
神経伝達物質センサーおよびそれを用いる方法
【課題】神経伝達物質シグナル伝達の分野に関し、より具体的には、蛍光共鳴エネルギー移動(FRET)を用いて神経伝達物質レベルの変化を測定および検出するためのバイオセンサーおよび方法の提供。
【解決手段】神経伝達物質バイオセンサーが開示され、神経伝達物質に結合した際に蛍光共鳴エネルギー移動の検出および測定を可能にするドナーおよび蛍光部分に結合した神経伝達物質結合ドメインを含むYbeJに基づくグルタミン酸結合バイオセンサーが含まれる。かかるバイオセンサーは、インビボおよび培養中の神経伝達物質濃度の検出に有用である。ここで、ドナー部分が励起され、グルタミン酸がグルタミン酸結合タンパク質部分に結合すると、ドナー部分とアクセプター部分との間の蛍光共鳴エネルギー移動(FRET)が変化する。
【解決手段】神経伝達物質バイオセンサーが開示され、神経伝達物質に結合した際に蛍光共鳴エネルギー移動の検出および測定を可能にするドナーおよび蛍光部分に結合した神経伝達物質結合ドメインを含むYbeJに基づくグルタミン酸結合バイオセンサーが含まれる。かかるバイオセンサーは、インビボおよび培養中の神経伝達物質濃度の検出に有用である。ここで、ドナー部分が励起され、グルタミン酸がグルタミン酸結合タンパク質部分に結合すると、ドナー部分とアクセプター部分との間の蛍光共鳴エネルギー移動(FRET)が変化する。
【発明の詳細な説明】
【技術分野】
【0001】
関連出願とのクロスリファレンス
本出願は、2004年10月14日出願の米国仮特許出願第60/618,179号および2005年1月14日出願の米国仮特許出願第60/643,576号からの優先権を享受し、これらはその内容全体を引用により本明細書に含める。
【0002】
本出願はまた、仮特許出願60/658,141号、仮特許出願第60/658,142号、仮特許出願第60/657,702号、PCT出願 [代理人整理番号056100-5053、“Phosphate Biosensors and Methods of Using the Same”]、PCT出願 [代理人整理番号056100-5054、“Methods of Reducing Repeat-Induced Silencing of Transgene Expression and Improved Fluorescent Biosensors”]、およびPCT出願 [代理人整理番号056100-5055、“Sucrose Biosensors and Methods of Using the Same”]にも関連し、これらはその内容全体を引用により本明細書に含める。
【0003】
連邦政府助成研究または開発に関する記述
本発明は2つの助成金から援助を受けており、それは、Duke University からのNIH 下請 (下請番号 SPSID 126632)および人間生体メカニズム解明計画助成金(契約番号 RGP0041/2004C)を含む。したがって、米国政府は本発明に対する一定の権利を有する。
【0004】
発明の分野
本発明は一般的に神経伝達物質シグナル伝達の分野に関し、より具体的には、蛍光共鳴エネルギー移動 (FRET)を用いて神経伝達物質レベルの変化を測定および検出するためのバイオセンサーおよび方法に関する。
【背景技術】
【0005】
発明の背景
本明細書中のすべての刊行物および特許出願は、個々の刊行物または特許出願が具体的かつ個別的に引用により明示されている程度に引用により本明細書にその内容を含める。
【0006】
本明細書に記載する刊行物は本出願の出願日より前のその開示を単に提供するものである。いかなる開示も本発明は以前の発明によりかかる刊行物から予測されるということを認めると解釈してはならない。
【0007】
グルタミン酸はアミノ酸であり、脳にみられる神経伝達物質の1つのタイプである。グルタミン酸作動性神経細胞は哺乳類脳において支配的な興奮性経路であり、中枢神経系におけるすべての高速興奮性シナプスの3分の1を占める(Cotman、C.W.、and Monaghan、D.T. (1986) Anatomical organization of excitatory amino acid receptors and their properties. Adv. Exp. Med. Biol. 203、237-252)。グルタミン酸によるシグナル伝達は多数かつ多様な受容体に媒介されており、以下のような受容体が挙げられる:活性化の際、共役したイオンチャンネルを通る細胞外カルシウムの通過を可能とするイオンチャンネル型受容体、および中間体分子、例えば、G タンパク質を活性化して細胞質カルシウム濃を上昇させる分子、例えば、IP3 の生産をもたらす代謝調節型受容体。神経細胞間の相互作用は興奮性または抑制性のいずれかであり得る。主な興奮性アミノ酸神経伝達物質は、グルタミン酸およびアスパラギン酸であり、一方、GABA (γ-アミノ酪酸)、グリシン (アミノ酢酸)、およびタウリンは抑制性である(Mark et al. (2001) American Journal of Neuroradiology 22:1813-1824)。
【0008】
グルタミン酸トランスポーターによる細胞外グルタミン酸のクリアランスは、グルタミン酸蓄積を妨げるために不可欠な工程であり、その工程がなければグルタミン酸蓄積により、グルタミン酸受容体の過剰刺激およびグルタミン酸興奮毒性が起こってしまう。興奮毒性損傷は、多数の神経系疾患を引き起こすかまたはかかる疾患に関与しており、神経性疾患としては、脳卒中、外傷、癲癇、および神経変性症状、例えば、ハンチントン病、エイズによる認知症、および筋萎縮性側索硬化症が挙げられる(Doble、A.、Louvel、E.、and Hugon、J. (1999) The role of excitotoxicity in neurodegenerative disease: implications for therapy、Pharmacol. Ther. 81(3):163-221; Waggie KS、Kahle PJ、Tolwani RJ. (1999) Neurons and mechanisms of neuronal death in neurodegenerative diseases: a brief review. Lab. Anim. Sci. 49:358-362)。グルタミン酸受容体過剰刺激は、イオンチャンネルを直接開けることにより細胞内カルシウムを上昇させ、カルシウムの流入を可能にし、膜脱分極を引き起こす。脱分極は次いで電位依存性カルシウムチャンネルを活性化し、それによりさらに細胞内カルシウムレベルが上昇する。グルタミン酸に誘導されたカルシウムレベルの上昇は以下を含む多数の酵素の過剰活性化を引き起こし、これらのなかには、毒性の遊離酸素ラジカルを産生するものや、神経細胞死をもたらす正のフィードバックループを引き起こすものもある:タンパク質キナーゼ C、カルシウム/カルモジュリン依存性タンパク質 キナーゼ II、ホスホリパーゼ、プロテアーゼ、ホスファターゼ、一酸化窒素シンターゼ、エンドヌクレアーゼおよびオルニチンデカルボキシラーゼ(Mark et al.、2001)。
【0009】
興奮毒性カスケードをトリガーする鍵となる因子はシナプス空間(synaptic space)におけるグルタミン酸の過剰な蓄積である。正常な細胞外グルタミン酸濃度は約 0.6μmol/Lであり、実質的な神経細胞の興奮毒性傷害は2〜5 μmol/Lのグルタミン酸濃度にて起こる。神経細胞に対する外傷性障害は、約 10 μmol/Lの細胞外空間への放出を伴う悲惨な結果をもたらしうる。後に続くカスケードを考えると、1つの神経細胞に対する傷害は、すべての近接する神経細胞を危険にさらすことになる(Mark et al.、2001)。
【0010】
多数の研究が、神経系疾患においてより高いグルタミン酸濃度が関与していることを示しているにもかかわらず、生細胞中のグルタミン酸濃度の測定はいまだに困難である。神経細胞のインビボでの機能を決定するのに必要なもっとも重要な手段の一つは、グルタミン酸の流動を直接に可視化することであろう。グルタミン酸の細胞外濃度は、インビボ微小透析技術によって測定されてきた(Faden、A.I.、Demediuk、P.、Panter、S.S.、and Vink、R. (1989) The role of excitatory amino acids and NMDA receptors in traumatic brain injury. Science 244、798-800; Fallgren、A.B.、and Paulsen、R.E. (1996) A microdyalysis study in rat brain of dihydrokainate、a glutamate uptake inhibitor. Neurochem Res 21、19-25)。しかし、微小透析は空間的および時間的分解能において制限されており、単一のシナプスの周囲の局在化した迅速な濃度変化を検出することはできない。さらに、インビボ微小透析技術は破壊的である。それはまた、生きている神経細胞または星状細胞の内側のグルタミン酸レベルの直接のモニタリングを可能にするものではない。
【0011】
蛍光共鳴エネルギー移動(FRET)の使用によるイオンおよび代謝産物のインビボでの測定を用いたカルシウム濃度変化の測定が成功しており、これはCFP、YFP、およびカルモジュリンおよびM13 ペプチドからなるレポータードメインを融合させることにより行われた(Zhang、J.、Campbell、R.E.、Ting、A.Y.、and Tsien、R.Y. (2002a) Creating new fluorescent probes for cell biology. Nat Rev Mol Cell Biol 3、906-918; Zhang、J.、Campbell、R.E.、Ting、A.Y.、and Tsien、R.Y. (2002b) Creating new fluprescent probes for cell biology. Nature Reviews Molecular Cell Biology 3、906-918)。カルシウムのカルモジュリンへの結合によりキメラの全体的な構造再編が起こり、その結果、ドナーおよびアクセプター蛍光部分によって媒介されてFRET 強度における変化が生じる。最近、基質が結合すると、2つの葉(lobe)がハエジゴクの様に閉じる多数の細菌周辺質結合タンパク質の、代謝産物 ナノセンサーの足場としての使用が成功している(Fehr、M.、Frommer、W.B.、and Lalonde、S. (2002) Visualization of maltose uptake in living yeast cells by fluprescent nanosensor. Proc. Natl. Acad. Sci. U S A 99、9846-9851; Fehr、M.、Lalonde、S.、Lager、I.、Wolff、M.W.、and Frommer、W.B. (2003) In vivo imaging of the dynamics of glucose uptake in the cytosol of COS-7 cells by fluorescent nanosensors. J. Biol. Chem. 278、19127-19133; Lager、I.、Fehr、M.、Frommer、W.B.、and Lalonde、S. (2003) Development of a fluorescent nanosensor for ribose. FEBS Lett 553、85-89)。
【0012】
グルタミン酸のためのナノセンサータンパク質を開発するために、本発明者らは、グルタミン酸が結合すると高次構造が変化するタンパク質を探索した。イオンチャンネル型グルタミン酸受容体(iGluRs)および代謝調節型グルタミン酸受容体 (mGluRs)のファミリーは、γ-アミノ酪酸 (GABA)B 受容体のリガンド結合ドメイン(Kaupmann、K.、Huggel、K.、Heid、J.、Flor、P.J.、Bischoff、S.、Mickel、S.J.、McMaster、G.、Angst、C.、Bittiger、H.、Froestl、W.、and Bettler、B. (1997) Expression cloning of GABA(B) receptors uncovers simirality to metabotropic glutamate receptors. Nature 386、239-246)および、細菌周辺質結合タンパク質に対して配列類似性を有する細胞外リガンド結合ドメインを有している (O'Hara、P.J.、Sheppard、P.O.、Thogersen、H.、Venezia、D.、Haldeman、B.A.、McGrane、V.、Houamed、K.M.、Thomsen、C.、Gilbert、T.L.、and Mulvihill、E.R. (1993) The ligand-binding domain in metabotropic glutamete receptors is related to bacterial periplasmic binding proteins. Neuron 11、41-52)。3種類の形態、1つはグルタミン酸との複合体、2つはリガンド非結合形態、にあるmGluR1 リガンド結合ドメインの結晶構造が決定され(Kunishima、N.、Shimada、Y.、Tsuji、Y.、Sato、T.、Yamamoto、M.、Kumasaka、T.、Nakanishi、S.、Jingami、H.、and Morikawa、K. (2000). Structural basis of glutamate recognition by a dimeric metaborotopic glutamate receptor. Nature 407、971-977)、その結果、グルタミン酸結合が閉じた高次構造を安定化することが示唆された。 Galvez et al. により、GABAB 受容体のリガンド結合ドメインでも2つの葉が閉じることが示唆された(Galvez、T.、Parmentier、M.L.、Joly、C.、Malitschek、B.、Kaupmann、K.、Kuhn、R.、Bittiger、H.、Froestl、W.、Bettler、B.、and Pin、J.P. (1999). Mutagenesis and modeling of the GABAB receptor extracellular domain support a venus flytrap mechanism for ligand binding. J Biol Chem 274、13362-13369)。本発明者らはそれゆえ、mGluRおよびGABAB 受容体を用いてFRET バイオセンサーの構築を試み、基質の添加によるFRET 効率の変化をアッセイした。しかし、FRET 効率の変化は観察されなかった。同様に、細菌由来のLIV ロイシン/イソロイシン/バリンアミノ酸結合タンパク質を操作しても機能的なFRET センサーは得られなかった。
【0013】
De Lorimier et alは、グルタミン-およびヒスチジン-結合タンパク質に対する配列相同性を有し、グルタミン酸代謝に関与するオペロンに位置する大腸菌由来のYbeJ タンパク質が、グルタミン酸およびアスパラギン酸に結合することを示した(de Lorimier、R.M.、Smith、J.J.、Dwyer、M.A.、Looger、L.L.、Sali、K.M.、Paavola、C.D.、Rizk、S.S.、Sadigov、S.、Conrad、D.W.、Loew、L.、and Hellinga、H.W. (2002) Construction of a fluorescent biosensor family Protein Sci 11、2655-2675)。細菌由来のグルタミンおよびヒスチジン結合タンパク質にYbeJが類似していることから、本発明者らは、これら2つのタンパク質の解明された結晶構造に基づく相同性モデルを作成した。グルタミンおよびヒスチジン結合タンパク質の3次元構造により、これらタンパク質のN-およびC- 末端が同じ葉に位置していることが示され、それゆえ基質の結合により2つの葉が閉じることは、N-およびC-末端の間の距離を変化させるわけではないようである。したがって、これらのタンパク質はいずれも同じ原理に基づくFRET センサーの構築を可能にするものではないはずである。実際、米国特許出願公開第20040118681号においてHellinga and Looger により提案されたセンサーは、単一のフルオロフォアを、リガンド結合の際の高次構造変化に応答するシステイン残基に結合させることを提案しており、これは、FRET に用いられる蛍光部分が2つであることとは全く異なる。
【発明の開示】
【発明が解決しようとする課題】
【0014】
しかし、本発明者らは驚くべきことに、大腸菌のYbeJ タンパク質が、両方の末端がタンパク質の同じ葉に位置しているという知見にもかかわらず、グルタミン酸結合の検出のための有効なFRET 足場であることを見いだした。これは、距離変化がFRET変化に変換されるという一般的な仮説に反するものである。
【課題を解決するための手段】
【0015】
発明の概要
本発明は、神経伝達物質濃度における変化を検出および測定するための神経伝達物質バイオセンサーを提供する。具体的には、本発明は、大腸菌 YbeJ由来のグルタミン酸結合タンパク質部分を含むグルタミン酸結合蛍光指標 (FLIP-E) をコードする単離核酸を提供し、ここで、該グルタミン酸結合タンパク質部分は遺伝子的にドナー蛍光タンパク質部分およびアクセプター蛍光タンパク質部分と融合しており、ここで、ドナー部分とアクセプター部分との間の蛍光共鳴エネルギー移動 (FRET)は、ドナー部分が励起され、グルタミン酸がグルタミン酸結合タンパク質部分に結合した場合に変化する。本発明の核酸を含むベクター、例えば、発現ベクターおよび宿主細胞も提供され、また、該核酸によりコードされるバイオセンサータンパク質も提供される。かかる核酸、ベクター、宿主細胞およびタンパク質は、神経伝達物質レベル、特に神経細胞サンプル中の細胞外グルタミン酸レベルの変化の検出方法、およびグルタミン酸興奮毒性を調節する化合物の同定方法に使用することができる。
【0016】
図面の簡単な説明
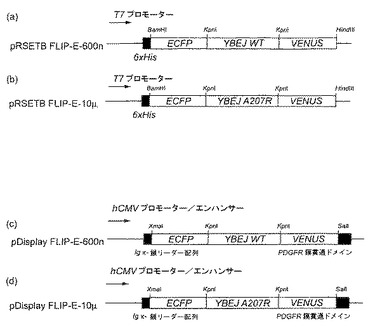
図 1.大腸菌 (AおよびB)および神経細胞培養物 (CおよびD) における発現に用いたFLIP-Eナノセンサーコンストラクト。
【0017】
図 2.3種類のグルタミン酸濃度でのFLIP-E 600n センサー (グルタミン酸に対するKdが600 nMである蛍光グルタミン酸ナノセンサー)のスペクトル: 0 mM (黒)、Kd 値の濃度(青)、および飽和濃度(赤)。これら曲線は520nmの等吸収点を共有する。
【0018】
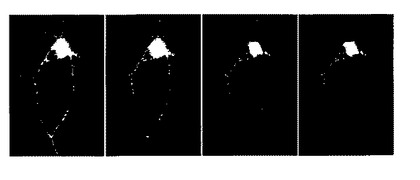
図 3. 1 mg/ml トリプシンで処理された海馬細胞。画像(A-D)は10 秒間隔で取得した。細胞表面上のシグナルが大きく消滅していることに注目されたい。
【0019】
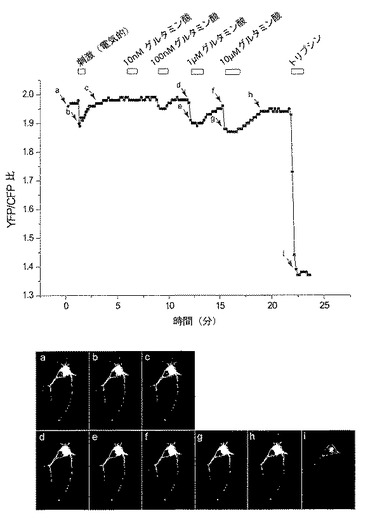
図 4. FLIP-E 600n センサーを発現する海馬細胞における発光強度比変化。画像は発光強度比変化を示すために疑似カラー表示している。グラフ (A)の上の白いバーは処理(刺激 /グルタミン酸の灌流)の時点を示す。矢印で示す時点での比画像(ratio image)をパネル(B)、a〜iに示す。発光強度比の変化が、電気的刺激とグルタミン酸の灌流との両方において観察された。比変化は低レベルの基質 (10 nM グルタミン酸)での灌流では観察されなかった。
【0020】
図 5. FLIP-E 10μ センサー(グルタミン酸に対するKdが10 μMである蛍光グルタミン酸ナノセンサー)を発現する海馬細胞における発光強度比変化。グラフ (A)の上の白いバーは、処理 (刺激 / グルタミン酸の灌流)の時点を示す。矢印で示す時点での比画像をパネル(B)、a〜gに示す。電気的刺激によっては発光強度比に大きい変化は起こらなかったが、100 μM グルタミン酸での灌流によると可逆的な比変化が引き起こされた (パネル (B)、c およびe)。
【0021】
発明の詳細な説明
以下の記載は本発明の理解に有用であり得る情報を含む。本明細書により提供される情報はいずれも従来技術であるとも本願請求項の発明と関連するものとも認めるものではなく、明示的または黙示的に引用するいずれの刊行物も従来技術であると認めるものではない。
【0022】
本発明のその他の目的、利点および特徴が本出願において提供する明細書および図面をみることにより当業者に明らかとなる。したがって、本発明のさらなる目的および利点は以下の記載から明らかであろう。
【0023】
バイオセンサー
本発明は、蛍光共鳴エネルギー移動 (FRET)を用いて神経伝達物質濃度の変化を検出および測定するための神経伝達物質バイオセンサーを提供する。神経伝達物質として作用する物質の3つの主なカテゴリーは以下の通りである:(1) アミノ酸 (主にグルタミン酸(glutamic acid または glutamate)、GABA、アスパラギン酸およびグリシン)、(2) ペプチド(バソプレッシン、ソマトスタチン、ニューロテンシン等) および (3) モノアミン (ノルエピネフリン、ドーパミンおよびセロトニン)ならびにアセチルコリン。具体的には、本発明は、グルタミン酸結合蛍光指標、特に、大腸菌グルタミン酸/アスパラギン酸受容体であるYbeJ由来のグルタミン酸結合タンパク質部分を含む指標を提供する。上記の神経伝達物質についてのさらなる神経伝達物質バイオセンサーも本明細書において提供するコンストラクトおよび方法を用いて調製することが出来る。
【0024】
したがって、本発明は、神経伝達物質に結合する蛍光指標をコードする単離核酸を提供する。とりわけ、一つの態様は、以下を含むグルタミン酸結合蛍光指標をコードする単離核酸である: グルタミン酸結合タンパク質部分、遺伝子的にグルタミン酸結合タンパク質部分と融合したドナー蛍光タンパク質部分、および遺伝子的にグルタミン酸結合タンパク質部分と融合したアクセプター蛍光タンパク質部分、ここでドナー部分とアクセプター部分との間のFRETは、ドナー部分が励起され、グルタミン酸がグルタミン酸結合タンパク質部分に結合すると変化する。好ましいグルタミン酸結合タンパク質部分は大腸菌 YbeJ由来のグルタミン酸結合タンパク質部分である。
【0025】
YbeJは当該技術分野においてYzzKおよびGltIとしても知られており、その DNA 配列 (配列番号1)およびタンパク質配列 (YbeJ、タンパク質受入番号 NP_415188、配列番号2) は既知である。グルタミン酸結合領域をコードするYbeJ DNA 配列のいずれの部分を本発明の核酸において用いてもよい。例えば、本発明の核酸における使用に好適な一つの領域は配列番号3により提供され、この配列は切型(truncated)グルタミン酸-アスパラギン酸結合タンパク質配列 (配列番号4)をコードし、シグナルペプチドを含まない成熟タンパク質をコードするものである。他の細菌種からの天然のホモログも用いることが出来、例えば緑膿菌(Pseudomonas aeruginosa )由来のPA5082遺伝子が挙げられ、その遺伝子産物は大腸菌のYbeJ タンパク質と70% 類似する。YbeJまたはそのいずれかのホモログのグルタミン酸結合部分を本明細書に記載するベクターにクローニングし、開示するアッセイにしたがって活性についてスクリーニングすればよい。
【0026】
測定可能なグルタミン酸結合機能を保持する、部位特異的突然変異、欠失または挿入を含む人工的に遺伝子操作された変異体に加えて、YbeJの天然の種変異体も用いることが出来る。本発明の核酸コンストラクトにおける使用に好適な変異体核酸配列は、好ましくは YbeJの遺伝子配列に対して少なくとも 70、75、80、85、90、95、または99%の類似性または同一性を有するであろう。好適な変異体核酸配列はまた、高度にストリンジェントなハイブリダイゼーション条件下でYbeJ遺伝子にハイブリダイズしうる。高度にストリンジェントな条件は当該技術分野において知られている;例えば、 Maniatis et al.、Molecular cloning: A Laboratory Manual、2d Edition、1989、および Short Protocols in Molecular Biology、ed. Ausubel、et alを参照されたい:これらはいずれも引用により本明細書に含める。ストリンジェントな条件は配列に依存し、異なる環境では異なるものとなろう。より長い配列はより高い温度で特異的にハイブリダイズする。核酸のハイブリダイゼーションについての詳細な説明は、Tijssen、Techniques in Biochemistry and Molecular Biology--Hybridyzation with Nucleic Acid Probes、"Overview of principles of Hybridyzation and the strategy of nucleic acid assays" (1993)にみられる。一般に、ストリンジェントな条件は規定されたイオン強度およびpHにおける特定の配列についての熱融解温度 (Tm)よりも約 5-10℃低くなるよう選択する。 (規定されたイオン強度、pHおよび核酸濃度における) Tmは、標的に相補的なプローブの50%が平衡状態で標的配列にハイブリダイズする温度である(Tm において標的配列が過剰に存在する場合、プローブの50%が平衡状態で占有される)。ストリンジェントな条件は、塩濃度が約 1.0M未満のナトリウムイオン、典型的には約 0.01〜1.0Mのナトリウムイオン濃度 (またはその他の塩)、pH 7.0 〜 8.3、そして短いプローブ(例えば、10〜 50 ヌクレオチオド)については少なくとも 約 30℃の温度、長いプローブ(例えば、50を超えるヌクレオチオド)については少なくとも 約 60℃の温度であろう。ストリンジェントな条件はまた、脱安定化剤、例えば、ホルムアミドを添加して達成してもよい。
【0027】
本発明の好ましい人工変異体は、YbeJに基づく、およびその他のグルタミン酸ナノセンサーにより測定できる濃度範囲を広げるために、グルタミン酸に対する親和性が上昇または低下しているものであり得る。とりわけ好ましい人工変異体としては、以下の突然変異を含むグルタミン酸結合領域が挙げられる:A207G、A207P、A207K A207M、A207S、A207C、A207R、A207V、A207L、A207Q、A207T、A207F、A207Y、A207N、A207W、A207H、A207D、および/またはS95W。グルタミン酸に対する結合親和性が低下または上昇しているさらなる人工変異体は、ランダムまたは部位特異的突然変異誘発およびその他の突然変異誘発技術により構築でき、本明細書に記載するベクターにクローニングし、開示するアッセイにしたがって活性についてスクリーニングすればよい。
【0028】
本発明の単離核酸は、組み合わせた場合にFRETにおけるドナーおよびアクセプター部分として作用することができるいずれの好適なドナーおよびアクセプター蛍光タンパク質 部分を組み込んでいてもよい。好ましいドナーおよびアクセプター部分は、GFP (緑色蛍光 タンパク質)、CFP (シアン蛍光タンパク質)、BFP (青色蛍光タンパク質)、YFP (黄色蛍光タンパク質)、およびそれらの増強された変異体からなる群から選択され、特に好ましい態様はドナー/アクセプター対CFP/YFP-Venusにより提供され、YFP-VenusはpH 耐性および熟成時間が改良されたYFPの変異体である (Nagai、T.、Ibata、K.、Park、E.S.、Kubota、M.、Mikoshiba、K.、and Miyawaki、A. (2002) A variant of yellow fluprescent protein with fast and efficient maturation for cell-biological applications. Nat. Biotechnol. 20、87-90)。あるいは好ましくはより高いpH 安定性およびより大きいスペクトル分離を示すMiCy/mKO 対である(Karasawa S、Araki T、Nagai T、Mizuno H、Miyawaki A. Cyan-emitting and/or ange-emitting fluprescent proteins as a donor/acceptor pair for fluorescence resonance energy transfer. Biochem J. 2004 381:307-12)。ドナーおよびアクセプター蛍光部分を選択する際に考慮すべき基準は当該技術分野において知られており、例えばUS 6,197,928に開示されており、その全体を引用により本明細書に含める。
【0029】
ドナーまたはアクセプターのいずれかとして好適なものとしてはまた、 Discosoma 種からのネイティブな DsRed、別の属からのDsRedのオルソログまたはネイティブな DsRedの性質を最適化した変異体(例えば、K83M 変異体またはDsRed2 (Clontechから入手可能))が挙げられる。 本明細書において用いる場合、「変異体」という用語は、ネイティブな蛍光分子に対して少なくとも 約 70%、より好ましくは少なくとも 75%の同一性、例えば、少なくとも 80%、90%、95%またはそれ以上の同一性を示すポリペプチドをいう。多くのかかる変異体が当該技術分野において知られているし、あるいはネイティブな蛍光分子のランダムまたは指向性突然変異誘発により容易に調製することができる(例えば、Fradkov et al.、FEBS Lett. 479:127-130 (2000)を参照)。
【0030】
バイオセンサーのフルオロフォアが(1または複数の)類似または関連配列のストレッチを含む場合、本発明者らは最近、遺伝子サイレンシングが特定の細胞および特に生物全体におけるバイオセンサーの発現に悪影響を与えうることを見いだした。かかる状況においては、フルオロフォアをコードする配列を1以上の各フルオロフォアのコドンの縮退またはゆらぎ位置にて改変し、フルオロフォアの核酸配列が改変されるがコードされるアミノ酸配列は変化しないようにすることが可能である。あるいは、フルオロフォアの機能に悪影響を与えない1以上の保存的置換を導入してもよい。PCT出願 [代理人整理番号056100-5054、“Methods of Reducing Repeat-Induced Silencing of Transgene Expression and Improved Fluorescent Biosensors”]を参照されたい。これはその全体を引用により本明細書に含める。
【0031】
本発明はさらに、神経伝達物質バイオセンサーポリペプチドをコードする単離核酸分子を含むベクターを提供する。例示的なベクターとしては、以下が挙げられる:ウイルス、例えば、バクテリオファージ、バキュロウイルスまたはレトロウイルス由来のベクター、および細菌または細菌配列とその他の生物由来の配列との組合せ由来のベクター、例えば、コスミドまたはプラスミド。かかるベクターとしては、神経伝達物質バイオセンサーをコードする核酸配列に作動可能に連結した発現制御配列を含む発現ベクターが挙げられる。ベクターは原核細胞、例えば、大腸菌またはその他の細菌において機能するのに適したものでもよいし、真核細胞、例えば酵母および動物細胞において機能するのに適したものでもよい。例えば、本発明のベクターは一般に以下のような要素を含むであろう:例えば、所望の宿主細胞に適合する複製起点、所望の宿主細胞に適合する1以上の選択可能なマーカーおよび1以上のマルチクローニングサイト。ベクターに含まれる特定の要素の選択は以下のような因子に依存するであろう:例えば、目的の宿主細胞、インサートのサイズ、挿入された配列の制御された発現が望まれるか否か、即ち、例えば、誘導可能または調節可能なプロモーターの使用を介するべきか否か、目的のベクターのコピー数、目的の選抜系等。異なる用途のための宿主細胞とベクターとの適合性を確証するのに関与する因子は当該技術分野において周知である。
【0032】
本発明において使用するために好ましいベクターは、ドナーおよびアクセプター蛍光分子をコードする核酸の間への神経伝達物質結合ドメインまたは受容体のクローニングを可能にし、その結果、ドナーおよびアクセプター蛍光分子に遺伝子的に融合した神経伝達物質結合ドメインを含むキメラまたは融合タンパク質の発現をもたらすものであろう。例示的なベクターとしては、細菌 pRSET-FLIP誘導体が挙げられ、これはFehr et al. (2002) (Visualization of maltose uptake in living yeast cells by fluprescent nanosensor. Proc. Natl. Acad. Sci. U S A 99、9846-9851)に開示されており、これはその全体を引用により本明細書に含める。あるいは、目的の神経伝達物質結合ドメインをまず蛍光ドナーおよびアクセプターをコードする配列に融合させ、その後で適当なベクターにクローニングしてもよく、これは米国特許第6,596,499号に記載されており、その全体を引用により本明細書に含める。
【0033】
本発明のキメラ核酸は好ましくは、ドナーおよびアクセプター間のFRETの変化が神経伝達物質の結合の際に検出されうるように、ドナーおよびアクセプター蛍光部分をコードする配列が、神経伝達物質結合ドメインの別々の末端に融合するように構築する。あるいは、該ドナーフルオロフォアおよび/または該アクセプターフルオロフォア部分の一方または両方をリガンド結合タンパク質部分に、該リガンド結合タンパク質部分の内部にて融合させてもよい。かかる融合は仮出願第60/658,141号に記載されており、これは引用により本出願に含める。ドナーおよびアクセプター部分はタンパク質の同じドメインまたは葉に含めてもよいが、好ましくは、ドナーおよびアクセプター部分はタンデムには融合されるべきではない。ドメインとは、特定の機能を担うタンパク質の部分であり、典型的には少なくとも 約 40〜約 50 アミノ酸の長さである。一つのタンパク質に複数のタンパク質ドメインが含まれうる。
【0034】
蛍光ドメインは所望の場合は1以上の可動性リンカー配列によって、神経伝達物質結合ドメインから離れていてもよい。かかるリンカー部分は、好ましくは約 1〜50 アミノ酸残基の長さであり、より好ましくは約 1〜30 アミノ酸残基の長さである。リンカー部分およびその用途は当該技術分野において周知であり、例えば、米国特許第5,998,204号および5,981,200号、およびNewton et al.、Biochemistry 35:545-553 (1996)に記載されている。あるいは、フルオロフォアまたは結合タンパク質を短くした形態のものを用いてもよい。
【0035】
例えば、本発明者らは、結合ドメインのコアタンパク質構造とフルオロフォアとを連結する配列を除くことにより、即ち、リンカー配列を除くことによるか、および/または、分析対象結合部分および/またはフルオロフォアの末端からアミノ酸を欠失させることにより、フルオロフォアのより近い結合 (coupling)が達成され、その結果比変化が大きくなることも見いだした。好ましくは、欠失は、リンカーまたはフルオロフォアまたはリガンド結合ドメインをコードする領域に位置するFRET バイオセンサーをコードする核酸コンストラクトにおいて少なくとも1、または少なくとも2、または少なくとも3、または少なくとも4、または少なくとも5、または少なくとも8、または少なくとも10、または少なくとも15ヌクレオチオドを欠失させることにより行う。異なる領域における欠失を一つのコンストラクトにおいて組み合わせることにより硬直性(rigidity)が増強した2以上の領域を作ることができる。アミノ酸をまた、付加または突然変異させて、例えば、プロリン残基またはその他の好適なアミノ酸を付加することによって捻れを導入することによって、バイオセンサーの硬直性を増強させ、感度を向上させることも出来る。感度の上昇はリガンド結合の際のFRET 蛍光の比変化によって測定でき、好ましくは感度は、該(1または複数の)欠失の結果少なくとも2倍上昇する。仮出願第60/658,141号を参照されたい。これはその全体を引用により本明細書に含める。
【0036】
本発明はまた、本発明のベクターまたは発現ベクターによってトランスフェクトされた宿主細胞も含み、例えば、原核細胞、例えば、大腸菌またはその他の細菌、あるいは真核細胞、例えば、酵母細胞または動物細胞が挙げられる。別の側面において、本発明は、神経伝達物質バイオセンサーの発現をコードする核酸配列の発現によって特徴づけられる表現型を有するトランスジェニック非ヒト動物を特徴とする。表現型は以下によって作成しうる動物の体細胞および生殖細胞に含まれる導入遺伝子によって与えられる:(a)動物の接合体(zygote)に、神経伝達物質バイオセンサーをコードするDNA コンストラクトを含む導入遺伝子を導入すること; (b)接合体を偽妊娠動物に移植すること; (c)接合体を出産(term)まで発生させること;および(d)導入遺伝子を含む少なくとも1つのトランスジェニック子孫を同定すること。導入遺伝子を胚に導入する工程は、導入遺伝子を含む胚性幹細胞を胚に導入すること、または、導入遺伝子を含むレトロウイルスを胚に感染させることによって行うことが出来る。好ましいトランスジェニック動物は脳において、コードされた神経伝達物質バイオセンサーを発現するであろう。本発明のトランスジェニック動物には、トランスジェニック線虫およびトランスジェニックマウスおよびその他の動物が含まれる。
【0037】
本発明はまた、本明細書に記載する性質を有する単離神経伝達物質バイオセンサー分子、特に、YbeJに基づくグルタミン酸結合蛍光指標も含む。かかるポリペプチドは、本明細書に記載する核酸コンストラクトを用いて組換えにより発現させてもよいし、成分となるドメインのいくつかまたはすべてを化学的に結合させることにより製造してもよい。発現したポリペプチドは、転写翻訳系または組換え細胞において産生させてもよいし、および/または転写翻訳系または組換え細胞から、当該技術分野において知られている生化学的および/または免疫学的精製方法によって単離してもよい。本発明のポリペプチドは、脂質二重層、例えば、細胞膜抽出物、または人工脂質二重層 (例えば、リポソーム小胞) またはナノ粒子に導入してもよい。
【0038】
神経伝達物質レベルの検出方法
本発明の核酸およびタンパク質は、動物の脳または神経系における神経伝達物質のレベルの変化、特にグルタミン酸興奮毒性に関連する障害または疾患のシグナルであり得る細胞外グルタミン酸のレベルの変化の検出および測定に有用である。一つの態様において、本発明は、以下を含む神経細胞のサンプルにおける細胞外グルタミン酸のレベルの変化を検出する方法を含む:(a)本明細書に記載するグルタミン酸結合バイオセンサーをコードする核酸を発現する細胞、および、神経細胞のサンプルを提供すること;および(b)それぞれグルタミン酸結合ドメインに共有結合している、ドナー蛍光タンパク質部分とアクセプター蛍光タンパク質部分との間のFRETの変化を検出すること、ここで、該ドナー部分と該アクセプター部分との間のFRETの変化は、神経細胞のサンプル中の細胞外グルタミン酸レベルの変化を示す。あるいは、タンパク質は異種宿主、例えば細菌で産生させ、精製し、器官に直接、または細胞間空間に注入してもよい。タンパク質またはその誘導体は、量子ドットなどの粒子に結合させて、細胞または区画に導入してもよい。
【0039】
FRETは当該技術分野において知られている様々な技術を用いて測定することが出来る。例えば、FRETの判定工程は、アクセプター蛍光タンパク質部分から放射される光の測定を含みうる。あるいは、FRETの判定工程は、ドナー蛍光タンパク質部分から放射される光を測定すること、アクセプター蛍光タンパク質部分から放射される光を測定すること、および、ドナー蛍光タンパク質部分から放射される光とアクセプター蛍光タンパク質部分から放射される光の比を計算することを含みうる。FRETの判定工程はまた、ドナー部分の励起状態寿命または異方性変化を測定することを含みうる (Squire A、Verveer PJ、Rocks O、Bastiaens PI. J Struct Biol. 2004 Jul;147(1):62-9 Red-edge anisotropy microscopy enables dynamic imaging of homo-FRET between green fluorescent protein in cells)。かかる方法は当該技術分野において知られており、US 6,197,928に一般的に記載されており、これはその全体を引用により本明細書に含める。
【0040】
神経細胞のサンプル中のグルタミン酸またはその他の神経伝達物質の量はFRETの程度を測定することにより決定することが出来る。第一に、FLIP-E センサーをサンプルに導入しなければならない。神経伝達物質濃度の変化は、所定の時間間隔でFRET 変化をモニタリングすることにより判定することが出来る。サンプル中の神経伝達物質の量は、例えばインビボでの滴定 (titration)により確立した検量線を用いて定量することが出来る。
【0041】
本発明の方法により分析すべき神経細胞サンプルは、インビボ、例えば、海馬神経細胞の表面上でのグルタミン酸流出の測定において含めてもよいし、または、グルタミン酸流出が神経細胞培養物にて測定される、インビトロに含めてもよい。あるいは、脳または1以上のシナプス空間からの液体抽出物を細胞外神経伝達物質が検出または測定されるサンプルとして用いてもよい。かかる測定は、該神経細胞に対する外傷性障害に伴う細胞外グルタミン酸の検出に用いてもよいし、グルタミン酸興奮毒性に関する神経障害、例えば、特に、脳卒中、癲癇、ハンチントン病、エイズによる認知症および筋萎縮性側索硬化症の可能性の指標として用いてもよい。
【0042】
本明細書に開示する神経伝達物質レベルの検出方法は、神経伝達物質濃度の調節に使用しうる化合物、特に、グルタミン酸興奮毒性の調節に有用な化合物のスクリーニングおよび同定に使用することができる。一つの態様において、とりわけ、本発明は以下を含むグルタミン酸興奮毒性を調節する化合物の同定方法を含む:(a)本明細書に開示するグルタミン酸バイオセンサーを発現する細胞および神経細胞のサンプルを1以上の被験化合物と接触させること、および(b)該ドナー蛍光ドメインと該アクセプター蛍光ドメインの間のFRETを該接触の後に測定すること、ここで該接触後のFRETの上昇または低下は、該被験化合物がグルタミン酸興奮毒性を調節する化合物であることを示す。「調節する」という用語は、かかる化合物がグルタミン酸興奮毒性を上昇または低下させうることを意味する。グルタミン酸レベルを上昇させる化合物は上記のようなグルタミン酸興奮毒性に関連する障害の治療的介入および処置のための標的である。グルタミン酸レベルを低下させる化合物はグルタミン酸興奮毒性に関連する障害の処置のための治療用品へと開発することが出来る。
【0043】
原形質膜の外側の葉(leaflet)に対するセンサーの標的化は、可能性のある用途の一つの態様に過ぎない。それは、ナノセンサーは特定の区画に標的化することが出来ることを示す。あるいは、その他の標的化配列を用いて、センサーをその他の区画、例えば、小胞、ER、液胞、などに発現させることが出来る。
【0044】
発現系にはラット神経細胞のみならず、ヒト細胞株、動物細胞および器官、真菌および植物細胞も含まれる。センサーはまた、グルタミン酸が重要な窒素化合物として役割を果たすのみならず、おそらくはシグナル伝達分子としても作用する、真菌および植物生物体におけるグルタミン酸レベルをモニターするのにも用いることが出来る。細菌における発現を用いて、感染部位または細菌が存在するかまたは導入される区画におけるグルタミン酸レベルをモニターすることができる。
【0045】
具体的には、センサーを発現する細菌または真菌は、バイオセンサーとして働くことができ、あるいは、上記の薬物スクリーニングについて記載したものと同様のスキームを用いて、新規殺虫剤の同定のためのツールとして働くことが出来る。
【0046】
さらなる有用性
本発明のバイオセンサーは、神経細胞の機能を判定するために動物細胞の表面に発現させてもよい。例えば、線虫において存在する神経細胞の多くはいまだに特定の機能がわかっていない。表面上でのバイオセンサーの発現により、刺激に応答した生きた線虫における神経細胞活性の可視化が可能となり、それにより神経細胞回路網の機能の決定および分析が可能となる。同様に、生きたマウスまたはラットの脳に多光子プローブを導入することにより、それらのプロセスを画像化することが可能となる。最後に、特定の神経細胞または膠細胞における発現により、 脳卒中またはアルツハイマー病などの現象の研究、および神経細胞の内側またはその表面におけるグルタミン酸レベルへのかかる障害の効果の研究が可能となる。さらに、局所脳領域または神経細胞回路網への薬物療法の効果をインビボで研究することが出来る。
【0047】
最後に、センサーを、例えば膜または人工脂質複合体上に提示される人工グルタミン酸捕捉剤として導入することにより、グルタミン酸流動を改変するツールとしてセンサーを使用することが可能であり、したがって脳または神経細胞機能を操作することが可能である。
【0048】
以下の実施例は本発明を説明および例示するために提供する。したがって、以下の実施例は本発明の範囲を限定するものと解釈してはならない。当業者であれば多くのその他の態様が本明細書および請求の範囲に記載する本発明の範囲内に包含されることを理解しているであろう。
【実施例】
【0049】
実施例 1. 核酸およびベクターの構築
シグナルペプチドを有さない成熟タンパク質をコードする切型グルタミン酸-アスパラギン酸結合タンパク質配列 (配列番号4)を、大腸菌 ゲノム DNAをテンプレートとして用いてPCRにより増幅した。用いたプライマーは、5’- ggtaccggaggcgccgcaggcagcacgctggacaaaatc -3’(配列番号5) および5’- accggtaccggcgccgttcagtgccttgtcattcggttc - 3’(配列番号6)であった。 PCR 断片をpRSET ベクター (Invitrogen)中のFLIPmal-25μ (Fehr et al. 2002)のKpnI 部位にクローニングし、マルトース結合タンパク質配列をYBEJ 配列に交換した。その結果得られたプラスミドをpRSET-FLIP-E-600nと命名した。
【0050】
pHおよびクロライド耐性を向上させ、センサータンパク質の成熟を向上させるために、pRSET-FLIP-E-600n 中の増強 YFP (EYFP、CLONTECH) 配列を含む断片をVenusの コード配列で置換した。VenusはpH 耐性と熟成時間が改良したYFPの変異体である(Nagai、T.、Ibata、K.、Park、E.S.、Kubota、M.、Mikoshiba、K.、and Miyawaki、A. (2002) A mutant of yellow fluorescent protein with fast and efficient maturation for cell-biological applications. Nat. Biotechnol. 20、87-90)。以下の置換を有する親和性突然変異体:A207G、A207P、A207K、A207M、A207S、A207C、A207R、A207V、A207L、A207Q、A207T、A207F、A207Y、A207N、A207W、A207H、A207D 、またはS95W:を部位特異的突然変異誘発により作成した (Kunkel、T.A.、Roberts、J.D.、and Zakour、R.A. (1987). Rapid and efficient site-specific mutagenesis without phenotypic selection. Methods Enzymol. 154、367-382)。
【0051】
pRSET-FLIP-E コンストラクトをエレクトロポレーションを用いて大腸菌 BL21(DE3)Gold (Stratagene) に移した(Sambrook、J.、Fritsch、E.F.、and Maniatis、T. (1989). Molecular Cloning. A laboratory manual. (Cold Spring Harbor NY: Cold Spring Harbor Laboratory Press)。BL21(DE3)Gold 株で発現したFLIP-E タンパク質を以前に記載されたようにして抽出し、精製した (Fehr et al. 2002)。ラット初代神経細胞培養物およびPC12 細胞培養物における発現のために、FLIP-E 600nおよび-10μ カセットを以下にようにしてpDisplay (Invitrogen)にクローニングした: XmaI 部位 およびSalI 部位をPCRによりFLIP-E カセットの5’- および3’-末端にそれぞれ導入した。用いたプライマーは、5’- gagcccgggatggtgagcaagggcgaggag -3’(配列番号7)および5’- gaggtcgaccttgtacagctcgtccatgccgag -3' (配列番号8)であった。PCR 断片を配列決定してPCRエラーの追加がないことを確認し、XmaI/SalIで消化し、pDisplay ベクターのXmaI/SalI 部位にクローニングした。細胞培養物に改変リン酸カルシウム トランスフェクションプロトコール(Xia、Z.、Dudek、H.、Miranti、C.K.、and Greenberg、M.E. (1996). Calcium influx via the NMDA receptor induces immediate early gene transcription by a MAP kinase/ERK-dependent mechanism. J. Neurosci. 16、5425-5436)またはLipofectamine (Invitrogen)を用いてトランスフェクトした。
【0052】
実施例 2. FLIP-E ナノセンサーのインビトロ特徴決定
成熟 YBEJ タンパク質をコードするDNA 断片をN- およびC-末端にてそれぞれECFPおよびVenus 配列に融合させた(図 1)。発光スペクトルおよび基質滴定曲線をモノクロメータ マイクロプレートリーダー Safire (Tecan、Austria)を用いて得た。 励起フィルターは433±12nmであり、CFP および YFP 発光についての発光フィルターはそれぞれ485±12、528nm±12nmであった。すべての分析は20mM リン酸ナトリウムバッファー、pH 7.0中で行った。
【0053】
グルタミン酸の添加の結果、CFP 発光が上昇し、YFP 発光が低下し、これは、グルタミン酸のYBEJへの結合の結果、フルオロフォアの双極子の配向の相対変化におそらく起因してキメラタンパク質の高次構造変化が起こったことを示唆する(図 2)。CFPおよびYFP 部分は同じ葉に結合していると考えられることから、本発明者らは、グルタミン酸結合は2つのフルオロフォアの双極子-双極子角の変化をもたらすと推測する。興味深いことに、比および比変化はいままでに作成されたその他のセンサーと比べて同様の範囲内であり(Fehr et al.、2002; Fehr et al.、2003; Lager et al.、2003)、これは距離変化はFRET 変化のメカニズムの根底にある第一の因子ではないであろうということを示唆する。3種類のグルタミン酸濃度(0、Kd、飽和)でのスペクトルにより520 nmの等吸収点が明らかである (図 2)。グルタミン酸についての結合定数 (Kd)は600nMであると判明し、その他の方法によって得られたデータと一致した(de Lorimier et al.、2002)。アスパラギン酸、グルタミン、アスパラギンについての結合定数はそれぞれ1μM、100μM、300μMであると判明した(以下の表1参照)。
【0054】
YBEJに基づくグルタミン酸ナノセンサーにより測定可能な濃度範囲を広げるために、YBEJ 部分に突然変異を誘発してグルタミン酸に対する親和性がより小さいナノセンサーを作成した。リガンド結合部位を形成するドメイン間間隙の周辺に位置する部位(「周囲立体(peristeric)」と称される)に様々なフルオロフォアを結合させることにより、周辺質結合タンパク質におけるリガンド結合親和性が変化することが以前に示されている(de Lorimier et al.、2002)。試験した残基のなかで、アラニン 207からリジン、メチオニン、セリン、システイン、アルギニン、バリン、ロイシン、グルタミン、スレオニン、フェニルアラニン、チロシン、アスパラギン、トリプトファン、ヒスチジン、アスパラギン酸への突然変異は結合親和性を有意に低下させた(表 1)。さらに、セリン 118から、グルタミン酸の窒素と相互作用することが示唆されるトリプトファンへの突然変異は、タンパク質の親和性を低下させることが判明した。したがって、FLIPE ナノセンサーに導入した突然変異により生理的グルタミン酸濃度の広い範囲を包含する好適な親和性突然変異体を作ることが出来る。
【0055】
表 1.
【表1】

【0056】
実施例 3. FLIP-Eのインビボ特徴決定
FLIP-E ナノセンサーのインビボ特徴決定のために、FLIPE-600n およびFLIPE-10μを哺乳類発現ベクター pDisplay (Invitrogen、USA)にクローニングした。pDisplay ベクター はタンパク質を分泌経路に向けるリーダー配列、および、タンパク質を原形質膜にアンカーする膜貫通ドメインを含み、タンパク質を細胞外表面に提示する。ラット海馬細胞およびPC12 細胞にpDisplay FLIPE-600nおよび -10μ コンストラクトをトランスフェクトした。FRETをトランスフェクションの24 - 48時間後に冷却 CoolSnap HQ デジタルカメラ (Photometrics)を備えた蛍光顕微鏡 (DM IRE2、Leica)で画像化した。二重(dual)発光強度比を436nmでの励起の後に同時に記録し、OI-5-EM フィルターセット (Optical Insights) およびMetafluor 6.1r1 software (Universal Imaging)を備えたDualViewによりCFPとVenus 発光とを分割した。
【0057】
FLIP-Eの発現はラット海馬細胞培養の原形質膜上に観察され、ある程度はおそらくは原形質膜タンパク質の原形質膜への標的化に関与する区画である細胞内区画にも観察された。1 mg/mL のトリプシンを含むタイロードバッファーで処理した場合、細胞表面の蛍光の大部分は消失し、pDisplay コンストラクトの性質から予測されるように、 FLIPE タンパク質が実際に原形質膜の細胞外表面に提示されていることが示された(図 3)。ナノセンサーはしたがって細胞表面近くの細胞外グルタミン酸レベルを測定しているはずである。
【0058】
CFPおよびVenus 発光の強度を定量するために、細胞周辺の2つのチャンネルにおける蛍光強度をピクセル毎に(pixel-by-pixel basis)積分し (integrated)、CFP/Venus 比を算出した。表面にFLIPE-600nを提示する海馬細胞に電流パルスを通すことにより電気的に刺激すると、CFP/Venus 発光比の低下が観察され(図 4 a-c)、グルタミン酸が膜脱分極によって海馬細胞から放出されていることが示唆された。比変化がグルタミン酸の細胞外濃度の変化に起因することを確認するために、細胞に様々な濃度のグルタミン酸を灌流した。発光強度比は濃度依存的に変化し(図 4 d-h)、細胞表面に提示されたFLIPE-600n が細胞外グルタミン酸を認識していることが示された。FLIP-E 600n センサー の動作範囲は100nM〜1μMであり、これはFLIPE-600n ナノセンサーのインビトロ動作範囲と一致する。CFP/Venus 比は外部培地を灌流により除くと上昇し、FRET 強度のインビボ変化は可逆的であることが示唆された。
【0059】
FLIPE/600n センサーを発現する細胞とは対照的に、CFP/Venus 発光強度変化はFLIPE-10μを発現する細胞において電気刺激によっては観察されなかった(図 5)。しかし比変化は細胞をより高濃度のグルタミン酸で灌流した場合に観察され(図 5 cおよびe)、細胞の脱分極により誘導されるグルタミン酸濃度変化はFLIP-E 10μ センサーの動作範囲を下回ることが示唆された。
【0060】
新規ナノセンサーはしたがって神経細胞表面のグルタミン酸を測定することが出来、前シナプス神経細胞のグルタミン酸分泌を直接追跡することが出来る。
【0061】
本明細書に記載するすべての刊行物、特許および特許出願は引用により本出願にその内容を含める。本発明を特定の態様に関して記載してきたが、さらなる改変が可能であり、本出願はあらゆる改変、使用または適用を包含する意図であることが理解されよう。 以下の、全体として本発明の主要部および本開示にはない本発明の属する公知または慣用技術も含めて、上記必須の特徴が適用することが可能であり、添付の請求の範囲に含まれる。
【図面の簡単な説明】
【0062】
【図1】図 1は、大腸菌 (AおよびB)および神経細胞培養物 (CおよびD) における発現に用いたFLIP-Eナノセンサーコンストラクトである。
【図2】図 2は、3種類のグルタミン酸濃度でのFLIP-E 600n センサー (グルタミン酸に対するKdが600 nMである蛍光グルタミン酸ナノセンサー)のスペクトルである: 0 mM (黒)、Kd 値の濃度(青)、および飽和濃度(赤)。これら曲線は520nmの等吸収点を共有する。
【図3】図 3は、 1 mg/ml トリプシンで処理された海馬細胞である。画像(A-D)は10 秒間隔で取得した。細胞表面上のシグナルが大きく消滅していることに注目されたい。
【図4】図 4は、FLIP-E 600n センサーを発現する海馬細胞における発光強度比変化である。画像は発光強度比変化を示すために疑似カラー表示している。グラフ (A)の上の白いバーは処理(刺激 /グルタミン酸の灌流)の時点を示す。矢印で示す時点での比画像(ratio image)をパネル(B)、a〜iに示す。発光強度比の変化が、電気的刺激とグルタミン酸の灌流との両方において観察された。比変化は低レベルの基質 (10 nM グルタミン酸)での灌流では観察されなかった。
【図5】図 5は、FLIP-E 10μ センサー(グルタミン酸に対するKdが10 μMである蛍光グルタミン酸ナノセンサー)を発現する海馬細胞における発光強度比変化である。グラフ (A)の上の白いバーは、処理 (刺激 / グルタミン酸の灌流)の時点を示す。矢印で示す時点での比画像をパネル(B)、a〜gに示す。電気的刺激によっては発光強度比に大きい変化は起こらなかったが、100 μM グルタミン酸での灌流によると可逆的な比変化が引き起こされた (パネル (B)、c およびe)。
【技術分野】
【0001】
関連出願とのクロスリファレンス
本出願は、2004年10月14日出願の米国仮特許出願第60/618,179号および2005年1月14日出願の米国仮特許出願第60/643,576号からの優先権を享受し、これらはその内容全体を引用により本明細書に含める。
【0002】
本出願はまた、仮特許出願60/658,141号、仮特許出願第60/658,142号、仮特許出願第60/657,702号、PCT出願 [代理人整理番号056100-5053、“Phosphate Biosensors and Methods of Using the Same”]、PCT出願 [代理人整理番号056100-5054、“Methods of Reducing Repeat-Induced Silencing of Transgene Expression and Improved Fluorescent Biosensors”]、およびPCT出願 [代理人整理番号056100-5055、“Sucrose Biosensors and Methods of Using the Same”]にも関連し、これらはその内容全体を引用により本明細書に含める。
【0003】
連邦政府助成研究または開発に関する記述
本発明は2つの助成金から援助を受けており、それは、Duke University からのNIH 下請 (下請番号 SPSID 126632)および人間生体メカニズム解明計画助成金(契約番号 RGP0041/2004C)を含む。したがって、米国政府は本発明に対する一定の権利を有する。
【0004】
発明の分野
本発明は一般的に神経伝達物質シグナル伝達の分野に関し、より具体的には、蛍光共鳴エネルギー移動 (FRET)を用いて神経伝達物質レベルの変化を測定および検出するためのバイオセンサーおよび方法に関する。
【背景技術】
【0005】
発明の背景
本明細書中のすべての刊行物および特許出願は、個々の刊行物または特許出願が具体的かつ個別的に引用により明示されている程度に引用により本明細書にその内容を含める。
【0006】
本明細書に記載する刊行物は本出願の出願日より前のその開示を単に提供するものである。いかなる開示も本発明は以前の発明によりかかる刊行物から予測されるということを認めると解釈してはならない。
【0007】
グルタミン酸はアミノ酸であり、脳にみられる神経伝達物質の1つのタイプである。グルタミン酸作動性神経細胞は哺乳類脳において支配的な興奮性経路であり、中枢神経系におけるすべての高速興奮性シナプスの3分の1を占める(Cotman、C.W.、and Monaghan、D.T. (1986) Anatomical organization of excitatory amino acid receptors and their properties. Adv. Exp. Med. Biol. 203、237-252)。グルタミン酸によるシグナル伝達は多数かつ多様な受容体に媒介されており、以下のような受容体が挙げられる:活性化の際、共役したイオンチャンネルを通る細胞外カルシウムの通過を可能とするイオンチャンネル型受容体、および中間体分子、例えば、G タンパク質を活性化して細胞質カルシウム濃を上昇させる分子、例えば、IP3 の生産をもたらす代謝調節型受容体。神経細胞間の相互作用は興奮性または抑制性のいずれかであり得る。主な興奮性アミノ酸神経伝達物質は、グルタミン酸およびアスパラギン酸であり、一方、GABA (γ-アミノ酪酸)、グリシン (アミノ酢酸)、およびタウリンは抑制性である(Mark et al. (2001) American Journal of Neuroradiology 22:1813-1824)。
【0008】
グルタミン酸トランスポーターによる細胞外グルタミン酸のクリアランスは、グルタミン酸蓄積を妨げるために不可欠な工程であり、その工程がなければグルタミン酸蓄積により、グルタミン酸受容体の過剰刺激およびグルタミン酸興奮毒性が起こってしまう。興奮毒性損傷は、多数の神経系疾患を引き起こすかまたはかかる疾患に関与しており、神経性疾患としては、脳卒中、外傷、癲癇、および神経変性症状、例えば、ハンチントン病、エイズによる認知症、および筋萎縮性側索硬化症が挙げられる(Doble、A.、Louvel、E.、and Hugon、J. (1999) The role of excitotoxicity in neurodegenerative disease: implications for therapy、Pharmacol. Ther. 81(3):163-221; Waggie KS、Kahle PJ、Tolwani RJ. (1999) Neurons and mechanisms of neuronal death in neurodegenerative diseases: a brief review. Lab. Anim. Sci. 49:358-362)。グルタミン酸受容体過剰刺激は、イオンチャンネルを直接開けることにより細胞内カルシウムを上昇させ、カルシウムの流入を可能にし、膜脱分極を引き起こす。脱分極は次いで電位依存性カルシウムチャンネルを活性化し、それによりさらに細胞内カルシウムレベルが上昇する。グルタミン酸に誘導されたカルシウムレベルの上昇は以下を含む多数の酵素の過剰活性化を引き起こし、これらのなかには、毒性の遊離酸素ラジカルを産生するものや、神経細胞死をもたらす正のフィードバックループを引き起こすものもある:タンパク質キナーゼ C、カルシウム/カルモジュリン依存性タンパク質 キナーゼ II、ホスホリパーゼ、プロテアーゼ、ホスファターゼ、一酸化窒素シンターゼ、エンドヌクレアーゼおよびオルニチンデカルボキシラーゼ(Mark et al.、2001)。
【0009】
興奮毒性カスケードをトリガーする鍵となる因子はシナプス空間(synaptic space)におけるグルタミン酸の過剰な蓄積である。正常な細胞外グルタミン酸濃度は約 0.6μmol/Lであり、実質的な神経細胞の興奮毒性傷害は2〜5 μmol/Lのグルタミン酸濃度にて起こる。神経細胞に対する外傷性障害は、約 10 μmol/Lの細胞外空間への放出を伴う悲惨な結果をもたらしうる。後に続くカスケードを考えると、1つの神経細胞に対する傷害は、すべての近接する神経細胞を危険にさらすことになる(Mark et al.、2001)。
【0010】
多数の研究が、神経系疾患においてより高いグルタミン酸濃度が関与していることを示しているにもかかわらず、生細胞中のグルタミン酸濃度の測定はいまだに困難である。神経細胞のインビボでの機能を決定するのに必要なもっとも重要な手段の一つは、グルタミン酸の流動を直接に可視化することであろう。グルタミン酸の細胞外濃度は、インビボ微小透析技術によって測定されてきた(Faden、A.I.、Demediuk、P.、Panter、S.S.、and Vink、R. (1989) The role of excitatory amino acids and NMDA receptors in traumatic brain injury. Science 244、798-800; Fallgren、A.B.、and Paulsen、R.E. (1996) A microdyalysis study in rat brain of dihydrokainate、a glutamate uptake inhibitor. Neurochem Res 21、19-25)。しかし、微小透析は空間的および時間的分解能において制限されており、単一のシナプスの周囲の局在化した迅速な濃度変化を検出することはできない。さらに、インビボ微小透析技術は破壊的である。それはまた、生きている神経細胞または星状細胞の内側のグルタミン酸レベルの直接のモニタリングを可能にするものではない。
【0011】
蛍光共鳴エネルギー移動(FRET)の使用によるイオンおよび代謝産物のインビボでの測定を用いたカルシウム濃度変化の測定が成功しており、これはCFP、YFP、およびカルモジュリンおよびM13 ペプチドからなるレポータードメインを融合させることにより行われた(Zhang、J.、Campbell、R.E.、Ting、A.Y.、and Tsien、R.Y. (2002a) Creating new fluorescent probes for cell biology. Nat Rev Mol Cell Biol 3、906-918; Zhang、J.、Campbell、R.E.、Ting、A.Y.、and Tsien、R.Y. (2002b) Creating new fluprescent probes for cell biology. Nature Reviews Molecular Cell Biology 3、906-918)。カルシウムのカルモジュリンへの結合によりキメラの全体的な構造再編が起こり、その結果、ドナーおよびアクセプター蛍光部分によって媒介されてFRET 強度における変化が生じる。最近、基質が結合すると、2つの葉(lobe)がハエジゴクの様に閉じる多数の細菌周辺質結合タンパク質の、代謝産物 ナノセンサーの足場としての使用が成功している(Fehr、M.、Frommer、W.B.、and Lalonde、S. (2002) Visualization of maltose uptake in living yeast cells by fluprescent nanosensor. Proc. Natl. Acad. Sci. U S A 99、9846-9851; Fehr、M.、Lalonde、S.、Lager、I.、Wolff、M.W.、and Frommer、W.B. (2003) In vivo imaging of the dynamics of glucose uptake in the cytosol of COS-7 cells by fluorescent nanosensors. J. Biol. Chem. 278、19127-19133; Lager、I.、Fehr、M.、Frommer、W.B.、and Lalonde、S. (2003) Development of a fluorescent nanosensor for ribose. FEBS Lett 553、85-89)。
【0012】
グルタミン酸のためのナノセンサータンパク質を開発するために、本発明者らは、グルタミン酸が結合すると高次構造が変化するタンパク質を探索した。イオンチャンネル型グルタミン酸受容体(iGluRs)および代謝調節型グルタミン酸受容体 (mGluRs)のファミリーは、γ-アミノ酪酸 (GABA)B 受容体のリガンド結合ドメイン(Kaupmann、K.、Huggel、K.、Heid、J.、Flor、P.J.、Bischoff、S.、Mickel、S.J.、McMaster、G.、Angst、C.、Bittiger、H.、Froestl、W.、and Bettler、B. (1997) Expression cloning of GABA(B) receptors uncovers simirality to metabotropic glutamate receptors. Nature 386、239-246)および、細菌周辺質結合タンパク質に対して配列類似性を有する細胞外リガンド結合ドメインを有している (O'Hara、P.J.、Sheppard、P.O.、Thogersen、H.、Venezia、D.、Haldeman、B.A.、McGrane、V.、Houamed、K.M.、Thomsen、C.、Gilbert、T.L.、and Mulvihill、E.R. (1993) The ligand-binding domain in metabotropic glutamete receptors is related to bacterial periplasmic binding proteins. Neuron 11、41-52)。3種類の形態、1つはグルタミン酸との複合体、2つはリガンド非結合形態、にあるmGluR1 リガンド結合ドメインの結晶構造が決定され(Kunishima、N.、Shimada、Y.、Tsuji、Y.、Sato、T.、Yamamoto、M.、Kumasaka、T.、Nakanishi、S.、Jingami、H.、and Morikawa、K. (2000). Structural basis of glutamate recognition by a dimeric metaborotopic glutamate receptor. Nature 407、971-977)、その結果、グルタミン酸結合が閉じた高次構造を安定化することが示唆された。 Galvez et al. により、GABAB 受容体のリガンド結合ドメインでも2つの葉が閉じることが示唆された(Galvez、T.、Parmentier、M.L.、Joly、C.、Malitschek、B.、Kaupmann、K.、Kuhn、R.、Bittiger、H.、Froestl、W.、Bettler、B.、and Pin、J.P. (1999). Mutagenesis and modeling of the GABAB receptor extracellular domain support a venus flytrap mechanism for ligand binding. J Biol Chem 274、13362-13369)。本発明者らはそれゆえ、mGluRおよびGABAB 受容体を用いてFRET バイオセンサーの構築を試み、基質の添加によるFRET 効率の変化をアッセイした。しかし、FRET 効率の変化は観察されなかった。同様に、細菌由来のLIV ロイシン/イソロイシン/バリンアミノ酸結合タンパク質を操作しても機能的なFRET センサーは得られなかった。
【0013】
De Lorimier et alは、グルタミン-およびヒスチジン-結合タンパク質に対する配列相同性を有し、グルタミン酸代謝に関与するオペロンに位置する大腸菌由来のYbeJ タンパク質が、グルタミン酸およびアスパラギン酸に結合することを示した(de Lorimier、R.M.、Smith、J.J.、Dwyer、M.A.、Looger、L.L.、Sali、K.M.、Paavola、C.D.、Rizk、S.S.、Sadigov、S.、Conrad、D.W.、Loew、L.、and Hellinga、H.W. (2002) Construction of a fluorescent biosensor family Protein Sci 11、2655-2675)。細菌由来のグルタミンおよびヒスチジン結合タンパク質にYbeJが類似していることから、本発明者らは、これら2つのタンパク質の解明された結晶構造に基づく相同性モデルを作成した。グルタミンおよびヒスチジン結合タンパク質の3次元構造により、これらタンパク質のN-およびC- 末端が同じ葉に位置していることが示され、それゆえ基質の結合により2つの葉が閉じることは、N-およびC-末端の間の距離を変化させるわけではないようである。したがって、これらのタンパク質はいずれも同じ原理に基づくFRET センサーの構築を可能にするものではないはずである。実際、米国特許出願公開第20040118681号においてHellinga and Looger により提案されたセンサーは、単一のフルオロフォアを、リガンド結合の際の高次構造変化に応答するシステイン残基に結合させることを提案しており、これは、FRET に用いられる蛍光部分が2つであることとは全く異なる。
【発明の開示】
【発明が解決しようとする課題】
【0014】
しかし、本発明者らは驚くべきことに、大腸菌のYbeJ タンパク質が、両方の末端がタンパク質の同じ葉に位置しているという知見にもかかわらず、グルタミン酸結合の検出のための有効なFRET 足場であることを見いだした。これは、距離変化がFRET変化に変換されるという一般的な仮説に反するものである。
【課題を解決するための手段】
【0015】
発明の概要
本発明は、神経伝達物質濃度における変化を検出および測定するための神経伝達物質バイオセンサーを提供する。具体的には、本発明は、大腸菌 YbeJ由来のグルタミン酸結合タンパク質部分を含むグルタミン酸結合蛍光指標 (FLIP-E) をコードする単離核酸を提供し、ここで、該グルタミン酸結合タンパク質部分は遺伝子的にドナー蛍光タンパク質部分およびアクセプター蛍光タンパク質部分と融合しており、ここで、ドナー部分とアクセプター部分との間の蛍光共鳴エネルギー移動 (FRET)は、ドナー部分が励起され、グルタミン酸がグルタミン酸結合タンパク質部分に結合した場合に変化する。本発明の核酸を含むベクター、例えば、発現ベクターおよび宿主細胞も提供され、また、該核酸によりコードされるバイオセンサータンパク質も提供される。かかる核酸、ベクター、宿主細胞およびタンパク質は、神経伝達物質レベル、特に神経細胞サンプル中の細胞外グルタミン酸レベルの変化の検出方法、およびグルタミン酸興奮毒性を調節する化合物の同定方法に使用することができる。
【0016】
図面の簡単な説明
図 1.大腸菌 (AおよびB)および神経細胞培養物 (CおよびD) における発現に用いたFLIP-Eナノセンサーコンストラクト。
【0017】
図 2.3種類のグルタミン酸濃度でのFLIP-E 600n センサー (グルタミン酸に対するKdが600 nMである蛍光グルタミン酸ナノセンサー)のスペクトル: 0 mM (黒)、Kd 値の濃度(青)、および飽和濃度(赤)。これら曲線は520nmの等吸収点を共有する。
【0018】
図 3. 1 mg/ml トリプシンで処理された海馬細胞。画像(A-D)は10 秒間隔で取得した。細胞表面上のシグナルが大きく消滅していることに注目されたい。
【0019】
図 4. FLIP-E 600n センサーを発現する海馬細胞における発光強度比変化。画像は発光強度比変化を示すために疑似カラー表示している。グラフ (A)の上の白いバーは処理(刺激 /グルタミン酸の灌流)の時点を示す。矢印で示す時点での比画像(ratio image)をパネル(B)、a〜iに示す。発光強度比の変化が、電気的刺激とグルタミン酸の灌流との両方において観察された。比変化は低レベルの基質 (10 nM グルタミン酸)での灌流では観察されなかった。
【0020】
図 5. FLIP-E 10μ センサー(グルタミン酸に対するKdが10 μMである蛍光グルタミン酸ナノセンサー)を発現する海馬細胞における発光強度比変化。グラフ (A)の上の白いバーは、処理 (刺激 / グルタミン酸の灌流)の時点を示す。矢印で示す時点での比画像をパネル(B)、a〜gに示す。電気的刺激によっては発光強度比に大きい変化は起こらなかったが、100 μM グルタミン酸での灌流によると可逆的な比変化が引き起こされた (パネル (B)、c およびe)。
【0021】
発明の詳細な説明
以下の記載は本発明の理解に有用であり得る情報を含む。本明細書により提供される情報はいずれも従来技術であるとも本願請求項の発明と関連するものとも認めるものではなく、明示的または黙示的に引用するいずれの刊行物も従来技術であると認めるものではない。
【0022】
本発明のその他の目的、利点および特徴が本出願において提供する明細書および図面をみることにより当業者に明らかとなる。したがって、本発明のさらなる目的および利点は以下の記載から明らかであろう。
【0023】
バイオセンサー
本発明は、蛍光共鳴エネルギー移動 (FRET)を用いて神経伝達物質濃度の変化を検出および測定するための神経伝達物質バイオセンサーを提供する。神経伝達物質として作用する物質の3つの主なカテゴリーは以下の通りである:(1) アミノ酸 (主にグルタミン酸(glutamic acid または glutamate)、GABA、アスパラギン酸およびグリシン)、(2) ペプチド(バソプレッシン、ソマトスタチン、ニューロテンシン等) および (3) モノアミン (ノルエピネフリン、ドーパミンおよびセロトニン)ならびにアセチルコリン。具体的には、本発明は、グルタミン酸結合蛍光指標、特に、大腸菌グルタミン酸/アスパラギン酸受容体であるYbeJ由来のグルタミン酸結合タンパク質部分を含む指標を提供する。上記の神経伝達物質についてのさらなる神経伝達物質バイオセンサーも本明細書において提供するコンストラクトおよび方法を用いて調製することが出来る。
【0024】
したがって、本発明は、神経伝達物質に結合する蛍光指標をコードする単離核酸を提供する。とりわけ、一つの態様は、以下を含むグルタミン酸結合蛍光指標をコードする単離核酸である: グルタミン酸結合タンパク質部分、遺伝子的にグルタミン酸結合タンパク質部分と融合したドナー蛍光タンパク質部分、および遺伝子的にグルタミン酸結合タンパク質部分と融合したアクセプター蛍光タンパク質部分、ここでドナー部分とアクセプター部分との間のFRETは、ドナー部分が励起され、グルタミン酸がグルタミン酸結合タンパク質部分に結合すると変化する。好ましいグルタミン酸結合タンパク質部分は大腸菌 YbeJ由来のグルタミン酸結合タンパク質部分である。
【0025】
YbeJは当該技術分野においてYzzKおよびGltIとしても知られており、その DNA 配列 (配列番号1)およびタンパク質配列 (YbeJ、タンパク質受入番号 NP_415188、配列番号2) は既知である。グルタミン酸結合領域をコードするYbeJ DNA 配列のいずれの部分を本発明の核酸において用いてもよい。例えば、本発明の核酸における使用に好適な一つの領域は配列番号3により提供され、この配列は切型(truncated)グルタミン酸-アスパラギン酸結合タンパク質配列 (配列番号4)をコードし、シグナルペプチドを含まない成熟タンパク質をコードするものである。他の細菌種からの天然のホモログも用いることが出来、例えば緑膿菌(Pseudomonas aeruginosa )由来のPA5082遺伝子が挙げられ、その遺伝子産物は大腸菌のYbeJ タンパク質と70% 類似する。YbeJまたはそのいずれかのホモログのグルタミン酸結合部分を本明細書に記載するベクターにクローニングし、開示するアッセイにしたがって活性についてスクリーニングすればよい。
【0026】
測定可能なグルタミン酸結合機能を保持する、部位特異的突然変異、欠失または挿入を含む人工的に遺伝子操作された変異体に加えて、YbeJの天然の種変異体も用いることが出来る。本発明の核酸コンストラクトにおける使用に好適な変異体核酸配列は、好ましくは YbeJの遺伝子配列に対して少なくとも 70、75、80、85、90、95、または99%の類似性または同一性を有するであろう。好適な変異体核酸配列はまた、高度にストリンジェントなハイブリダイゼーション条件下でYbeJ遺伝子にハイブリダイズしうる。高度にストリンジェントな条件は当該技術分野において知られている;例えば、 Maniatis et al.、Molecular cloning: A Laboratory Manual、2d Edition、1989、および Short Protocols in Molecular Biology、ed. Ausubel、et alを参照されたい:これらはいずれも引用により本明細書に含める。ストリンジェントな条件は配列に依存し、異なる環境では異なるものとなろう。より長い配列はより高い温度で特異的にハイブリダイズする。核酸のハイブリダイゼーションについての詳細な説明は、Tijssen、Techniques in Biochemistry and Molecular Biology--Hybridyzation with Nucleic Acid Probes、"Overview of principles of Hybridyzation and the strategy of nucleic acid assays" (1993)にみられる。一般に、ストリンジェントな条件は規定されたイオン強度およびpHにおける特定の配列についての熱融解温度 (Tm)よりも約 5-10℃低くなるよう選択する。 (規定されたイオン強度、pHおよび核酸濃度における) Tmは、標的に相補的なプローブの50%が平衡状態で標的配列にハイブリダイズする温度である(Tm において標的配列が過剰に存在する場合、プローブの50%が平衡状態で占有される)。ストリンジェントな条件は、塩濃度が約 1.0M未満のナトリウムイオン、典型的には約 0.01〜1.0Mのナトリウムイオン濃度 (またはその他の塩)、pH 7.0 〜 8.3、そして短いプローブ(例えば、10〜 50 ヌクレオチオド)については少なくとも 約 30℃の温度、長いプローブ(例えば、50を超えるヌクレオチオド)については少なくとも 約 60℃の温度であろう。ストリンジェントな条件はまた、脱安定化剤、例えば、ホルムアミドを添加して達成してもよい。
【0027】
本発明の好ましい人工変異体は、YbeJに基づく、およびその他のグルタミン酸ナノセンサーにより測定できる濃度範囲を広げるために、グルタミン酸に対する親和性が上昇または低下しているものであり得る。とりわけ好ましい人工変異体としては、以下の突然変異を含むグルタミン酸結合領域が挙げられる:A207G、A207P、A207K A207M、A207S、A207C、A207R、A207V、A207L、A207Q、A207T、A207F、A207Y、A207N、A207W、A207H、A207D、および/またはS95W。グルタミン酸に対する結合親和性が低下または上昇しているさらなる人工変異体は、ランダムまたは部位特異的突然変異誘発およびその他の突然変異誘発技術により構築でき、本明細書に記載するベクターにクローニングし、開示するアッセイにしたがって活性についてスクリーニングすればよい。
【0028】
本発明の単離核酸は、組み合わせた場合にFRETにおけるドナーおよびアクセプター部分として作用することができるいずれの好適なドナーおよびアクセプター蛍光タンパク質 部分を組み込んでいてもよい。好ましいドナーおよびアクセプター部分は、GFP (緑色蛍光 タンパク質)、CFP (シアン蛍光タンパク質)、BFP (青色蛍光タンパク質)、YFP (黄色蛍光タンパク質)、およびそれらの増強された変異体からなる群から選択され、特に好ましい態様はドナー/アクセプター対CFP/YFP-Venusにより提供され、YFP-VenusはpH 耐性および熟成時間が改良されたYFPの変異体である (Nagai、T.、Ibata、K.、Park、E.S.、Kubota、M.、Mikoshiba、K.、and Miyawaki、A. (2002) A variant of yellow fluprescent protein with fast and efficient maturation for cell-biological applications. Nat. Biotechnol. 20、87-90)。あるいは好ましくはより高いpH 安定性およびより大きいスペクトル分離を示すMiCy/mKO 対である(Karasawa S、Araki T、Nagai T、Mizuno H、Miyawaki A. Cyan-emitting and/or ange-emitting fluprescent proteins as a donor/acceptor pair for fluorescence resonance energy transfer. Biochem J. 2004 381:307-12)。ドナーおよびアクセプター蛍光部分を選択する際に考慮すべき基準は当該技術分野において知られており、例えばUS 6,197,928に開示されており、その全体を引用により本明細書に含める。
【0029】
ドナーまたはアクセプターのいずれかとして好適なものとしてはまた、 Discosoma 種からのネイティブな DsRed、別の属からのDsRedのオルソログまたはネイティブな DsRedの性質を最適化した変異体(例えば、K83M 変異体またはDsRed2 (Clontechから入手可能))が挙げられる。 本明細書において用いる場合、「変異体」という用語は、ネイティブな蛍光分子に対して少なくとも 約 70%、より好ましくは少なくとも 75%の同一性、例えば、少なくとも 80%、90%、95%またはそれ以上の同一性を示すポリペプチドをいう。多くのかかる変異体が当該技術分野において知られているし、あるいはネイティブな蛍光分子のランダムまたは指向性突然変異誘発により容易に調製することができる(例えば、Fradkov et al.、FEBS Lett. 479:127-130 (2000)を参照)。
【0030】
バイオセンサーのフルオロフォアが(1または複数の)類似または関連配列のストレッチを含む場合、本発明者らは最近、遺伝子サイレンシングが特定の細胞および特に生物全体におけるバイオセンサーの発現に悪影響を与えうることを見いだした。かかる状況においては、フルオロフォアをコードする配列を1以上の各フルオロフォアのコドンの縮退またはゆらぎ位置にて改変し、フルオロフォアの核酸配列が改変されるがコードされるアミノ酸配列は変化しないようにすることが可能である。あるいは、フルオロフォアの機能に悪影響を与えない1以上の保存的置換を導入してもよい。PCT出願 [代理人整理番号056100-5054、“Methods of Reducing Repeat-Induced Silencing of Transgene Expression and Improved Fluorescent Biosensors”]を参照されたい。これはその全体を引用により本明細書に含める。
【0031】
本発明はさらに、神経伝達物質バイオセンサーポリペプチドをコードする単離核酸分子を含むベクターを提供する。例示的なベクターとしては、以下が挙げられる:ウイルス、例えば、バクテリオファージ、バキュロウイルスまたはレトロウイルス由来のベクター、および細菌または細菌配列とその他の生物由来の配列との組合せ由来のベクター、例えば、コスミドまたはプラスミド。かかるベクターとしては、神経伝達物質バイオセンサーをコードする核酸配列に作動可能に連結した発現制御配列を含む発現ベクターが挙げられる。ベクターは原核細胞、例えば、大腸菌またはその他の細菌において機能するのに適したものでもよいし、真核細胞、例えば酵母および動物細胞において機能するのに適したものでもよい。例えば、本発明のベクターは一般に以下のような要素を含むであろう:例えば、所望の宿主細胞に適合する複製起点、所望の宿主細胞に適合する1以上の選択可能なマーカーおよび1以上のマルチクローニングサイト。ベクターに含まれる特定の要素の選択は以下のような因子に依存するであろう:例えば、目的の宿主細胞、インサートのサイズ、挿入された配列の制御された発現が望まれるか否か、即ち、例えば、誘導可能または調節可能なプロモーターの使用を介するべきか否か、目的のベクターのコピー数、目的の選抜系等。異なる用途のための宿主細胞とベクターとの適合性を確証するのに関与する因子は当該技術分野において周知である。
【0032】
本発明において使用するために好ましいベクターは、ドナーおよびアクセプター蛍光分子をコードする核酸の間への神経伝達物質結合ドメインまたは受容体のクローニングを可能にし、その結果、ドナーおよびアクセプター蛍光分子に遺伝子的に融合した神経伝達物質結合ドメインを含むキメラまたは融合タンパク質の発現をもたらすものであろう。例示的なベクターとしては、細菌 pRSET-FLIP誘導体が挙げられ、これはFehr et al. (2002) (Visualization of maltose uptake in living yeast cells by fluprescent nanosensor. Proc. Natl. Acad. Sci. U S A 99、9846-9851)に開示されており、これはその全体を引用により本明細書に含める。あるいは、目的の神経伝達物質結合ドメインをまず蛍光ドナーおよびアクセプターをコードする配列に融合させ、その後で適当なベクターにクローニングしてもよく、これは米国特許第6,596,499号に記載されており、その全体を引用により本明細書に含める。
【0033】
本発明のキメラ核酸は好ましくは、ドナーおよびアクセプター間のFRETの変化が神経伝達物質の結合の際に検出されうるように、ドナーおよびアクセプター蛍光部分をコードする配列が、神経伝達物質結合ドメインの別々の末端に融合するように構築する。あるいは、該ドナーフルオロフォアおよび/または該アクセプターフルオロフォア部分の一方または両方をリガンド結合タンパク質部分に、該リガンド結合タンパク質部分の内部にて融合させてもよい。かかる融合は仮出願第60/658,141号に記載されており、これは引用により本出願に含める。ドナーおよびアクセプター部分はタンパク質の同じドメインまたは葉に含めてもよいが、好ましくは、ドナーおよびアクセプター部分はタンデムには融合されるべきではない。ドメインとは、特定の機能を担うタンパク質の部分であり、典型的には少なくとも 約 40〜約 50 アミノ酸の長さである。一つのタンパク質に複数のタンパク質ドメインが含まれうる。
【0034】
蛍光ドメインは所望の場合は1以上の可動性リンカー配列によって、神経伝達物質結合ドメインから離れていてもよい。かかるリンカー部分は、好ましくは約 1〜50 アミノ酸残基の長さであり、より好ましくは約 1〜30 アミノ酸残基の長さである。リンカー部分およびその用途は当該技術分野において周知であり、例えば、米国特許第5,998,204号および5,981,200号、およびNewton et al.、Biochemistry 35:545-553 (1996)に記載されている。あるいは、フルオロフォアまたは結合タンパク質を短くした形態のものを用いてもよい。
【0035】
例えば、本発明者らは、結合ドメインのコアタンパク質構造とフルオロフォアとを連結する配列を除くことにより、即ち、リンカー配列を除くことによるか、および/または、分析対象結合部分および/またはフルオロフォアの末端からアミノ酸を欠失させることにより、フルオロフォアのより近い結合 (coupling)が達成され、その結果比変化が大きくなることも見いだした。好ましくは、欠失は、リンカーまたはフルオロフォアまたはリガンド結合ドメインをコードする領域に位置するFRET バイオセンサーをコードする核酸コンストラクトにおいて少なくとも1、または少なくとも2、または少なくとも3、または少なくとも4、または少なくとも5、または少なくとも8、または少なくとも10、または少なくとも15ヌクレオチオドを欠失させることにより行う。異なる領域における欠失を一つのコンストラクトにおいて組み合わせることにより硬直性(rigidity)が増強した2以上の領域を作ることができる。アミノ酸をまた、付加または突然変異させて、例えば、プロリン残基またはその他の好適なアミノ酸を付加することによって捻れを導入することによって、バイオセンサーの硬直性を増強させ、感度を向上させることも出来る。感度の上昇はリガンド結合の際のFRET 蛍光の比変化によって測定でき、好ましくは感度は、該(1または複数の)欠失の結果少なくとも2倍上昇する。仮出願第60/658,141号を参照されたい。これはその全体を引用により本明細書に含める。
【0036】
本発明はまた、本発明のベクターまたは発現ベクターによってトランスフェクトされた宿主細胞も含み、例えば、原核細胞、例えば、大腸菌またはその他の細菌、あるいは真核細胞、例えば、酵母細胞または動物細胞が挙げられる。別の側面において、本発明は、神経伝達物質バイオセンサーの発現をコードする核酸配列の発現によって特徴づけられる表現型を有するトランスジェニック非ヒト動物を特徴とする。表現型は以下によって作成しうる動物の体細胞および生殖細胞に含まれる導入遺伝子によって与えられる:(a)動物の接合体(zygote)に、神経伝達物質バイオセンサーをコードするDNA コンストラクトを含む導入遺伝子を導入すること; (b)接合体を偽妊娠動物に移植すること; (c)接合体を出産(term)まで発生させること;および(d)導入遺伝子を含む少なくとも1つのトランスジェニック子孫を同定すること。導入遺伝子を胚に導入する工程は、導入遺伝子を含む胚性幹細胞を胚に導入すること、または、導入遺伝子を含むレトロウイルスを胚に感染させることによって行うことが出来る。好ましいトランスジェニック動物は脳において、コードされた神経伝達物質バイオセンサーを発現するであろう。本発明のトランスジェニック動物には、トランスジェニック線虫およびトランスジェニックマウスおよびその他の動物が含まれる。
【0037】
本発明はまた、本明細書に記載する性質を有する単離神経伝達物質バイオセンサー分子、特に、YbeJに基づくグルタミン酸結合蛍光指標も含む。かかるポリペプチドは、本明細書に記載する核酸コンストラクトを用いて組換えにより発現させてもよいし、成分となるドメインのいくつかまたはすべてを化学的に結合させることにより製造してもよい。発現したポリペプチドは、転写翻訳系または組換え細胞において産生させてもよいし、および/または転写翻訳系または組換え細胞から、当該技術分野において知られている生化学的および/または免疫学的精製方法によって単離してもよい。本発明のポリペプチドは、脂質二重層、例えば、細胞膜抽出物、または人工脂質二重層 (例えば、リポソーム小胞) またはナノ粒子に導入してもよい。
【0038】
神経伝達物質レベルの検出方法
本発明の核酸およびタンパク質は、動物の脳または神経系における神経伝達物質のレベルの変化、特にグルタミン酸興奮毒性に関連する障害または疾患のシグナルであり得る細胞外グルタミン酸のレベルの変化の検出および測定に有用である。一つの態様において、本発明は、以下を含む神経細胞のサンプルにおける細胞外グルタミン酸のレベルの変化を検出する方法を含む:(a)本明細書に記載するグルタミン酸結合バイオセンサーをコードする核酸を発現する細胞、および、神経細胞のサンプルを提供すること;および(b)それぞれグルタミン酸結合ドメインに共有結合している、ドナー蛍光タンパク質部分とアクセプター蛍光タンパク質部分との間のFRETの変化を検出すること、ここで、該ドナー部分と該アクセプター部分との間のFRETの変化は、神経細胞のサンプル中の細胞外グルタミン酸レベルの変化を示す。あるいは、タンパク質は異種宿主、例えば細菌で産生させ、精製し、器官に直接、または細胞間空間に注入してもよい。タンパク質またはその誘導体は、量子ドットなどの粒子に結合させて、細胞または区画に導入してもよい。
【0039】
FRETは当該技術分野において知られている様々な技術を用いて測定することが出来る。例えば、FRETの判定工程は、アクセプター蛍光タンパク質部分から放射される光の測定を含みうる。あるいは、FRETの判定工程は、ドナー蛍光タンパク質部分から放射される光を測定すること、アクセプター蛍光タンパク質部分から放射される光を測定すること、および、ドナー蛍光タンパク質部分から放射される光とアクセプター蛍光タンパク質部分から放射される光の比を計算することを含みうる。FRETの判定工程はまた、ドナー部分の励起状態寿命または異方性変化を測定することを含みうる (Squire A、Verveer PJ、Rocks O、Bastiaens PI. J Struct Biol. 2004 Jul;147(1):62-9 Red-edge anisotropy microscopy enables dynamic imaging of homo-FRET between green fluorescent protein in cells)。かかる方法は当該技術分野において知られており、US 6,197,928に一般的に記載されており、これはその全体を引用により本明細書に含める。
【0040】
神経細胞のサンプル中のグルタミン酸またはその他の神経伝達物質の量はFRETの程度を測定することにより決定することが出来る。第一に、FLIP-E センサーをサンプルに導入しなければならない。神経伝達物質濃度の変化は、所定の時間間隔でFRET 変化をモニタリングすることにより判定することが出来る。サンプル中の神経伝達物質の量は、例えばインビボでの滴定 (titration)により確立した検量線を用いて定量することが出来る。
【0041】
本発明の方法により分析すべき神経細胞サンプルは、インビボ、例えば、海馬神経細胞の表面上でのグルタミン酸流出の測定において含めてもよいし、または、グルタミン酸流出が神経細胞培養物にて測定される、インビトロに含めてもよい。あるいは、脳または1以上のシナプス空間からの液体抽出物を細胞外神経伝達物質が検出または測定されるサンプルとして用いてもよい。かかる測定は、該神経細胞に対する外傷性障害に伴う細胞外グルタミン酸の検出に用いてもよいし、グルタミン酸興奮毒性に関する神経障害、例えば、特に、脳卒中、癲癇、ハンチントン病、エイズによる認知症および筋萎縮性側索硬化症の可能性の指標として用いてもよい。
【0042】
本明細書に開示する神経伝達物質レベルの検出方法は、神経伝達物質濃度の調節に使用しうる化合物、特に、グルタミン酸興奮毒性の調節に有用な化合物のスクリーニングおよび同定に使用することができる。一つの態様において、とりわけ、本発明は以下を含むグルタミン酸興奮毒性を調節する化合物の同定方法を含む:(a)本明細書に開示するグルタミン酸バイオセンサーを発現する細胞および神経細胞のサンプルを1以上の被験化合物と接触させること、および(b)該ドナー蛍光ドメインと該アクセプター蛍光ドメインの間のFRETを該接触の後に測定すること、ここで該接触後のFRETの上昇または低下は、該被験化合物がグルタミン酸興奮毒性を調節する化合物であることを示す。「調節する」という用語は、かかる化合物がグルタミン酸興奮毒性を上昇または低下させうることを意味する。グルタミン酸レベルを上昇させる化合物は上記のようなグルタミン酸興奮毒性に関連する障害の治療的介入および処置のための標的である。グルタミン酸レベルを低下させる化合物はグルタミン酸興奮毒性に関連する障害の処置のための治療用品へと開発することが出来る。
【0043】
原形質膜の外側の葉(leaflet)に対するセンサーの標的化は、可能性のある用途の一つの態様に過ぎない。それは、ナノセンサーは特定の区画に標的化することが出来ることを示す。あるいは、その他の標的化配列を用いて、センサーをその他の区画、例えば、小胞、ER、液胞、などに発現させることが出来る。
【0044】
発現系にはラット神経細胞のみならず、ヒト細胞株、動物細胞および器官、真菌および植物細胞も含まれる。センサーはまた、グルタミン酸が重要な窒素化合物として役割を果たすのみならず、おそらくはシグナル伝達分子としても作用する、真菌および植物生物体におけるグルタミン酸レベルをモニターするのにも用いることが出来る。細菌における発現を用いて、感染部位または細菌が存在するかまたは導入される区画におけるグルタミン酸レベルをモニターすることができる。
【0045】
具体的には、センサーを発現する細菌または真菌は、バイオセンサーとして働くことができ、あるいは、上記の薬物スクリーニングについて記載したものと同様のスキームを用いて、新規殺虫剤の同定のためのツールとして働くことが出来る。
【0046】
さらなる有用性
本発明のバイオセンサーは、神経細胞の機能を判定するために動物細胞の表面に発現させてもよい。例えば、線虫において存在する神経細胞の多くはいまだに特定の機能がわかっていない。表面上でのバイオセンサーの発現により、刺激に応答した生きた線虫における神経細胞活性の可視化が可能となり、それにより神経細胞回路網の機能の決定および分析が可能となる。同様に、生きたマウスまたはラットの脳に多光子プローブを導入することにより、それらのプロセスを画像化することが可能となる。最後に、特定の神経細胞または膠細胞における発現により、 脳卒中またはアルツハイマー病などの現象の研究、および神経細胞の内側またはその表面におけるグルタミン酸レベルへのかかる障害の効果の研究が可能となる。さらに、局所脳領域または神経細胞回路網への薬物療法の効果をインビボで研究することが出来る。
【0047】
最後に、センサーを、例えば膜または人工脂質複合体上に提示される人工グルタミン酸捕捉剤として導入することにより、グルタミン酸流動を改変するツールとしてセンサーを使用することが可能であり、したがって脳または神経細胞機能を操作することが可能である。
【0048】
以下の実施例は本発明を説明および例示するために提供する。したがって、以下の実施例は本発明の範囲を限定するものと解釈してはならない。当業者であれば多くのその他の態様が本明細書および請求の範囲に記載する本発明の範囲内に包含されることを理解しているであろう。
【実施例】
【0049】
実施例 1. 核酸およびベクターの構築
シグナルペプチドを有さない成熟タンパク質をコードする切型グルタミン酸-アスパラギン酸結合タンパク質配列 (配列番号4)を、大腸菌 ゲノム DNAをテンプレートとして用いてPCRにより増幅した。用いたプライマーは、5’- ggtaccggaggcgccgcaggcagcacgctggacaaaatc -3’(配列番号5) および5’- accggtaccggcgccgttcagtgccttgtcattcggttc - 3’(配列番号6)であった。 PCR 断片をpRSET ベクター (Invitrogen)中のFLIPmal-25μ (Fehr et al. 2002)のKpnI 部位にクローニングし、マルトース結合タンパク質配列をYBEJ 配列に交換した。その結果得られたプラスミドをpRSET-FLIP-E-600nと命名した。
【0050】
pHおよびクロライド耐性を向上させ、センサータンパク質の成熟を向上させるために、pRSET-FLIP-E-600n 中の増強 YFP (EYFP、CLONTECH) 配列を含む断片をVenusの コード配列で置換した。VenusはpH 耐性と熟成時間が改良したYFPの変異体である(Nagai、T.、Ibata、K.、Park、E.S.、Kubota、M.、Mikoshiba、K.、and Miyawaki、A. (2002) A mutant of yellow fluorescent protein with fast and efficient maturation for cell-biological applications. Nat. Biotechnol. 20、87-90)。以下の置換を有する親和性突然変異体:A207G、A207P、A207K、A207M、A207S、A207C、A207R、A207V、A207L、A207Q、A207T、A207F、A207Y、A207N、A207W、A207H、A207D 、またはS95W:を部位特異的突然変異誘発により作成した (Kunkel、T.A.、Roberts、J.D.、and Zakour、R.A. (1987). Rapid and efficient site-specific mutagenesis without phenotypic selection. Methods Enzymol. 154、367-382)。
【0051】
pRSET-FLIP-E コンストラクトをエレクトロポレーションを用いて大腸菌 BL21(DE3)Gold (Stratagene) に移した(Sambrook、J.、Fritsch、E.F.、and Maniatis、T. (1989). Molecular Cloning. A laboratory manual. (Cold Spring Harbor NY: Cold Spring Harbor Laboratory Press)。BL21(DE3)Gold 株で発現したFLIP-E タンパク質を以前に記載されたようにして抽出し、精製した (Fehr et al. 2002)。ラット初代神経細胞培養物およびPC12 細胞培養物における発現のために、FLIP-E 600nおよび-10μ カセットを以下にようにしてpDisplay (Invitrogen)にクローニングした: XmaI 部位 およびSalI 部位をPCRによりFLIP-E カセットの5’- および3’-末端にそれぞれ導入した。用いたプライマーは、5’- gagcccgggatggtgagcaagggcgaggag -3’(配列番号7)および5’- gaggtcgaccttgtacagctcgtccatgccgag -3' (配列番号8)であった。PCR 断片を配列決定してPCRエラーの追加がないことを確認し、XmaI/SalIで消化し、pDisplay ベクターのXmaI/SalI 部位にクローニングした。細胞培養物に改変リン酸カルシウム トランスフェクションプロトコール(Xia、Z.、Dudek、H.、Miranti、C.K.、and Greenberg、M.E. (1996). Calcium influx via the NMDA receptor induces immediate early gene transcription by a MAP kinase/ERK-dependent mechanism. J. Neurosci. 16、5425-5436)またはLipofectamine (Invitrogen)を用いてトランスフェクトした。
【0052】
実施例 2. FLIP-E ナノセンサーのインビトロ特徴決定
成熟 YBEJ タンパク質をコードするDNA 断片をN- およびC-末端にてそれぞれECFPおよびVenus 配列に融合させた(図 1)。発光スペクトルおよび基質滴定曲線をモノクロメータ マイクロプレートリーダー Safire (Tecan、Austria)を用いて得た。 励起フィルターは433±12nmであり、CFP および YFP 発光についての発光フィルターはそれぞれ485±12、528nm±12nmであった。すべての分析は20mM リン酸ナトリウムバッファー、pH 7.0中で行った。
【0053】
グルタミン酸の添加の結果、CFP 発光が上昇し、YFP 発光が低下し、これは、グルタミン酸のYBEJへの結合の結果、フルオロフォアの双極子の配向の相対変化におそらく起因してキメラタンパク質の高次構造変化が起こったことを示唆する(図 2)。CFPおよびYFP 部分は同じ葉に結合していると考えられることから、本発明者らは、グルタミン酸結合は2つのフルオロフォアの双極子-双極子角の変化をもたらすと推測する。興味深いことに、比および比変化はいままでに作成されたその他のセンサーと比べて同様の範囲内であり(Fehr et al.、2002; Fehr et al.、2003; Lager et al.、2003)、これは距離変化はFRET 変化のメカニズムの根底にある第一の因子ではないであろうということを示唆する。3種類のグルタミン酸濃度(0、Kd、飽和)でのスペクトルにより520 nmの等吸収点が明らかである (図 2)。グルタミン酸についての結合定数 (Kd)は600nMであると判明し、その他の方法によって得られたデータと一致した(de Lorimier et al.、2002)。アスパラギン酸、グルタミン、アスパラギンについての結合定数はそれぞれ1μM、100μM、300μMであると判明した(以下の表1参照)。
【0054】
YBEJに基づくグルタミン酸ナノセンサーにより測定可能な濃度範囲を広げるために、YBEJ 部分に突然変異を誘発してグルタミン酸に対する親和性がより小さいナノセンサーを作成した。リガンド結合部位を形成するドメイン間間隙の周辺に位置する部位(「周囲立体(peristeric)」と称される)に様々なフルオロフォアを結合させることにより、周辺質結合タンパク質におけるリガンド結合親和性が変化することが以前に示されている(de Lorimier et al.、2002)。試験した残基のなかで、アラニン 207からリジン、メチオニン、セリン、システイン、アルギニン、バリン、ロイシン、グルタミン、スレオニン、フェニルアラニン、チロシン、アスパラギン、トリプトファン、ヒスチジン、アスパラギン酸への突然変異は結合親和性を有意に低下させた(表 1)。さらに、セリン 118から、グルタミン酸の窒素と相互作用することが示唆されるトリプトファンへの突然変異は、タンパク質の親和性を低下させることが判明した。したがって、FLIPE ナノセンサーに導入した突然変異により生理的グルタミン酸濃度の広い範囲を包含する好適な親和性突然変異体を作ることが出来る。
【0055】
表 1.
【表1】
【0056】
実施例 3. FLIP-Eのインビボ特徴決定
FLIP-E ナノセンサーのインビボ特徴決定のために、FLIPE-600n およびFLIPE-10μを哺乳類発現ベクター pDisplay (Invitrogen、USA)にクローニングした。pDisplay ベクター はタンパク質を分泌経路に向けるリーダー配列、および、タンパク質を原形質膜にアンカーする膜貫通ドメインを含み、タンパク質を細胞外表面に提示する。ラット海馬細胞およびPC12 細胞にpDisplay FLIPE-600nおよび -10μ コンストラクトをトランスフェクトした。FRETをトランスフェクションの24 - 48時間後に冷却 CoolSnap HQ デジタルカメラ (Photometrics)を備えた蛍光顕微鏡 (DM IRE2、Leica)で画像化した。二重(dual)発光強度比を436nmでの励起の後に同時に記録し、OI-5-EM フィルターセット (Optical Insights) およびMetafluor 6.1r1 software (Universal Imaging)を備えたDualViewによりCFPとVenus 発光とを分割した。
【0057】
FLIP-Eの発現はラット海馬細胞培養の原形質膜上に観察され、ある程度はおそらくは原形質膜タンパク質の原形質膜への標的化に関与する区画である細胞内区画にも観察された。1 mg/mL のトリプシンを含むタイロードバッファーで処理した場合、細胞表面の蛍光の大部分は消失し、pDisplay コンストラクトの性質から予測されるように、 FLIPE タンパク質が実際に原形質膜の細胞外表面に提示されていることが示された(図 3)。ナノセンサーはしたがって細胞表面近くの細胞外グルタミン酸レベルを測定しているはずである。
【0058】
CFPおよびVenus 発光の強度を定量するために、細胞周辺の2つのチャンネルにおける蛍光強度をピクセル毎に(pixel-by-pixel basis)積分し (integrated)、CFP/Venus 比を算出した。表面にFLIPE-600nを提示する海馬細胞に電流パルスを通すことにより電気的に刺激すると、CFP/Venus 発光比の低下が観察され(図 4 a-c)、グルタミン酸が膜脱分極によって海馬細胞から放出されていることが示唆された。比変化がグルタミン酸の細胞外濃度の変化に起因することを確認するために、細胞に様々な濃度のグルタミン酸を灌流した。発光強度比は濃度依存的に変化し(図 4 d-h)、細胞表面に提示されたFLIPE-600n が細胞外グルタミン酸を認識していることが示された。FLIP-E 600n センサー の動作範囲は100nM〜1μMであり、これはFLIPE-600n ナノセンサーのインビトロ動作範囲と一致する。CFP/Venus 比は外部培地を灌流により除くと上昇し、FRET 強度のインビボ変化は可逆的であることが示唆された。
【0059】
FLIPE/600n センサーを発現する細胞とは対照的に、CFP/Venus 発光強度変化はFLIPE-10μを発現する細胞において電気刺激によっては観察されなかった(図 5)。しかし比変化は細胞をより高濃度のグルタミン酸で灌流した場合に観察され(図 5 cおよびe)、細胞の脱分極により誘導されるグルタミン酸濃度変化はFLIP-E 10μ センサーの動作範囲を下回ることが示唆された。
【0060】
新規ナノセンサーはしたがって神経細胞表面のグルタミン酸を測定することが出来、前シナプス神経細胞のグルタミン酸分泌を直接追跡することが出来る。
【0061】
本明細書に記載するすべての刊行物、特許および特許出願は引用により本出願にその内容を含める。本発明を特定の態様に関して記載してきたが、さらなる改変が可能であり、本出願はあらゆる改変、使用または適用を包含する意図であることが理解されよう。 以下の、全体として本発明の主要部および本開示にはない本発明の属する公知または慣用技術も含めて、上記必須の特徴が適用することが可能であり、添付の請求の範囲に含まれる。
【図面の簡単な説明】
【0062】
【図1】図 1は、大腸菌 (AおよびB)および神経細胞培養物 (CおよびD) における発現に用いたFLIP-Eナノセンサーコンストラクトである。
【図2】図 2は、3種類のグルタミン酸濃度でのFLIP-E 600n センサー (グルタミン酸に対するKdが600 nMである蛍光グルタミン酸ナノセンサー)のスペクトルである: 0 mM (黒)、Kd 値の濃度(青)、および飽和濃度(赤)。これら曲線は520nmの等吸収点を共有する。
【図3】図 3は、 1 mg/ml トリプシンで処理された海馬細胞である。画像(A-D)は10 秒間隔で取得した。細胞表面上のシグナルが大きく消滅していることに注目されたい。
【図4】図 4は、FLIP-E 600n センサーを発現する海馬細胞における発光強度比変化である。画像は発光強度比変化を示すために疑似カラー表示している。グラフ (A)の上の白いバーは処理(刺激 /グルタミン酸の灌流)の時点を示す。矢印で示す時点での比画像(ratio image)をパネル(B)、a〜iに示す。発光強度比の変化が、電気的刺激とグルタミン酸の灌流との両方において観察された。比変化は低レベルの基質 (10 nM グルタミン酸)での灌流では観察されなかった。
【図5】図 5は、FLIP-E 10μ センサー(グルタミン酸に対するKdが10 μMである蛍光グルタミン酸ナノセンサー)を発現する海馬細胞における発光強度比変化である。グラフ (A)の上の白いバーは、処理 (刺激 / グルタミン酸の灌流)の時点を示す。矢印で示す時点での比画像をパネル(B)、a〜gに示す。電気的刺激によっては発光強度比に大きい変化は起こらなかったが、100 μM グルタミン酸での灌流によると可逆的な比変化が引き起こされた (パネル (B)、c およびe)。
【特許請求の範囲】
【請求項1】
以下を含むグルタミン酸結合蛍光指標をコードする単離核酸:
・大腸菌 YbeJ由来グルタミン酸結合タンパク質部分;
・グルタミン酸結合タンパク質部分に共有結合したドナー蛍光タンパク質部分;および、
・グルタミン酸結合タンパク質部分に共有結合したアクセプター蛍光タンパク質部分;
ここで、ドナー部分が励起され、グルタミン酸がグルタミン酸結合タンパク質部分に結合すると、ドナー部分とアクセプター部分との間の蛍光共鳴エネルギー移動 (FRET)が変化する。
【請求項2】
該グルタミン酸結合タンパク質部分が配列番号2または4の配列を含む、請求項 1の単離核酸。
【請求項3】
該ドナー蛍光タンパク質部分が、GFP、CFP、BFP、YFPおよびdsREDからなる群から選択される請求項 1の単離核酸。
【請求項4】
該アクセプター蛍光タンパク質部分が、GFP、CFP、BFP、YFPおよびdsREDからなる群から選択される請求項 1の単離核酸。
【請求項5】
該ドナー蛍光タンパク質部分が CFPであり、該アクセプター蛍光タンパク質部分がYFP Venusである、請求項 1の単離核酸。
【請求項6】
さらに少なくとも1つのリンカー部分を含む請求項 1の単離核酸。
【請求項7】
請求項 1の核酸を発現する細胞。
【請求項8】
請求項 1の核酸を含む発現ベクター。
【請求項9】
請求項 8のベクターを含む細胞。
【請求項10】
原核細胞において機能するのに適した請求項 8の発現ベクター。
【請求項11】
真核細胞において機能するのに適した請求項 8の発現ベクター。
【請求項12】
原核細胞である請求項 7の細胞。
【請求項13】
大腸菌である請求項 7の細胞。
【請求項14】
真核細胞である請求項 7の細胞。
【請求項15】
酵母細胞である請求項 7の細胞。
【請求項16】
動物細胞である請求項 7の細胞。
【請求項17】
請求項 1の核酸を発現するトランスジェニック動物。
【請求項18】
該トランスジェニック動物が線虫である請求項 17のトランスジェニック動物。
【請求項19】
グルタミン酸結合タンパク質部分のグルタミン酸に対する親和性を低下させる1以上の核酸置換をさらに含む請求項 1の単離核酸。
【請求項20】
該1以上の核酸置換が、A207G、A207P、A207K、A207M、A207S、A207C、A207R、A207V、A207L、A207Q、A207T、A207F、A207Y、A207N、A207W、A207H、A207D、およびS95Wからなる群から選択される請求項 1の単離核酸。
【請求項21】
請求項 1の核酸によりコードされるグルタミン酸結合蛍光指標。
【請求項22】
以下の工程を含む、神経細胞のサンプルにおける細胞外グルタミン酸レベルの変化の検出方法:
(a)請求項 1の核酸を発現する細胞および神経細胞のサンプルを提供する工程;および、
(b)該ドナー蛍光タンパク質部分と該アクセプター蛍光タンパク質部分との間のFRETの変化を検出する工程、
ここで、該ドナー部分と該アクセプター部分との間のFRETの変化は、神経細胞のサンプルにおける細胞外グルタミン酸レベルの変化を示す。
【請求項23】
FRETの判定工程がアクセプター蛍光タンパク質部分から放射される光を測定することを含む、請求項 22の方法。
【請求項24】
FRET の判定工程が、ドナー蛍光タンパク質部分から放射される光を測定すること、アクセプター蛍光タンパク質部分から放射される光を測定すること、およびドナー蛍光タンパク質部分から放射される光とアクセプター蛍光タンパク質部分から放射される光の比を計算することを含む、請求項 22の方法。
【請求項25】
FRETの判定工程がドナー部分の励起状態寿命を測定することを含む、請求項 22の方法。
【請求項26】
該神経細胞のサンプルがインビボに含まれる、請求項 22の方法。
【請求項27】
該神経細胞のサンプルがインビトロに含まれる、請求項 22の方法。
【請求項28】
該細胞外グルタミン酸レベルの変化が該神経細胞に対する外傷性障害によって起こる、請求項 26の方法。
【請求項29】
該細胞外グルタミン酸レベルの変化が、脳卒中、癲癇、ハンチントン病、エイズによる認知症および筋萎縮性側索硬化症からなる群から選択される障害に関連する、請求項 22の方法。
【請求項30】
以下の工程を含むグルタミン酸興奮毒性を調節する化合物を同定する方法:
(a)請求項 1の核酸を発現する細胞および神経細胞のサンプルを1以上の被験化合物と接触させる工程; および、
(b)該接触の後に該ドナー蛍光ドメインと該アクセプター蛍光ドメインの間のFRETを測定する工程、
ここで、該接触の後のFRETの上昇または低下は、該被験化合物がグルタミン酸興奮毒性を調節する化合物であることを示す。
【請求項1】
以下を含むグルタミン酸結合蛍光指標をコードする単離核酸:
・大腸菌 YbeJ由来グルタミン酸結合タンパク質部分;
・グルタミン酸結合タンパク質部分に共有結合したドナー蛍光タンパク質部分;および、
・グルタミン酸結合タンパク質部分に共有結合したアクセプター蛍光タンパク質部分;
ここで、ドナー部分が励起され、グルタミン酸がグルタミン酸結合タンパク質部分に結合すると、ドナー部分とアクセプター部分との間の蛍光共鳴エネルギー移動 (FRET)が変化する。
【請求項2】
該グルタミン酸結合タンパク質部分が配列番号2または4の配列を含む、請求項 1の単離核酸。
【請求項3】
該ドナー蛍光タンパク質部分が、GFP、CFP、BFP、YFPおよびdsREDからなる群から選択される請求項 1の単離核酸。
【請求項4】
該アクセプター蛍光タンパク質部分が、GFP、CFP、BFP、YFPおよびdsREDからなる群から選択される請求項 1の単離核酸。
【請求項5】
該ドナー蛍光タンパク質部分が CFPであり、該アクセプター蛍光タンパク質部分がYFP Venusである、請求項 1の単離核酸。
【請求項6】
さらに少なくとも1つのリンカー部分を含む請求項 1の単離核酸。
【請求項7】
請求項 1の核酸を発現する細胞。
【請求項8】
請求項 1の核酸を含む発現ベクター。
【請求項9】
請求項 8のベクターを含む細胞。
【請求項10】
原核細胞において機能するのに適した請求項 8の発現ベクター。
【請求項11】
真核細胞において機能するのに適した請求項 8の発現ベクター。
【請求項12】
原核細胞である請求項 7の細胞。
【請求項13】
大腸菌である請求項 7の細胞。
【請求項14】
真核細胞である請求項 7の細胞。
【請求項15】
酵母細胞である請求項 7の細胞。
【請求項16】
動物細胞である請求項 7の細胞。
【請求項17】
請求項 1の核酸を発現するトランスジェニック動物。
【請求項18】
該トランスジェニック動物が線虫である請求項 17のトランスジェニック動物。
【請求項19】
グルタミン酸結合タンパク質部分のグルタミン酸に対する親和性を低下させる1以上の核酸置換をさらに含む請求項 1の単離核酸。
【請求項20】
該1以上の核酸置換が、A207G、A207P、A207K、A207M、A207S、A207C、A207R、A207V、A207L、A207Q、A207T、A207F、A207Y、A207N、A207W、A207H、A207D、およびS95Wからなる群から選択される請求項 1の単離核酸。
【請求項21】
請求項 1の核酸によりコードされるグルタミン酸結合蛍光指標。
【請求項22】
以下の工程を含む、神経細胞のサンプルにおける細胞外グルタミン酸レベルの変化の検出方法:
(a)請求項 1の核酸を発現する細胞および神経細胞のサンプルを提供する工程;および、
(b)該ドナー蛍光タンパク質部分と該アクセプター蛍光タンパク質部分との間のFRETの変化を検出する工程、
ここで、該ドナー部分と該アクセプター部分との間のFRETの変化は、神経細胞のサンプルにおける細胞外グルタミン酸レベルの変化を示す。
【請求項23】
FRETの判定工程がアクセプター蛍光タンパク質部分から放射される光を測定することを含む、請求項 22の方法。
【請求項24】
FRET の判定工程が、ドナー蛍光タンパク質部分から放射される光を測定すること、アクセプター蛍光タンパク質部分から放射される光を測定すること、およびドナー蛍光タンパク質部分から放射される光とアクセプター蛍光タンパク質部分から放射される光の比を計算することを含む、請求項 22の方法。
【請求項25】
FRETの判定工程がドナー部分の励起状態寿命を測定することを含む、請求項 22の方法。
【請求項26】
該神経細胞のサンプルがインビボに含まれる、請求項 22の方法。
【請求項27】
該神経細胞のサンプルがインビトロに含まれる、請求項 22の方法。
【請求項28】
該細胞外グルタミン酸レベルの変化が該神経細胞に対する外傷性障害によって起こる、請求項 26の方法。
【請求項29】
該細胞外グルタミン酸レベルの変化が、脳卒中、癲癇、ハンチントン病、エイズによる認知症および筋萎縮性側索硬化症からなる群から選択される障害に関連する、請求項 22の方法。
【請求項30】
以下の工程を含むグルタミン酸興奮毒性を調節する化合物を同定する方法:
(a)請求項 1の核酸を発現する細胞および神経細胞のサンプルを1以上の被験化合物と接触させる工程; および、
(b)該接触の後に該ドナー蛍光ドメインと該アクセプター蛍光ドメインの間のFRETを測定する工程、
ここで、該接触の後のFRETの上昇または低下は、該被験化合物がグルタミン酸興奮毒性を調節する化合物であることを示す。
【図1】

【図2】

【図3】
【図4】
【図5】

【図2】
【図3】
【図4】
【図5】
【公開番号】特開2013−59335(P2013−59335A)
【公開日】平成25年4月4日(2013.4.4)
【国際特許分類】
【外国語出願】
【出願番号】特願2012−246314(P2012−246314)
【出願日】平成24年11月8日(2012.11.8)
【分割の表示】特願2007−536924(P2007−536924)の分割
【原出願日】平成17年10月14日(2005.10.14)
【出願人】(500026234)カーネギー インスチチューション オブ ワシントン (25)
【Fターム(参考)】
【公開日】平成25年4月4日(2013.4.4)
【国際特許分類】
【出願番号】特願2012−246314(P2012−246314)
【出願日】平成24年11月8日(2012.11.8)
【分割の表示】特願2007−536924(P2007−536924)の分割
【原出願日】平成17年10月14日(2005.10.14)
【出願人】(500026234)カーネギー インスチチューション オブ ワシントン (25)
【Fターム(参考)】
[ Back to top ]