERCA2に関して詳細に本明細書に記載する本発明の方法は、CHF心臓でその発現及び活性が損傷されている他の心臓蛋白質を送達するために使用するように適合させることができるので有利であることも、当業者なら理解するであろう。特に、哺乳動物のカルシウム/ナトリウム交換体をコードしている遺伝子は、SERCA2ポリヌクレオチドの送達に関して本明細書に記載したのと同じ方法で、本発明の方法に従って心臓に送達するための特に魅力的な対象である。
【0026】
本開示に含まれるSERCA2配列及び当技術分野で公知のものを入手し使用するために、DNA及びRNAを当技術分野で公知の自動核酸合成装置を使用して合成することができる。公知のポリメラーゼ連鎖反応(PCR)の使用が、ポリヌクレオチド混合物を生成するのに特に好ましい。Ausubel他、Current Protocols in Molecular Biology、2章及び4章、(Wiley Interscience、1989年)に記載のプロトコールのような当技術分野で公知の手段によって、ゲノム核酸を調製することができる。cDNAは、当技術分野で公知の手段によって合成することができる(例えば、Maniatis他、Molecular Cloning、A Laboratory Manual(Cold Spring Harbor Lab、米国ニューヨーク州、1982年)を参照のこと)。目的のポリヌクレオチドを含むcDNAの発現ライブラリを、当技術分野で公知の手段によってスクリーニングすることもできる。
【0027】
例えば、これらの手段には、それだけには限らないが、1)共通のヌクレオチド配列を検出するための、ゲノム又はcDNAライブラリへのプローブのハイブリダイゼーション、2)共通の構造的特徴を検出するための発現ライブラリの抗体スクリーニング、及び3)ポリメラーゼ連鎖反応(PCR)による合成が含まれる。特定のDNAコード配列又はその断片の開発は、1)ゲノムDNAからの二重鎖DNA配列の単離、2)目的のポリペプチドのために必要なコドンを提供するDNA配列の化学的作製、及び3)真核性ドナー細胞から単離したmRNAの逆転写による二重鎖DNA配列のin vitro合成によって行うことができる。後者の場合、一般にcDNAと呼ばれるmRNAの二重鎖DNA相補物が、最終的に形成される。
【0028】
ハイブリダイゼーション手順は、混合した合成オリゴヌクレオチド標識プローブを使用して、組換えクローンをスクリーニングするのに有用であり、この場合、各プローブは変性させた二重鎖DNAの不均一な混合物を含むハイブリダイゼーション試料中にある特定のDNA配列の完全な相補物である可能性がある。このようなスクリーニングの場合、一本鎖DNA又は変性した二重鎖DNA上でハイブリダイゼーションを行うことが好ましい。ハイブリダイゼーションは、目的のポリペプチドに関係するmRNA配列が極めて少量しか存在しない供給源に由来するcDNAクローンの検出に特に有用である。言い換えれば、非特異的な結合を避けるようにした厳密なハイブリダイゼーション条件を使用することにより、例えば、混合物中のただ一つのプローブに標的DNAをハイブリダイゼーションすることによって、特定のcDNAクローンをオートラジオグラフィーで可視化させることが可能である。
【0029】
目的のポリヌクレオチドを含むと考えられるcDNAライブラリは、cDNAから誘導した様々なmRNAを卵母細胞に注入し、十分な時間を与えてcDNA遺伝子産物を発現させ、例えば目的のポリヌクレオチドにコードされたペプチドに特異的な抗体、又は目的のポリヌクレオチドにコードされたペプチドの特徴である繰り返しモチーフ及び組織の発現パターンを調べるためのプローブを使用して、所望のcDNA発現産物が存在するかどうか試験することによって、スクリーニングすることができる。或いは、cDNAライブラリは、少なくとも1種のエピトープを有する治療用及び/又は免疫原性ペプチドの発現について、それらのペプチドに特異的な抗体を使用して、間接的にスクリーニングすることもできる。これらの抗体は、ポリクローナル型又はモノクローナル型を作製し、目的のcDNAの存在を示す発現産物を検出するのに使用することができる。
【0030】
適切なプローブが利用可能であるならば、核酸ハイブリダイゼーションを利用するスクリーニング手順によって、任意の生物から任意の遺伝子配列を単離することが可能になる。問題の蛋白質をコードする配列の一部分に相当するオリゴヌクレオチドプローブを、化学的に合成することができる。このためには、アミノ酸配列の短いオリゴペプチド区間がわかっていなければならない。蛋白質をコードしているDNAの配列は、遺伝コードから推測できるが、コードの縮重を考慮しなければならない。配列が縮重しているときは、混合付加反応を行うことができる。これは、変性した二重鎖DNAの不均一な混合物を含む。このようなスクリーニングの場合、一本鎖DNA又は変性した二重鎖DNA上でハイブリダイゼーションを行うことが好ましい。
【0031】
本発明で使用するSERCA2ポリヌクレオチドはDNA又はRNAでよいが、相補的DNA(cDNA)配列であることが好ましい。本発明で使用するポリヌクレオチドは、(a)発現可能で、且つ(b)複製不能、又は宿主ゲノム内に入って複製しないように当技術分野で公知の手段によって改変されたものでなければならない。SERCA2蛋白質を作動的にコードするポリヌクレオチドを組換え発現ベクター、最も好ましくはアデノウイルス構築体の一部として使用することが好ましい。
【0032】
以下は、本発明で使用するのに適するポリヌクレオチドの調製の例であり、どのように特定のポリヌクレオチド組成物が作成されたかを示す特定の実施例が以下に提供される。しかし、複製不能なポリヌクレオチドを調製する他の既知の手段も適し得ることは、当業者には明らかであろう。
【0033】
本発明のポリヌクレオチドは、SERCA2蛋白質を作動的にコードする既知のポリヌクレオチドの機能的誘導体を含む。「機能的誘導体」とは、SERCA2の「断片」、「変異体」、「類似体」、又は「化学的誘導体」をコードするポリヌクレオチドを意味する。分子の「断片」は、分子の任意のペプチド部分を含む。このような分子の「変異体」とは、分子全体又はその断片にかなり類似した天然の分子を意味する。分子の「類似体」とは、分子全体又はその断片にかなり類似した非天然の分子を意味する。
【0034】
本明細書では、ある分子が普通ならその一部分ではない追加の化学的部分を含むとき、別の分子の「化学誘導体」であると言う。これらの部分は、分子の溶解性、吸収性、生物的半減期などを改善することができる。或いは、これらの部分は、分子の毒性を弱め、分子の望ましくない副作用を無くし又は減らすこともできる。このような効果をもたらすことができる当技術分野で既知の部分は、例えば、Remington’s Pharmaceutical Sciences、16版、Mack Publishing Co.、ペンシルベニア州、イーストン(1980年)に開示されている。
【0035】
SERCA2ポリヌクレオチドは、これらのポリペプチドの発現を制御する調節蛋白質を作動的にコードする他のポリヌクレオチドに結合させ又はそれらと一緒に使用し、或いは、認識配列、プロモーター配列、及び分泌配列を含むことができる。当業者なら、過度に実験をすることなく、調節ポリヌクレオチドを選別し、それらを本発明のSERCA2ポリヌクレオチドに組み込むことができるであろう。例えば、ネズミ又はヒトの系で使用するのに適したプロモーター及びその使用法が、前述のCurrent Protocols in Molecular Biologyの1章に記載されている。
【0036】
本発明で使用するためのSERCA2ポリヌクレオチドの特に好ましい形態は、組換え発現ベクターに組み込まれたものとなる。組換え発現ベクターを使用すると、標的組織における遺伝子の発現が長くなる。
【0037】
適切な組換え発現ベクターは当技術分野で公知であり、前述のCurrent Protocols in Molecular Biologyの1章に記載されているベクターが含まれる。2種の特に好ましいプラスミドプロモーターベクターは、pRSV(ラウス肉腫ウイルス(Rous sarcoma virus))及びpCMV(サイトメガロウイルス(cytomegalovirus))であり、特に後者である。この選択は、CMVプロモーターを使用すると、より高レベルの発現がこの状況で得られるという観察結果に基づく。
【0038】
RSVプロモーターを単離し、プラスミドベクターの構築にそれを使用するための適切なプロトコールは、Gorman他、Proc.Natl.Acad.Sci.USA、79:6777、(1982年)に記載されている。他の好ましいプラスミドベクターは、カリフォルニア州サンディエゴのインビトロゲン(Invitrogen)社から市販されているpREP7及びpREVである。ポリヌクレオチドのクローニングにおいて、mRNAの産生に特に適したプラスミドは、Kreig他、Nucleic Acids Res.、12:7057〜7070(1984年)に記載のpSP64Tクローニングベクターである。従来技術を使用して、開始コドンを含む任意のcDNAをこのプラスミドに導入し、発現されたDNA鋳型からmRNAを調製することができる。
【0039】
組換え発現ベクターで使用するための特徴が良く分かっている「on/off」スイッチは、抗生物質(テトラサイクリン)で調節されるプロモーター系である。このような系を構築する手段は、当技術分野で公知である。この点に関する総説について、当業者なら、Furth他、Proc.Natl.Acad.Sci.USA、91:9302〜9306、1994年(トランスジェニックマウスにおける、テトラサイクリンに調節された遺伝子発現制御)、Fishman他、J.Clin.Invest.、93:1864〜1868、1993年(心臓の遺伝子発現のテトラサイクリンによる制御)、及びNiwa他、Gene、108:193〜200、1991年(高発現のトランスフェクトタントのためのプロモーター系の使用)を参照できよう。テトラサイクリンプロモーター系の制御下でSERCA2をコードするプラスミドを、実施例4に記載する。
【0040】
本発明で利用できる様々なウイルスベクターには、アデノ随伴ウイルス(AAV)、アデノウイルス、ヘルペスウイルス、ワクシニア、又はレトロウイルスなどのRNAウイルスが含まれる。単一の外来遺伝子を挿入できるレトロウイルスベクターの例には、それだけには限らないが、モロニーマウス白血病ウイルス(Moloney murine leukemiavirus)(MoMuLV)、ハーベイマウス肉腫ウイルス(Harvey murine sarcoma virus)(HaMuSV)、マウス乳癌ウイルス(murine mammary tumor virus)(MuMTV)及びラウス肉腫ウイルス(Rous Sarcoma virus)(RSV)が含まれる。いくつかの他のレトロウイルスベクターは、複数の遺伝子を組み込むことができる。これらのベクターの全ては、形質導入された細胞を同定及び生成できるように、選択マーカーとして遺伝子を移入又は組み込むことができる。
【0041】
組換えレトロウイルスは欠損があるため、感染性のベクター粒子を産生するためには助けを必要とする。例えば、LTR内の調節配列の制御下にあるレトロウイルスの構造遺伝子の全てをコードしているプラスミドを含むヘルパー細胞系を使用して助けることができる。これらのプラスミドは、パッケージング機序でキャプシド形成のためのRNA転写物を認識するのを可能にするヌクレオチド配列を欠失している。パッケージングシグナルを欠失しているヘルパー細胞系には、それだけに限らないが、例えば、ψ2、PA317、及びPA12が含まれる。これらの細胞系は、ゲノムが全くパッケージされない(詰め込まれない)ため、空のウイルス粒子を産生する。パッケージングシグナルは損なわれていないが構造遺伝子が目的の他の遺伝子で置き換えられているこのようなヘルパー細胞系にレトロウイルスベクターが導入されると、ベクターはパッケージされることができ、ベクターウイルス粒子を産生することができる。
【0042】
DNA発現ベクターのうちでは、トランスフェクション効率(心筋細胞で最大70%)の点からAAVベクターが好ましい。このようなベクターは、外来DNAの比較的大きなセグメントを受け入れることができ、低い免疫原性で高タイターで生成することができるので好ましい。AAVベクターは、(例えばSERCA2ポリヌクレオチドのポリリジン結合体と比べて)冠動脈内輸注法に良く耐えるので有利である。
【0043】
アデノ随伴ウイルス(AAV)は、単純な非病原性の一本鎖DNAウイルスである。パッケージング配列を含むそのcap及びrep遺伝子は、ウイルスの先頭及び末尾を決めている逆方向反復配列の間に挟まれている。cap遺伝子は、ウイルスのキャプシド(capsil)(莢膜)蛋白質をコードし、rep遺伝子産物は、ウイルスの複製及び組み込みに関与している。AAVは、複製のために、ヘルパーウイルス、例えばアデノウイルス又は単純ヘルペスウイルスによって提供される追加の遺伝子を必要とする。AAVは様々な細胞型に感染する。そのウイルスDNAは、ヒト染色体19に優先的に組み込まれることができる。
【0044】
rep及びcap遺伝子を導入遺伝子で置き換えることによって、AAVベクターを作製し、遺伝子治療用ベクターとして使用することができる。ヒト因子IXのcDNAを含むAAVベクターが構築され、免疫適格性マウスの肝臓細胞及び筋肉細胞に感染させるために使用されている。これらのマウスは、6ヶ月以上の間、治療効果のある量の因子IX蛋白質を血液中に産生することができた。AAVベクターを大量調製するための組成物及び方法は、米国特許第6,416,992号に開示されている。
【0045】
本発明で使用するのに特に好ましいAAV構築体を、実施例1に示す。この構築体は、プロモーター、ポリリンカー、及びその他のものを含むシャトルベクター中にSERCA2ポリヌクレオチドをクローニングすることによって、形成される。試料中で使用する特定のアデノ随伴シャトルベクターは、Samulski他によって記載の(Journal of Virology 63(9):3822〜3828、1989年)プラスミドpSub201から誘導した。このプラスミドのXba I断片を、マルチクローニングサイト及びポリアデニル化シグナルの前にある572塩基対のヒトCMVエンハンサー/プロモーター断片に連結させた。このプラスミドをAAV−シャトルと呼び、サイズは約5000塩基対である。
【0046】
3’末端でフラッグシグナルペプチドに融着させたラットSERCA2aのコード配列が報告されている(He他、J.Clin.Invest 100:974〜980、1999年)。Kpn I/Xba I又はHind III/Xba酵素を使用して、緑蛍光蛋白質(GFP)をAAV−シャトル中に好都合にクローニングすることができる(He他、J.Clin.Invest 100:974〜980、1999年)。組換えクローンはDNA配列決定により確認され、大量のプラスミドpDGが記載され、CsCl大量調製プロトコールによって精製されている(Grimm他、Human Gene Therapy 9:2745〜2760、1998年)。
【0047】
純粋な感染性ウイルス粒子を調製するために、挿入されたcDNAを含むAAV−シャトルプラスミド及びpDGプラスミドは共に、CaPO4共沈法を使用して細胞にトランスフェクトされる。トランスフェクションに続いて、細胞は新鮮な培地で培養され、次いでウイルス粒子が回収され精製される。クローニングスキーム及び適切な制限部位の略図を図4に示す。
【0048】
(以下のセクションCで記載の)リポソーム送達のためのSERCA2のカプセル化も、複製可能なAAV構築体を使用する場合と同様に、ウイルスの免疫破壊を制限するはずである。しかし、ウイルスDNAの宿主ゲノムへの組み込みに伴う可能性のある危険性を考えると、このようなベクターの使用は、ベクター産生物の長期間の発現が患者の生存に極めて重要である状況に限られることになる可能性が高い。
【0049】
例えば、特定の標的細胞上の受容体に対するリガンドをコードするもう1つの遺伝子といっしょに1種又は複数の目的の配列をウイルスベクターに挿入することによって、ベクターを標的特異性にすることができる。例えば、糖、糖脂質、又は蛋白質をコードするポリヌクレオチドを挿入することによって、レトロウイルスベクターを標的特異性にすることができる。
【0050】
心臓に特異的なプロモーターを使用することにより、標的筋細胞以外の心筋細胞への本発明の組換え発現ベクターの非特異的トランスフェクションを回避することが好ましい。いくつかのこのようなプロモーターが現在知られており、トリβ−アクチンが挙げられる(実施例1を参照のこと)。当業者なら、ウイルスゲノムに挿入され、目的のSERCA2ポリヌクレオチドを含むウイルスベクターの標的特異的な送達を可能にすることができる他の特定のポリヌクレオチド配列について知っているか、過度に実験をすることなく容易に確認することができよう。
【0051】
心臓組織へのSERCA2導入遺伝子の送達のためにAAVベクターを使用することの利点は、このベクターに対して免疫応答が起こらず、導入遺伝子の発現が最大7ヶ月持続することである。SERCA2導入遺伝子の送達のためのAAVベクターの使用に関する詳細な説明を、以下の実施例1に示す。
【0052】
C.SERCA2ポリヌクレオチドの薬剤調製物
SERCA2ポリヌクレオチドの組成物及びSERCA2ポリヌクレオチドの混合物は、製薬上許容される懸濁液、溶液、又は乳濁液中に入れてよい。適切な媒体は生理食塩水を含み、SERCA2ポリヌクレオチドを標的組織に送達するために抗原提示細胞を使用しない実施形態の場合は、リポソーム調製物を含んでよい。
【0053】
より詳細には、製薬上許容される担体は、無菌の水性又は非水性溶液、懸濁液、及び乳濁液を含んでよい。非水溶媒の例としては、プロピレングリコール、ポリエチレングリコール、オリーブ油などの植物油、及びエチルオレエートなどの注射可能な有機エステルがある。水性の担体には、生理食塩水及び緩衝処理した媒体を含めて、水、アルコール性/水性溶液、乳濁液、又は懸濁液が含まれる。非経口用ビヒクル(vehicle)には、塩化ナトリウム溶液、ブドウ糖入りリンガー液、ブドウ糖及び塩化ナトリウム溶液、乳酸化リンガー液、又は不揮発性油が含まれる。静脈内用ビヒクルには、(ブドウ糖入りリンガー液をベースとしたものなどの)流動性で栄養性の補充液及び電解質補充液などが含まれる。例えば、抗菌剤、抗酸化剤、キレート化剤、不活性ガスなどの保存剤及び他の添加剤も存在してよい。さらに、SERCA2ポリヌクレオチドの組成物を、本発明に従って後で再構成し使用するために、当技術分野で公知の手段を使用して凍結乾燥してもよい。
【0054】
上記に論じた標的化ベクター送達系に加えて、コロイド分散系も標的化送達に使用してよい。コロイド分散系には、水中油乳濁液、ミセル、混合ミセル、及びリポソームを含めて、高分子複合体、ナノカプセル、微粒子、ビーズ、及び脂質ベースの系が含まれる。本発明の好ましいコロイド系はリポソームである。
【0055】
リポソームは、in vitro及びin vivoの送達運搬体として有用な人工膜小胞である。大きさ0.2〜4.0nmの大きな一枚膜小胞(LUV)が、大きな高分子を含む水性緩衝液のかなりの割合をカプセル化できることが示された。RNA、DNA、及び無傷のウイルス粒子は、水性の内部にカプセル化され、生物活性のある形で細胞に送達されることができる(Fraley他、Trends Biochem.Sci.、6:77、1981年)。リポソームは、哺乳動物細胞に加えて、作動的にコードするポリヌクレオチドを植物、酵母、細菌細胞中に送達するのに使用されている。リポソームが効率的な遺伝子移入運搬体となるためには、以下の特徴がなければならない。すなわち、(1)アンチセンスポリヌクレオチドをコードしている遺伝子を高効率でカプセル化するが、それらの生物活性は損なわないこと、(2)標的ではない細胞と比べて標的細胞に優先的且つ有意に結合すること、(3)運搬体中の水性内容物を、高効率で標的細胞の細胞質に送達すること、及び(4)遺伝情報を正確且つ有効に発現することである(Mannino他、Biotechniques、6:682、1988年)。
【0056】
リポソームの組成物は、通常ステロイド、特にコレステロールと組み合わせた、リン脂質、特に相転移温度の高いリン脂質の組み合わせである。他のリン脂質又は他の脂質も使用してよい。リポソームの物理的特性は、pH、イオン強度、及び二価の陽イオンの有無によって決まる。
【0057】
リポソーム調製に有用な脂質の例には、ホスファチジルグリセロール、ホスファチジルコリン、ホスファチジルセリン、ホスファチジルエタノールアミン、スフィンゴ脂質、セレブロシド、及びガングリオシドなどのホスファチジル化合物が含まれる。特に、脂質部分が14〜18個の炭素原子、特に16〜18個の炭素原子を含み、飽和されているジアシルホスファチジルグリセロールが有用である。リン脂質の例には、卵ホスファチジルコリン、ジパルミトイルホスファチジルコリン、及びジステアロイルホスファチジルコリンが含まれる。
【0058】
リポソームの標的化は、解剖学的及び機械的要因に基づいて分類することができる。解剖学的分類は、選択性のレベル、例えば、器官特異性、細胞特異性、及び細胞小器官特異性に基づく。機械的標的化は、それが受動的であるか能動的であるかに基づいて区別することができる。受動的標的化は、洞様毛細血管を含む器官において細網内皮系(RES)の細胞に分布するというリポソームの本来の傾向を利用する。一方、能動的標的化では、モノクローナル抗体、糖、糖脂質、蛋白質など特定のリガンドにリポソームを結合させること、又は本来の所在位置以外の器官及び細胞型を標的させるためにリポソームの組成又は大きさを変更することによってリポソームを改変する。
【0059】
標的化送達系の表面は、様々な方法で改変してよい。リポソームによる標的化送達系の場合は、リポソームの二重層と安定に結合した状態で標的リガンドを維持するために、脂質類をリポソームの脂質二重層に組み込むことができる。標的リガンドに脂質鎖を結合させるために、様々な結合基を使用できる。
【0060】
これらの技術(及び薬物送達を容易にするために通常使用される他の技術)は、当業者によって過度に実験をすることなく、本発明の方法で使用するためのSERCA2ポリヌクレオチドの調製に適合されることができると予想される。特に、上記のパラグラフで論じた手法は、発明者の知る限りでは、SERCA2ポリヌクレオチドをin vivoで筋細胞に送達するためにこれまで使用されていないが、この目的に使用するのに適していると考えられる。このため、上記に挙げた参照は、本発明の方法に必須ではないが、参照により本明細書に組み込む。それが適していることを示す特定の実施例を、以下に示す。
【0061】
D.心臓のSERCA2活性をIN VIVOで増大させるための方法
本発明では、SERCA2ポリヌクレオチドが、それがコードする生物的に活性なペプチドを発現させるのに十分な用量で供給されれば十分である。実現される発現レベルが、「通常の」内在性SERCA2の活性を実質的に補充するのに十分であることが好ましい。SERCA2ポリヌクレオチドが、組換え発現ベクター、好ましくはAAVベクターに含まれ、(セクションCで前述したように)製薬上許容される組成物に配合されると有利である。
【0062】
「通常の」SERCA2レベル及び活性は個体間で様々であり、従って、特定の種について絶対的に決定することはできない。しかし、SERCA2蛋白質の治療前後のレベル、並びにCHF心臓における増強された収縮性及び心臓性能の臨床的兆候をモニターすることによって、特定の治療目的を達成するために望ましいSERCA2発現レベル(「治療上有益な量」)を確認し、それを許容される臨床制限内に維持することができる。SERCA2レベルをモニターする手段は、以下のセクションFに説明する。
【0063】
心臓性能、及びCHFに伴うストレスに対する適応力の改善の臨床的兆候は、心臓病分野の当業者には公知であり、例えば、血流、心臓のポンプ容量、及び心室圧(例えば、血管造影及び心エコー検査による)、カルシウム輸送速度(心臓液試料のin vitro評価による)、耐性試験(例えば、心臓への圧力過負荷なストレスに対する心拍のモニタリングによる)、及びCHF症状の減退の一般的な臨床的兆候(例えば、宿主の持久力が増え、呼吸が容易になる)をモニターすることによって、判定することができる。投与用量は、個々の治療目的を達成するために変更してよい(例えば、急性のCHF病態の宿主においてカルシウム輸送を増大するためのSERCA2の過剰発現)。最大及び最小範囲は、前述のモデル及び大型の哺乳動物種の使用などから得られるヒト以外の動物データの結果を外挿することによっても決定される。
【0064】
SERCA2ポリヌクレオチド送達は、静脈内又は冠動脈内輸注法により(好ましくは、ポリヌクレオチドの大動脈起始部への逆流を最小限にするためにカテーテル法を使用して)行うことが好ましい。或いは、より強くより即時的なSERCA2発現を実現するために、SERCA2ポリヌクレオチドを(例えば、開胸技術を使って)心室壁に注入しても、(例えば、血管造影カテーテル法によって)直接心室に導入してもよい。その後、SERCA2発現をモニターし、必要なだけ送達を繰り返す。処理した宿主を、ポリヌクレオチド又はSERCA2に対する免疫応答などの副作用、及び(例えば、拡張期の過度の延長によって示されるような)過度のSERCA2発現がないかどうかについても注意深くモニターすべきである。
【0065】
しかし、通常、以下の実施例で記載する結果に基づくと、in vivoで筋細胞にSERCA2ポリヌクレオチドをAAV送達することに関係した細胞変性又は他の有害な影響を生じる危険性は低いようであり、野生型のベクターの混入物を排除すること及び他の方法によって(リポソームによるベクターのカプセル化などによって)ベクターの免疫原性を低減させることによって最小限に抑えることができる。
【0066】
E.本発明の方法を試験するための動物モデル
ラットが、その心臓には側副循環がないが、ヒトCHFの状態を満足できる程度に予測できる、うっ血性心不全の再現性のある実験モデルであることが明らかになった。特に、冠動脈の外科的結紮を受けたラットは、ヒトの心筋梗塞後のCHFの特に良いモデルである。CHF動物モデルとしてのラットを作製及び使用するための実験プロトコールは、当技術分野で十分に記載されており、参照文献については、当業者なら、Pfieffer他、Am.J.Med.、76:99〜103、1984年;Johns及びOlsen、Ann.Surg.、140:675〜682、1954年;並びに、Selye他、Angiology、11:398〜407、1960年(その開示内容を、CHF動物モデルの開発及び使用に関する当技術分野の知識を例示するために参照により本明細書に組み込む)を参照できよう。さらに、甲状腺の状態が減退したマウス、又は心臓の収縮性が低減しカルシウムトランジェントが遅くなっている甲状腺機能低下マウスも、再現性のある実験モデルとして使用することができる。
【0067】
さらに、本発明の方法によってSERCA2活性を増大させたCHF心臓における心臓性能への効果を特に予測するトランスジェニック動物モデルが開発された。これらの(SERCA2導入遺伝子を発現する)トランスジェニック動物を複製するのに有用なプロトコールを以下に記載し、実施例1で説明する。このプロトコールは、発現可能な導入遺伝子を哺乳動物に導入するための従来技術におおむね従う。当技術分野の当業者なら、これらの適用例を熟知しており、過度に実験をすることなく本発明の状況でそれらの技術を適用することができよう。
【0068】
例えば、様々な発達段階の胚性の標的細胞を、導入遺伝子を導入するのに使用することができる。胚性の標的細胞の発達段階に応じて様々な方法が使用される。受精卵は、マイクロインジェクションの最も良い標的である。マウスでは、オスの前核の大きさは直径約20マイクロメートルに達するので1〜2plのDNA溶液を複製可能に注入することができる。遺伝子移入の標的として受精卵を使用することは、多くの場合注入されたDNAが第一卵割の前に宿主遺伝子に組み込まれることになるという点で、大きな利点がある(Brinster他、Proc.Natl.Acad.Sci.USA、82:4438〜4442、1985年)。その結果、トランスジェニック非ヒト動物の全細胞が、組み込まれた導入遺伝子を持つことになる。これはまた、通常、生殖細胞の50%がその導入遺伝子を含むことになるため、初代の子孫への導入遺伝子の効率的な伝達として反映されることにもなる。受精卵のマイクロインジェクションは、本発明を実施する際に導入遺伝子を組み込むための好ましい方法である。
【0069】
レトロウイルス感染も、非ヒト動物に導入遺伝子を導入するのに使用することができる。発達途中の非ヒト胚は、胚盤胞段階までin vitroで培養することができる。この間、割球をレトロウイルス感染の標的とすることができる(Jaenisch、Proc.Natl.Acad.Sci USA、73:1260〜1264、1976年)。酵素処理によって透明帯を除去することにより、割球の効率的な感染が得られる(Hogan他、Manipulating the Mouse Embryo、Cold Spring Harbor Laboratory Press、Cold Spring Harbor 、米国ニューヨーク州、1986年)。導入遺伝子を導入するのに使用するウイルスベクター系は、通常、この導入遺伝子を含む複製欠損型レトロウイルスである(Jahner他、Proc.Natl.Acad.Sci.USA、82:6927〜6931、1985年;Vander Putten 他、Proc.Natl.Acad.Sci USA、82:6148〜6152)。ウイルス産生細胞の単層上で割球を培養することによって、容易に且つ効率的にトランスフェクションが得られる(前述のVan der Putten;Steward他、EMBO J.、6:383〜388、1987年)。
【0070】
或いは、もっと後の段階で感染を実施してもよい。ウイルス又はウイルス産生細胞を分割腔に注入することができる(Jahner他、Nature、298:623〜628、1982年)。組込みはトランスジェニック非ヒト動物を形成する細胞の一部分でしか起こらないため、初代のほとんどは、導入遺伝子についてモザイク状となる。さらに、初代は、通常子孫に分かれていくゲノムの様々な位置に、導入遺伝子の様々なレトロウイルス挿入物を含むことができる。さらに、効率は低いが、妊娠中期の胚に子宮内でレトロウイルスを感染させて生殖細胞系に導入遺伝子を導入することも可能である(前述のJahner他、1982年)。
【0071】
導入遺伝子を導入するための第3の標的細胞は、胚性肝細胞(ES)である。ES細胞は、着床前の胚から得られ、in vitroで培養し胚と融合される(Evans他、Nature、292:154〜156、1981年;Bradley他、Nature、309:255〜258、1984年;Gossler他、Proc.Natl.Acad.Sci USA、83:9065〜9069、1986年;及びRobertson他、Nature、322:445〜448、1986年)。導入遺伝子は、DNAトランスフェクション又はレトロウイルスの媒介による形質導入によって、ES細胞に効率的に導入することができる。その後に、これらの形質転換されたES細胞を非ヒト動物由来の胚盤胞に結合させることができる。ES細胞は、その後、胚に定着し、結果として得られるキメラ動物の生殖系の一部になる(総説については、Jaenisch、Science、240:1468〜1474、1988年を参照のこと)。
【0072】
本発明の状況で動物モデルとして使用するために、最適の導入遺伝子は、SERCA2の比較的高い発現レベルをもたらすことができるプロモーターを含むものが好ましい。この点で使用するのに好ましいプロモーターは、β−アクチンイントロンを含む、(例えば鳥類由来の)β−アクチンプロモーターに連結したヒトCMVエンハンサーである。導入遺伝子の発現活性を検出するために、前述したフラッグ抗原エピトープのコード領域を、SERCA2のC末端領域をコードする導入遺伝子の領域に含ませた。実施例1で説明するように、導入遺伝子を発現した初代動物の子孫は成体になるまで生存し、導入遺伝子が初代系列の少なくとも約15%の子孫に含まれていると予想できる。
【0073】
F.SERCA2の発現をIN VIVOでモニターする方法
SERCA2の発現をモニターするために、本発明に従って宿主に導入されるSERCA2ポリヌクレオチドを、既知のレポーター遺伝子を含むように改変してよい。例えば、Norton他、Mol.Cell.Biol.、5:281、1985年に記載のpRSV lac−Z DNAベクターは、蛋白質発現とともにβ−ガラクシターゼを産生する。ルシフェラーゼ及びクロラムフェニコールアセチル基転移酵素(「CAT」;pRSV−CATプラスミドの構築については、例えば前述のGorman他を参照のこと)も使用できる。別の有用なレポーター分子は、免疫測定法で容易に検出することができるフラッグ抗原性ペプチドである。フラッグ抗原性ペプチドの8個のアミノ酸配列及びそれをコードする領域は、当技術分野で既知である。この点に関して参照するためには、当業者なら、Chiang他、Peptide Res.、6:62〜64、1993年を参照できよう。(例えば、開始コドンから約15塩基対の位置に遺伝子を挿入することにより)コード用のレポーター遺伝子をSERCA2蛋白質のC末端に挿入しても、SERCA2触媒活性は妨害されない。このようなレポーター遺伝子の発現を検出する手段は当技術分野で公知であり、詳細には記載しないが、以下に要約する。
【0074】
例えば、本発明に従ってSERCA2ポリヌクレオチドを導入した後にin vivoで発現されたSERCA2を、免疫測定法によって検出することができる。この免疫測定法では、SERCA2蛋白質を液相中で利用し又は固相担体に結合させることができる。さらに、これらのアッセイで利用するSERCA2蛋白質は、様々な方法で検出可能に標識することができる。さらに、SERCA2又はレポーター遺伝子産物に対する抗体を、血液や血清などのアッセイ試料中でのSERCA2ポリヌクレオチド発現を検出するために利用することもできる。
【0075】
簡単に言うと、このような抗体は、当技術分野で公知の手段によって調製することができる。例えば、SERCA2又はレポーター遺伝子産物に特異的な抗体は、抗原性のSERCA2又はレポーター遺伝子ペプチドで非ヒト生物を免疫することによって調製することができる。このようなペプチドは天然の供給源から単離することができ(例えば、総説としてその開示内容を本明細書に組み込んだPopovich他、Am.J.Physiol、261:E377〜E381、1991年に報告されているラットSERCA2を単離するのに使用された方法を参照のこと)、或いは、α−アミノ基のt−BOC又はFMOC保護などの通常使用される方法により、過度に実験をすることなく合成することもできる。
【0076】
後者の方法は、ペプチドのC末端から出発して、各段階で1つのアミノ酸が加えられる段階的合成である(Coligan他、Current Protocols in Immunology、Wiley Interscience、991、ユニット9を参照のこと)。この場合に使用するペプチドは、Merrifield、J.Am.Chem.Soc.、85:2149、1962年、並びにStewart及びYoung、固相ペプチド合成法(Solid Phase Peptides Synthesis)(Freeman、サンフランシスコ、27〜62、1969年)に記載されているものなど様々な公知の固相ペプチド合成方法により、ポリマー1g当たり0.1〜1.0mMolのアミンを含む共重合体(スチレン−ジビニルベンゼン)を使用して、合成することができる。
【0077】
化学合成が完了するとすぐに、0℃で約1/4〜1時間、フッ化水素酸10%アニソール液で処理することにより、ペプチドを脱保護しポリマーから切断することができる。試薬を蒸発させた後、ペプチドを1%酢酸溶液でポリマーから抽出し、次いで凍結乾燥させて未精製物質を得る。通常、溶媒として5%酢酸を使用するSephadex G−15を用いたゲル濾過などの技術によってこれを精製することができる。カラムの適切な画分を凍結乾燥すると、均質なペプチド又はペプチド誘導体が得られ、これらは、次いで、アミノ酸分析、薄層クロマトグラフィー、高速液体クロマトグラフィー、紫外吸収分光法、モル旋光度、溶解度などの標準の技術により特徴を明らかにし、固相エドマン分解法によって定量することができる。目的のペプチドの抗原性は、そのペプチドで免疫された動物の抗体反応の大きさを判定する従来技術によって決定することができる。
【0078】
抗原性のSERCA2ペプチドを調製した後で、(ウサギ、マウス、又はラットなどの)哺乳動物にペプチドを導入することにより、免疫ペプチドに対する抗体を作製する。複数回注入による免疫化プロトコールが、抗原性ペプチドで動物を免疫する際に使用するのに好ましい(例えば、Langone他編、「小用量の免疫原生物による抗血清の調製:皮内複数回注入(Production of Antisera with Small Doses of Immunogen:Multiple Intradermal Injections)」、Methods of Enzymology(Acad.Press、1981年を参照のこと)。例えば、フロイント完全アジュバントに乳濁させた1mgの抗原性ペプチドを皮内注射し、その数週間後にフロイント不完全アジュバント中の同じ抗原を1回又は複数回追加することにより、通常、ウサギで良い抗体反応が得られる。
【0079】
望むなら、免疫ペプチドは、当技術分野で公知の技術を使用して結合することにより、キャリア蛋白質に結合させてよい。ペプチドに化学的に結合されるこれらの普通に使用されるキャリアには、キーホールリンペットヘモシアニン(KLH)、チログロブリン、ウシ血清アルブミン(BSA)、及び破傷風トキソイドが含まれる。次に、結合されたペプチドを使用して動物(例えばマウス又はウサギ)を免疫する。SERCA2は、現在、哺乳動物種間でかなり良く保存されていると考えられているので、SERCA2蛋白質の免疫原性を増大させるためにキャリア蛋白質を使用することが好ましい。
【0080】
動物に産生されたポリクローナル抗体を、例えば、抗体が産生された対象のペプチドが結合されているマトリクスに結合させ、マトリクスから溶出させることによって、さらに精製することができる。当業者なら、ポリクローナル抗体、並びにモノクローナル抗体の精製及び/又は濃縮に関する免疫学分野では普通の様々な技術について知っていよう(例えば、Coligan他、ユニット9,Current Protocols in Immunology、Wiley Interscience,1991年を参照のこと)。
【0081】
モノクローナル抗体は、特異性があり調製が容易であるため、SERCA2発現の検出に使用するのに好ましい。モノクローナル抗体を調製するためには、マウス又はラットの免疫が好ましい。本発明で使用する「抗体」という用語は、無傷の分子、並びに、例えば、エピトープ決定基を結合することができるFab及びF(ab’)2’などのその断片も含むものとする。また、この文脈では、「本発明のmAb’s」という用語は、SERCA2又はレポーター遺伝子産物に特異的なモノクローナル抗体を意味する。
【0082】
モノクローナル抗体(「mAb’s」)を分泌するハイブリドーマの調製に使用する通常の方法は、公知である(Kohler及びMilstein、Nature、256:495、1975年)。簡単に言うと、Kohler及びMilsteinに記載のように、この技術は、外科的検体から得、プールし、次いでSHFP−1に融合させた、黒色腫、奇形癌、子宮頸癌、神経膠腫、又は肺癌である5名の別々の癌患者の排出性(draining)所属リンパ節から単離したリンパ球を含む。ハイブリドーマは、癌細胞系に結合する抗体を産生するかどうかでスクリーニングした。
【0083】
mAb’sの抗原特異性の確認は、(酵素結合免疫測定法、即ち「ELISA」などの)比較的よく使うスクリーニング技術を使用して目的のmAbの基本的な反応パターンを判定することによって、実現できる。あるmAbを評価して、本発明のmAbと同じ特異性を有するかどうか判定することも、前述のように単離した目的の抗原に本発明のmAbが結合するのを試験対象のmAbが妨げるかどうか判定することによって、過度に実験をすることなく、可能である。本発明のmAbによる結合の低減により試験対象のmAbが本発明のmAbに競合することが示される場合は、それら2種のモノクローナル抗体が同じ或いは密接に関係するエピトープに結合する可能性が高い。
【0084】
あるmAbが本発明のmAbの特異性を有するかどうか判定するさらに別の方法では、本発明のmAbを通常反応性の抗原と一緒にプレインキュベートし、試験対象のmAbがその抗原に結合する能力を阻害されるかどうか判定する。そのとき試験対象のmAbが阻害される場合は、高い確率で、本発明のmAbと同じ或いは密接に関係するエピトープ特異性がある。
【0085】
MHCα及びMHCβに共通するエピトープを認識するSERCA2モノクローナル抗体は、検出するSERCA2の構造が本質的に改変されていないかどうか判定するのに特に有用である。このようなmAb’sは、その開示内容を総説及び参考文献として使用するために本明細書に組み込んだ、Dorn他、Am.J.Physiol.、267:H400〜H405、1994年に、再現するのに十分な詳しさで記載されている。
【0086】
SERCA2発現を検出するために使用できる免疫測定法の例は、直接式又は間接式の競合及び非競合免疫測定法である。このような免疫測定法の例は、放射性免疫測定法(RIA)、サンドイッチ法(イムノメトリックアッセイ(immunometric assay))、及びウェスタンブロット法がある。SERCA2又はレポーター遺伝子産物に結合する抗体の検出は、生理試料の免疫組織化学的分析を含めて、順方向、逆方向、又は同時に両方向に作動する免疫測定法を利用して実施することができる。lac−Z又はβ−gal(例えば、β−ガラクトシダーゼ)などのレポーター遺伝子産物を検出するための市販の分析キットが、この点で、使用するのに特に好ましい。
【0087】
使用する抗原又は抗体の濃度は、免疫測定法のタイプ及び使用する検出可能な標識の性質に応じて変わる。しかし、使用する免疫測定法のタイプにかかわらず、利用する抗原又は抗体の濃度は、当業者が常法の実験を実施して容易に決定することができる。
【0088】
このような抗原又は抗体は、多数の様々なキャリアに結合し、それに特異的に反応する抗原又は抗体の存在を検出するのに使用することができる。公知のキャリアの例には、ガラス、ポリスチレン、ポリ塩化ビニル、ポリプロピレン、ポリエチレン、ポリカーボネート、デキストラン、ナイロン、アミラーゼ、天然及び修飾セルロース、ポリアクリルアミド、アガロース、並びに磁鉄鉱が含まれる。本発明では、キャリアの性質は可溶性でも不溶性でもよい。当業者なら、これに関して、使用するのに適した他のキャリアについて知っているか、常法の実験を実施してそれらを確認することができよう。
【0089】
当業者には既知の多くの様々な標識及び標識方法がある。本発明で使用できる標識のタイプの例には、酵素、ラジオアイソトープ、コロイド金属、蛍光性化合物、化学発光化合物、及び生物発光化合物が含まれる。
【0090】
或いは、SERCA2ポリヌクレオチドは、その一般的技術を以下に要約する、当技術分野で既知の定量的ポリメラーゼ連鎖反応(PCR)プロトコールを使用して(好ましくは従来の心臓組織の生検技術を使用して得た標的細胞試料中で)検出することができる。
【0091】
精製又は未精製の任意の組織標本に由来する核酸を、それが標的核酸を含む特定の核酸配列を含む又は含むと推測されることを条件として、1種又は複数の出発核酸として利用することができる。従って、このプロセスは、例えば、メッセンジャーRNA(mRNA)を含めて、DNA又はRNAを使用してよく、DNA又はRNAは1本鎖でも2本鎖でもよい。RNAを鋳型として使用する場合は、その鋳型をDNAに逆転写するのに最適な酵素及び/又は条件を利用することになる。さらに、それぞれ1本の鎖を含むDNA−RNAハイブリッドも利用してよい。また、核酸の混合物を使用しても、以前の増幅反応で同じ又は別のプライマーを使用してここで作製した核酸をそのように使用してもよい。増幅するヌクレオチド配列は、大分子の一部分でもよく、或いは、別々の分子として最初に存在することもでき、従って、この特定の配列が核酸全体を構成する。増幅する配列は、最初に精製した形で存在する必要はなく、ヒトDNA全体に含まれるような複雑な混合物のごく一部分でよい。
【0092】
試料の標的ヌクレオチド配列が2本の鎖を含む場合、鋳型として使用できるように核酸の鎖を分離することが必要である。鎖の分離は、別々のステップとして又はプライマー伸張生成物の合成と同時に実施することができる。この鎖の分離は、物理的、化学的、又は酵素的手段を含めて、様々な適切な変性条件を使用して実現することができる。「変性させる」という語は、これら全ての手段を含む。核酸の鎖を分離する1つの物理的方法は、核酸を変性するまで加熱するものである。通常の加熱変性は、約1〜10分間、約80〜105℃の温度を必要とする。鎖の分離は、ヘリカーゼとして知られている酵素群のうちの1つの酵素又はヘリカーゼ活性のある酵素RecAにより、DNAを変性することが知られているリボATPの存在下で誘導してもよい。ヘリカーゼを用いた核酸鎖の分離に適した反応条件は、Kuhn Hoffmann−Berling(CSH−Quantitative Biology、43:63、1978年)に記載されており、RecAを使用する技術はC.Radding(Ann.Rev.Genetics、16:405〜437、1982年)に総説がある。
【0093】
増幅する標的核酸を含む核酸が一本鎖の場合、1種又は2種のオリゴヌクレオチドプライマーを加えることにより、その相補鎖が合成される。単一のプライマーを利用する場合は、プライマー、重合用作用物質、及び後述する4種のヌクレオシド3リン酸の存在下で、プライマー伸長生成物を合成する。この生成物は、一本鎖核酸に相補的なものとなり、一本鎖核酸にハイブリダイズして、長さが不揃いな鎖の二重鎖を形成する。次にこの二重鎖を一本鎖に分離させて、一本に分離された相補鎖2本を生成することができる。或いは、2種のプライマーを一本鎖核酸に加え、記載したように反応を実施してもよい。
【0094】
核酸の相補鎖を分離すると、その核酸がもともと二本鎖であったか一本鎖であったかにかかわらず、分離された鎖は、追加の核酸鎖を合成するための鋳型としていつでも使用できる。この合成は、プライマーが鋳型にハイブリダイズできる条件の下で実施する。通常、合成は好ましくはpH7〜9、最も好ましくはpH8の緩衝水溶液中で実施する。過剰モル(ゲノム核酸の場合、通常、プライマーと鋳型の比が約108対1)の2種のオリゴヌクレオチドプライマーを、分離された鋳型鎖を含む緩衝液に加えることが好ましい。しかし、本発明のプロセスを診断で適用するのに使用する場合は、相補鎖の量が分からない可能性があり、従って、相補鎖に対するプライマーの量を確実に決定することはできない。しかし、増幅しようとする配列が複雑で長い核酸鎖の混合物に含まれるとき、実用的には、加えるプライマーの量は通常、相補鎖(鋳型)の量に対して過剰モルである。プロセスの効率を高めるためには、大過剰モルが好ましい。
【0095】
いくつかの増幅実施形態では、基質、例えばデオキシリボヌクレオチド3リン酸のdATP、dCTP、dGTP、及びdTTPを適量、プライマーと別に又は一緒に、合成混合物に加え、得られる溶液を約1〜10分間、好ましくは1〜4分間、約90〜100℃に加熱する。この加熱期間の後、溶液をプライマーのハイブリダイズにとって好ましい室温まで自然冷却させる。冷却した混合物に、プライマー伸長反応を実施するのに適した作用物質(本明細書では「重合用作用物質」と呼ぶ)を加え、当技術分野で既知の条件下で反応を起こさせる。重合用作用物質は、熱安定性である場合は、他の試薬とともに加えてもよい。この合成(即ち増幅)反応は、室温から、最高ではそれを超えると重合用作用物質が機能しなくなる温度までで行うことができる。従って、例えばDNAポリメラーゼが作用物質として使用される場合は、通常、温度は約40℃以下である。
【0096】
重合用作用物質は、酵素を含めて、プライマー伸長生成物の合成を実現するように機能するどんな化合物又は系でもよい。この目的に適した酵素には、例えば、大腸菌(E.coli)DNAポリメラーゼI、Taqポリメラーゼ、大腸菌(E.coli)DNAポリメラーゼIのクレノウ断片、T4 DNA ポリメラーゼ、他の入手可能なDNAポリメラーゼ、ポリメラーゼ突然変異蛋白質、逆転写酵素、リガーゼ、及び、熱安定性酵素(即ち、変性を引き起こすのに十分な高い温度下に置かれた後でも、プライマー伸長を行う酵素)を含めて、他の酵素が含まれる。適切な酵素を使用すると、ヌクレオチドの正しい組み合わせがうまく進み、各変異ヌクレオチド鎖に相補的なプライマー伸長生成物が形成される。通常、合成は各プライマーの3’末端で開始され、合成が終わるまで鋳型鎖に沿って5’方向に進行し、様々な長さの分子を生成する。しかし、前述したのと同じプロセスを使用して、5’末端で合成を開始し、別の方向に進行する重合用作用物質があるかもしれない。
【0097】
新しく合成された変異ヌクレオチド及びその相補的な核酸鎖は、前述のハイブリダイズする条件下で二本鎖分子を形成し、このハイブリッドをプロセスの後続のステップで使用する。次のステップでは、新しく合成された二本鎖分子を前述した手順のいずれかを使用して変性条件において、一本鎖分子を生成させる。
【0098】
上記のプロセスを一本鎖分子に対して繰り返す。反応が先に規定した条件下で進行するように、必要なら、追加の重合用作用物質、ヌクレオシド、及びプライマーを加えてよい。やはり、合成各オリゴヌクレオチドプライマーの一端で開始され、鋳型の一本鎖に沿って進行し、追加の核酸を生成する。このステップの後、伸長生成物の半分は、2種のプライマーが結合した特定の核酸配列からなるはずである。
【0099】
変性及び伸長生成物合成のステップは、標的の変異ヌクレオチド配列を検出に必要な量まで増幅するのに必要なだけ繰り返すことができる。生成される変異ヌクレオチド配列の量は、指数関数的に増加する。
【0100】
増幅した生成物は、放射性プローブを使用せずに、サザンブロット分析により検出することができる。このようなプロセスでは、例えば、標的ヌクレオチド配列を極めて低いレベルで含むDNAの少量のサンプルを増幅し、サザンブロット法により分析する。シグナルが高レベルで増幅されると、非放射性プローブ又は標識の使用が容易になる。
【0101】
定量的PCRを実施するための好ましい方法は、1種又は複数の塩基対に誘導された突然変異を含み、その結果、配列及び大きさが標的の鋳型遺伝子と異なる競合物になった競合鋳型を使用して実施する競合的PCR法である。プライマーのうちの1種をビオチン標識し、或いは、好ましくは、結果として得られるPCR生成物の一方の鎖(通常アンチセンス鎖)を、アミノ−カルボキシル、アミノ−アミノ、ビオチン−ストレプトアビジン、又は他の適度に緊密な結合によって、適切な反応物に緊密に結合された固相の支持体に固定化できるように、アミノ化する。PCR生成物、固相の支持体、及び反応物の間の結合が共有結合であり、従って、変性条件下で結合を解けにくいものにすることが最も好ましい。
【0102】
PCR生成物のアミノ化又はビオチン標識した鎖を固定化した後、結合していない相補鎖をアルカリ性の変性洗浄液中で分離し、反応環境から除去する。標的に対応する配列特異的なオリゴヌクレオチド(「SSO’s」)及び競合核酸を検出タグで標識する。次に、SSOを、除去された結合していないセンス鎖との競合が無い状況でアンチセンス鎖にハイブリダイズさせる。適切な分析試薬を加え、使用した検出タグ及び固相支持体の手段に適したELISA測定法、好ましくはELISAマイクロプレートリーダーによって、ハイブリダイゼーションの程度を測定する。標的鋳型及び競合鋳型を含む鋳型を増幅するPCR反応から別々に得られた標準曲線を用いて測定値を比較して、標的の核酸含有量を導出する。
【0103】
この方法は、定量的で、PCRサイクルの数によって変わらず、SSOプローブとPCR生成物の相補鎖との競合に影響を及ぼされないという点で有利である。
【0104】
或いは、重合ステップ及びハイブリダイゼーションの全ステップを固相の支持体上で実施することもできる。この方法では、固相支持体上に捕捉されるのは、PCR生成物の鎖ではなく、ヌクレオチド重合プライマー(好ましくはオリゴヌクレオチド)である。次に、標的及び競合核酸PCR生成物を、溶液の状態で固相支持体に加え、重合ステップを実施する。重合生成物の結合されていないセンス鎖を、前述の変性条件下で除去する。
【0105】
標的核酸と競合核酸の比を、適切な測定手段(好ましくはELISAリーダー)を使用して標識されたオリゴヌクレオチドSSOプローブを検出することによって決定することができる。この方法の効率は非常に高くできるため、重合ステップの連鎖反応が不必要になり、従って、この方法を実施するために必要な時間が短縮できる可能性がある。重合の最終生成物をハイブリダイゼーションのために反応管から固相支持体に移動させる必要がなく、従って、それらが損失又は損傷する潜在的可能性が限られてくるため、この方法の正確性も高くなっている。しかし、特定の試料にとって必要なら、PCRを使用して標的及び競合核酸を別々の反応管で増幅させ、続いて固相支持体上で最終の重合反応を実施してよい。
【0106】
結合したPCR生成物の形成を示す、当業者に既知の様々な検出可能のシグナルを提供することができる分子(標的及び競合PCR生成物の形成を示す別の色を呈する、標識したヌクレオチド発色団など)を、反応の最後の数サイクルの進行中に反応溶液に加えることができる。標的核酸と競合核酸の比は、ELISAによって又は他の適切な測定手段及び固定化したハイブリダイゼーションプライマーの3’末端に結合させた検出タグに反応する試薬によって、求めることができる。この方法は、従来の非競合的PCRプロトコールを実施することによって、(定量せずに)特定の遺伝子が試料中に存在するかどうか検出できるように適合させることもできる。
【0107】
当業者なら、前述の方法で使用するのに適したプライマーを選択する方法を知っているか、容易に確認できよう。前述の技術に関するさらに詳細な内容については、Kohsaka他、Nuc.Acids Res.、21:3469〜3472、1993年;Bunn他、米国特許第5,213,961号の開示内容、並びにInnis他、PCRプロトコール:方法と適用のガイド(PCR Protocols:A Guide to Methods And Applications)、Acad.Press、1990年を参照できよう。これらの開示内容は、定量的PCRプロトコールに関する当技術分野の状況を例示するためだけに、本明細書に組み込む。
【0108】
PCR、オリゴマー制限(Saiki他、Bio/Technology、3:1008〜1012、1985年)、対立遺伝子特異的オリゴヌクレオチド(ASO)プローブ分析(Conner他、Proc.Natl.Acad.Sci.USA、80:278、1983年)、オリゴヌクレオチドライゲーション分析(OLA)(Landegren他、Science、241:1077、1988年)などの、特定のDNA配列の検出に通常適用されるどんな方法によっても、前述したように検出したSERCA2ポリヌクレオチドをさらに、溶液の状態で又は固体支持体に結合させた後で、評価、検出、クローン化、配列決定などすることができる。DNA分析に関する分子技術の総説があり、当技術分野で公知である(Landegren他、Science、242:229〜237、1988年)。
【0109】
本発明を十分に説明してきたが、その実施を例示する実施例を以下に記載する。しかし、これらの実施例は、添付の特許請求の範囲によって定義される本発明の範囲を限定するものとみなされるべきではない。
【0110】
実施例において、略語「min」は分を、「hrs」及び「h」は時間を意味し、(mlなどの)測定単位は標準の略語によって示す。
【実施例1】
【0111】
AAVベクターによるSERCA2のクローニング及び発現並びにマウス心筋の心筋細胞機能の評価
SERCA2 cDNAを従来のクローニング技術を使用してアデノ随伴ウイルスベクター中にクローニングした。SERCA2を発現するアデノ随伴ウイルス又はGFPレポーター配列の構築は、基本的に以下のように実施した。
【0112】
アデノ随伴シャトルベクターをSamulski他、Journal of Virology 63:3822〜3828、1989年に記載されているプラスミドpSub201から誘導した。このプラスミドのXba I断片を、マルチクローニングサイト及びポリアデニル化シグナルの前にある572塩基対のヒトCMVエンハンサー/プロモーター断片に連結した。このプラスミドをAAV−シャトルと呼び、サイズは約5000塩基対である。
【0113】
3’末端でフラッグシグナルペプチドに融着させたラットSERCA2a、及びKpn I/Xba I酵素又はHind III/Xba酵素を使用してAAV−シャトル中にクローニングした緑蛍光蛋白質(GFP)のコード配列が、それぞれ報告されている(He他、J.Clin.Invest 100:974〜980、1999年)。組換えクローンをDNA配列決定により確認し、大量のプラスミドをCsCl大量調製(maxiprep)プロトコールによって精製した。プラスミドpDGはGrimm他によって記載されており(Human Gene Therapy 9:2745〜2760、1998年)、同様にCsCl精製した。
【0114】
純粋な感染性ウイルス粒子を生成するために、挿入されたcDNAを含むAAV−シャトルプラスミド及びpDGプラスミドを共に、CaPO4共沈法を使用して293T細胞にトランスフェクトした。各ウイルス調製物のために、サブコンフルエントな293T細胞を蒔いた直径15cmの50セル培養プレートを使用した。各15cmプレートに、18マイクログラムのシャトルプラスミド及び70マイクログラムのpDGプラスミドをトランスフェクトした。トランスフェクションの翌日に10%ウシ胎児血清を含む新鮮なDMEMに培地を変更し、細胞を、5%CO2、摂氏37度でさらに2日間インキュベートした。
【0115】
ウイルス粒子の回収は、2.5mlのDMEM中で各プレートから293T細胞をかきとることによって行った。次に、この細胞を遠心し、25mlのDMEM中に再懸濁させ、摂氏−80度で凍結させた。さらに2回の凍結融解サイクルの後、100マイクログラムのDNアーゼ I及びRNアーゼ Hを加え、懸濁液を摂氏37度で30分間インキュベートした。もう一度3000gで遠心した後、上清を0.5%デオキシコレートに加え、摂氏37度で30分間インキュベートした。次に、この溶液を、5マイクロメーター及び0.8マイクロメーターのシリンジフィルターに順番に通して濾過し、3mlのヘパリン−アガロース懸濁液とともに室温で1時間回転台の上で回転させた。次に、この懸濁液をガラスカラムの上に載せ、ヘパリン−アガロース樹脂を0.254M NaClを含むPBS25mlで洗浄した。洗浄後、ウイルス粒子を0.554M NaClを含むPBSで溶出し、タイター評価のために少量の画分で保存した。最高のタイターの画分をプールし、PBSで透析し、摂氏4度で保存した。クローニングスキーム及び適切な制限部位の略図を図4に示す。
【0116】
次に、甲状腺機能低下マウスの左心室壁にウイルスベクターを投与した。甲状腺の状態が減退したマウス又は甲状腺機能低下マウスは、心臓の収縮性が低減しカルシウムトランジェントが遅くなっている。アデノウイルスベクターを、5〜10μlのアリコートで送り込み、左心室壁の異なる5つの部分に注入した。
【0117】
標的の導入遺伝子を含むAAVの発現効率は一定で、当技術分野で公知である。これにより、心筋細胞の約50%で導入遺伝子が発現する。核コードβGalを発現するAAV又はアデノ随伴ウイルスをマウスの心臓の左心室自由壁に注入することによっても、同様の結果が得られた(図5)。ここでは、アデノウイルス又はAAV注入の2、4、6、又は8週間後に動物を屠殺した。マウスの心臓からのデータにより、AAVの強い発現は注入後6又は8週間持続するが、アデノウイルスの発現は有意に低減したことが示されている。
【0118】
SERCA2発現AAVベクターの注入4週間後に、いわゆるランゲルドルフ単離灌流装置で心臓の収縮性能を判定した。肺静脈を通して左心室に風船を挿入し、収縮機能、特に収縮期の圧力上昇又はdP/dtの最高値、拡張期の圧力低下又はdP/dtの最小値、及び収縮期全体の圧力を、対照としてGFP CREを発現するAAVを注入した心臓で判定した。甲状腺機能低下性マウスの心臓において、全ての収縮性周辺部で非常に有意な上昇が得られ、収縮機能が正常範囲に回復した。図6は、フラッグ標識したSERCA2を発現するAAVを注入した甲状腺機能低下性マウスから単離したマウス心臓の収縮性能についてのデータを示す。収縮期の収縮速度dP/dtの最大値は、GFP発現AAVを注入したマウスと比べて、有意に上昇した。同様に、拡張期の弛緩速度即ち−dP/dtは、AAV SERCAを注入された甲状腺機能低下性マウスで有意に上昇し、収縮期圧力も、SERCA2aを発現するAAVを注入したマウス心臓で有意に上昇した。
【実施例2】
【0119】
SERCA2発現トランスジェニック動物及び対照動物における圧力過負荷モデル及び心臓肥大モデル
CHF心臓の条件を模倣した条件下で導入遺伝子を含む動物におけるSERCA2の追加発現の効果を試験するために、性別、年齢、体重が同程度の動物群を処理して、心臓狭窄を誘発させ、腹大動脈狭窄を発症させ、食塩負荷させた。CHF心臓で顕著なこのような条件下で、これらの動物は心肥大(CH)を発症し、心臓への圧力過負荷(PO)に伴う心臓異常を起こす。導入遺伝子発現を心臓組織に対して特異的にするために、SERCA2 CMV/β−アクチントランスジェニック動物を使用した。
【0120】
簡単に言うと、このような動物を開発するために、実施例1で記載したSERCA2/AAVベクター構築体を使用して開発したトランスジェニック動物(体重約200〜250gのラット)及び対照の動物を腹腔動脈のレベルで腹大動脈を分離するために麻酔し、トランスジェニック動物では、ステンレススチール製のヘモクリップ(Edward Weck&Co.、ノースカロライナ州)を使用して腎動脈の上で大動脈を外科的に狭窄した。後者の手順により、大動脈管腔の大きさが約50%小さくなった。
【0121】
手術の約2日後から、大動脈を狭窄したトランスジェニック動物に、ゴマ油に懸濁させたデオキシコルチコステロンを1週2回筋肉内注射し(25mg/g体重)、飲料水中1%塩化ナトリウムを与えた。通常は手術後1〜2週間内に生じるCHを発症するかどうか、全ての動物を心臓超音波(心エコー検査)を用いてモニターする。さらに、これらの動物は、圧力過負荷をかけることにより急性の損傷期を経験させる。
【実施例3】
【0122】
トランスジェニック動物におけるCHF状態に対する適応反応
AAV/SERCA2構築体を使用して開発したトランスジェニック動物は、手術の6週間後に試験すると(SERCA2のmRNAレベルを検出して判定された)SERCA2遺伝子転写が有意に減少し、通常、10〜12週で心不全を発症し始める。SERCA2 mRNAレベル及び蛋白質レベルを、前述のノーザンブロット法及びウェスタンブロット法又は他の分析技術(例えば、やはり前述したPCR)によって決定した。トランスジェニック動物及び対照動物の(SRカルシウムトランジェントに関係している)酵素活性レベル及び左心室機能を下記に記載するように測定した。他のCHF動物モデル及び本発明の方法による治療を受けているヒトにおいて、以下に記載するプロトコールに従って、心臓性能に関する同じパラメータを測定することができる。
【0123】
心臓収縮機能の評価
A.心室の機能性を判定するための心エコー検査のプロトコール(in vivo)
心エコー図の撮像は、7Mhzで作動する2周波フェーズドアレイトランスデューサー、及び積算ドプラー出力機能を備えた「ACCUSON(登録商標)」128超音波コンソールを用いて、適切な動物モデルで実施する。心エコー事象のタイミングは、皮下電極から得られる同時の心電図記録と相関がある。撮像の深さは2cm、セクタ角度はフレームレートが50/秒になる60にセットする。パワー設定は75dBとする。2次元イメージを使用して、Mモード記録(1000ライン/秒で得られる)に適したカーソル位置を選択することができる。
【0124】
軽く麻酔した動物をうつ伏せに置き、胸骨近くの左の肋間隙を通して撮像を実施する。傍胸骨長軸図及び短軸図を得、ビデオテープに録画し、上述したように、2次元図を使用して心室の中央レベルで実施するMモード記録のためのカーソル位置を導く。全ての関連した測定値は、American Society of Echocardiography推奨の臨床的慣行に関する判断基準を使用して、2次元化したMモード記録から得ることができる。隔壁の厚さは、拡張末期における立ち上がりから立ち下がりまでを測定する。
【0125】
後壁心内膜は、収縮期の傾きの最も大きい線で特定され、拡張末期における後壁の立ち上がりから心外膜境界の立ち上がりまでを測定する。左心室拡張末期径(LVEDD)は、心室径が最大の時点で、心室中隔の立ち下がりから後壁の立ち上がりまでを測定する。左心室収縮末期径(LVESD)は、同じ判断基準を用いて、心室径が最小の時点で測定する。収縮機能の指標としての心室内径短縮率(FS)は、[(LVEDD−LVESD)/(LVEDD 0)]×100で算出する。
【0126】
B.試料液を抜き取り、心臓内の血行動態パラメータを決定するためのマイクロマノメータ付きカテーテルによる処置のためのプロトコール(in vivo)
マウスを例にすると、体重20〜30gの成体マウス(PO有り又は無しの対象及びトランスジェニック)をケタミン(100mg/kg、腹腔内投与(IP))及びキシラジン(5mg/kg、腹腔内投与(IP))の混合物で麻酔する。解剖顕微鏡下で、動物をうつ伏せにし、正中線頚部切開を行って気管及び頚動脈を露出させる。次に、20ゲージの鈍針を、総(tital)容量0.2ml、呼吸数100/分のボリュームサイクル方式げっ歯類用人工呼吸器(ハーバードアパレイタス(Harvard Apparatus)社製)に連結されている気管カニューレを補助するように気管に通す。換気の妥当性を、胸部の膨張を目視検査して判定する。挿管後、大動脈圧を測定するために、1本の頚動脈に、液体入りカテーテル(管状になるよう火炎で延伸したポリエチレン(PE))付きカニューレを挿入する。次に、胸部を開き、2F高忠実度のマイクロマノメータ(ミラー(Millar)社)を僧帽弁から挿入して、LV中に取り付ける。
【0127】
(例えば、オンライン検出装置を使用して)LV収縮期圧力及び拡張期圧力、dP/dtの最大値及び最小値、並びに大動脈圧を継続的に測定して、血行動態パラメータをモニターする。データは、1500試料/秒の速度で得られ、高忠実度のカテーテルの周波数応答は10,000Hz超まではフラットである。マウスの心拍速度が速い(300〜500/分)ため、そのdP/dtを正確に測定するためにこれらの高いサンプルレートが必要である。
【0128】
血行動態パラメータのオフライン分析は、LV圧力低下の時定数(タウ(τ))の計算に加えて最小及び最大LV圧力並びに大動脈血圧の測定を含むことができる。特に拡張期弛緩の最初の等容性期を反映するタウ(τ)の値の決定は、SERCA2機能に密接に関連づけられている。
【0129】
C.単離した筋肉生検組織において筋細胞の短縮を判定するためのプロトコール(in vitro)
このプロトコールをラットで例示すると、切除による損傷を最小限にし拘縮を防ぐために25mM BDMを含む酸素化タイロード液中で左心室乳頭筋を切除し、受動的伸張を避ける。その端を短い5−0シルク縫合糸で結びつけて調製物の心室壁及び弁の端に付着させ、過剰伸張を防ぐ。筋肉の一方の端はCambridge400 Isometric force transducer(等尺性力トランスデューサー)(フルスケール5g、分解能100μg)のガラスロッドに付着し、もう一方の端はトランスデューサー台(Newport423、±1μm)上の小型のステンレススチールフックに付着するようにする。都合よく見られるように、1mlの試験浴をOlympus SZ45ステレオ顕微鏡の視野内におく。浴の温度を33℃に維持する。
【0130】
次に、静止時の力が最初にトランスデューサーチャネルに記録されるゆるんだ長さに筋肉を調整し、BDMを洗い流し、(例えば、Gras刺激装置を使用して)双極性の刺激用電極を浴中に沈める。筋肉を、0.2Hz(閾値より20%高い)で刺激し、30分後、灌流液を[Ca2+]0.5mM溶液から[Ca2+]2.0mM溶液に切り換える。全ての灌流液を100%O2でバブリングする。
【0131】
定常状態での等尺性張力の発生は、筋肉長の増加で測定し、発生した張力が最大になるLmaxに対する割合として表す。(結果をin vivoの筋肉組織性能に外挿する際に)使用する筋肉の長さが短いことを補正するために、張力のピーク時に短くなる歪みを、筋肉上に置いた対の表面細粒を使用して測定し、ビデオカメラで録画する。
【0132】
NIH Image 1.47を使用してData Translationビデオフレームグラッバーボード付きMacintosh Centris 650上でマーカー位置を検出しデジタル化することが好ましい。任意の固定した基準長に関する歪みを使用して長さの縮尺を補正すると、長さ−張力曲線の下降を打ち消すことが示された。
【0133】
収縮性は、等尺性の応力−歪み曲線の傾きで表され、このときの応力は発生した張力(静止期全体)を筋肉の断面積で割った値から計算する。この断面積は、機械的試験後に(その重量から決定した)標本の体積を、両端の間の無負荷の筋肉の長さで割ることにより推定する。細胞外Ca2+濃度0.5〜2.5mMの関数としてこれらのプロトコールを繰り返すことにより、これらの調製物の潜在的なCa2+感度を特徴付けることが可能になる。
【0134】
これらの実験のSERCA2機能及びEC結合をさらに特徴づけるために、Na/Ca交換がNaを含まないタイロード液で遮断されたとき、及び筋小胞体のカルシウム蓄積が10mMカフェインで遮断されたときの弛緩時間を測定することによって、SECRA2のポンプ機能を試験する。Na/Ca交換を測定すると、弛緩時間は約30%遅くなる。試験筋肉の弛緩特性は、チャートレコーダ(例えば、Gould 2200レコーダ)及び高速(40mm/秒)のマイクロコンピュータ取得システム(例えば、市販のStrawberry Tree)に記録された等尺性張力の追跡から、筋肉長とカルシウムの関数として決定される。測定されるパラメータには、ピーク張力から90%回復までの時間及び等尺性の力の低減の指数時定数が含まれる。筋節の長さが損傷されていない中央の筋肉を一定に保つように筋肉長が制御されると、等尺性弛緩が有意に延長されることが研究により示されているので、これらの測定値は前述のアーチファクトにも当てはまり、補正可能である。
【0135】
前述したのと同じ手法を用いて、解剖顕微鏡下でラットの左心室から単離した長くて薄い均質な梁柱を使用して、収縮の研究を実施することもできる。梁柱を使用して研究すると、従来の乳頭筋調製物からは得られない筋肉の一連の弾性があるため、拡張期収縮機序における絶対的変化に関する、アーチファクトとは無関係なより具体的な情報が得られる。左心室壁の他の部分領域を、辺縁検出及び細胞内Ca2+トランジェントの研究をするために心筋細胞を単離するのに使用できる。
【0136】
D.心筋細胞の単離及びカルシウムトランジェントレベルの評価のためのプロトコール(in vitro)
心筋細胞は、コラゲナーゼ灌流法を使用して成体ラットの心臓から調製することができる。この筋細胞は、4%ウシ胎児血清の不在下又は存在下で少なくとも4日間維持し、成体の心筋細胞の形態及び代謝的特徴を維持することができる。1匹の成体ラットの心臓から、9〜10x106個の生存可能な筋細胞を得ることができる。
【0137】
心筋細胞は、心室壁の小片から調製することもできる。これにより、筋細胞機能、乳頭筋機能、並びに同じ心臓から得られるSERCA2遺伝子発現及びSERCA2機能に関係するパラメータについて判断することが可能になる。このために、心室組織を細かく刻み、25mM NaHCO3、3.4mM MgCl2、30mMタウリン、及び2mMカルニチンを追加したCa2+を含まない改変Joklik最少必須培養液(MEM)中で洗浄する。次に、この組織を0.1% コラゲナーゼ タイプII(ワーシントン(Worthington)社)、0.1%BSAフラクションV、及び25μMCaCl2を含むMEMで消化する。連続的な酸素添加のもと、32℃で組織を穏やかに攪拌しながら、酵素消化を30〜40分間継続する。口径の大きな血清ピペットで穏やかに粉砕することによって、最終的に細胞を完全に分散させる。
【0138】
次に、細胞を300μMナイロンメッシュに通して濾過し、洗浄する。次に、カルシウムレベルを徐々にその場の濃度が1μMになるまで上昇させる。この手順により、最大2x106細胞/心臓を得ることができ、その60〜70%が以下に記載する研究で使用可能な生存可能な棒形の筋細胞である。
【0139】
単離した成体ラットの心筋細胞(非培養)を118mM NaCl、4.8mM KCL、1.2mM NaH2PO4、1.2mM MgSO4、1mM CaCl2、11mMグルコース、及び25mM Na−N−2−ヒドロキシエチルピペラジン−N−2−エタンスルホン酸を含む10mlの溶液中に懸濁させ、500μlの細胞懸濁液を1.5mlの普通のタイロード液に加え、Indo Iのアセトキシメチルエステル誘導体のDMSO中1mM溶液5μlを加える。細胞を室温で15〜20分間インキュベートする。次に、Indo I添加の均一性、及びきれいな横紋を伴う棒形細胞の形態に基づき細胞を選別する。石英光学素子を装備したNikon Diaphot顕微鏡の台上に取り付けた小型灌流チャンバーに細胞を移す。細胞を灌流チャンバーの底に自然沈降させ、90%O2−5%CO2でガス補給し37℃で維持したタイロード液を用いて5ml/分で連続的に灌流する。細胞を外部の白金電極を用いて0.2、0.33、1.0、2.0、及び5.0Hzで駆動し、Ca2+トランジェントを蛍光定量によって測定する。同様の研究を、新生児期のラット又はマウスの筋細胞を使用して実施することもできる。Adv SERCA2を新生児筋細胞に感染させると、カルシウムトランジェントが顕著に低減する。
【0140】
カルシウムトランジェントレベルに関しては、カルシウム2++トランジェントが最も低減するための時間は、SERCA2導入遺伝子を含む陽性の筋細胞において、in vitroで観察されるのと同様の程度まで有意に短縮された(実施例4、図3、p<0.05)。心室圧に関しては、左心室で得られた最大圧力(LVdP/dt最大値)は、トランスジェニック動物において有意に大きかったが、最小レベルの場合、心室の筋肉は、トランスジェニック動物において拡張期弛緩がより速くなる傾向を示した(n=3、コントロール(n=5)と比べて)。動物で検出したSERCA2 mRNA及び蛋白質レベルを図1及び図2に示す。これらのデータは、心筋のカルシウム輸送及び拡張期弛緩が、予測用動物モデルで本発明の方法を適用することにより、有利な影響を受けるという原理を証明している。
【実施例4】
【0141】
SERCA2発現アデノウイルスベクターでトランスフェクションした後の心筋細胞機能のIN VITROでの評価
簡単に言うと、新生児の心筋細胞をSERCA2アデノウイルスベクターでトランスフェクトし、実施例3で記載したようにカルシウムトランジェントを測定し、対照(トランスフェクトされていない)細胞と比べる。図3において、細胞#42は対照の新生児心筋細胞の挙動の平均を表し、細胞#27は、トランスフェクトした新生児心筋細胞の挙動の平均を表す(ADV)。図3で示されるように、カルシウムトランジェントの最大減少の半分までの時間は、対照細胞と比べてトランスフェクトした細胞で33±19%短縮された(n=4、p≦0.01)。これらのデータは、カルシウムトランジェントがSERCA2 ADVでトランスフェクトした筋細胞で増大され、その結果、in vivoで測定すれば、心筋でより速い拡張期弛緩が促進されるはずであるという原理の証拠を提供する。
【0142】
本発明のいくつかの実施形態を記載してきた。しかし、本発明の趣旨及び範囲を逸脱することなく様々に改変を加えることができることが理解されよう。従って、本発明は例示した特定の実施形態によってではなく、添付の特許請求の範囲によってのみ限定されることが理解されよう。
【図面の簡単な説明】
【0143】
【図1】(例として、アデノウイルスのSERCA2構築体を使用して開発した)マウスにおけるSERCA2導入遺伝子の発現を示したウェスタンブロットを示す図である。レーン1〜2はα−筋節アクチンの発現を示し、レーン3は、分子量マーカーを示し、レーン4〜5はSERCA2発現を示す。
【図2】SERCA2をコードする数種のアデノウイルス、すなわち、例えばCMVプロモーター(HH−1、HH−2、及びSAI−3)又はTKプロモーター(TK)の制御下にあるベクターなどのうちの1種でトランスフェクトしたマウス由来の心筋細胞におけるラットSERCA2 mRNAの存在を示したノーザンブロットを示す図である。対照の構築体は、SERCA2ポリヌクレオチドを含まなかった(ctrl)。( )内の数字は、各構築体のプラーク形成単位(pfu)の数を示し、L6細胞をアデノウイルス構築体で48時間感染させ、抽出し、細胞性RNA全体をゲル上で分離し、次いでSERCA2のcDNAをRNAにハイブリダイズさせるノーザン分析法によって決定した。
【図3】例として、アデノウイルスベクターでトランスフェクトした新生児の筋細胞において検出されたカルシウムトランジェント(平均値は細胞#27で表されている)及びトランスフェクトされていない新生児の筋細胞において検出したカルシウムトランジェント(平均値は細胞#42で表されている)を示す図である。カルシウムトランジェントは蛍光定量法で測定し、時間(x軸)の関数として表される。
【図4】例として、ベクターにSERCA2a配列を挿入するためのクローニングスキーム及び適切な制限部位を示す図である。具体的には、ラットSERCA2aを3’末端でフラッグシグナルペプチドに融着させたSERCA2a−フラッグが、ヒトCMVプロモーターのもと、XbaI/BamHI及びBgI II/KpnI制限部位に挿入される。
【図5】マウスの心臓の左心室自由壁にAAV又はアデノ随伴ウイルスを注入し、注入の2、4、6、及び8週間後に発現している核コードβgalを示す図である。
【図6】rAAV遺伝子送達4週間後に、単離して灌流させた心臓の機能測定値に対するSERCA2a−フラッグ発現の効果を甲状腺低下症マウスのGFP対照と比較して示す図である。*は、AAV−GFP処理したマウスと有意に(p<0.05)異なることを示す。rAAV−SERCA−フラッグ処理をしたマウスの機能が、AAV−GFP処理をしたマウスの機能より改善していることに留意されたい。
【技術分野】
【0001】
関連特許出願へのクロスリファレンス
本発明は、1995年11月4日出願の現在も係属中の米国特許出願第08/420,306号の一部継続出願である。
【0002】
1.発明の分野
本発明は、心筋収縮性を調節するための方法に関する。より詳細には、本発明は、SERCA2蛋白質を作動的にコードする遺伝子をin vivoで送達することによって、心臓の筋小胞体(SR)カルシウム2++ATPアーゼ(SERCA2)のin vivoのレベルを上昇させる方法に関する。
【背景技術】
【0003】
2.先行技術の変遷
うっ血性心不全は、米国において成人の主な死亡原因のうちの1つである。心虚血(心臓への血液供給が妨害され又は失われることにより生じる急性の事象)と比べて、うっ血性心不全は、心筋の収縮性及びストレスに対する心臓の適応性の漸進的な消失を伴う比較的潜行性の事象である。結局、有効な治療法はなく、CHF心臓は、身体の代謝要求を満たすのに十分な速度で血液をポンプ輸送する能力を失う。
【0004】
うっ血性心不全(CHF)に付随する心機能の異常は様々であるが、収縮性蛋白質の活性化に必要とされるCa2++イオンのSRからの放出の低減は、CHF症候群の共通の特徴である。この消失の意味は、カルシウム輸送が心臓の正常な機能活動において果たしている役割という文脈で最も良く理解することができる。
【0005】
簡単に言うと、SRは心筋の各筋原線維を取り囲む膜性構造物である。SERCA2は、SRの膜内部に含まれており、心筋の拡張期弛緩の間にSR内部空間に遊離カルシウムイオンの70〜80%を能動輸送する働きをする。輸送に利用可能な残存カルシウムイオンの多くは、SRのナトリウム/カルシウム交換輸送系、並びに程度ははるかに低いが、筋細胞膜カルシウムイオンATPアーゼが触媒するATP加水分解によって駆動される輸送、及びミトコンドリアのカルシウム取り込みによって、細胞質から取り除かれる(Bassani他、J.Physiol.、453:591〜608、1992年、及びCarafoli,E.、Ann.Rev.Biochem.、56:395〜433、1987年)。
【0006】
SERCA2のATP加水分解活性とSERCA2 mRNAの絶対レベルの双方がCHF心臓において低下することから(Hasenfuss他、Circ.Res.、75:434〜442、1994年、及びStuder他、Circ.Res.、75:443〜453、1994年)、CHF心臓の低圧力で血液を受け取る能力の損傷は、SERCA2媒介による、収縮を活性化するカルシウムイオンのSRへの輸送の遅れに直接関連づけられ、その結果心臓の拡張期弛緩が遅れると広く想定されている(例えば、Grossman,W.、N.Engl.J.Med.、325:1557〜1564、1991年;Lorell,BH、Ann.Rev.Med.、42:411〜436、1991年;及びArai他、Circ.Res.、74:555〜564、1994年を参照のこと)。これらの知見は、特にmRNAのSERCA2コードレベルの低下に関して、ヒト並びに他の哺乳動物種において確かめられている。(ヒトSERCA2 mRNAレベルについては、Arai他の上記論文、及びMercadier他、J.Clin.Invest.、85:305〜309、1990年を参照のこと。また、他の哺乳動物種の肥大心臓組織におけるSERCA2 mRNAレベルの低減については、Wang他、Am.J.Physiol.、267:H918〜H924、1994年(フェレット);Afzal及びDhella、Am.J.Physiol.、262:H868〜H874、1992年(げっ歯類);及びFeldman他、Circ.Res.、73:184〜192、1993年(げっ歯類)を参照のこと)。
【0007】
CHFにおけるカルシウム輸送の役割に関して近年関心が持たれているにもかかわらず、SERCA2カルシウム輸送活性の損傷の分子的原理は少ししか理解されておらず、CHFの治療レジメンにまだ利用されていない。そうではなくて、CHF症候群のための現在の治療様式は、CHF心臓の不全につながる機能異常を引き起こさないとしてもそれに付随すると考えられている生化学的事象及び分子的事象を直接標的としていないという意味で、ほとんど非特異的である。例えば、アドレナリン様薬物を投与することによるCHFの薬剤治療は、心筋収縮を刺激するが、筋肉の収縮性の減退を引き起こした根底にある状態を是正しない。
【0008】
従って、in vivoの心臓蛋白質レベルを補充及び/又は上昇させることは、ヒトのCHFの進行を治療及び制御するための魅力的な代替方法である。しかし、心臓組織に心臓蛋白質又は心臓ペプチドを導入することによってこの目標を実現することは、成功する可能性が低い。特に蛋白質に対する生体反応を起こすのに十分な用量で、潜在的な毒性を生じる危険性が主要な懸念である。実用的な観点からは、これらの蛋白質を単離し精製すること、又は合成することに伴うコストの問題もある。さらに、標的組織中に存在するプロテアーゼによって分解されるため、in vivoでの半減期が比較的短いことにより、これらの蛋白質の臨床上の効果も限られるはずである。
【0009】
これらの理由により、この蛋白質を発現する遺伝子の送達により、患者に蛋白質を導入することは、蛋白質それ自体の投与に代わる魅力的な代替方法である。この目的で、標的細胞に外来遺伝子を導入するための様々な戦略が開発されている。ヒトで使用するために今日までに提案されている遺伝子治療プロトコールの大半は、ex vivoの遺伝子移入に焦点を合わせている。例えば、標的組織に移植するための細胞のレトロウイルスによるトランスフェクションなどがある(例えば、Anderson,WF、Science,256:808〜813、1992年、及びMiller,AD、Nature、357:455〜460、1992年(アデノシン脱アミノ酵素欠損の治療)を参照のこと)。しかし、これらのプロトコールの有用性は、蛋白質発現が比較的非効率的であること並びに標的器官及び組織への到達性が限られていることにより、限定的であることが判明した。
【0010】
従って、in vivoの遺伝子送達方法は、当技術分野で関心の高いテーマである。この目的で、「裸の(naked)」ポリヌクレオチド(プラスミド)、ウイルスに連結したプラスミド、ウイルスに取り込ませた(cointernalized)プラスミドの導入、並びにリポソームによる遺伝子構築体のカプセル化及び送達を含めて、この目標を達成するためにいくつかの系が開発されている。
【0011】
例えば、リス肝炎の裸のクローン化プラスミドDNAをリスに肝臓内注入したところ、ウイルス感染及び抗ウイルス性抗体の形成が共にリスで生じたというNIH(米国国立衛生研究所)の研究が1984年に報告された(Seeger他、Proc.Nat’l.Acad.Sci USA、81:5849〜5852、1984年)。数年後、Felgner他は、骨格筋組織に注入した「裸の」ポリヌクレオチド(すなわち、リポソーム又はウイルス発現ベクターを伴わないDNA又はRNA)から蛋白質発現を得たことを報告した(Felgner他、Science、247:1465、1990年。また、PCT国際特許出願公開第90/11092号も参照のこと)。Felgner他は、筋組織が、多核細胞、筋小胞体、及び筋細胞の深部まで伸びる横行の管状系からなる独特な構造を有するため、筋細胞がポリヌクレオチドを効率的に取込み発現すると推測した。
【0012】
標的組織に直接遺伝子を送達するための同様の系が、Stribling他、Proc.Natl.Acad.Sci.USA、89:11277〜11281、1992年(リポゾームにカプセル化した遺伝子のエアロゾル送達後に検出された蛋白質発現)及びTang他、Nature、356:152〜154、1992年(ワクチン「銃」を用いてコロイド性の金粒に結合させたhGHプラスミドをマウスの皮膚に注入した結果、注入された遺伝子に対する免疫応答を誘発せずにhGH蛋白質を発現した)によって報告されている。筋細胞内で高レベルの蛋白質発現を起こすのに一般に有効ではあるが、長期治療のための筋組織へのDNA又はRNAの直接注入では、遺伝子の分解による発現の損失を補うために注入を繰り返し利用する必要がある。この手法は、時間がかかり高価であるだけでなく、注入部位及びその周辺で炎症が引き起こされるため、長期治療には非実用的であるかもしれない。従って、短期治療又は救急治療では有用であるが、標的組織に発現可能な遺伝子を導入するための侵襲性のより低い手段が、標的組織への直接注入よりも一般に好ましい。
【0013】
さらに、組換え発現ベクターを使用しないin vivoの遺伝子送達方法の大半は、標的細胞のトランスフェクションが非効率的で蛋白質発現が比較的弱いという欠点がある。従って、組換え発現ベクター(特に複製不能型のベクター)が現在もなおin vivoの遺伝子送達のための好ましい運搬体(vehicle)である。
【0014】
心筋細胞は、in vivo遺伝子送達に適した標的であることが示されている。例えば、CHFの治療に対する近年の1つの提案を用いれば、CHF心臓の筋細胞の活性β2−アドレナリン受容体の数が補充及び/又は増加されるはずである。マウスでの研究により、直接移植技術を使用してこれらの受容体をコードする遺伝子を導入したところ、外因性のアドレナリン供給源の不在下でさえ心筋の収縮性をin vivoで増大させたことが示されている(Lefkowitz他、Science、264:582〜586、1994年)。特に、アデノ随伴ウイルスベクターが、心筋細胞への遺伝子送達のための成功裡の運搬体として示されている(例えば、Guzman他、Circ.Res.、73:1202〜1207、1993年、及びMuhlhauser他、Circulation、88(Part2):1〜475、1993年を参照のこと)。
【発明の開示】
【発明が解決しようとする課題】
【0015】
本発明の好ましい実施形態の詳細を、添付図及び以下の説明に示す。本発明の詳細が知られれば、非常に多くのさらなる革新及び変更が当業者には明らかになるであろう。
【0016】
本発明は、SERCA2を作動的にコードするポリヌクレオチドを心筋などの組織に導入することにより触媒活性のあるSERCA2の発現をin vivoで高めることによって、心筋のSRへのカルシウム輸送を増大させる方法である。標的細胞のトランスフェクション効率の点で好ましい実施形態では、SERCA2をコードしているポリヌクレオチドを、複製不能型のウイルス組換え発現ベクター、最も好ましくはアデノ随伴ウイルスベクター(AAV)構築体を用いて心臓組織に送達する。
【0017】
或いは、SERCA2コード遺伝子を、リポソームによるカプセル化又は「裸の」ヌクレオチドの投与によって提供してもよい。SERCA2遺伝子の送達は、外科的に(即ち、ベクター又はトランスフェクトした細胞の標的組織への直接導入によって)又は冠動脈内輸注法によって行ってよい。
【0018】
SR中のSERCA2活性上昇によってもたらされるSRへのカルシウムイオン輸送の増大により、心臓の筋原線維収縮の活性化が緩和されることになる。従って、本発明の方法の特に有利な点は、特に心筋の拡張期弛緩の初期段階を短くすることにより、CHFに関連する異常に対して心臓が調整するのを助けることである。
【0019】
本発明の方法の一使用は、インターベンション治療(例えば移植)を待機している、且つ/又は既にCHF治療を受けている患者のCHFの短期改善を目的としている。本発明の方法の使用はまた、CHFに関連する心機能の異常を生じるリスクがある、或いは既に異常を生じ始めている患者に恩恵をもたらすと予想される。
【0020】
本発明の他の態様では、本発明の方法で使用するためのベクターを開発する際及び本発明の方法の適用を試験する際に使用するためのトランスジェニック動物が提供される。CHFの治療に使用したときの本発明の方法の有効性をモニターする方法も説明する。
【発明を実施するための最良の形態】
【0021】
この説明を通じて、ここに示す好ましい実施形態及び実施例は例とみなすべきで、本発明を限定するものとみなすべきではない。
【0022】
A.定義
以下の定義は、本発明の考察を簡単にするために提供される。しかし、これらの定義が本発明の正当な範囲又は趣旨から逸脱しない等価物を含むように拡張できることを、当業者なら理解するであろう。この理由から、これらの定義は本発明を限定するものと解釈されるべきではない。
1.「SERCA2ポリヌクレオチド」とは、DNA又はRNAを意味し、本発明に従って行う治療の目的に適したセンス鎖及びアンチセンス鎖を含むことができる。本明細書では、「ポリヌクレオチド」とは、分離した断片の形又は大きな構築体の成分としての、デオキシリボヌクレオチド又はリボヌクレオチドの重合体を意味する。ポリヌクレオチドの配列は、遺伝コードから推測できるが、コードの縮重を考慮しなければならない。本発明のポリヌクレオチドは、遺伝コードの結果として縮重した配列を含み、その配列は当業者によって容易に決定することができる。
2.「作動的にコードする」とは、所望の翻訳産物、例えばペプチド又は蛋白質の発現及び望むならその分泌に必要なプロモーター及び他の配列を含むように改変されたポリヌクレオチドを意味する。本発明の全ての実施形態は、公知の組換え発現ベクターを使用して実施することができる。これらのベクターは、所望の翻訳産物をコードするcDNAを含むことが好ましい。従って、文脈上別の意味で解釈する必要がない限り、「ポリヌクレオチド」とは、適切な組換え発現ベクター中に含まれ作動的にコードする配列を意味すると想定され、その例が本明細書で提供される。
3.「合成」とは、ポリヌクレオチド配列及びポリペプチド配列を合成する公知の手段を意味し、天然のポリヌクレオチド及び蛋白質の単離及び精製を含むことができる。
4.「ペプチド」とは、in vivoで所望の生体効果を有する小型ペプチド、ポリペプチド、オリゴペプチド、及び蛋白質を意味する。
5.「送達」とは、作動的にコードするポリヌクレオチドを宿主に導入する既知の方法を意味する。当業者なら、これらの送達手段を熟知しており、或いは容易に特定することができる。しかし、特に有用な送達手段に関しては、「新規の薬物送達系(Novel Drug Delivery Systems)」(Marcel Dekker、1992年)を参照することができ、その関連する開示内容を、薬物送達技術に関する当技術分野の知識状態を例示するために参照により本明細書に組み込む。
6.「宿主」とは、本発明に従って行われる治療の受容者を意味する。宿主はどんな脊椎動物でもよいが、好ましくは哺乳動物である。哺乳動物の場合は、宿主はヒトが好ましいが、飼育された家畜でもペット動物でもよい。
7.「標的組織」とは、作動的にコードするポリヌクレオチドの発現を得ようとする、宿主の組織を意味する。
8.「抗体」とは、任意のクラスの免疫グロブリン全体、キメラ抗体、2つ又は複数の抗原特異性を有するハイブリッド抗体、及びハイブリッド断片を含む断片を意味する。また、このような断片の結合体、例えば米国特許第4,704,692号に記載されているいわゆる抗原結合性蛋白質(単鎖抗体)、及び、例えば米国特許第4,699,880号に記載されている抗イディオタイプ抗体(他の抗体に結合する抗体)も、「抗体」の意味の範囲内に含まれる。
9.「組換え発現ベクター」は、真核生物又は原核生物で発現可能なポリペプチドを作動的にコードするポリヌクレオチドの系を意味する。真核生物又はウイルスの配列を有するDNA配列を原核生物で発現する方法は当技術分野で公知である。宿主中で発現及び複製できる、生物的に機能的なウイルス及びプラスミドDNAベクターも当技術分野で公知である。宿主としては、微生物、酵母、昆虫、及び哺乳類生物を挙げることができる。
【0023】
B.本発明の方法で使用するポリヌクレオチド構築体
ラットSERCA2のゲノムクローンのヌクレオチド配列(Rohrer他、J.Biol.Chem.、263:6941〜6944、1988年(ラットSERCA2 mRNA)も参照のこと。この論文で報告された配列を、参照のために本明細書に組み込む)。このクローンは、Rohrer他、J.Biol.Chem.、266:8638〜8646、1991年(その開示内容を参照のために本明細書に組み込む)に記載されているような従来のハイブリダイゼーション技術によって得られた(実施例1を参照のこと)。この論文は、クローンを転写するための開始コドン及び終止コドンも記載している。
【0024】
カルシウムATPアーゼのSERCA2アイソフォームのヌクレオチド配列(すなわち、横紋筋の「速筋型」アイソフォームに対する骨格筋の「遅筋型」アイソフォーム)は、哺乳動物種間で90%以上保存されている。従って、SERCA2は、ヒト(GENBANK #J4025、又、GENBANK M23114〜23116、M23277〜23279、及びLyttonとMacLennan、J.Biol.Chem.、263:15024〜15031、1988年も参照のこと)、及びウサギ(MacLennan,DH、Nature、316:697〜700、1985年、及びGENBANK M33834)を含めて、げっ歯類以外の哺乳動物の骨格筋組織からかなり容易に同定され、配列決定されている。当業者なら、ヒトSERCA2ポリヌクレオチドの使用がヒトの治療では大いに好ましいことを理解するであろうが、ラットSERCA2ポリヌクレオチドが、実施例で記載した実験のためには使用しやすく好ましい。
【0025】
SERCA2に関して詳細に本明細書に記載する本発明の方法は、CHF心臓でその発現及び活性が損傷されている他の心臓蛋白質を送達するために使用するように適合させることができるので有利であることも、当業者なら理解するであろう。特に、哺乳動物のカルシウム/ナトリウム交換体をコードしている遺伝子は、SERCA2ポリヌクレオチドの送達に関して本明細書に記載したのと同じ方法で、本発明の方法に従って心臓に送達するための特に魅力的な対象である。
【0026】
本開示に含まれるSERCA2配列及び当技術分野で公知のものを入手し使用するために、DNA及びRNAを当技術分野で公知の自動核酸合成装置を使用して合成することができる。公知のポリメラーゼ連鎖反応(PCR)の使用が、ポリヌクレオチド混合物を生成するのに特に好ましい。Ausubel他、Current Protocols in Molecular Biology、2章及び4章、(Wiley Interscience、1989年)に記載のプロトコールのような当技術分野で公知の手段によって、ゲノム核酸を調製することができる。cDNAは、当技術分野で公知の手段によって合成することができる(例えば、Maniatis他、Molecular Cloning、A Laboratory Manual(Cold Spring Harbor Lab、米国ニューヨーク州、1982年)を参照のこと)。目的のポリヌクレオチドを含むcDNAの発現ライブラリを、当技術分野で公知の手段によってスクリーニングすることもできる。
【0027】
例えば、これらの手段には、それだけには限らないが、1)共通のヌクレオチド配列を検出するための、ゲノム又はcDNAライブラリへのプローブのハイブリダイゼーション、2)共通の構造的特徴を検出するための発現ライブラリの抗体スクリーニング、及び3)ポリメラーゼ連鎖反応(PCR)による合成が含まれる。特定のDNAコード配列又はその断片の開発は、1)ゲノムDNAからの二重鎖DNA配列の単離、2)目的のポリペプチドのために必要なコドンを提供するDNA配列の化学的作製、及び3)真核性ドナー細胞から単離したmRNAの逆転写による二重鎖DNA配列のin vitro合成によって行うことができる。後者の場合、一般にcDNAと呼ばれるmRNAの二重鎖DNA相補物が、最終的に形成される。
【0028】
ハイブリダイゼーション手順は、混合した合成オリゴヌクレオチド標識プローブを使用して、組換えクローンをスクリーニングするのに有用であり、この場合、各プローブは変性させた二重鎖DNAの不均一な混合物を含むハイブリダイゼーション試料中にある特定のDNA配列の完全な相補物である可能性がある。このようなスクリーニングの場合、一本鎖DNA又は変性した二重鎖DNA上でハイブリダイゼーションを行うことが好ましい。ハイブリダイゼーションは、目的のポリペプチドに関係するmRNA配列が極めて少量しか存在しない供給源に由来するcDNAクローンの検出に特に有用である。言い換えれば、非特異的な結合を避けるようにした厳密なハイブリダイゼーション条件を使用することにより、例えば、混合物中のただ一つのプローブに標的DNAをハイブリダイゼーションすることによって、特定のcDNAクローンをオートラジオグラフィーで可視化させることが可能である。
【0029】
目的のポリヌクレオチドを含むと考えられるcDNAライブラリは、cDNAから誘導した様々なmRNAを卵母細胞に注入し、十分な時間を与えてcDNA遺伝子産物を発現させ、例えば目的のポリヌクレオチドにコードされたペプチドに特異的な抗体、又は目的のポリヌクレオチドにコードされたペプチドの特徴である繰り返しモチーフ及び組織の発現パターンを調べるためのプローブを使用して、所望のcDNA発現産物が存在するかどうか試験することによって、スクリーニングすることができる。或いは、cDNAライブラリは、少なくとも1種のエピトープを有する治療用及び/又は免疫原性ペプチドの発現について、それらのペプチドに特異的な抗体を使用して、間接的にスクリーニングすることもできる。これらの抗体は、ポリクローナル型又はモノクローナル型を作製し、目的のcDNAの存在を示す発現産物を検出するのに使用することができる。
【0030】
適切なプローブが利用可能であるならば、核酸ハイブリダイゼーションを利用するスクリーニング手順によって、任意の生物から任意の遺伝子配列を単離することが可能になる。問題の蛋白質をコードする配列の一部分に相当するオリゴヌクレオチドプローブを、化学的に合成することができる。このためには、アミノ酸配列の短いオリゴペプチド区間がわかっていなければならない。蛋白質をコードしているDNAの配列は、遺伝コードから推測できるが、コードの縮重を考慮しなければならない。配列が縮重しているときは、混合付加反応を行うことができる。これは、変性した二重鎖DNAの不均一な混合物を含む。このようなスクリーニングの場合、一本鎖DNA又は変性した二重鎖DNA上でハイブリダイゼーションを行うことが好ましい。
【0031】
本発明で使用するSERCA2ポリヌクレオチドはDNA又はRNAでよいが、相補的DNA(cDNA)配列であることが好ましい。本発明で使用するポリヌクレオチドは、(a)発現可能で、且つ(b)複製不能、又は宿主ゲノム内に入って複製しないように当技術分野で公知の手段によって改変されたものでなければならない。SERCA2蛋白質を作動的にコードするポリヌクレオチドを組換え発現ベクター、最も好ましくはアデノウイルス構築体の一部として使用することが好ましい。
【0032】
以下は、本発明で使用するのに適するポリヌクレオチドの調製の例であり、どのように特定のポリヌクレオチド組成物が作成されたかを示す特定の実施例が以下に提供される。しかし、複製不能なポリヌクレオチドを調製する他の既知の手段も適し得ることは、当業者には明らかであろう。
【0033】
本発明のポリヌクレオチドは、SERCA2蛋白質を作動的にコードする既知のポリヌクレオチドの機能的誘導体を含む。「機能的誘導体」とは、SERCA2の「断片」、「変異体」、「類似体」、又は「化学的誘導体」をコードするポリヌクレオチドを意味する。分子の「断片」は、分子の任意のペプチド部分を含む。このような分子の「変異体」とは、分子全体又はその断片にかなり類似した天然の分子を意味する。分子の「類似体」とは、分子全体又はその断片にかなり類似した非天然の分子を意味する。
【0034】
本明細書では、ある分子が普通ならその一部分ではない追加の化学的部分を含むとき、別の分子の「化学誘導体」であると言う。これらの部分は、分子の溶解性、吸収性、生物的半減期などを改善することができる。或いは、これらの部分は、分子の毒性を弱め、分子の望ましくない副作用を無くし又は減らすこともできる。このような効果をもたらすことができる当技術分野で既知の部分は、例えば、Remington’s Pharmaceutical Sciences、16版、Mack Publishing Co.、ペンシルベニア州、イーストン(1980年)に開示されている。
【0035】
SERCA2ポリヌクレオチドは、これらのポリペプチドの発現を制御する調節蛋白質を作動的にコードする他のポリヌクレオチドに結合させ又はそれらと一緒に使用し、或いは、認識配列、プロモーター配列、及び分泌配列を含むことができる。当業者なら、過度に実験をすることなく、調節ポリヌクレオチドを選別し、それらを本発明のSERCA2ポリヌクレオチドに組み込むことができるであろう。例えば、ネズミ又はヒトの系で使用するのに適したプロモーター及びその使用法が、前述のCurrent Protocols in Molecular Biologyの1章に記載されている。
【0036】
本発明で使用するためのSERCA2ポリヌクレオチドの特に好ましい形態は、組換え発現ベクターに組み込まれたものとなる。組換え発現ベクターを使用すると、標的組織における遺伝子の発現が長くなる。
【0037】
適切な組換え発現ベクターは当技術分野で公知であり、前述のCurrent Protocols in Molecular Biologyの1章に記載されているベクターが含まれる。2種の特に好ましいプラスミドプロモーターベクターは、pRSV(ラウス肉腫ウイルス(Rous sarcoma virus))及びpCMV(サイトメガロウイルス(cytomegalovirus))であり、特に後者である。この選択は、CMVプロモーターを使用すると、より高レベルの発現がこの状況で得られるという観察結果に基づく。
【0038】
RSVプロモーターを単離し、プラスミドベクターの構築にそれを使用するための適切なプロトコールは、Gorman他、Proc.Natl.Acad.Sci.USA、79:6777、(1982年)に記載されている。他の好ましいプラスミドベクターは、カリフォルニア州サンディエゴのインビトロゲン(Invitrogen)社から市販されているpREP7及びpREVである。ポリヌクレオチドのクローニングにおいて、mRNAの産生に特に適したプラスミドは、Kreig他、Nucleic Acids Res.、12:7057〜7070(1984年)に記載のpSP64Tクローニングベクターである。従来技術を使用して、開始コドンを含む任意のcDNAをこのプラスミドに導入し、発現されたDNA鋳型からmRNAを調製することができる。
【0039】
組換え発現ベクターで使用するための特徴が良く分かっている「on/off」スイッチは、抗生物質(テトラサイクリン)で調節されるプロモーター系である。このような系を構築する手段は、当技術分野で公知である。この点に関する総説について、当業者なら、Furth他、Proc.Natl.Acad.Sci.USA、91:9302〜9306、1994年(トランスジェニックマウスにおける、テトラサイクリンに調節された遺伝子発現制御)、Fishman他、J.Clin.Invest.、93:1864〜1868、1993年(心臓の遺伝子発現のテトラサイクリンによる制御)、及びNiwa他、Gene、108:193〜200、1991年(高発現のトランスフェクトタントのためのプロモーター系の使用)を参照できよう。テトラサイクリンプロモーター系の制御下でSERCA2をコードするプラスミドを、実施例4に記載する。
【0040】
本発明で利用できる様々なウイルスベクターには、アデノ随伴ウイルス(AAV)、アデノウイルス、ヘルペスウイルス、ワクシニア、又はレトロウイルスなどのRNAウイルスが含まれる。単一の外来遺伝子を挿入できるレトロウイルスベクターの例には、それだけには限らないが、モロニーマウス白血病ウイルス(Moloney murine leukemiavirus)(MoMuLV)、ハーベイマウス肉腫ウイルス(Harvey murine sarcoma virus)(HaMuSV)、マウス乳癌ウイルス(murine mammary tumor virus)(MuMTV)及びラウス肉腫ウイルス(Rous Sarcoma virus)(RSV)が含まれる。いくつかの他のレトロウイルスベクターは、複数の遺伝子を組み込むことができる。これらのベクターの全ては、形質導入された細胞を同定及び生成できるように、選択マーカーとして遺伝子を移入又は組み込むことができる。
【0041】
組換えレトロウイルスは欠損があるため、感染性のベクター粒子を産生するためには助けを必要とする。例えば、LTR内の調節配列の制御下にあるレトロウイルスの構造遺伝子の全てをコードしているプラスミドを含むヘルパー細胞系を使用して助けることができる。これらのプラスミドは、パッケージング機序でキャプシド形成のためのRNA転写物を認識するのを可能にするヌクレオチド配列を欠失している。パッケージングシグナルを欠失しているヘルパー細胞系には、それだけに限らないが、例えば、ψ2、PA317、及びPA12が含まれる。これらの細胞系は、ゲノムが全くパッケージされない(詰め込まれない)ため、空のウイルス粒子を産生する。パッケージングシグナルは損なわれていないが構造遺伝子が目的の他の遺伝子で置き換えられているこのようなヘルパー細胞系にレトロウイルスベクターが導入されると、ベクターはパッケージされることができ、ベクターウイルス粒子を産生することができる。
【0042】
DNA発現ベクターのうちでは、トランスフェクション効率(心筋細胞で最大70%)の点からAAVベクターが好ましい。このようなベクターは、外来DNAの比較的大きなセグメントを受け入れることができ、低い免疫原性で高タイターで生成することができるので好ましい。AAVベクターは、(例えばSERCA2ポリヌクレオチドのポリリジン結合体と比べて)冠動脈内輸注法に良く耐えるので有利である。
【0043】
アデノ随伴ウイルス(AAV)は、単純な非病原性の一本鎖DNAウイルスである。パッケージング配列を含むそのcap及びrep遺伝子は、ウイルスの先頭及び末尾を決めている逆方向反復配列の間に挟まれている。cap遺伝子は、ウイルスのキャプシド(capsil)(莢膜)蛋白質をコードし、rep遺伝子産物は、ウイルスの複製及び組み込みに関与している。AAVは、複製のために、ヘルパーウイルス、例えばアデノウイルス又は単純ヘルペスウイルスによって提供される追加の遺伝子を必要とする。AAVは様々な細胞型に感染する。そのウイルスDNAは、ヒト染色体19に優先的に組み込まれることができる。
【0044】
rep及びcap遺伝子を導入遺伝子で置き換えることによって、AAVベクターを作製し、遺伝子治療用ベクターとして使用することができる。ヒト因子IXのcDNAを含むAAVベクターが構築され、免疫適格性マウスの肝臓細胞及び筋肉細胞に感染させるために使用されている。これらのマウスは、6ヶ月以上の間、治療効果のある量の因子IX蛋白質を血液中に産生することができた。AAVベクターを大量調製するための組成物及び方法は、米国特許第6,416,992号に開示されている。
【0045】
本発明で使用するのに特に好ましいAAV構築体を、実施例1に示す。この構築体は、プロモーター、ポリリンカー、及びその他のものを含むシャトルベクター中にSERCA2ポリヌクレオチドをクローニングすることによって、形成される。試料中で使用する特定のアデノ随伴シャトルベクターは、Samulski他によって記載の(Journal of Virology 63(9):3822〜3828、1989年)プラスミドpSub201から誘導した。このプラスミドのXba I断片を、マルチクローニングサイト及びポリアデニル化シグナルの前にある572塩基対のヒトCMVエンハンサー/プロモーター断片に連結させた。このプラスミドをAAV−シャトルと呼び、サイズは約5000塩基対である。
【0046】
3’末端でフラッグシグナルペプチドに融着させたラットSERCA2aのコード配列が報告されている(He他、J.Clin.Invest 100:974〜980、1999年)。Kpn I/Xba I又はHind III/Xba酵素を使用して、緑蛍光蛋白質(GFP)をAAV−シャトル中に好都合にクローニングすることができる(He他、J.Clin.Invest 100:974〜980、1999年)。組換えクローンはDNA配列決定により確認され、大量のプラスミドpDGが記載され、CsCl大量調製プロトコールによって精製されている(Grimm他、Human Gene Therapy 9:2745〜2760、1998年)。
【0047】
純粋な感染性ウイルス粒子を調製するために、挿入されたcDNAを含むAAV−シャトルプラスミド及びpDGプラスミドは共に、CaPO4共沈法を使用して細胞にトランスフェクトされる。トランスフェクションに続いて、細胞は新鮮な培地で培養され、次いでウイルス粒子が回収され精製される。クローニングスキーム及び適切な制限部位の略図を図4に示す。
【0048】
(以下のセクションCで記載の)リポソーム送達のためのSERCA2のカプセル化も、複製可能なAAV構築体を使用する場合と同様に、ウイルスの免疫破壊を制限するはずである。しかし、ウイルスDNAの宿主ゲノムへの組み込みに伴う可能性のある危険性を考えると、このようなベクターの使用は、ベクター産生物の長期間の発現が患者の生存に極めて重要である状況に限られることになる可能性が高い。
【0049】
例えば、特定の標的細胞上の受容体に対するリガンドをコードするもう1つの遺伝子といっしょに1種又は複数の目的の配列をウイルスベクターに挿入することによって、ベクターを標的特異性にすることができる。例えば、糖、糖脂質、又は蛋白質をコードするポリヌクレオチドを挿入することによって、レトロウイルスベクターを標的特異性にすることができる。
【0050】
心臓に特異的なプロモーターを使用することにより、標的筋細胞以外の心筋細胞への本発明の組換え発現ベクターの非特異的トランスフェクションを回避することが好ましい。いくつかのこのようなプロモーターが現在知られており、トリβ−アクチンが挙げられる(実施例1を参照のこと)。当業者なら、ウイルスゲノムに挿入され、目的のSERCA2ポリヌクレオチドを含むウイルスベクターの標的特異的な送達を可能にすることができる他の特定のポリヌクレオチド配列について知っているか、過度に実験をすることなく容易に確認することができよう。
【0051】
心臓組織へのSERCA2導入遺伝子の送達のためにAAVベクターを使用することの利点は、このベクターに対して免疫応答が起こらず、導入遺伝子の発現が最大7ヶ月持続することである。SERCA2導入遺伝子の送達のためのAAVベクターの使用に関する詳細な説明を、以下の実施例1に示す。
【0052】
C.SERCA2ポリヌクレオチドの薬剤調製物
SERCA2ポリヌクレオチドの組成物及びSERCA2ポリヌクレオチドの混合物は、製薬上許容される懸濁液、溶液、又は乳濁液中に入れてよい。適切な媒体は生理食塩水を含み、SERCA2ポリヌクレオチドを標的組織に送達するために抗原提示細胞を使用しない実施形態の場合は、リポソーム調製物を含んでよい。
【0053】
より詳細には、製薬上許容される担体は、無菌の水性又は非水性溶液、懸濁液、及び乳濁液を含んでよい。非水溶媒の例としては、プロピレングリコール、ポリエチレングリコール、オリーブ油などの植物油、及びエチルオレエートなどの注射可能な有機エステルがある。水性の担体には、生理食塩水及び緩衝処理した媒体を含めて、水、アルコール性/水性溶液、乳濁液、又は懸濁液が含まれる。非経口用ビヒクル(vehicle)には、塩化ナトリウム溶液、ブドウ糖入りリンガー液、ブドウ糖及び塩化ナトリウム溶液、乳酸化リンガー液、又は不揮発性油が含まれる。静脈内用ビヒクルには、(ブドウ糖入りリンガー液をベースとしたものなどの)流動性で栄養性の補充液及び電解質補充液などが含まれる。例えば、抗菌剤、抗酸化剤、キレート化剤、不活性ガスなどの保存剤及び他の添加剤も存在してよい。さらに、SERCA2ポリヌクレオチドの組成物を、本発明に従って後で再構成し使用するために、当技術分野で公知の手段を使用して凍結乾燥してもよい。
【0054】
上記に論じた標的化ベクター送達系に加えて、コロイド分散系も標的化送達に使用してよい。コロイド分散系には、水中油乳濁液、ミセル、混合ミセル、及びリポソームを含めて、高分子複合体、ナノカプセル、微粒子、ビーズ、及び脂質ベースの系が含まれる。本発明の好ましいコロイド系はリポソームである。
【0055】
リポソームは、in vitro及びin vivoの送達運搬体として有用な人工膜小胞である。大きさ0.2〜4.0nmの大きな一枚膜小胞(LUV)が、大きな高分子を含む水性緩衝液のかなりの割合をカプセル化できることが示された。RNA、DNA、及び無傷のウイルス粒子は、水性の内部にカプセル化され、生物活性のある形で細胞に送達されることができる(Fraley他、Trends Biochem.Sci.、6:77、1981年)。リポソームは、哺乳動物細胞に加えて、作動的にコードするポリヌクレオチドを植物、酵母、細菌細胞中に送達するのに使用されている。リポソームが効率的な遺伝子移入運搬体となるためには、以下の特徴がなければならない。すなわち、(1)アンチセンスポリヌクレオチドをコードしている遺伝子を高効率でカプセル化するが、それらの生物活性は損なわないこと、(2)標的ではない細胞と比べて標的細胞に優先的且つ有意に結合すること、(3)運搬体中の水性内容物を、高効率で標的細胞の細胞質に送達すること、及び(4)遺伝情報を正確且つ有効に発現することである(Mannino他、Biotechniques、6:682、1988年)。
【0056】
リポソームの組成物は、通常ステロイド、特にコレステロールと組み合わせた、リン脂質、特に相転移温度の高いリン脂質の組み合わせである。他のリン脂質又は他の脂質も使用してよい。リポソームの物理的特性は、pH、イオン強度、及び二価の陽イオンの有無によって決まる。
【0057】
リポソーム調製に有用な脂質の例には、ホスファチジルグリセロール、ホスファチジルコリン、ホスファチジルセリン、ホスファチジルエタノールアミン、スフィンゴ脂質、セレブロシド、及びガングリオシドなどのホスファチジル化合物が含まれる。特に、脂質部分が14〜18個の炭素原子、特に16〜18個の炭素原子を含み、飽和されているジアシルホスファチジルグリセロールが有用である。リン脂質の例には、卵ホスファチジルコリン、ジパルミトイルホスファチジルコリン、及びジステアロイルホスファチジルコリンが含まれる。
【0058】
リポソームの標的化は、解剖学的及び機械的要因に基づいて分類することができる。解剖学的分類は、選択性のレベル、例えば、器官特異性、細胞特異性、及び細胞小器官特異性に基づく。機械的標的化は、それが受動的であるか能動的であるかに基づいて区別することができる。受動的標的化は、洞様毛細血管を含む器官において細網内皮系(RES)の細胞に分布するというリポソームの本来の傾向を利用する。一方、能動的標的化では、モノクローナル抗体、糖、糖脂質、蛋白質など特定のリガンドにリポソームを結合させること、又は本来の所在位置以外の器官及び細胞型を標的させるためにリポソームの組成又は大きさを変更することによってリポソームを改変する。
【0059】
標的化送達系の表面は、様々な方法で改変してよい。リポソームによる標的化送達系の場合は、リポソームの二重層と安定に結合した状態で標的リガンドを維持するために、脂質類をリポソームの脂質二重層に組み込むことができる。標的リガンドに脂質鎖を結合させるために、様々な結合基を使用できる。
【0060】
これらの技術(及び薬物送達を容易にするために通常使用される他の技術)は、当業者によって過度に実験をすることなく、本発明の方法で使用するためのSERCA2ポリヌクレオチドの調製に適合されることができると予想される。特に、上記のパラグラフで論じた手法は、発明者の知る限りでは、SERCA2ポリヌクレオチドをin vivoで筋細胞に送達するためにこれまで使用されていないが、この目的に使用するのに適していると考えられる。このため、上記に挙げた参照は、本発明の方法に必須ではないが、参照により本明細書に組み込む。それが適していることを示す特定の実施例を、以下に示す。
【0061】
D.心臓のSERCA2活性をIN VIVOで増大させるための方法
本発明では、SERCA2ポリヌクレオチドが、それがコードする生物的に活性なペプチドを発現させるのに十分な用量で供給されれば十分である。実現される発現レベルが、「通常の」内在性SERCA2の活性を実質的に補充するのに十分であることが好ましい。SERCA2ポリヌクレオチドが、組換え発現ベクター、好ましくはAAVベクターに含まれ、(セクションCで前述したように)製薬上許容される組成物に配合されると有利である。
【0062】
「通常の」SERCA2レベル及び活性は個体間で様々であり、従って、特定の種について絶対的に決定することはできない。しかし、SERCA2蛋白質の治療前後のレベル、並びにCHF心臓における増強された収縮性及び心臓性能の臨床的兆候をモニターすることによって、特定の治療目的を達成するために望ましいSERCA2発現レベル(「治療上有益な量」)を確認し、それを許容される臨床制限内に維持することができる。SERCA2レベルをモニターする手段は、以下のセクションFに説明する。
【0063】
心臓性能、及びCHFに伴うストレスに対する適応力の改善の臨床的兆候は、心臓病分野の当業者には公知であり、例えば、血流、心臓のポンプ容量、及び心室圧(例えば、血管造影及び心エコー検査による)、カルシウム輸送速度(心臓液試料のin vitro評価による)、耐性試験(例えば、心臓への圧力過負荷なストレスに対する心拍のモニタリングによる)、及びCHF症状の減退の一般的な臨床的兆候(例えば、宿主の持久力が増え、呼吸が容易になる)をモニターすることによって、判定することができる。投与用量は、個々の治療目的を達成するために変更してよい(例えば、急性のCHF病態の宿主においてカルシウム輸送を増大するためのSERCA2の過剰発現)。最大及び最小範囲は、前述のモデル及び大型の哺乳動物種の使用などから得られるヒト以外の動物データの結果を外挿することによっても決定される。
【0064】
SERCA2ポリヌクレオチド送達は、静脈内又は冠動脈内輸注法により(好ましくは、ポリヌクレオチドの大動脈起始部への逆流を最小限にするためにカテーテル法を使用して)行うことが好ましい。或いは、より強くより即時的なSERCA2発現を実現するために、SERCA2ポリヌクレオチドを(例えば、開胸技術を使って)心室壁に注入しても、(例えば、血管造影カテーテル法によって)直接心室に導入してもよい。その後、SERCA2発現をモニターし、必要なだけ送達を繰り返す。処理した宿主を、ポリヌクレオチド又はSERCA2に対する免疫応答などの副作用、及び(例えば、拡張期の過度の延長によって示されるような)過度のSERCA2発現がないかどうかについても注意深くモニターすべきである。
【0065】
しかし、通常、以下の実施例で記載する結果に基づくと、in vivoで筋細胞にSERCA2ポリヌクレオチドをAAV送達することに関係した細胞変性又は他の有害な影響を生じる危険性は低いようであり、野生型のベクターの混入物を排除すること及び他の方法によって(リポソームによるベクターのカプセル化などによって)ベクターの免疫原性を低減させることによって最小限に抑えることができる。
【0066】
E.本発明の方法を試験するための動物モデル
ラットが、その心臓には側副循環がないが、ヒトCHFの状態を満足できる程度に予測できる、うっ血性心不全の再現性のある実験モデルであることが明らかになった。特に、冠動脈の外科的結紮を受けたラットは、ヒトの心筋梗塞後のCHFの特に良いモデルである。CHF動物モデルとしてのラットを作製及び使用するための実験プロトコールは、当技術分野で十分に記載されており、参照文献については、当業者なら、Pfieffer他、Am.J.Med.、76:99〜103、1984年;Johns及びOlsen、Ann.Surg.、140:675〜682、1954年;並びに、Selye他、Angiology、11:398〜407、1960年(その開示内容を、CHF動物モデルの開発及び使用に関する当技術分野の知識を例示するために参照により本明細書に組み込む)を参照できよう。さらに、甲状腺の状態が減退したマウス、又は心臓の収縮性が低減しカルシウムトランジェントが遅くなっている甲状腺機能低下マウスも、再現性のある実験モデルとして使用することができる。
【0067】
さらに、本発明の方法によってSERCA2活性を増大させたCHF心臓における心臓性能への効果を特に予測するトランスジェニック動物モデルが開発された。これらの(SERCA2導入遺伝子を発現する)トランスジェニック動物を複製するのに有用なプロトコールを以下に記載し、実施例1で説明する。このプロトコールは、発現可能な導入遺伝子を哺乳動物に導入するための従来技術におおむね従う。当技術分野の当業者なら、これらの適用例を熟知しており、過度に実験をすることなく本発明の状況でそれらの技術を適用することができよう。
【0068】
例えば、様々な発達段階の胚性の標的細胞を、導入遺伝子を導入するのに使用することができる。胚性の標的細胞の発達段階に応じて様々な方法が使用される。受精卵は、マイクロインジェクションの最も良い標的である。マウスでは、オスの前核の大きさは直径約20マイクロメートルに達するので1〜2plのDNA溶液を複製可能に注入することができる。遺伝子移入の標的として受精卵を使用することは、多くの場合注入されたDNAが第一卵割の前に宿主遺伝子に組み込まれることになるという点で、大きな利点がある(Brinster他、Proc.Natl.Acad.Sci.USA、82:4438〜4442、1985年)。その結果、トランスジェニック非ヒト動物の全細胞が、組み込まれた導入遺伝子を持つことになる。これはまた、通常、生殖細胞の50%がその導入遺伝子を含むことになるため、初代の子孫への導入遺伝子の効率的な伝達として反映されることにもなる。受精卵のマイクロインジェクションは、本発明を実施する際に導入遺伝子を組み込むための好ましい方法である。
【0069】
レトロウイルス感染も、非ヒト動物に導入遺伝子を導入するのに使用することができる。発達途中の非ヒト胚は、胚盤胞段階までin vitroで培養することができる。この間、割球をレトロウイルス感染の標的とすることができる(Jaenisch、Proc.Natl.Acad.Sci USA、73:1260〜1264、1976年)。酵素処理によって透明帯を除去することにより、割球の効率的な感染が得られる(Hogan他、Manipulating the Mouse Embryo、Cold Spring Harbor Laboratory Press、Cold Spring Harbor 、米国ニューヨーク州、1986年)。導入遺伝子を導入するのに使用するウイルスベクター系は、通常、この導入遺伝子を含む複製欠損型レトロウイルスである(Jahner他、Proc.Natl.Acad.Sci.USA、82:6927〜6931、1985年;Vander Putten 他、Proc.Natl.Acad.Sci USA、82:6148〜6152)。ウイルス産生細胞の単層上で割球を培養することによって、容易に且つ効率的にトランスフェクションが得られる(前述のVan der Putten;Steward他、EMBO J.、6:383〜388、1987年)。
【0070】
或いは、もっと後の段階で感染を実施してもよい。ウイルス又はウイルス産生細胞を分割腔に注入することができる(Jahner他、Nature、298:623〜628、1982年)。組込みはトランスジェニック非ヒト動物を形成する細胞の一部分でしか起こらないため、初代のほとんどは、導入遺伝子についてモザイク状となる。さらに、初代は、通常子孫に分かれていくゲノムの様々な位置に、導入遺伝子の様々なレトロウイルス挿入物を含むことができる。さらに、効率は低いが、妊娠中期の胚に子宮内でレトロウイルスを感染させて生殖細胞系に導入遺伝子を導入することも可能である(前述のJahner他、1982年)。
【0071】
導入遺伝子を導入するための第3の標的細胞は、胚性肝細胞(ES)である。ES細胞は、着床前の胚から得られ、in vitroで培養し胚と融合される(Evans他、Nature、292:154〜156、1981年;Bradley他、Nature、309:255〜258、1984年;Gossler他、Proc.Natl.Acad.Sci USA、83:9065〜9069、1986年;及びRobertson他、Nature、322:445〜448、1986年)。導入遺伝子は、DNAトランスフェクション又はレトロウイルスの媒介による形質導入によって、ES細胞に効率的に導入することができる。その後に、これらの形質転換されたES細胞を非ヒト動物由来の胚盤胞に結合させることができる。ES細胞は、その後、胚に定着し、結果として得られるキメラ動物の生殖系の一部になる(総説については、Jaenisch、Science、240:1468〜1474、1988年を参照のこと)。
【0072】
本発明の状況で動物モデルとして使用するために、最適の導入遺伝子は、SERCA2の比較的高い発現レベルをもたらすことができるプロモーターを含むものが好ましい。この点で使用するのに好ましいプロモーターは、β−アクチンイントロンを含む、(例えば鳥類由来の)β−アクチンプロモーターに連結したヒトCMVエンハンサーである。導入遺伝子の発現活性を検出するために、前述したフラッグ抗原エピトープのコード領域を、SERCA2のC末端領域をコードする導入遺伝子の領域に含ませた。実施例1で説明するように、導入遺伝子を発現した初代動物の子孫は成体になるまで生存し、導入遺伝子が初代系列の少なくとも約15%の子孫に含まれていると予想できる。
【0073】
F.SERCA2の発現をIN VIVOでモニターする方法
SERCA2の発現をモニターするために、本発明に従って宿主に導入されるSERCA2ポリヌクレオチドを、既知のレポーター遺伝子を含むように改変してよい。例えば、Norton他、Mol.Cell.Biol.、5:281、1985年に記載のpRSV lac−Z DNAベクターは、蛋白質発現とともにβ−ガラクシターゼを産生する。ルシフェラーゼ及びクロラムフェニコールアセチル基転移酵素(「CAT」;pRSV−CATプラスミドの構築については、例えば前述のGorman他を参照のこと)も使用できる。別の有用なレポーター分子は、免疫測定法で容易に検出することができるフラッグ抗原性ペプチドである。フラッグ抗原性ペプチドの8個のアミノ酸配列及びそれをコードする領域は、当技術分野で既知である。この点に関して参照するためには、当業者なら、Chiang他、Peptide Res.、6:62〜64、1993年を参照できよう。(例えば、開始コドンから約15塩基対の位置に遺伝子を挿入することにより)コード用のレポーター遺伝子をSERCA2蛋白質のC末端に挿入しても、SERCA2触媒活性は妨害されない。このようなレポーター遺伝子の発現を検出する手段は当技術分野で公知であり、詳細には記載しないが、以下に要約する。
【0074】
例えば、本発明に従ってSERCA2ポリヌクレオチドを導入した後にin vivoで発現されたSERCA2を、免疫測定法によって検出することができる。この免疫測定法では、SERCA2蛋白質を液相中で利用し又は固相担体に結合させることができる。さらに、これらのアッセイで利用するSERCA2蛋白質は、様々な方法で検出可能に標識することができる。さらに、SERCA2又はレポーター遺伝子産物に対する抗体を、血液や血清などのアッセイ試料中でのSERCA2ポリヌクレオチド発現を検出するために利用することもできる。
【0075】
簡単に言うと、このような抗体は、当技術分野で公知の手段によって調製することができる。例えば、SERCA2又はレポーター遺伝子産物に特異的な抗体は、抗原性のSERCA2又はレポーター遺伝子ペプチドで非ヒト生物を免疫することによって調製することができる。このようなペプチドは天然の供給源から単離することができ(例えば、総説としてその開示内容を本明細書に組み込んだPopovich他、Am.J.Physiol、261:E377〜E381、1991年に報告されているラットSERCA2を単離するのに使用された方法を参照のこと)、或いは、α−アミノ基のt−BOC又はFMOC保護などの通常使用される方法により、過度に実験をすることなく合成することもできる。
【0076】
後者の方法は、ペプチドのC末端から出発して、各段階で1つのアミノ酸が加えられる段階的合成である(Coligan他、Current Protocols in Immunology、Wiley Interscience、991、ユニット9を参照のこと)。この場合に使用するペプチドは、Merrifield、J.Am.Chem.Soc.、85:2149、1962年、並びにStewart及びYoung、固相ペプチド合成法(Solid Phase Peptides Synthesis)(Freeman、サンフランシスコ、27〜62、1969年)に記載されているものなど様々な公知の固相ペプチド合成方法により、ポリマー1g当たり0.1〜1.0mMolのアミンを含む共重合体(スチレン−ジビニルベンゼン)を使用して、合成することができる。
【0077】
化学合成が完了するとすぐに、0℃で約1/4〜1時間、フッ化水素酸10%アニソール液で処理することにより、ペプチドを脱保護しポリマーから切断することができる。試薬を蒸発させた後、ペプチドを1%酢酸溶液でポリマーから抽出し、次いで凍結乾燥させて未精製物質を得る。通常、溶媒として5%酢酸を使用するSephadex G−15を用いたゲル濾過などの技術によってこれを精製することができる。カラムの適切な画分を凍結乾燥すると、均質なペプチド又はペプチド誘導体が得られ、これらは、次いで、アミノ酸分析、薄層クロマトグラフィー、高速液体クロマトグラフィー、紫外吸収分光法、モル旋光度、溶解度などの標準の技術により特徴を明らかにし、固相エドマン分解法によって定量することができる。目的のペプチドの抗原性は、そのペプチドで免疫された動物の抗体反応の大きさを判定する従来技術によって決定することができる。
【0078】
抗原性のSERCA2ペプチドを調製した後で、(ウサギ、マウス、又はラットなどの)哺乳動物にペプチドを導入することにより、免疫ペプチドに対する抗体を作製する。複数回注入による免疫化プロトコールが、抗原性ペプチドで動物を免疫する際に使用するのに好ましい(例えば、Langone他編、「小用量の免疫原生物による抗血清の調製:皮内複数回注入(Production of Antisera with Small Doses of Immunogen:Multiple Intradermal Injections)」、Methods of Enzymology(Acad.Press、1981年を参照のこと)。例えば、フロイント完全アジュバントに乳濁させた1mgの抗原性ペプチドを皮内注射し、その数週間後にフロイント不完全アジュバント中の同じ抗原を1回又は複数回追加することにより、通常、ウサギで良い抗体反応が得られる。
【0079】
望むなら、免疫ペプチドは、当技術分野で公知の技術を使用して結合することにより、キャリア蛋白質に結合させてよい。ペプチドに化学的に結合されるこれらの普通に使用されるキャリアには、キーホールリンペットヘモシアニン(KLH)、チログロブリン、ウシ血清アルブミン(BSA)、及び破傷風トキソイドが含まれる。次に、結合されたペプチドを使用して動物(例えばマウス又はウサギ)を免疫する。SERCA2は、現在、哺乳動物種間でかなり良く保存されていると考えられているので、SERCA2蛋白質の免疫原性を増大させるためにキャリア蛋白質を使用することが好ましい。
【0080】
動物に産生されたポリクローナル抗体を、例えば、抗体が産生された対象のペプチドが結合されているマトリクスに結合させ、マトリクスから溶出させることによって、さらに精製することができる。当業者なら、ポリクローナル抗体、並びにモノクローナル抗体の精製及び/又は濃縮に関する免疫学分野では普通の様々な技術について知っていよう(例えば、Coligan他、ユニット9,Current Protocols in Immunology、Wiley Interscience,1991年を参照のこと)。
【0081】
モノクローナル抗体は、特異性があり調製が容易であるため、SERCA2発現の検出に使用するのに好ましい。モノクローナル抗体を調製するためには、マウス又はラットの免疫が好ましい。本発明で使用する「抗体」という用語は、無傷の分子、並びに、例えば、エピトープ決定基を結合することができるFab及びF(ab’)2’などのその断片も含むものとする。また、この文脈では、「本発明のmAb’s」という用語は、SERCA2又はレポーター遺伝子産物に特異的なモノクローナル抗体を意味する。
【0082】
モノクローナル抗体(「mAb’s」)を分泌するハイブリドーマの調製に使用する通常の方法は、公知である(Kohler及びMilstein、Nature、256:495、1975年)。簡単に言うと、Kohler及びMilsteinに記載のように、この技術は、外科的検体から得、プールし、次いでSHFP−1に融合させた、黒色腫、奇形癌、子宮頸癌、神経膠腫、又は肺癌である5名の別々の癌患者の排出性(draining)所属リンパ節から単離したリンパ球を含む。ハイブリドーマは、癌細胞系に結合する抗体を産生するかどうかでスクリーニングした。
【0083】
mAb’sの抗原特異性の確認は、(酵素結合免疫測定法、即ち「ELISA」などの)比較的よく使うスクリーニング技術を使用して目的のmAbの基本的な反応パターンを判定することによって、実現できる。あるmAbを評価して、本発明のmAbと同じ特異性を有するかどうか判定することも、前述のように単離した目的の抗原に本発明のmAbが結合するのを試験対象のmAbが妨げるかどうか判定することによって、過度に実験をすることなく、可能である。本発明のmAbによる結合の低減により試験対象のmAbが本発明のmAbに競合することが示される場合は、それら2種のモノクローナル抗体が同じ或いは密接に関係するエピトープに結合する可能性が高い。
【0084】
あるmAbが本発明のmAbの特異性を有するかどうか判定するさらに別の方法では、本発明のmAbを通常反応性の抗原と一緒にプレインキュベートし、試験対象のmAbがその抗原に結合する能力を阻害されるかどうか判定する。そのとき試験対象のmAbが阻害される場合は、高い確率で、本発明のmAbと同じ或いは密接に関係するエピトープ特異性がある。
【0085】
MHCα及びMHCβに共通するエピトープを認識するSERCA2モノクローナル抗体は、検出するSERCA2の構造が本質的に改変されていないかどうか判定するのに特に有用である。このようなmAb’sは、その開示内容を総説及び参考文献として使用するために本明細書に組み込んだ、Dorn他、Am.J.Physiol.、267:H400〜H405、1994年に、再現するのに十分な詳しさで記載されている。
【0086】
SERCA2発現を検出するために使用できる免疫測定法の例は、直接式又は間接式の競合及び非競合免疫測定法である。このような免疫測定法の例は、放射性免疫測定法(RIA)、サンドイッチ法(イムノメトリックアッセイ(immunometric assay))、及びウェスタンブロット法がある。SERCA2又はレポーター遺伝子産物に結合する抗体の検出は、生理試料の免疫組織化学的分析を含めて、順方向、逆方向、又は同時に両方向に作動する免疫測定法を利用して実施することができる。lac−Z又はβ−gal(例えば、β−ガラクトシダーゼ)などのレポーター遺伝子産物を検出するための市販の分析キットが、この点で、使用するのに特に好ましい。
【0087】
使用する抗原又は抗体の濃度は、免疫測定法のタイプ及び使用する検出可能な標識の性質に応じて変わる。しかし、使用する免疫測定法のタイプにかかわらず、利用する抗原又は抗体の濃度は、当業者が常法の実験を実施して容易に決定することができる。
【0088】
このような抗原又は抗体は、多数の様々なキャリアに結合し、それに特異的に反応する抗原又は抗体の存在を検出するのに使用することができる。公知のキャリアの例には、ガラス、ポリスチレン、ポリ塩化ビニル、ポリプロピレン、ポリエチレン、ポリカーボネート、デキストラン、ナイロン、アミラーゼ、天然及び修飾セルロース、ポリアクリルアミド、アガロース、並びに磁鉄鉱が含まれる。本発明では、キャリアの性質は可溶性でも不溶性でもよい。当業者なら、これに関して、使用するのに適した他のキャリアについて知っているか、常法の実験を実施してそれらを確認することができよう。
【0089】
当業者には既知の多くの様々な標識及び標識方法がある。本発明で使用できる標識のタイプの例には、酵素、ラジオアイソトープ、コロイド金属、蛍光性化合物、化学発光化合物、及び生物発光化合物が含まれる。
【0090】
或いは、SERCA2ポリヌクレオチドは、その一般的技術を以下に要約する、当技術分野で既知の定量的ポリメラーゼ連鎖反応(PCR)プロトコールを使用して(好ましくは従来の心臓組織の生検技術を使用して得た標的細胞試料中で)検出することができる。
【0091】
精製又は未精製の任意の組織標本に由来する核酸を、それが標的核酸を含む特定の核酸配列を含む又は含むと推測されることを条件として、1種又は複数の出発核酸として利用することができる。従って、このプロセスは、例えば、メッセンジャーRNA(mRNA)を含めて、DNA又はRNAを使用してよく、DNA又はRNAは1本鎖でも2本鎖でもよい。RNAを鋳型として使用する場合は、その鋳型をDNAに逆転写するのに最適な酵素及び/又は条件を利用することになる。さらに、それぞれ1本の鎖を含むDNA−RNAハイブリッドも利用してよい。また、核酸の混合物を使用しても、以前の増幅反応で同じ又は別のプライマーを使用してここで作製した核酸をそのように使用してもよい。増幅するヌクレオチド配列は、大分子の一部分でもよく、或いは、別々の分子として最初に存在することもでき、従って、この特定の配列が核酸全体を構成する。増幅する配列は、最初に精製した形で存在する必要はなく、ヒトDNA全体に含まれるような複雑な混合物のごく一部分でよい。
【0092】
試料の標的ヌクレオチド配列が2本の鎖を含む場合、鋳型として使用できるように核酸の鎖を分離することが必要である。鎖の分離は、別々のステップとして又はプライマー伸張生成物の合成と同時に実施することができる。この鎖の分離は、物理的、化学的、又は酵素的手段を含めて、様々な適切な変性条件を使用して実現することができる。「変性させる」という語は、これら全ての手段を含む。核酸の鎖を分離する1つの物理的方法は、核酸を変性するまで加熱するものである。通常の加熱変性は、約1〜10分間、約80〜105℃の温度を必要とする。鎖の分離は、ヘリカーゼとして知られている酵素群のうちの1つの酵素又はヘリカーゼ活性のある酵素RecAにより、DNAを変性することが知られているリボATPの存在下で誘導してもよい。ヘリカーゼを用いた核酸鎖の分離に適した反応条件は、Kuhn Hoffmann−Berling(CSH−Quantitative Biology、43:63、1978年)に記載されており、RecAを使用する技術はC.Radding(Ann.Rev.Genetics、16:405〜437、1982年)に総説がある。
【0093】
増幅する標的核酸を含む核酸が一本鎖の場合、1種又は2種のオリゴヌクレオチドプライマーを加えることにより、その相補鎖が合成される。単一のプライマーを利用する場合は、プライマー、重合用作用物質、及び後述する4種のヌクレオシド3リン酸の存在下で、プライマー伸長生成物を合成する。この生成物は、一本鎖核酸に相補的なものとなり、一本鎖核酸にハイブリダイズして、長さが不揃いな鎖の二重鎖を形成する。次にこの二重鎖を一本鎖に分離させて、一本に分離された相補鎖2本を生成することができる。或いは、2種のプライマーを一本鎖核酸に加え、記載したように反応を実施してもよい。
【0094】
核酸の相補鎖を分離すると、その核酸がもともと二本鎖であったか一本鎖であったかにかかわらず、分離された鎖は、追加の核酸鎖を合成するための鋳型としていつでも使用できる。この合成は、プライマーが鋳型にハイブリダイズできる条件の下で実施する。通常、合成は好ましくはpH7〜9、最も好ましくはpH8の緩衝水溶液中で実施する。過剰モル(ゲノム核酸の場合、通常、プライマーと鋳型の比が約108対1)の2種のオリゴヌクレオチドプライマーを、分離された鋳型鎖を含む緩衝液に加えることが好ましい。しかし、本発明のプロセスを診断で適用するのに使用する場合は、相補鎖の量が分からない可能性があり、従って、相補鎖に対するプライマーの量を確実に決定することはできない。しかし、増幅しようとする配列が複雑で長い核酸鎖の混合物に含まれるとき、実用的には、加えるプライマーの量は通常、相補鎖(鋳型)の量に対して過剰モルである。プロセスの効率を高めるためには、大過剰モルが好ましい。
【0095】
いくつかの増幅実施形態では、基質、例えばデオキシリボヌクレオチド3リン酸のdATP、dCTP、dGTP、及びdTTPを適量、プライマーと別に又は一緒に、合成混合物に加え、得られる溶液を約1〜10分間、好ましくは1〜4分間、約90〜100℃に加熱する。この加熱期間の後、溶液をプライマーのハイブリダイズにとって好ましい室温まで自然冷却させる。冷却した混合物に、プライマー伸長反応を実施するのに適した作用物質(本明細書では「重合用作用物質」と呼ぶ)を加え、当技術分野で既知の条件下で反応を起こさせる。重合用作用物質は、熱安定性である場合は、他の試薬とともに加えてもよい。この合成(即ち増幅)反応は、室温から、最高ではそれを超えると重合用作用物質が機能しなくなる温度までで行うことができる。従って、例えばDNAポリメラーゼが作用物質として使用される場合は、通常、温度は約40℃以下である。
【0096】
重合用作用物質は、酵素を含めて、プライマー伸長生成物の合成を実現するように機能するどんな化合物又は系でもよい。この目的に適した酵素には、例えば、大腸菌(E.coli)DNAポリメラーゼI、Taqポリメラーゼ、大腸菌(E.coli)DNAポリメラーゼIのクレノウ断片、T4 DNA ポリメラーゼ、他の入手可能なDNAポリメラーゼ、ポリメラーゼ突然変異蛋白質、逆転写酵素、リガーゼ、及び、熱安定性酵素(即ち、変性を引き起こすのに十分な高い温度下に置かれた後でも、プライマー伸長を行う酵素)を含めて、他の酵素が含まれる。適切な酵素を使用すると、ヌクレオチドの正しい組み合わせがうまく進み、各変異ヌクレオチド鎖に相補的なプライマー伸長生成物が形成される。通常、合成は各プライマーの3’末端で開始され、合成が終わるまで鋳型鎖に沿って5’方向に進行し、様々な長さの分子を生成する。しかし、前述したのと同じプロセスを使用して、5’末端で合成を開始し、別の方向に進行する重合用作用物質があるかもしれない。
【0097】
新しく合成された変異ヌクレオチド及びその相補的な核酸鎖は、前述のハイブリダイズする条件下で二本鎖分子を形成し、このハイブリッドをプロセスの後続のステップで使用する。次のステップでは、新しく合成された二本鎖分子を前述した手順のいずれかを使用して変性条件において、一本鎖分子を生成させる。
【0098】
上記のプロセスを一本鎖分子に対して繰り返す。反応が先に規定した条件下で進行するように、必要なら、追加の重合用作用物質、ヌクレオシド、及びプライマーを加えてよい。やはり、合成各オリゴヌクレオチドプライマーの一端で開始され、鋳型の一本鎖に沿って進行し、追加の核酸を生成する。このステップの後、伸長生成物の半分は、2種のプライマーが結合した特定の核酸配列からなるはずである。
【0099】
変性及び伸長生成物合成のステップは、標的の変異ヌクレオチド配列を検出に必要な量まで増幅するのに必要なだけ繰り返すことができる。生成される変異ヌクレオチド配列の量は、指数関数的に増加する。
【0100】
増幅した生成物は、放射性プローブを使用せずに、サザンブロット分析により検出することができる。このようなプロセスでは、例えば、標的ヌクレオチド配列を極めて低いレベルで含むDNAの少量のサンプルを増幅し、サザンブロット法により分析する。シグナルが高レベルで増幅されると、非放射性プローブ又は標識の使用が容易になる。
【0101】
定量的PCRを実施するための好ましい方法は、1種又は複数の塩基対に誘導された突然変異を含み、その結果、配列及び大きさが標的の鋳型遺伝子と異なる競合物になった競合鋳型を使用して実施する競合的PCR法である。プライマーのうちの1種をビオチン標識し、或いは、好ましくは、結果として得られるPCR生成物の一方の鎖(通常アンチセンス鎖)を、アミノ−カルボキシル、アミノ−アミノ、ビオチン−ストレプトアビジン、又は他の適度に緊密な結合によって、適切な反応物に緊密に結合された固相の支持体に固定化できるように、アミノ化する。PCR生成物、固相の支持体、及び反応物の間の結合が共有結合であり、従って、変性条件下で結合を解けにくいものにすることが最も好ましい。
【0102】
PCR生成物のアミノ化又はビオチン標識した鎖を固定化した後、結合していない相補鎖をアルカリ性の変性洗浄液中で分離し、反応環境から除去する。標的に対応する配列特異的なオリゴヌクレオチド(「SSO’s」)及び競合核酸を検出タグで標識する。次に、SSOを、除去された結合していないセンス鎖との競合が無い状況でアンチセンス鎖にハイブリダイズさせる。適切な分析試薬を加え、使用した検出タグ及び固相支持体の手段に適したELISA測定法、好ましくはELISAマイクロプレートリーダーによって、ハイブリダイゼーションの程度を測定する。標的鋳型及び競合鋳型を含む鋳型を増幅するPCR反応から別々に得られた標準曲線を用いて測定値を比較して、標的の核酸含有量を導出する。
【0103】
この方法は、定量的で、PCRサイクルの数によって変わらず、SSOプローブとPCR生成物の相補鎖との競合に影響を及ぼされないという点で有利である。
【0104】
或いは、重合ステップ及びハイブリダイゼーションの全ステップを固相の支持体上で実施することもできる。この方法では、固相支持体上に捕捉されるのは、PCR生成物の鎖ではなく、ヌクレオチド重合プライマー(好ましくはオリゴヌクレオチド)である。次に、標的及び競合核酸PCR生成物を、溶液の状態で固相支持体に加え、重合ステップを実施する。重合生成物の結合されていないセンス鎖を、前述の変性条件下で除去する。
【0105】
標的核酸と競合核酸の比を、適切な測定手段(好ましくはELISAリーダー)を使用して標識されたオリゴヌクレオチドSSOプローブを検出することによって決定することができる。この方法の効率は非常に高くできるため、重合ステップの連鎖反応が不必要になり、従って、この方法を実施するために必要な時間が短縮できる可能性がある。重合の最終生成物をハイブリダイゼーションのために反応管から固相支持体に移動させる必要がなく、従って、それらが損失又は損傷する潜在的可能性が限られてくるため、この方法の正確性も高くなっている。しかし、特定の試料にとって必要なら、PCRを使用して標的及び競合核酸を別々の反応管で増幅させ、続いて固相支持体上で最終の重合反応を実施してよい。
【0106】
結合したPCR生成物の形成を示す、当業者に既知の様々な検出可能のシグナルを提供することができる分子(標的及び競合PCR生成物の形成を示す別の色を呈する、標識したヌクレオチド発色団など)を、反応の最後の数サイクルの進行中に反応溶液に加えることができる。標的核酸と競合核酸の比は、ELISAによって又は他の適切な測定手段及び固定化したハイブリダイゼーションプライマーの3’末端に結合させた検出タグに反応する試薬によって、求めることができる。この方法は、従来の非競合的PCRプロトコールを実施することによって、(定量せずに)特定の遺伝子が試料中に存在するかどうか検出できるように適合させることもできる。
【0107】
当業者なら、前述の方法で使用するのに適したプライマーを選択する方法を知っているか、容易に確認できよう。前述の技術に関するさらに詳細な内容については、Kohsaka他、Nuc.Acids Res.、21:3469〜3472、1993年;Bunn他、米国特許第5,213,961号の開示内容、並びにInnis他、PCRプロトコール:方法と適用のガイド(PCR Protocols:A Guide to Methods And Applications)、Acad.Press、1990年を参照できよう。これらの開示内容は、定量的PCRプロトコールに関する当技術分野の状況を例示するためだけに、本明細書に組み込む。
【0108】
PCR、オリゴマー制限(Saiki他、Bio/Technology、3:1008〜1012、1985年)、対立遺伝子特異的オリゴヌクレオチド(ASO)プローブ分析(Conner他、Proc.Natl.Acad.Sci.USA、80:278、1983年)、オリゴヌクレオチドライゲーション分析(OLA)(Landegren他、Science、241:1077、1988年)などの、特定のDNA配列の検出に通常適用されるどんな方法によっても、前述したように検出したSERCA2ポリヌクレオチドをさらに、溶液の状態で又は固体支持体に結合させた後で、評価、検出、クローン化、配列決定などすることができる。DNA分析に関する分子技術の総説があり、当技術分野で公知である(Landegren他、Science、242:229〜237、1988年)。
【0109】
本発明を十分に説明してきたが、その実施を例示する実施例を以下に記載する。しかし、これらの実施例は、添付の特許請求の範囲によって定義される本発明の範囲を限定するものとみなされるべきではない。
【0110】
実施例において、略語「min」は分を、「hrs」及び「h」は時間を意味し、(mlなどの)測定単位は標準の略語によって示す。
【実施例1】
【0111】
AAVベクターによるSERCA2のクローニング及び発現並びにマウス心筋の心筋細胞機能の評価
SERCA2 cDNAを従来のクローニング技術を使用してアデノ随伴ウイルスベクター中にクローニングした。SERCA2を発現するアデノ随伴ウイルス又はGFPレポーター配列の構築は、基本的に以下のように実施した。
【0112】
アデノ随伴シャトルベクターをSamulski他、Journal of Virology 63:3822〜3828、1989年に記載されているプラスミドpSub201から誘導した。このプラスミドのXba I断片を、マルチクローニングサイト及びポリアデニル化シグナルの前にある572塩基対のヒトCMVエンハンサー/プロモーター断片に連結した。このプラスミドをAAV−シャトルと呼び、サイズは約5000塩基対である。
【0113】
3’末端でフラッグシグナルペプチドに融着させたラットSERCA2a、及びKpn I/Xba I酵素又はHind III/Xba酵素を使用してAAV−シャトル中にクローニングした緑蛍光蛋白質(GFP)のコード配列が、それぞれ報告されている(He他、J.Clin.Invest 100:974〜980、1999年)。組換えクローンをDNA配列決定により確認し、大量のプラスミドをCsCl大量調製(maxiprep)プロトコールによって精製した。プラスミドpDGはGrimm他によって記載されており(Human Gene Therapy 9:2745〜2760、1998年)、同様にCsCl精製した。
【0114】
純粋な感染性ウイルス粒子を生成するために、挿入されたcDNAを含むAAV−シャトルプラスミド及びpDGプラスミドを共に、CaPO4共沈法を使用して293T細胞にトランスフェクトした。各ウイルス調製物のために、サブコンフルエントな293T細胞を蒔いた直径15cmの50セル培養プレートを使用した。各15cmプレートに、18マイクログラムのシャトルプラスミド及び70マイクログラムのpDGプラスミドをトランスフェクトした。トランスフェクションの翌日に10%ウシ胎児血清を含む新鮮なDMEMに培地を変更し、細胞を、5%CO2、摂氏37度でさらに2日間インキュベートした。
【0115】
ウイルス粒子の回収は、2.5mlのDMEM中で各プレートから293T細胞をかきとることによって行った。次に、この細胞を遠心し、25mlのDMEM中に再懸濁させ、摂氏−80度で凍結させた。さらに2回の凍結融解サイクルの後、100マイクログラムのDNアーゼ I及びRNアーゼ Hを加え、懸濁液を摂氏37度で30分間インキュベートした。もう一度3000gで遠心した後、上清を0.5%デオキシコレートに加え、摂氏37度で30分間インキュベートした。次に、この溶液を、5マイクロメーター及び0.8マイクロメーターのシリンジフィルターに順番に通して濾過し、3mlのヘパリン−アガロース懸濁液とともに室温で1時間回転台の上で回転させた。次に、この懸濁液をガラスカラムの上に載せ、ヘパリン−アガロース樹脂を0.254M NaClを含むPBS25mlで洗浄した。洗浄後、ウイルス粒子を0.554M NaClを含むPBSで溶出し、タイター評価のために少量の画分で保存した。最高のタイターの画分をプールし、PBSで透析し、摂氏4度で保存した。クローニングスキーム及び適切な制限部位の略図を図4に示す。
【0116】
次に、甲状腺機能低下マウスの左心室壁にウイルスベクターを投与した。甲状腺の状態が減退したマウス又は甲状腺機能低下マウスは、心臓の収縮性が低減しカルシウムトランジェントが遅くなっている。アデノウイルスベクターを、5〜10μlのアリコートで送り込み、左心室壁の異なる5つの部分に注入した。
【0117】
標的の導入遺伝子を含むAAVの発現効率は一定で、当技術分野で公知である。これにより、心筋細胞の約50%で導入遺伝子が発現する。核コードβGalを発現するAAV又はアデノ随伴ウイルスをマウスの心臓の左心室自由壁に注入することによっても、同様の結果が得られた(図5)。ここでは、アデノウイルス又はAAV注入の2、4、6、又は8週間後に動物を屠殺した。マウスの心臓からのデータにより、AAVの強い発現は注入後6又は8週間持続するが、アデノウイルスの発現は有意に低減したことが示されている。
【0118】
SERCA2発現AAVベクターの注入4週間後に、いわゆるランゲルドルフ単離灌流装置で心臓の収縮性能を判定した。肺静脈を通して左心室に風船を挿入し、収縮機能、特に収縮期の圧力上昇又はdP/dtの最高値、拡張期の圧力低下又はdP/dtの最小値、及び収縮期全体の圧力を、対照としてGFP CREを発現するAAVを注入した心臓で判定した。甲状腺機能低下性マウスの心臓において、全ての収縮性周辺部で非常に有意な上昇が得られ、収縮機能が正常範囲に回復した。図6は、フラッグ標識したSERCA2を発現するAAVを注入した甲状腺機能低下性マウスから単離したマウス心臓の収縮性能についてのデータを示す。収縮期の収縮速度dP/dtの最大値は、GFP発現AAVを注入したマウスと比べて、有意に上昇した。同様に、拡張期の弛緩速度即ち−dP/dtは、AAV SERCAを注入された甲状腺機能低下性マウスで有意に上昇し、収縮期圧力も、SERCA2aを発現するAAVを注入したマウス心臓で有意に上昇した。
【実施例2】
【0119】
SERCA2発現トランスジェニック動物及び対照動物における圧力過負荷モデル及び心臓肥大モデル
CHF心臓の条件を模倣した条件下で導入遺伝子を含む動物におけるSERCA2の追加発現の効果を試験するために、性別、年齢、体重が同程度の動物群を処理して、心臓狭窄を誘発させ、腹大動脈狭窄を発症させ、食塩負荷させた。CHF心臓で顕著なこのような条件下で、これらの動物は心肥大(CH)を発症し、心臓への圧力過負荷(PO)に伴う心臓異常を起こす。導入遺伝子発現を心臓組織に対して特異的にするために、SERCA2 CMV/β−アクチントランスジェニック動物を使用した。
【0120】
簡単に言うと、このような動物を開発するために、実施例1で記載したSERCA2/AAVベクター構築体を使用して開発したトランスジェニック動物(体重約200〜250gのラット)及び対照の動物を腹腔動脈のレベルで腹大動脈を分離するために麻酔し、トランスジェニック動物では、ステンレススチール製のヘモクリップ(Edward Weck&Co.、ノースカロライナ州)を使用して腎動脈の上で大動脈を外科的に狭窄した。後者の手順により、大動脈管腔の大きさが約50%小さくなった。
【0121】
手術の約2日後から、大動脈を狭窄したトランスジェニック動物に、ゴマ油に懸濁させたデオキシコルチコステロンを1週2回筋肉内注射し(25mg/g体重)、飲料水中1%塩化ナトリウムを与えた。通常は手術後1〜2週間内に生じるCHを発症するかどうか、全ての動物を心臓超音波(心エコー検査)を用いてモニターする。さらに、これらの動物は、圧力過負荷をかけることにより急性の損傷期を経験させる。
【実施例3】
【0122】
トランスジェニック動物におけるCHF状態に対する適応反応
AAV/SERCA2構築体を使用して開発したトランスジェニック動物は、手術の6週間後に試験すると(SERCA2のmRNAレベルを検出して判定された)SERCA2遺伝子転写が有意に減少し、通常、10〜12週で心不全を発症し始める。SERCA2 mRNAレベル及び蛋白質レベルを、前述のノーザンブロット法及びウェスタンブロット法又は他の分析技術(例えば、やはり前述したPCR)によって決定した。トランスジェニック動物及び対照動物の(SRカルシウムトランジェントに関係している)酵素活性レベル及び左心室機能を下記に記載するように測定した。他のCHF動物モデル及び本発明の方法による治療を受けているヒトにおいて、以下に記載するプロトコールに従って、心臓性能に関する同じパラメータを測定することができる。
【0123】
心臓収縮機能の評価
A.心室の機能性を判定するための心エコー検査のプロトコール(in vivo)
心エコー図の撮像は、7Mhzで作動する2周波フェーズドアレイトランスデューサー、及び積算ドプラー出力機能を備えた「ACCUSON(登録商標)」128超音波コンソールを用いて、適切な動物モデルで実施する。心エコー事象のタイミングは、皮下電極から得られる同時の心電図記録と相関がある。撮像の深さは2cm、セクタ角度はフレームレートが50/秒になる60にセットする。パワー設定は75dBとする。2次元イメージを使用して、Mモード記録(1000ライン/秒で得られる)に適したカーソル位置を選択することができる。
【0124】
軽く麻酔した動物をうつ伏せに置き、胸骨近くの左の肋間隙を通して撮像を実施する。傍胸骨長軸図及び短軸図を得、ビデオテープに録画し、上述したように、2次元図を使用して心室の中央レベルで実施するMモード記録のためのカーソル位置を導く。全ての関連した測定値は、American Society of Echocardiography推奨の臨床的慣行に関する判断基準を使用して、2次元化したMモード記録から得ることができる。隔壁の厚さは、拡張末期における立ち上がりから立ち下がりまでを測定する。
【0125】
後壁心内膜は、収縮期の傾きの最も大きい線で特定され、拡張末期における後壁の立ち上がりから心外膜境界の立ち上がりまでを測定する。左心室拡張末期径(LVEDD)は、心室径が最大の時点で、心室中隔の立ち下がりから後壁の立ち上がりまでを測定する。左心室収縮末期径(LVESD)は、同じ判断基準を用いて、心室径が最小の時点で測定する。収縮機能の指標としての心室内径短縮率(FS)は、[(LVEDD−LVESD)/(LVEDD 0)]×100で算出する。
【0126】
B.試料液を抜き取り、心臓内の血行動態パラメータを決定するためのマイクロマノメータ付きカテーテルによる処置のためのプロトコール(in vivo)
マウスを例にすると、体重20〜30gの成体マウス(PO有り又は無しの対象及びトランスジェニック)をケタミン(100mg/kg、腹腔内投与(IP))及びキシラジン(5mg/kg、腹腔内投与(IP))の混合物で麻酔する。解剖顕微鏡下で、動物をうつ伏せにし、正中線頚部切開を行って気管及び頚動脈を露出させる。次に、20ゲージの鈍針を、総(tital)容量0.2ml、呼吸数100/分のボリュームサイクル方式げっ歯類用人工呼吸器(ハーバードアパレイタス(Harvard Apparatus)社製)に連結されている気管カニューレを補助するように気管に通す。換気の妥当性を、胸部の膨張を目視検査して判定する。挿管後、大動脈圧を測定するために、1本の頚動脈に、液体入りカテーテル(管状になるよう火炎で延伸したポリエチレン(PE))付きカニューレを挿入する。次に、胸部を開き、2F高忠実度のマイクロマノメータ(ミラー(Millar)社)を僧帽弁から挿入して、LV中に取り付ける。
【0127】
(例えば、オンライン検出装置を使用して)LV収縮期圧力及び拡張期圧力、dP/dtの最大値及び最小値、並びに大動脈圧を継続的に測定して、血行動態パラメータをモニターする。データは、1500試料/秒の速度で得られ、高忠実度のカテーテルの周波数応答は10,000Hz超まではフラットである。マウスの心拍速度が速い(300〜500/分)ため、そのdP/dtを正確に測定するためにこれらの高いサンプルレートが必要である。
【0128】
血行動態パラメータのオフライン分析は、LV圧力低下の時定数(タウ(τ))の計算に加えて最小及び最大LV圧力並びに大動脈血圧の測定を含むことができる。特に拡張期弛緩の最初の等容性期を反映するタウ(τ)の値の決定は、SERCA2機能に密接に関連づけられている。
【0129】
C.単離した筋肉生検組織において筋細胞の短縮を判定するためのプロトコール(in vitro)
このプロトコールをラットで例示すると、切除による損傷を最小限にし拘縮を防ぐために25mM BDMを含む酸素化タイロード液中で左心室乳頭筋を切除し、受動的伸張を避ける。その端を短い5−0シルク縫合糸で結びつけて調製物の心室壁及び弁の端に付着させ、過剰伸張を防ぐ。筋肉の一方の端はCambridge400 Isometric force transducer(等尺性力トランスデューサー)(フルスケール5g、分解能100μg)のガラスロッドに付着し、もう一方の端はトランスデューサー台(Newport423、±1μm)上の小型のステンレススチールフックに付着するようにする。都合よく見られるように、1mlの試験浴をOlympus SZ45ステレオ顕微鏡の視野内におく。浴の温度を33℃に維持する。
【0130】
次に、静止時の力が最初にトランスデューサーチャネルに記録されるゆるんだ長さに筋肉を調整し、BDMを洗い流し、(例えば、Gras刺激装置を使用して)双極性の刺激用電極を浴中に沈める。筋肉を、0.2Hz(閾値より20%高い)で刺激し、30分後、灌流液を[Ca2+]0.5mM溶液から[Ca2+]2.0mM溶液に切り換える。全ての灌流液を100%O2でバブリングする。
【0131】
定常状態での等尺性張力の発生は、筋肉長の増加で測定し、発生した張力が最大になるLmaxに対する割合として表す。(結果をin vivoの筋肉組織性能に外挿する際に)使用する筋肉の長さが短いことを補正するために、張力のピーク時に短くなる歪みを、筋肉上に置いた対の表面細粒を使用して測定し、ビデオカメラで録画する。
【0132】
NIH Image 1.47を使用してData Translationビデオフレームグラッバーボード付きMacintosh Centris 650上でマーカー位置を検出しデジタル化することが好ましい。任意の固定した基準長に関する歪みを使用して長さの縮尺を補正すると、長さ−張力曲線の下降を打ち消すことが示された。
【0133】
収縮性は、等尺性の応力−歪み曲線の傾きで表され、このときの応力は発生した張力(静止期全体)を筋肉の断面積で割った値から計算する。この断面積は、機械的試験後に(その重量から決定した)標本の体積を、両端の間の無負荷の筋肉の長さで割ることにより推定する。細胞外Ca2+濃度0.5〜2.5mMの関数としてこれらのプロトコールを繰り返すことにより、これらの調製物の潜在的なCa2+感度を特徴付けることが可能になる。
【0134】
これらの実験のSERCA2機能及びEC結合をさらに特徴づけるために、Na/Ca交換がNaを含まないタイロード液で遮断されたとき、及び筋小胞体のカルシウム蓄積が10mMカフェインで遮断されたときの弛緩時間を測定することによって、SECRA2のポンプ機能を試験する。Na/Ca交換を測定すると、弛緩時間は約30%遅くなる。試験筋肉の弛緩特性は、チャートレコーダ(例えば、Gould 2200レコーダ)及び高速(40mm/秒)のマイクロコンピュータ取得システム(例えば、市販のStrawberry Tree)に記録された等尺性張力の追跡から、筋肉長とカルシウムの関数として決定される。測定されるパラメータには、ピーク張力から90%回復までの時間及び等尺性の力の低減の指数時定数が含まれる。筋節の長さが損傷されていない中央の筋肉を一定に保つように筋肉長が制御されると、等尺性弛緩が有意に延長されることが研究により示されているので、これらの測定値は前述のアーチファクトにも当てはまり、補正可能である。
【0135】
前述したのと同じ手法を用いて、解剖顕微鏡下でラットの左心室から単離した長くて薄い均質な梁柱を使用して、収縮の研究を実施することもできる。梁柱を使用して研究すると、従来の乳頭筋調製物からは得られない筋肉の一連の弾性があるため、拡張期収縮機序における絶対的変化に関する、アーチファクトとは無関係なより具体的な情報が得られる。左心室壁の他の部分領域を、辺縁検出及び細胞内Ca2+トランジェントの研究をするために心筋細胞を単離するのに使用できる。
【0136】
D.心筋細胞の単離及びカルシウムトランジェントレベルの評価のためのプロトコール(in vitro)
心筋細胞は、コラゲナーゼ灌流法を使用して成体ラットの心臓から調製することができる。この筋細胞は、4%ウシ胎児血清の不在下又は存在下で少なくとも4日間維持し、成体の心筋細胞の形態及び代謝的特徴を維持することができる。1匹の成体ラットの心臓から、9〜10x106個の生存可能な筋細胞を得ることができる。
【0137】
心筋細胞は、心室壁の小片から調製することもできる。これにより、筋細胞機能、乳頭筋機能、並びに同じ心臓から得られるSERCA2遺伝子発現及びSERCA2機能に関係するパラメータについて判断することが可能になる。このために、心室組織を細かく刻み、25mM NaHCO3、3.4mM MgCl2、30mMタウリン、及び2mMカルニチンを追加したCa2+を含まない改変Joklik最少必須培養液(MEM)中で洗浄する。次に、この組織を0.1% コラゲナーゼ タイプII(ワーシントン(Worthington)社)、0.1%BSAフラクションV、及び25μMCaCl2を含むMEMで消化する。連続的な酸素添加のもと、32℃で組織を穏やかに攪拌しながら、酵素消化を30〜40分間継続する。口径の大きな血清ピペットで穏やかに粉砕することによって、最終的に細胞を完全に分散させる。
【0138】
次に、細胞を300μMナイロンメッシュに通して濾過し、洗浄する。次に、カルシウムレベルを徐々にその場の濃度が1μMになるまで上昇させる。この手順により、最大2x106細胞/心臓を得ることができ、その60〜70%が以下に記載する研究で使用可能な生存可能な棒形の筋細胞である。
【0139】
単離した成体ラットの心筋細胞(非培養)を118mM NaCl、4.8mM KCL、1.2mM NaH2PO4、1.2mM MgSO4、1mM CaCl2、11mMグルコース、及び25mM Na−N−2−ヒドロキシエチルピペラジン−N−2−エタンスルホン酸を含む10mlの溶液中に懸濁させ、500μlの細胞懸濁液を1.5mlの普通のタイロード液に加え、Indo Iのアセトキシメチルエステル誘導体のDMSO中1mM溶液5μlを加える。細胞を室温で15〜20分間インキュベートする。次に、Indo I添加の均一性、及びきれいな横紋を伴う棒形細胞の形態に基づき細胞を選別する。石英光学素子を装備したNikon Diaphot顕微鏡の台上に取り付けた小型灌流チャンバーに細胞を移す。細胞を灌流チャンバーの底に自然沈降させ、90%O2−5%CO2でガス補給し37℃で維持したタイロード液を用いて5ml/分で連続的に灌流する。細胞を外部の白金電極を用いて0.2、0.33、1.0、2.0、及び5.0Hzで駆動し、Ca2+トランジェントを蛍光定量によって測定する。同様の研究を、新生児期のラット又はマウスの筋細胞を使用して実施することもできる。Adv SERCA2を新生児筋細胞に感染させると、カルシウムトランジェントが顕著に低減する。
【0140】
カルシウムトランジェントレベルに関しては、カルシウム2++トランジェントが最も低減するための時間は、SERCA2導入遺伝子を含む陽性の筋細胞において、in vitroで観察されるのと同様の程度まで有意に短縮された(実施例4、図3、p<0.05)。心室圧に関しては、左心室で得られた最大圧力(LVdP/dt最大値)は、トランスジェニック動物において有意に大きかったが、最小レベルの場合、心室の筋肉は、トランスジェニック動物において拡張期弛緩がより速くなる傾向を示した(n=3、コントロール(n=5)と比べて)。動物で検出したSERCA2 mRNA及び蛋白質レベルを図1及び図2に示す。これらのデータは、心筋のカルシウム輸送及び拡張期弛緩が、予測用動物モデルで本発明の方法を適用することにより、有利な影響を受けるという原理を証明している。
【実施例4】
【0141】
SERCA2発現アデノウイルスベクターでトランスフェクションした後の心筋細胞機能のIN VITROでの評価
簡単に言うと、新生児の心筋細胞をSERCA2アデノウイルスベクターでトランスフェクトし、実施例3で記載したようにカルシウムトランジェントを測定し、対照(トランスフェクトされていない)細胞と比べる。図3において、細胞#42は対照の新生児心筋細胞の挙動の平均を表し、細胞#27は、トランスフェクトした新生児心筋細胞の挙動の平均を表す(ADV)。図3で示されるように、カルシウムトランジェントの最大減少の半分までの時間は、対照細胞と比べてトランスフェクトした細胞で33±19%短縮された(n=4、p≦0.01)。これらのデータは、カルシウムトランジェントがSERCA2 ADVでトランスフェクトした筋細胞で増大され、その結果、in vivoで測定すれば、心筋でより速い拡張期弛緩が促進されるはずであるという原理の証拠を提供する。
【0142】
本発明のいくつかの実施形態を記載してきた。しかし、本発明の趣旨及び範囲を逸脱することなく様々に改変を加えることができることが理解されよう。従って、本発明は例示した特定の実施形態によってではなく、添付の特許請求の範囲によってのみ限定されることが理解されよう。
【図面の簡単な説明】
【0143】
【図1】(例として、アデノウイルスのSERCA2構築体を使用して開発した)マウスにおけるSERCA2導入遺伝子の発現を示したウェスタンブロットを示す図である。レーン1〜2はα−筋節アクチンの発現を示し、レーン3は、分子量マーカーを示し、レーン4〜5はSERCA2発現を示す。
【図2】SERCA2をコードする数種のアデノウイルス、すなわち、例えばCMVプロモーター(HH−1、HH−2、及びSAI−3)又はTKプロモーター(TK)の制御下にあるベクターなどのうちの1種でトランスフェクトしたマウス由来の心筋細胞におけるラットSERCA2 mRNAの存在を示したノーザンブロットを示す図である。対照の構築体は、SERCA2ポリヌクレオチドを含まなかった(ctrl)。( )内の数字は、各構築体のプラーク形成単位(pfu)の数を示し、L6細胞をアデノウイルス構築体で48時間感染させ、抽出し、細胞性RNA全体をゲル上で分離し、次いでSERCA2のcDNAをRNAにハイブリダイズさせるノーザン分析法によって決定した。
【図3】例として、アデノウイルスベクターでトランスフェクトした新生児の筋細胞において検出されたカルシウムトランジェント(平均値は細胞#27で表されている)及びトランスフェクトされていない新生児の筋細胞において検出したカルシウムトランジェント(平均値は細胞#42で表されている)を示す図である。カルシウムトランジェントは蛍光定量法で測定し、時間(x軸)の関数として表される。
【図4】例として、ベクターにSERCA2a配列を挿入するためのクローニングスキーム及び適切な制限部位を示す図である。具体的には、ラットSERCA2aを3’末端でフラッグシグナルペプチドに融着させたSERCA2a−フラッグが、ヒトCMVプロモーターのもと、XbaI/BamHI及びBgI II/KpnI制限部位に挿入される。
【図5】マウスの心臓の左心室自由壁にAAV又はアデノ随伴ウイルスを注入し、注入の2、4、6、及び8週間後に発現している核コードβgalを示す図である。
【図6】rAAV遺伝子送達4週間後に、単離して灌流させた心臓の機能測定値に対するSERCA2a−フラッグ発現の効果を甲状腺低下症マウスのGFP対照と比較して示す図である。*は、AAV−GFP処理したマウスと有意に(p<0.05)異なることを示す。rAAV−SERCA−フラッグ処理をしたマウスの機能が、AAV−GFP処理をしたマウスの機能より改善していることに留意されたい。
【特許請求の範囲】
【請求項1】
SERCA2をコードする導入遺伝子を含むアデノ随伴ウイルス(AAV)ベクターを宿主の心筋に送達することを含む、宿主の心筋細胞の筋小胞体へのSERCA2媒介カルシウムイオン輸送を増大させるための方法であって、前記導入遺伝子の発現が、処理されていない筋細胞と比べて処理された筋細胞で、SERCA2媒介カルシウムイオン輸送の増大を誘起する方法。
【請求項2】
前記AAVベクターがコロイド分散系に含まれている請求項1記載の方法。
【請求項3】
前記宿主がうっ血性心不全と診断されたことがある請求項1記載の方法。
【請求項4】
前記宿主がヒトである請求項3記載の方法。
【請求項1】
SERCA2をコードする導入遺伝子を含むアデノ随伴ウイルス(AAV)ベクターを宿主の心筋に送達することを含む、宿主の心筋細胞の筋小胞体へのSERCA2媒介カルシウムイオン輸送を増大させるための方法であって、前記導入遺伝子の発現が、処理されていない筋細胞と比べて処理された筋細胞で、SERCA2媒介カルシウムイオン輸送の増大を誘起する方法。
【請求項2】
前記AAVベクターがコロイド分散系に含まれている請求項1記載の方法。
【請求項3】
前記宿主がうっ血性心不全と診断されたことがある請求項1記載の方法。
【請求項4】
前記宿主がヒトである請求項3記載の方法。
【図1】


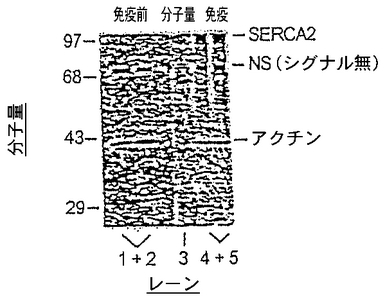
【図2】

【図3】

【図4】
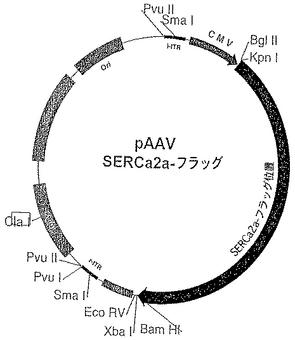
【図5】

【図6】

【図2】
【図3】
【図4】
【図5】
【図6】
【公表番号】特表2006−518996(P2006−518996A)
【公表日】平成18年8月24日(2006.8.24)
【国際特許分類】
【出願番号】特願2006−500911(P2006−500911)
【出願日】平成16年1月12日(2004.1.12)
【国際出願番号】PCT/US2004/000719
【国際公開番号】WO2004/062618
【国際公開日】平成16年7月29日(2004.7.29)
【公序良俗違反の表示】
(特許庁注:以下のものは登録商標)
Macintosh
【出願人】(304056899)ザ リージェンツ オブ ザ ユニバーシティ オブ カリフォルニア (9)
【氏名又は名称原語表記】THE REGENTS OF THE UNIVERSITY OF CALIFORNIA
【住所又は居所原語表記】1111 Franklin Street, Oakland, CA 94607−5200, United States of America
【Fターム(参考)】
【公表日】平成18年8月24日(2006.8.24)
【国際特許分類】
【出願日】平成16年1月12日(2004.1.12)
【国際出願番号】PCT/US2004/000719
【国際公開番号】WO2004/062618
【国際公開日】平成16年7月29日(2004.7.29)
【公序良俗違反の表示】
(特許庁注:以下のものは登録商標)
Macintosh
【出願人】(304056899)ザ リージェンツ オブ ザ ユニバーシティ オブ カリフォルニア (9)
【氏名又は名称原語表記】THE REGENTS OF THE UNIVERSITY OF CALIFORNIA
【住所又は居所原語表記】1111 Franklin Street, Oakland, CA 94607−5200, United States of America
【Fターム(参考)】
[ Back to top ]