
高活性糖タンパク質−製造条件、及びその効率的製造方法
【課題】最適化されたシアル酸付加度(sialylation degree)を有する高活性糖タンパク質、診断又は治療に用いるための該糖タンパク質を含む医薬組成物、高活性糖タンパク質及びその製造のための条件の決定方法、高活性糖タンパク質の製造方法、並びに分泌性糖タンパク質の示差的シアル酸付加方法の提供。
【解決手段】糖タンパク質を発現させるための、少なくとも1つのシアル酸前駆体添加物の所望の濃度を決定する。シアル酸の糖ヌクレオチド生合成経路における、細胞のシアル酸付加能の低下を引き起こす欠損を有する発現細胞系の作製。シアル酸付加された糖タンパク質を発現細胞系の細胞により分泌させる。
【解決手段】糖タンパク質を発現させるための、少なくとも1つのシアル酸前駆体添加物の所望の濃度を決定する。シアル酸の糖ヌクレオチド生合成経路における、細胞のシアル酸付加能の低下を引き起こす欠損を有する発現細胞系の作製。シアル酸付加された糖タンパク質を発現細胞系の細胞により分泌させる。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、最適化されたシアル酸付加度(sialylation degree)を有する高活性糖タンパク質、診断又は治療に用いるための該糖タンパク質を含む医薬組成物、高活性糖タンパク質及びその製造のための条件の決定方法、高活性糖タンパク質の製造方法、並びに分泌性糖タンパク質の示差的シアル酸付加方法に関する。また本発明は、生物学的目的のため、並びに、疾患(特に骨髄移植、好中球減少症、血球減少症、AML及び骨髄異形成症候群、癌、HIV及び/又は造血系疾患)の予防的及び/若しくは治療的処置、又は診断のための、組換え発現された高活性糖タンパク質の使用に関する。
【背景技術】
【0002】
糖タンパク質は、機能及び発生が各糖タンパク質間で大きく異なっている多様な一群である。潜在的治療能を有するタンパク質のうちのほとんどのタンパク質はグリコシル化されており、これは、例えば多数のホルモン(例えば、成長ホルモン、グリカゴン、FSH及びLH)、増殖因子(例えば、GM−CSF、G−CSF、VEGF及びエリスロポエチン)、サイトカイン(例えば、IL−2、IL−7、インターフェロンα及びβ、TNFα)、抗凝固剤(例えば、レピルジン(Lepirudin)、デシルジン(Desirudin))、血液凝固因子(例えば、第VII因子、第VIII因子及び第IX因子)、ワクチン(例えば、B型肝炎抗原)及び抗体で見られる。そのようなタンパク質を製造するための確立された細胞産生系は、元のヒトグリコシル化を有するタンパク質を産生することができない。原核生物(例えば、細菌)及びほとんどの真核細胞系(例えば、酵母、昆虫及び植物細胞)は、グリコシル化を欠くタンパク質を合成し、あるいはヒト糖鎖とは大きく異なるグリカンを含有するタンパク質を合成する。チャイニーズハムスター卵巣(CHO)細胞は、ヒト細胞と同様にタンパク質をグリコシル化しうる一般に使用されている産生系であるが、CHO細胞は、例えば、ガラクトシル化、フコシル化、特に、Nアセチルグリコサミンでのグリコシル化及び特にシアル酸付加の種々の態様において、いくつかの重要な相違を有する。
【0003】
これらの産生系が確立された時点においては、少なくとも或る程度活性である治療用タンパク質が産生されることで十分であった。今日では、(i)治療用タンパク質の適用用量の回数及び濃度を減少させるために、(ii)治療のコストを減少させるために、並びに(iii)副作用を軽減するために、治療用タンパク質の活性を改善することに集中した多大な努力がなされている。
【0004】
1つの方法は、発現系の生産性を増加させるものであるが、ほとんどの場合には、これはグリコシル化を全くもたらさないか非常に異常なグリコシル化しかもたらさず、ほとんどの場合には、それはタンパク質には不適当なものである。他の問題点、例えば分子の安定性及びフォールディングの低下も生じ、このため、しばしば、追加的な製造工程が必要となり、これは時間及びコストを浪費し、無駄、副作用及び最適未満の生物活性を与える。
【0005】
タンパク質の生物活性を改善するための主な方法は、その血清半減期を延長させ、したがってそのバイオアベイラビリティを向上させるものである。これは、PEG付加(PEGylation)と称される方法により行われ、この場合、産生されたタンパク質に或る形態のポリエチレングリコールが化学的に付加される。PEGは分子量を増加させ、したがって血清半減期を延長させる。しかし、この方法にはいくつかの問題が伴う。例えば、ほとんどすべての場合において、PEG付加はその細胞エフェクター機能によりタンパク質の活性を減少させ、ヒトにおける反復投与は、しばしば、中和抗体としての有害な免疫応答を引き起こし、及び/又は、該製造方法は追加的な化学修飾を要し、さらなるコスト、無駄及び時間の浪費を招く多段階の方法となる。
【0006】
糖鎖の修飾は、組換え発現されたタンパク質の血清半減期を改善することに焦点が合わされている。したがって、この技術は、組換え糖タンパク質のシアル酸付加度を最大にすることに焦点が合わされている。シアル酸は、真核細胞の表面上に最もよく見られる末端単糖であり、糖タンパク質のシアル酸付加が多くなればなるほど、循環中のその血清半減期は長くなる、と一般には考えられている。これは、循環非シアル酸付加タンパク質に結合し分解のためにそれを細胞内に導く肝臓内のアシアロタンパク質−受容体としての或る受容体の存在に基づくものである。
【0007】
組換え糖タンパク質のシアル酸付加度を可能な限り高くまで増加させるためのいくつかの方法が提示されており、これらには例えば、(i)ほとんどの末端にシアル酸付加をもたらすのに寄与する特定のシアリルトランスフェラーゼ又はガラクトシルトランスフェラーゼを過剰発現する組換え宿主細胞系を得ること、(ii)糖加水分解酵素を、例えばCuイオンを培地に加えて調節することにより、宿主細胞シアル酸の分解を妨げること、(iii)オリゴ糖産生を増強するための追加的な複合糖質及び他の基質の存在下、組換え糖タンパク質を産生する宿主細胞を培養すること、(iv)培養条件を種々変化させること、(v)追加的な糖鎖を導入するために、したがってより多数のシアル酸を導入するために、該タンパク質を突然変異させること、並びに(vi)該糖タンパク質の産生後、精製されたシアリルトランスフェラーゼでin vitroシアル酸付加を完了させることが含まれる。これらのすべての技術は一連の欠点を伴い、シアル酸付加度を最大にすることにより血清半減期を延長させるよう意図されているに過ぎない。
【0008】
前記のとおり、先行技術は一連の決定的な欠点を有し、生物活性の改善、副作用又は毒性の軽減のためのグリコシル化の可能性のごく一部しか検討しておらず、したがって糖タンパク質を何らかの用途に結びつけるものではない。対応する組換え糖タンパク質に関する適当な試験系及び産生系が全く利用できないため、先行技術は、糖タンパク質の生物活性がシアル酸付加度と相関しているかどうかについての認識をほとんど完全に欠いている。
【発明の概要】
【0009】
本発明は、シアル酸の糖ヌクレオチド生合成経路における少なくとも1つの欠損を含有し、かつ糖タンパク質をコードする核酸でトランスフェクトされた発現細胞系において、ある濃度の少なくとも1つのシアル酸前駆体添加物を添加した培地内で高活性糖タンパク質を発現させることを含む方法により製造される高活性糖タンパク質であって、
該濃度が、
(i)種々の濃度の少なくとも1つのシアル酸前駆体を使用する示差的シアル酸付加により、複数の異なるシアル酸付加形態の糖タンパク質を発現させること、
及び
(ii)適当なバイオアッセイにおいて、参照糖タンパク質と比較して、前記の異なるシアル酸付加形態の活性を測定すること、
及び
(iii)より高い/最高の活性を有するシアル酸付加形態を選択し、該糖タンパク質のより高い/最高の活性レベルと相関する、該シアル酸前駆体添加物の濃度を決定すること
を含む方法により決定される、高活性糖タンパク質を提供することにより、この課題を解決するものである。
【0010】
したがって、また本発明は方法に関する。
【図面の簡単な説明】
【0011】
【図1】NM−wt細胞のグライコエンジニアリング(GlycoEngineering)及び得られたNM−F9細胞の特徴決定を示す。
【図2】NM−wt細胞並びに誘導体NM−F9及びNM−D4によるTF、sTF、Tn、sTn、グリコホリンA(GPA)、アシアログリコホリンA(AGPA)、Lex、sLex及びLeyの発現を示す。
【図3】(A)レクチンPNA、SNA、MAA及びUEAIを使用するフローサイトメトリーによる、あるいは(B)チオバルビツール酸法による、NM−wt細胞並びに誘導体NM−F9及びNM−D4上の膜複合糖質結合シアル酸の測定を示す。
【図4】(A)mRNA発現分析による、及び(B)酵素活性分析による、NM−F9細胞の遺伝的欠損の分析を示す。
【図5】代謝的相補によるシアル酸付加の復元を示す。
【図6】遺伝的相補によるシアル酸付加の復元を示す。
【図7】NM−F9におけるrhGM−CSFの発現を示す。
【図8】種々の分泌シグナルペプチドを使用した場合の、NM−F9細胞による、組換え発現された分泌性AGPAの発現を示す。
【図9】代謝改変による膜複合糖質の示差的シアル酸付加を示す。
【図10】代謝的相補による細胞表面上の膜タンパク質の示差的シアル酸付加を示す。
【図11】代謝的相補による分泌性rhGM−CSFの示差的シアル酸付加を示す。
【図12】TF1細胞上の示差的シアル酸付加rhGM−CSFの増殖活性を示す。
【図13】樹状細胞上の示差的シアル酸付加rhGM−CSFの増殖活性を示す。
【図14】rhGM−CSF濃度に対する増殖活性の依存性を示す。
【図15】rhGM−CSF増殖活性の抑制を示す。
【発明を実施するための形態】
【0012】
本発明において、「参照糖タンパク質」という用語は、本発明の糖タンパク質と同一又は類似のタンパク質配列を有する糖タンパク質、好ましくは、同じタンパク質配列を有する糖タンパク質を意味する。グリコシル化構造を欠くことも可能である。参照糖タンパク質は一定の活性を有し、天然由来又は組換え発現されたものでありうる。好ましい参照体は公知の組換え糖タンパク質である。活性、例えばin vitroにおける受容体媒介活性の改善をもたらす、先行技術の特定のタンパク質配列の突然変異の場合には、本発明のより高い活性の糖タンパク質は、この点に関してより高い活性であることが可能であり、及び/又は、前記活性はより良くなることなく別の生物活性、例えば免疫原性に関してより高い活性でありうる。これらの具体例においては、免疫原性におけるより高い活性は、哺乳動物、好ましくはヒトにおける使用に有利であり、したがって本発明における、より高い活性の糖タンパク質に匹敵する。当業者であれば、前臨床及び/又は臨床試験計画における適当な選択基準を選択することが可能である。
【0013】
本発明において、「発現細胞系」なる語は、当業者に公知の又は本明細書に記載の技術による、標的遺伝子の更なる組換え発現又はウイルス若しくは当業者に公知の適当な物質の感染を伴う又は伴わない、タンパク質、糖タンパク質、ウイルス又は他の生物物質の発現に使用しうる細胞又は細胞系を意味する。
【0014】
本発明において、「シアル酸の糖ヌクレオチド生合成経路における少なくとも1つの欠損を含有する発現細胞系」なる表現は、CMP−シアル酸の合成に関与する酵素に欠損を含有する発現細胞系を意味する。欠損は、対応する酵素活性が減少しているか又は完全に存在しないことを意味し、例えば、酵素の遺伝子、遺伝子の発現、酵素の活性、又はその機能に関連した酵素のバイオアベイラビリティのレベルでの種々の主要欠損によるものでありうる。酵素としては、例えばUDP−N−アセチルグルコサミン−2−エピメラーゼ、キナーゼ(例えば、N−アセチルマンノサミンキナーゼ、N−アセチルグルコサミンキナーゼ)、N−アセチルノイラミン酸−9−P(Neu5Ac−9−P)−シンテターゼ、Neu5Ac−9−P−ホスファターゼ及びCMP−Neu5Acシンテターゼが挙げられる。シアル酸の糖ヌクレオチド生合成経路における、より好ましい欠損は、エピメラーゼの突然変異であり、より一層好ましいのは、UDP−N−アセチルグルコサミン−2−エピメラーゼにおける、そのmRNA発現の欠損を引き起こす欠損である。
【0015】
該欠損は、標準的な条件下及び/又は無血清条件下での細胞のシアル酸付加の軽減又はシアル酸付加のほぼ完全な若しくは完全な欠如をもたらす。このシアル酸の軽減は、シアル酸前駆体添加物の添加により部分的又は完全に復元されうる。この欠損は、CMP−シアル酸の合成に関与する1以上の酵素の欠損に限定されるものではなく、シアル酸前駆体添加物の添加により該効果が部分的又は完全に復元されうるものである限り他の酵素の欠損でありうる。したがって、限定的なものではないが、一例としては、CMP−シアル酸輸送体としての、対応する糖輸送酵素が挙げられる。当業者であれば、どのような単一の欠損又は欠損の組合せが本発明に適しているかを判定することが可能であり、それらのいくつかは以下に更に詳しく説明されている。
【0016】
本発明において、「トランスフェクション」という用語は、ウイルス感染又はトランスフェクション方法(限定的なものではないが、リン酸カルシウム共沈法、エレクトロポレーション、DEAE−デキストラン又はカチオン性脂質試薬との複合体形成及びマイクロインジェクションが含まれる)により組換えタンパク質の発現のために細胞内へ遺伝物質を送達し、それにより発現細胞系の細胞内に核酸を導入するのに用いられうる当業者に公知の方法を意味する。
【0017】
本発明において、「糖タンパク質をコードする核酸」なる表現は、対象の哺乳動物糖タンパク質又はその活性断片及び/若しくは突然変異体(したがって、任意の糖タンパク質、好ましくは、ヒト由来の任意の糖タンパク質が使用されうる)をコードする核酸配列を意味する。哺乳動物糖タンパク質の具体例には、サイトカインのような分子及びそれらの受容体、例えば、腫瘍壊死因子TNF−α及びTNF−β;レニン;ヒト成長ホルモン及びウシ成長ホルモン;成長ホルモン放出因子;副甲状腺ホルモン;甲状腺刺激ホルモン;リポタンパク質;α−1−抗トリプシン;インスリンA鎖及びB鎖;性腺刺激ホルモン、例えば濾胞刺激ホルモン(FSH)、黄体形成ホルモン(LH)、甲状腺刺激ホルモン及びヒト絨毛性性腺刺激ホルモン(hCG);カルシトニン;グルカゴン;凝固因子、例えば第VIIC因子、第IX因子、第VII因子、組織因子及びフォンビルブラント因子;抗凝固因子、例えばプロテインC;心房性ナトリウム利尿因子;肺界面活性物質;プラスミノーゲン活性化因子、例えばウロキナーゼ、ヒト尿及び組織型プラスミノーゲン活性化因子;ボンベシン;トロンビン;造血増殖因子;エンケファリナーゼ;ヒトマクロファージ炎症性タンパク質;血清アルブミン、例えばヒト血清アルブミン;ミュラー抑制性物質;レラキシンA鎖及びB鎖;プロレラキシン;マウス性腺刺激ホルモン関連ペプチド;血管内皮増殖因子;ホルモン又は増殖因子に対する受容体;インテグリン;プロテインA及びD;リウマチ因子;神経栄養因子、例えば骨由来神経栄養因子、ニューロトロフィン3、4、5、6及び神経成長因子β;血小板由来増殖因子;繊維芽細胞増殖因子;表皮増殖因子;トランスフォーミング増殖因子、例えばTGFα及びTGFβ;インスリン様増殖因子I及びII;インスリン様増殖因子結合タンパク質;CDタンパク質、例えばCD3、CD4、CD8及びCD9;エリスロポエチン(EPO);骨誘導因子;イムノトキシン;骨形成タンパク質;インターフェロン、例えばインターフェロンα、β及びγ;コロニー刺激因子(CSF)、例えばM−CSF、GM−CSF及びG−CSF;インターロイキン(IL)、例えばIL−1〜IL−12;スーパーオキシドジスムターゼ;T細胞受容体;表面膜タンパク質;崩壊促進因子;抗体及びイムノアドヘシン;グリコホリンA;MUC1が含まれる。
【0018】
好ましい糖タンパク質は、グリコホリンA、EPO、G−CSF、GM−CSF、FSH、hCG、LH、インターフェロン、インターロイキン、抗体及び/又はそれらのフラグメントを含む群から選ばれる。
【0019】
本発明において、「糖タンパク質」という用語は、ウイルス、ウイルス粒子又はウイルスタンパク質としても定義される。
【0020】
本発明の好ましい実施形態においては、核酸は糖タンパク質の分泌性形態又はその断片をコードしている。好ましい実施形態においては、該分泌性形態は膜貫通ドメインを欠く。別の好ましい実施形態においては、該分泌性形態をコードする核酸は少なくとも1つの分泌シグナルを含む。最も好ましい実施形態においては、該分泌シグナルはGM−CSF由来である。他の実施形態及び詳細は以下で更に詳しく説明されている。
【0021】
本発明において、「シアル酸前駆体添加物」なる語は、糖タンパク質の部分的又は完全なシアル酸付加をもたらす、シアル酸の糖ヌクレオチド生合成経路における欠損を部分的又は完全に復元しうる任意の天然又は合成化合物を意味する。本発明の好ましい実施形態においては、種々の濃度のシアル酸前駆体添加物の添加は、種々のシアル酸付加度を有する糖タンパク質の一連の異なるシアル酸形態を与える。それにより、糖タンパク質のシアル酸付加形態は単一のグライコフォーム又は一群のグライコフォームから構成されうる。これは、単一のシアル酸前駆体添加物を使用することにより、又は2以上の前駆体添加物の組合せを使用することにより達成されうる。本発明の好ましい実施形態においては、シアル酸前駆体添加物として、シアル酸中間体及び/又はシアル酸含有糖タンパク質を使用する。別の好ましい実施形態においては、シアル酸前駆体添加物はManNAc、アセチル化ManNAc、パーアセチル化ManNAc又はフェツインである。別の好ましい実施形態においては、ManNAc、アセチル化ManNAc又はパーアセチル化ManNAcとフェツインとの組合せを使用する。
【0022】
好ましいのは、天然シアル酸修飾を有する糖タンパク質を与えるシアル酸前駆体添加物である。天然シアル酸修飾は、出発物質、中間体グライコフォーム又は最終グライコフォームのような、生物において見出されうる修飾である。さらに、天然糖タンパク質、グリコシル化形態(これらは、生物体内に天然で存在するグライコフォームに可能な限り近い)が好ましい。グライコフォーム、例えば、ヒトにおいて見出される糖鎖又はヒト由来の任意の生物学的物質に類似した、シアル酸を有する糖鎖を有する、ヒト糖タンパク質のグライコフォームが使用されるであろう。それらの好ましい形態は、生物において見出されない非天然側鎖により修飾されたシアル酸を含有しない。例えば、糖タンパク質又は糖脂質内に組込まれるシアル酸は、化学合成により導入されうる、置換された若しくは欠失した若しくは付加された若しくは伸長した若しくは短縮した側鎖、及び/又は生物において見出されない、シアル酸への新規鎖間連結(interlinkage)を含まない。
【0023】
本発明の好ましい形態においては、いずれの細胞生成物のグリコシル化も、非天然シアル酸修飾を与える化学的に操作された前駆体又は代謝産物によって変化させたり又は修飾するのではなく、天然グリコシル化、好ましくはシアル酸付加の度合を最適化することにより変化させたり又は修飾する。化学修飾されたシアル酸代謝産物は、天然グリコシル化に影響を及ぼさない様態でのみ使用されうる。例えば、アセチル化ManNAc又はパーアセチル化ManNAcは天然N−アセチルマンノサミン(ManNAc)より遥かに容易に細胞により取り込まれるため、アセチル化ManNAc又はパーアセチル化ManNAcを添加した細胞培養培地としてManNAcは置換されうる。細胞内で、追加的なアセチル残基が切り離され、CMP−シアル酸の生合成のためにManNAcがプロセシングされる。したがって、アセチル化ManNAcで処理された細胞により産生される糖タンパク質は、合成により修飾された炭水化物(例えば、合成によりアセチル化されたシアル酸)を何ら示さない。
【0024】
本発明において、「異なるシアル酸付加形態」なる表現は、少なくとも1つのシアル酸が別のグライコフォームとは異なる糖タンパク質のグライコフォームを意味する。シアル酸における相違は、シアル酸の量、及び/又は該分子の残部上の少なくとも1つのシアル酸の位置におけるものでありうる。個々のシアル酸付加形態は、糖タンパク質の単一のグライコフォーム又はいくつかのグライコフォームの混合物から構成されることが可能であり、それにより、異なるシアル酸付加形態はお互いに、少なくとも1つのグライコフォームにおいて及び/又はグライコフォームの相対組成において異なる。
【0025】
少なくとも1つのシアル酸付加形態のシアル酸付加度は、当業者に公知の生化学的及び/又は化学的方法により測定することが可能である。例えば、迅速な方法は、シアル酸特異的抗体又はレクチン、例えばSNA及びMAAで、あるいはシアル酸を含まないグリカンを認識する抗体又はレクチン、例えばA78−G/A7及びNemod−TF1又はレクチンPNAで、細胞又は糖タンパク質を染色するものである。染色された細胞はフローサイトメトリーにより分析し、染色された糖タンパク質は、糖タンパク質が精製されている場合には通常のELISAにより、あるいは分析対象の糖タンパク質を捕捉するために糖タンパク質特異的抗体を使用するサンドイッチELISAにより分析することが可能である。当業者に公知のこれらのすべての方法は、細胞又は糖タンパク質の示差的シアル酸付加のモニターを可能にする。したがって、結合したシアル酸特異的抗体又はレクチンの量は、細胞又は糖タンパク質に結合したシアル酸の量と相関し、一方、シアル酸を含まないグリカンを認識する結合抗体又はレクチンの量は、細胞又は糖タンパク質に結合したシアル酸の量と逆相関する。シアル酸含量を定量するためのもう1つの方法は、ノイラミニダーゼを使用することにより酵素的に、あるいは酸加水分解により化学的に、糖タンパク質に結合したすべてのシアル酸を遊離させるものである。ついでサンプル中のシアル酸の濃度を、1,2−ジアミノ−4,5−メチレンジオキシベンゼン(DMB)でのシアル酸の蛍光測定染色、及び蛍光測定検出器を使用するHPLC分析により測定する。あるいは、遊離したシアル酸の量を、チオバルビツレート及びβ−ホルミルピルベートの存在下、549nmで比色検出されうるピンク色色素にシアル酸を変換することにより定量する。N−又はO−グリカンに結合したシアル酸の数に関するデータを得るためには、全N−又はO−グリカンを酵素的又は化学的に遊離させる。アンテナ性(antennarity)に関する遊離グリカン(例えば、HPAEC−PADにより)、結合シアル酸の数(例えば、陰イオン交換クロマトグラフィーと組合されたHPAEC−PADにより)、単糖の組成(例えば、グリカンの酸加水分解の後のHPAEC−PADにより)及び/又は厳密な構造(例えば、質量分析)を分析するためのいくつかの方法が当業者に利用可能である。
【0026】
本発明における定義において、「活性」は、生物学的環境中で分子により発揮される機能又は一連の機能である。一般には、本発明の方法により製造される糖タンパク質は、先行技術により製造される糖タンパク質より活性が高く及び/又はより有効な生物学的産物である。したがって、本発明は、ヒト高活性糖タンパク質、及び該糖タンパク質を含む診断、治療又は他の生物学的系及び試験におけるその使用に関する。本発明における定義においては、「高活性糖タンパク質」なる語、その内容の均等物及びその文法的均等物は、選択された用途に関して、改善された又は最適の活性として理解されるべきであり、したがって、該活性は極限値に近似されることが可能であり、例えば最小化又は最大化されることが可能であり、あるいは、先行技術により製造された対応糖タンパク質及び/又は本発明の示差的シアル酸付加により製造された別のシアル酸付加形態より高い又はより低い活性を表す中間値に設定されることが可能であろう。本発明における改善された又はより高い活性はまた、その生物学的及び/又は薬学的意味において好ましい活性を意味する。例えば、糖タンパク質の生物活性は、有害な生物学的影響を軽減することにより(例えば、有害な免疫作用の刺激の低減又は免疫原性の低減により)或る程度は増強されうるであろう。本発明の方法は、最適化された血清半減期、薬物動態、安定性、抗原性及び/又は免疫原性を有する糖タンパク質を製造するために有効に使用されうる。
【0027】
本発明において、「異なるシアル酸付加形態の活性の測定」なる表現は、糖タンパク質の活性を測定しうる適当なバイオアッセイにおける活性の測定を意味する。当業者であれば、適当なバイオアッセイを決定し、適当なバイオアッセイを構成することが可能である。本発明においては、そのようなバイオアッセイとしては、例えば、生物学的in vitroアッセイ、例えば細胞若しくは分子又は混合アッセイ、例えば増殖アッセイ、アポトーシスアッセイ、細胞接着アッセイ、シグナル伝達アッセイ、遊走アッセイ、細胞毒性アッセイ、食作用アッセイ、細胞溶解アッセイ及び結合アッセイが挙げられる。そのようなバイオアッセイにはまた、動物モデル又はヒトを使用するin vivoアッセイ、例えば生体内分布、薬物動態、薬動力学試験、血清半減期試験、バイオアベイラビリティに関する試験、有効性試験、局在性試験、疾患の治療及び予防試験(臨床研究を含む)が含まれる。そのようなバイオアッセイには、化学的、物理的、物理化学的、生物物理学的及び生化学的試験、例えば温度、せん断応力、圧力、pH、複合体形成(コンジュゲーション)などに対する安定性も含まれる。そのようなバイオアッセイには、糖タンパク質の臨床的使用に関する該糖タンパク質の特性を改善するための免疫原性及び/又は抗原性に関する試験も含まれる。当業者であれば、糖タンパク質及びそのシアル酸付加形態の活性又は記載されている活性の組合せを決定することが可能である。
【0028】
好ましい実施形態においては、糖タンパク質のより高い活性は、少なくとも1つのin vitroモデルにおける、り高い活性、及び/又は少なくとも1つのin vivoモデルにおけるより高い活性、及び/又はより高い安定性、及び/又はより長い血清半減期、及び/又はより長いバイオアベイラビリティ、及び/又は免疫原性の改善、及び/又は抗原性の改善(少なくとも1つのバイオアッセイにより測定される)を特徴とする。全活性における改善(これは本明細書中では、より高い活性とも称される)は、例えば、ヒト又は対応生物において使用される場合の該生成物の、より低い投与量、より長い投与時間間隔、より低い副作用、及び全く無い又はより低い毒性をもたらして、大きく改善された医薬を与えうる。
【0029】
本発明において、「より高い活性を有するシアル酸付加形態の選択」という用語は、バイオアッセイの少なくとも1つにおける比較において、より高い活性を有する糖タンパク質のシアル酸付加形態の選択を意味する。この比較は、本発明の方法により発現される少なくとも1つのシアル酸付加形態を先行技術のグリコシル化形態の糖タンパク質と比較することにより、あるいは本発明の方法により発現される少なくとも2つのシアル酸付加形態を比較することにより行いうる。好ましくは、相当に最適化されたグライコフォームを達成し、同定し、製造するためには、高活性糖タンパク質は3以上の異なるシアル酸付加形態から選ばれる。当業者であれば、どのシアル酸付加形態が、特定のバイオアッセイに関して、より高い活性を有するかを決定することが可能である。好ましい実施形態においては、より高い活性を有するシアル酸付加形態を選択するためには、バイオアッセイの組合せの活性を考慮する。その場合、それぞれのバイオアッセイが、より高い活性を示す必要があるわけではなく、個々の糖タンパク質の用途及び特徴に応じて、いくつかの好ましい生物学的作用が、それほど好ましくはない他のものを補うことが可能であり、本発明においては尚も全体的にはより高い活性のグライコフォームをもたらしうる。例えば、あるグライコフォームは、細胞に対してその受容体に結合することにより、より遥かに高い活性をもたらして、例えば増殖の誘導のような副次的効果を誘導しうるが、若干減少した血清半減期を示す。したがって、組合せとしては、該受容体をより惹起するより高い活性は、全体的な生物活性においてバイオアベイラビリティが短くなることを補う。もう1つの例においては、より短い半減期、及び受容体惹起をもたらす、より高い活性は、どちらも有利である。さらにもう1つの例においては、in vivoでの活性は改善されないが、in vitroでの安定性は糖タンパク質の製造及び貯蔵を改善する。さらにもう1つの例においては、長い半減期の一方で、より低い活性が必要とされる。したがって、実際の有利なシアル酸付加形態を、特定の糖タンパク質に基づいて、その後の使用に関して決定する必要がある。本発明は、この統合された系において、これを初めて可能にするものである。このすべての態様は、決定すべき単一の糖タンパク質に左右され、これらは大きく異なり、予想できず、本発明の後述する方法を必要とする。本発明を使用することにより、当業者は、適当なバイオアッセイ及び/又はバイオアッセイの組合せ、並びに前記の点に関して糖タンパク質の所望の用途のためのより高い活性を有する糖タンパク質のシアル酸付加形態を選択することが可能であり、したがって、標準的な方法により製造されうるグリコシル化形態と比べて糖タンパク質を改善することが可能である。本発明を使用することにより、当業者は、該糖タンパク質のより高い活性レベルと相関する、シアル酸前駆体添加物又はシアル酸前駆体添加物の組合せの濃度を決定することが可能である。
【0030】
本発明において、示差的シアル酸付加により発現させバイオアッセイにおいて比較する複数の異なるシアル酸形態という表現は、バイオアッセイにおいて試験した場合に、先行技術により得られる少なくとも1つの異なるシアル酸付加又はグリコシル化形態と比べて高い活性を示す、本発明の方法により発現される1つのシアル酸形態をも意味する。
【0031】
前記のすべての定義及び説明は、前記及び後記の糖タンパク質及び方法にも有効である。
【0032】
驚くべきことに、本発明の組換え発現された糖タンパク質は、シアル酸前駆体添加物の特定の濃度に対応する特定の度合でシアル酸付加された場合に、最高の生物活性を示すことを見出した。最高の生物活性と相関するこのシアル酸付加度は、分析する各糖タンパク質によって異なり、予測可能ではなく、最高のシアル酸付加度が必ずしも達成可能なわけではない。本発明に従って糖タンパク質のシアル酸付加度を同定するためには、細胞欠損によってシアル酸付加の復元及び種々のシアル酸付加度の達成が可能となるようにシアル酸付加能が欠損した細胞を使用及び/又は作製する。シアル酸付加の復元を達成するための1つの方法は、シアル酸生合成経路のシアル酸前駆体添加物を該細胞の培地に添加することによるものである。該欠損細胞及び少なくとも1つのシアル酸前駆体添加物を使用して、該培地に加えるシアル酸前駆体添加物の量を変化させることにより、該細胞により産生される本発明の任意の糖タンパク質のシアル酸付加度を示差的に制御することが可能である。少なくとも2つの異なるシアル酸付加形態が達成されうる。僅か2つの異なる濃度の前記添加物を使用する場合には、異なるシアル酸付加形態の作製が保証されるよう、濃度勾配は十分大きくなければならない。好ましくは、最高値が方法の特徴(例えば、粘度、溶解度、aw値など)により定められる高濃度及び低濃度が選ばれる。示差的にシアル酸付加された糖タンパク質の少なくとも2つの形態、好ましくは複数の形態(異なるシアル酸形態)を単離し、グリコシル化、好ましくはシアル酸付加、活性、血清半減期、薬物動態、薬動力学、in vivo生物活性、抗原性及び免疫原性のような特性に関して分析することが可能である。そのようにして、該糖タンパク質の、その活性又は任意の他の特性のための最適なシアル酸付加度は、前記の基準の1つ又は組合せにより同定され、その最適シアル酸付加糖タンパク質の製造のための条件は、生産細胞として上記欠損細胞を使用する製造プロセスに移されうる。
【0033】
本発明はまた、高活性糖タンパク質の製造のための特別な方法及び高活性糖タンパク質そのものを教示する。これらの特別な場合には、シアル酸前駆体添加物の最適濃度が他の技術により決定され又は当業者の論理的思考から推定されうる場合には(例えば、後記実施例に記載のとおり)、個々のシアル酸付加形態は、それを、異なる濃度のシアル酸前駆体添加物の対応発現細胞系内での発現により得られる第2の形態と比較することを要することなく、製造されうる。これらの特別な場合も、高活性糖タンパク質及び本発明の方法において、明らかに本発明に包含される。これらの場合、本発明の基本的な発明原理は尚も、本方法及び得られる高活性糖タンパク質の中心的かつ決定的な部分である。これに関しては、前記及び後記の他の説明も参照されたい。一例としては、シアル酸付加を全く又は最小限度しか伴わないGPAが挙げられ、これは後記の実施例の1つにおいて更に詳しく説明されており、この場合の目的は、初めから、最小限度のシアル酸付加及び最大限度のトムセン−フリーデンライヒ(Thomsen-Friedenreich)を伴う形態の作製でありうる。したがって、シアル酸前駆体添加物を加えることなく、好ましくは、培地内にいずれのシアロ糖タンパク質も存在することなく、最低限度のシアル酸付加を伴う形態のみを作製することで十分でありうる。更なる使用のためのこの形態の適合性は、それを、少なくとも1つの他のシアル酸付加形態と比較することにより判定されうるが、この代わりに、ヒト由来のシアル酸付加又は低シアル酸付加GPAとも比較しうる。もう1つの例は、最高シアル酸付加が望ましいことが判明しており、最大限度のシアル酸前駆体添加物を使用することで十分でありうる場合である。しかし、後者の場合には、ほとんどすべての場合において、本明細書中の種々のシアル酸付加形態を作製することによりシアル酸前駆体添加物の組成及び濃度を最適化することにより最高シアル酸付加を最適化することが有利である。この特別な場合には、単一のバイオアッセイとしてのシアル酸度の測定のための技術を用いることで十分かもしれないが、他のバイオアッセイとの組合せが好ましい。
【0034】
新規技術は、新規糖タンパク質の作製及び種々の糖タンパク質の改善(改良バイオジェネリックス(biogenerics)の作製を含む)のための高い潜在性を保有する。
【0035】
以下において、シアル酸の糖ヌクレオチド生合成経路における欠損を更に詳しく説明する。本発明においては、細胞のシアル酸付加能の低下を引き起こす欠損(好ましくはシアル酸付加能の完全な消失)を有し該欠損が復元されうることにより示差的制御シアル酸付加を可能にする欠損を有する発現細胞系により、高活性糖タンパク質を製造する。「シアル酸の糖ヌクレオチド生合成経路における欠損」とも称される該細胞の分子的欠損は、シアル酸の代謝に関与するタンパク質(例えば、糖輸送における例えばCMP−シアル酸輸送体、又はCMP−シアル酸の生合成における例えばキナーゼ、デヒドロゲナーゼ、ホスファターゼ、シンテターゼ、トランスケトラーゼ、トランスアルドラーゼ、イソメラーゼ、トランスフェラーゼ及びエピメラーゼ)の機能喪失(loss-of-function)でありうる。また、シアル酸付加の軽減又は欠如を伴うシアル酸付加不能細胞を与える分子的欠損は、シアル酸の代謝に関与するタンパク質(例えば、炭水化物からシアル酸を遊離させる酵素、例えば、ノイラミニダーゼ)の機能獲得(gain-of-function)でありうる。好ましい標的は、CMP−シアル酸の輸送に寄与するタンパク質、及びシアル酸生合成経路における律速段階を触媒する酵素である。より好ましくは、該欠損はエピメラーゼ、最も好ましくはUDP−GlcNAc−2−エピメラーゼに関するものである。
【0036】
先行技術は、N−アセチルマンノサミン(ManNAc)がCMP−シアル酸の生合成において重要な役割を果たすことを教示している。ManNAcは、シアル酸の細胞生合成における律速段階を代表するUDP−GlcNAc−2−エピメラーゼの作用により、UDP−N−アセチルグルコサミン(UDP−GlcNAc)から形成される。ついでManNAcは特異的キナーゼ(例えば、N−アセチルマンノサミンキナーゼ、N−アセチルグルコサミンキナーゼ)によりリン酸化される。ManNAc−6−PはNeu5Ac−9−P−シンテターゼによりホスホエノールピルビン酸と縮合してN−アセチルノイラミン酸−9−P(NeuAc−9−P)となり、ついで該ホスフェートはNeu5Ac−9−P−ホスファターゼにより遊離され、Neu5AcはCMP−Neu5Acシンテターゼにより活性化される。ついでCMP−Neu5Acは特異的輸送体(例えば、CMP−シアル酸輸送体)により細胞のゴルジ区画内へ輸送され、その部位において、それは、シアル酸を糖タンパク質又は糖脂質の末端ガラクトース(Gal)−、N−アセチルガラクトサミン(GalNAc)−又はN−アセチルグルコサミン(GlcNAc)−残基へ転移させる種々のシアリルトランスフェラーゼ、例えばST6GlcNAcシアリルトランスフェラーゼ、ST6GalNAc−I〜VI、ST3Gal−I〜VI及びST6Gal−Iのための供与体として働く。
【0037】
シアル酸生合成の各酵素、例えばUDP−GlcNAc−2−エピメラーゼ、キナーゼ、Neu5Ac−9−P−シンテターゼ、Neu5Ac−9−P−ホスファターゼ及びCMP−Neu5Acシンテターゼ、又はゴルジ区画内への輸送に必要な各タンパク質は、欠損細胞に欠損酵素の産物を供給することにより該欠損が復元されうるように、細胞のシアル酸付加能を改変するよう、好ましくは軽減するよう、より好ましくは完全に遮断されるよう影響を受けることが可能である。細胞のシアル酸付加能は、シアリルトランスフェラーゼにより触媒される炭水化物へのシアル酸の転移における変化によっても影響されうる。
【0038】
該細胞欠損を生じさせ、遺伝的、転写、翻訳又はタンパク質レベルでタンパク質の合成及び機能を標的化するよう特異的に該細胞欠損を作製するには、多数の選択肢が存在する。
【0039】
特定のタンパク質をコードする遺伝子における突然変異は、しばしば、そのタンパク質の機能喪失を引き起こし、あるいは、時には、機能を獲得するタンパク質を与える。例えば、1個若しくは2個のヌクレオチドの置換により又は(1+n)個若しくは(2+n)個のヌクレオチド(ここで、nは3の倍数である正の整数)の導入若しくは欠失により引き起こされうるフレームシフトにより、遺伝子のコード領域内に停止コドンを与える突然変異が挙げられる。多数の他の「機能喪失」及び「機能獲得」遺伝子突然変異が当業者に公知である。これらの遺伝子突然変異は遺伝子の転写にも影響を及ぼしうる。化学的突然変異原、例えばメタンスルホン酸エチル(EMS)又は物理的突然変異原、例えばUV光により遺伝的突然変異をランダムに誘発するための多数の方法が当業者に公知である。また、現代の分子生物学的技術によって、当業者は突然変異により遺伝子を特異的に標的化することが可能である。これらの技術を用いて、当業者は特定の遺伝子をノックアウトしたり、あるいは新たな遺伝子を導入して機能獲得突然変異体を作製することが可能である。
【0040】
転写レベルでは、特定のタンパク質をコードするmRNAの合成が制御される。遺伝子転写における欠損又は変化は、特定の酵素のmRNA合成のスイッチをオフ又はオンにすることができる。例えば、シアル酸の生合成に関与する酵素をコードするDNAのメチル化、又はDNA結合タンパク質、例えばヒストン、転写因子若しくは転写装置のタンパク質の修飾、例えばアセチル化は転写を妨げたり、あるいは特定の遺伝子の転写のスイッチをオンにすることが可能であろう。前記の及び当業者に公知の遺伝学的及び分子生物学的方法による突然変異の誘発は、任意の遺伝子の転写効率に影響を及ぼすための1つの選択肢である。NM−F9細胞内のUDP−GlcNAc−2−エピメラーゼmRNAの欠如(その欠損はEMS突然変異原での野生型細胞の処理により作製される)は、機能喪失欠損を作製するための転写の影響の一例である。
【0041】
翻訳レベルでは、特定の酵素の、合成されたmRNAを標的化して、好ましくは、リボソームタンパク質の生合成を妨げ又は軽減する。具体例としては、共に特異的mRNA分子を標的化するアンチセンス分子又は小さな干渉性RNA分子を使用する翻訳の阻止及び/又はRNaseによる特定のmRNAの分解が挙げられる。どちらの技術も当業者に公知である。
【0042】
タンパク質レベルでは、酵素活性の欠如は遺伝子突然変異により引き起こされうるであろう。可能な突然変異には、少なくとも1つのアミノ酸の置換、欠失及び/又は修飾が含まれる。該突然変異は、新たな及び非天然構造が基質の結合及び/又はその変換を許容しないタンパク質の構造的再折りたたみによる、あるいは基質との相互作用に寄与する活性中心内の残基を変化させることによる、酵素の触媒特性に関するものでありうる。そのような突然変異を作製するための技術は前記で説明されている。
【0043】
CMP−シアル酸の生合成又は輸送は特定のインヒビターによっても影響されうる。これらのインヒビターは、CMP−シアル酸生合成に関与する酵素若しくはタンパク質の活性を遮断するシアル酸類似体、又はCMP−シアル酸の転移に必要ないずれの輸送体の活性を遮断するシアル酸類似体でありうる。これらのインヒビターは、酵素又はタンパク質の調節中心及び/又は触媒中心に結合して、基質の結合及び/又はその変換若しくは輸送を妨げうる。インヒビターは、シアル酸類似体、小有機分子、ペプチドリガンド、抗体及びそのフラグメント、DNAアプタマー、RNAアプタマー、スピーゲルマー(Spiegelmer)などを含む群から選ばれる高い結合アフィニティーを有する分子である。
【0044】
Nemod Biotherapeutics GmbH & Co. KG, Robert-Rossle-Str. 10, 13125 Berlin(ドイツ)により2003年8月14日付けでBraunschweig(ドイツ)の“DSMZ-Deutsche Sammlung von Mikroorganismen und Zellkulturen GmbH”にDSM ACC2606(NM−F9)及びDSM ACC2605(NM−D4)として寄託されているNM−F9及び/又はNM−D4細胞は、本発明の高活性糖タンパク質の発現のために及び本発明の後記方法における使用のために理想的に適合化された発現細胞系であることが判明している糖改変細胞である。なぜなら、NM−F9及びNM−D4細胞は、代謝的相補により復元されうるようにシアル酸付加能を喪失しているからである。したがって、本発明においては、NM−F9及び/又はNM−D4細胞は、本発明の高活性糖タンパク質及び後記の方法のための、シアル酸の糖ヌクレオチド生合成経路における欠損を有する最も好ましい細胞系である。
【0045】
NM−F9及びNM−D4細胞におけるシアル酸付加の細胞欠損はUDP−GlcNAc−2−エピメラーゼ活性の欠如により引き起こされ、それにより、シアル酸付加は、該エピメラーゼにより触媒される酵素反応の上流のシアル酸生合成の細胞産物での培地補足(代謝的相補とも称される)により復元されうる。そのようなシアル酸中間体はManNAc、ManNAc−6−P、NeuAc−9−P、Neu5Ac及びCMP−Neu5Acでありうる。ManNAcが好ましい。驚くべきことに、細胞シアル酸付加度は、細胞培養培地に加えるManNAcの量により制御することが可能であった。また、細胞シアル酸付加度だけでなく、より一層驚くべきことに、細胞から分泌又は遊離される糖タンパク質の度合も、そのように調節されうる。驚くべきことに、たいていは培地へのウシ胎仔血清(FCS)の添加によりもたらされる、細胞培養培地内のシアル酸付加血清タンパク質の存在は、細胞又は分泌性糖タンパク質のシアル酸付加度に影響を及ぼす。これは妥当だと思われる。なぜなら、該細胞は、CMP−シアル酸を産生するためのそれ自体の生合成経路を要することなく、ある程度は、de novoグリコシル化のために、取り込まれた糖タンパク質からのシアル酸を利用するようであり、あるいは異化経路によりシアル酸をシアル酸の公知又は未知前駆体生合成経路へと導入するためである。したがって、該細胞をFCS非含有培地内で又は更にはシアル酸付加タンパク質非含有培地内で培養する場合には、非常に低い度合のシアル酸付加が得られる。シアル酸付加度は常に、FCSの存在下では、FCSを含有しない同じ細胞培養条件下より高い。したがって、与えられた細胞系において最高のシアル酸付加度を達成するためには、FCS、又はより好ましくはシアル酸付加糖タンパク質、又は最も好ましくはシアル酸付加糖タンパク質フェツインを培地に加える。このようにして、NM−F9細胞又は分泌性糖タンパク質上のシアル酸付加度は、元のNM−wt細胞のシアル酸付加に比べて20%未満、好ましくは10%未満、最も好ましくはほぼ0%のシアル酸付加から、エピメラーゼ欠損を有さない元のNM−wt細胞シアル酸付加と比べてシアル酸付加を完了する(100%)シアル酸付加まで制御されうるであろう。
【0046】
この範囲のなかで、シアル酸付加度を示差的に制御することが可能であり、これは、一定の糖中間体、例えば0〜200mM、好ましくは0〜140mM、より好ましくは0〜90mMの濃度範囲のManNAcを培地に添加することにより行うことが可能である。非常に高いシアル酸付加度を達成するためには、シアロ糖タンパク質を培地に加えることが可能である。異なるシアル酸付加度を有する各糖タンパク質を単離し、異なるシアル酸付加度を有する同じ糖タンパク質とは別々に分析することが可能である。したがって、本明細書に記載のとおり、あるシアル酸付加度は、単一のグライコフォーム又は複数のグライコフォームの組成物を含みうるシアル酸付加形態に対応する。
【0047】
したがって、本発明は、後記で更に詳しく説明するとおり、糖タンパク質の示差的シアル酸付加のための方法又はプロセスを提供する。
【0048】
前記のとおり、異なるシアル酸付加度を有する各糖タンパク質の活性を分析することにより、シアル酸前駆体添加物の最適化濃度を決定することが可能である。分析対象の活性は、目的の糖タンパク質に左右される。例えば、糖タンパク質がサイトカインである場合には、免疫細胞(好ましくは、そのサイトカインにより刺激されることが公知の細胞、より好ましくは、該サイトカインの活性を測定するための標準的なアッセイにおいて使用される細胞)を刺激する該糖タンパク質の能力を分析することになろう。もう1つの例において、糖タンパク質が増殖因子である場合には、細胞増殖(好ましくは、その増殖因子により刺激されることが公知の細胞の細胞増殖、より好ましくは、該増殖因子の増殖活性を測定するための標準的なアッセイにおいて使用される細胞の増殖細胞)を誘導する該糖タンパク質の能力を分析することになろう。もう1つの例においては、血清半減期が対象となる。なぜなら、それは、治療用に適用される糖タンパク質の生物活性に著しく影響を及ぼすからである。本発明は、種々のシアル酸付加度を特徴とする1つの糖タンパク質の種々の形態の発現及び単離を可能にするものであり、該糖タンパク質の任意の活性を分析するために任意のアッセイを適用することにより、該糖タンパク質の活性のための最適なシアル酸付加度を決定することが可能である。
【0049】
以下においては、糖タンパク質及び高活性糖タンパク質を更に詳しく説明する。糖タンパク質の具体例は前記に挙げられている。前記糖タンパク質の多くは「サイトカイン」に属し、サイトカインは本明細書においては、免疫系の細胞において見出されるホルモンの一般的クラスを意味し、リンホカイン及びモノカインの両方並びにその他を含む。この定義は、局所的に作用し血中に循環せず、本発明に従い使用される場合に個体の免疫応答の改変を引き起こすホルモンを含む(これらに限定されるものではない)と意図される。他の適当な免疫調節性サイトカインの具体例には、限定的なものではないがインターフェロン(例えば、IFN−α、IFN−β及びIFN−γ)、インターロイキン(例えば、IL−1、IL−2、IL−3、IL−4、IL−5、IL−6、IL−7、IL−8、IL−9、IL−10及びIL−12)、腫瘍壊死因子(例えば、TNFα及びTNFβ)、エリスロポエチン(EPO)、FLT−3リガンド、マクロファージコロニー刺激因子(M−CSF)、顆粒球コロニー刺激因子(G−CSF)、顆粒球−マクロファージコロニー刺激因子(GM−CSF)、CD2及びICAMが含まれる。エリスロポエチンに関しては、この分子は前駆細胞を赤血球へ成熟させると考えられている。一方、トロンボポエチンは前駆細胞を血小板経路に沿って誘導すると考えられている。CSFは、骨髄内に見出される前駆細胞を成熟血液細胞の特定の型へ分化するように誘導するリンホイシンのファミリーを意味する。前駆細胞から生じる成熟血液細胞の個々の型は存在するCSFの型よって異なる。同様に、顆粒球−マクロファージコロニーの形成はGM−CSFの存在に左右される。また、IL−2、GM−CSF、TNF−αなどのヒト形態に対して実質的な相同性を有する他の哺乳動物のサイトカインは、免疫系に対して同様の活性を示すことが証明されている場合には、本発明において有用であろう。同様に、いずれかの特定のサイトカインに実質的に類似しているがタンパク質配列の比較的わずかな変化を有するタンパク質も本発明において有用であろう。タンパク質分子の機能的能力を妨げることなくタンパク質配列内のいくつかのわずかな変化が可能でありうることはよく知られており、したがって、本発明においてサイトカインとして機能するが現在公知の配列とは若干異なるタンパク質が製造されうる。最後に、本出願における「サイトカイン」なる語の単数形又は複数形の使用は決定的なものではなく、本発明及び特許請求の範囲の解釈を限定するものではない。接着若しくはアクセサリー分子又はそれらの組合せは単独で又はサイトカインと組合せて使用されうる。
【0050】
好ましい糖タンパク質は、グリコホリンA、EPO、G−CSF、GM−CSF、FSH、hCG、LH、インターフェロン、インターロイキン、抗体及び/又はそのフラグメントを含む群から選ばれる。
【0051】
本発明においては、高活性糖タンパク質は内因性タンパク質、好ましくは、発現細胞系において組換え手法により製造されるタンパク質でありうる。したがって、目的の糖タンパク質をコードするcDNA又は遺伝子を分子生物学的方法により発現ベクター内にクローニングし、発現細胞系内で発現させる。この発現は、好ましくは、安定発現である。すべてのその技術は当業者に公知である。多数の発現ベクターが、当業者に公知の哺乳動物細胞における発現に利用可能であり適している。好ましい実施形態においては、糖タンパク質は細胞により分泌される。したがって、発現ベクターは、糖タンパク質の分泌を促進するように構築される。分泌性糖タンパク質は可溶性糖タンパク質、又は膜貫通ドメインが欠失した膜タンパク質でありうる。好ましい実施形態においては、分泌シグナルペプチドをコードする核酸を、糖タンパク質をコードする核酸内に含ませ、これは、該タンパク質の分泌を促進及び/又は増強する。それらのシグナルペプチド及びそれらのコード核酸は当業者に公知である。本発明の好ましい実施形態においては、シグナルペプチドの核酸は配列番号1の配列又は該配列の突然変異、欠失、挿入及び伸長並びに該配列を含有する融合タンパク質を有する。
【0052】
本発明においては、NM−F9細胞内での2つの高活性糖タンパク質、すなわち可溶性タンパク質としての顆粒球マクロファージコロニー刺激因子(GM−CSF)及び膜タンパク質としてのグリコホリンA(GPA)の組換え発現が実施例として記載されている。
【0053】
GM−CSFは、先天性及び適応免疫系の活性化において重要な役割を果たす、骨髄細胞の強力な種特異的増殖因子である。該因子は顆粒球、マクロファージ、好酸球及びそれらの前駆細胞の増殖、発生、分化及び活性化を刺激する。さらに、GM−CSFは、体液性及び細胞性免疫応答にとって中心的な樹状前駆細胞の分化及び成熟に決定的に重要である。赤芽球及び巨核球前駆細胞の増殖におけるエリスロポエチン(EPO)との相乗作用、並びに種々の他のサイトカイン(例えば、IL−1、IL−3、IL−4、G−CSF)との相乗作用が観察される。GM−CSFは好中球、好酸球及びマクロファージの殺微生物活性、酸化的代謝、食作用活性及び細胞傷害性を増強し、好塩基球からのヒスタミン及びロイコトリエンC4の遊離を誘導する。骨髄移植、好中球減少症(特に化学療法及び/又は放射線療法後)並びに化学療法後の感染性疾患及び出血に関する血球減少症の患者を治療する一方で標準的な療法として適用されるヒトGM−CSFのいくつかの臨床的適応が存在する。さらに、それは回収のための幹細胞の末梢プールを増殖させ、急性骨髄球様白血病及び骨髄形成異常症候群を引き起こす白血病細胞の、化学療法中の細胞周期特異的薬に対する感受性を誘導する。組換えヒトGM−CSFは種々の疾患における造血系の再構成を支持する。また、いくつかの実験的療法が実施されており、例えば、照射の致死効果を無効にし造血を回復させる、癌治療におけるGM−CSF/放射線療法の併用、並びに細胞毒性薬(例えば、癌化学療法、HIV)に対する許容度を増強し、より高い投与量及び有意な罹病率減少を可能にするGM−CSF/化学療法の併用が挙げられる。rhGM−CSFは癌及び感染性疾患に対する細胞ワクチン及び癌ワクチンのための有望なアジュバントであり、腫瘍及び感染性疾患に対する、樹状細胞に基づくワクチンの重要成分である。それは養子T細胞移入にも関与している。一般には、rhGM−CSFは先天性免疫系を増強し刺激するであろう。
【0054】
本発明のヒトGM−CSFは、より高い特異的活性を有するため、先行技術により作製されたGM−CSFと比較して有利である。この増強された活性は、用量当たり患者当たりの、より少ない物質の使用、及び/又はより少ない投与、及び/又は望ましくない副作用(例えば、組換えGM−CSFの治療的使用中に観察されている毛細管漏出症候群)の軽減若しくは欠如、及び/又はより低い製造コストを可能にする。
【0055】
本発明においては、rhGM−CSFをUDP−GlcNAc−2−エピメラーゼ欠損細胞、例えばNM−F9において組換え発現させ、ほとんど存在しないシアル酸付加から最大限度のシアル酸付加までの種々のシアル酸付加度を有する種々の形態のrhGM−CSFを、漸増量のシアル酸代謝産物、好ましくはManNAc、例えば0〜90mMのManNAc、90mM ManNAcに加えて1及び3mg/mlのシアロ糖タンパク質フェツインの存在下での代謝的相補により製造した。rhGM−CSFの種々のシアロ形態の活性を分析するために、細胞増殖(好ましくは、TF−1細胞及びヒト単球樹状細胞系NemodDC(www.nemod.com))を刺激する能力を測定する標準的なアッセイを選択した。驚くべきことに、rhGM−CSFの細胞増殖刺激活性はrhGM−CSFのシアル酸付加度に強く左右され、高度にシアル酸付加されているが最大限度にはシアル酸付加されていないrhGM−CSFの1つのシアロ形態が最高活性を示す。商業的に入手可能なrhGM−CSF、好ましくは、細菌及び酵母において発現されるLeukomax(登録商標)及びLeukine(登録商標)の細胞増殖活性を同じアッセイで試験したところ、NM−F9細胞内で発現される最も活性なシアロ形態より低いことが判明した。Leukomax(登録商標)又はLeukine(登録商標)の最高活性を達成するためには、対応in vitro試験において、500倍未満までの、NM−F9細胞からの最適シアロ形態が必要な可能性がある。Leukine(登録商標)、Leukomax(登録商標)、又はrhGM−CSFの最適シアロ形態の同一濃度で活性を分析したところ、活性の増加は1.5倍まで、好ましくは2倍まで、より好ましくは3倍まで、最も好ましくは4倍まで、非常に好ましくは5倍までであった。先行技術は、rhGM−CSFの活性がそのグリコシル化に左右されることは示していなかったため、これらの知見は特に驚くべきものである。
【0056】
rhGM−CSFの最適シアロ形態で見出される、より高い活性に加えて、そのシアロ形態の比較的高いシアル酸付加度は、治療適用における、より長い血清半減期を保証する。有利には、より長い時間間隔の、より低い用量の投与は、免疫原性及び副作用の軽減又は完全な消失を伴う。後者の2つは更に、ヒト関連グリコシル化パターンにより正の様態で影響される。高活性rhGM−CSFの高いシアル酸付加度は薬物動態学的特性を大きく改善する。
【0057】
本発明は、種々の臨床用途だけでなくin vitro用途においても(例えば、分化樹状細胞及び活性化T細胞の作製のために)前記のとおりに有利に使用されうる高活性糖タンパク質分泌性rhGM−CSFを提供する。
【0058】
本発明の新規技術は、他の増殖因子及びホルモン、例えばEPO、FSH及びhCGの改善のために、並びに種々の他の糖タンパク質、例えばサイトカイン及び抗体(前記のとおり)、及び本明細書に記載のその他のものの改善のために非常に有益である。
【0059】
もう1つの例として、アシアログリコホリンA(AGPA)をNM−F9細胞内で組換え発現させた。血液サンプルから単離し次いで酵素により脱シアル酸してトムセン−フリーデンライヒ(Thomsen-Friedenreich)(TF、コア−1)抗原を露出させたグリコホリンA(GPA)に基づき、TF陽性腫瘍の治療のためのワクチンの開発に成功した。
【0060】
GPA及びAGPAは内在性膜タンパク質である。製造方法の確立のためには、任意の細胞系により、好ましくはNM−F9及びNM−D4細胞により細胞培養培地内に分泌されうるGPAの組換え断片を発現させることが非常に有利であろう。したがって、GPAの細胞外グリコシル化ドメインを含有しGPAの膜貫通ドメインと細胞内ドメインとを欠くGPA断片をコードする発現ベクターsecGPAを構築した。最初の実験では、安定にトランスフェクトされたNM−F9細胞の培地においては分泌性AGPAは少量しか検出され得ないことが示された(実施例を参照されたい)。分泌率を上昇させるためには、いくつかの異種シグナルペプチドをGPA断片に融合させて、内因性シグナルペプチドを除去した。驚くべきことに、配列番号1により特徴付けられるアミノ酸配列を含むGM−CSF由来のシグナルペプチドは、GPAの内因性シグナルペプチド並びにT細胞受容体及び抗体κ軽鎖由来のその他の異種シグナルペプチドと比べて最も有効なシグナルペプチドであった。後者の2つのシグナルペプチドのみが分泌性タンパク質の組換え発現における使用に関して利用可能であり当業者に公知である。
【0061】
本発明は、シアル酸付加を欠損した任意の細胞又は細胞系により発現又は産生される高活性糖タンパク質分泌性アシアログリコホリンAを提供し、好ましくは、該細胞はヒト細胞であり、より好ましくは該細胞はNM−F9又はNM−D4細胞であり、それは、シアル酸付加による遮蔽を伴うことなく好ましい密度でTF抗原を含有する。マウスモデルにおいて抗腫瘍ワクチンとして使用される場合には、それは強力な抗TF及び抗腫瘍応答を示し、それはヒトin vitro研究において強力なTF特異的抗腫瘍応答をも示し、これはGPAのシアル酸付加又は他のグリコシル化形態によっては達成されず、このことは、これが本発明における高活性糖タンパク質形態であることを示している。その強力な抗腫瘍応答は、それがTF特異的腫瘍に対する強力な抗腫瘍ワクチン成分として治療又は予防場面において有効に使用されうることを示している。
【0062】
TF糖鎖は、多数の腫瘍組織上には存在するが健康なヒトの正常細胞上には存在しない周知の腫瘍抗原である。グリコホリンA(GPA)は、シアル酸付加により遮蔽されたTF抗原の担体である。血液サンプルから単離し次いで酵素により脱シアル酸してTF抗原を露出させたGPAに基づき、TF陽性腫瘍患者の治療のためのワクチンの開発に成功した。本発明においては、GPAの唯一のグリコシル化部分でもある細胞外ドメインのみを、シアル酸付加を欠損するヒト細胞(好ましくはNM−F9及びNM−D4細胞)内で分泌形態(secGPA、前記を参照されたい)として組換え産生させる。シアル酸付加欠損細胞、例えばNM−F9又はNM−D4細胞を使用する利点は、その細胞内で任意の形態で発現されるGPA上に存在するTF抗原がシアル酸を含有せず、したがって酵素前処理無しで、規定の標準化されうるワクチンとして使用されうることである。
【0063】
本発明はまた、糖タンパク質をコードする核酸配列並びに配列番号1の核酸配列又は該配列の突然変異、欠失、挿入及び伸長並びに該配列を含有する融合タンパク質を含む核酸を提供する。配列番号1又は該配列の突然変異、欠失、挿入及び伸長並びに該配列を含有する融合タンパク質により特徴付けられるGM−CSF由来のシグナルペプチドは、哺乳動物細胞、好ましくはヒト細胞、より好ましくはNM−wt細胞のサブクローン、最も好ましくはNM−F9及びNM−D4細胞内での任意の哺乳動物遺伝子産物(例えば、GPA)の分泌性発現を駆動するために使用されうる。該シグナルペプチドは、当業者に公知の技術、例えば、目的タンパク質をコードするcDNA内へ該コード領域のフレームと合わせて、シグナルペプチドをコードするヌクレオチド配列を分子クローニングすることにより、目的タンパク質内に組込まれる。それにより、該内因性シグナルペプチドは完全又は部分的に除去される。この構築物を、当業者に公知の任意の発現ベクター内にサブクローニングし、当業者に公知の任意の方法、例えば化学的トランスフェクション、リポフェクション、エレクトロポレーション又は感染により細胞内に導入すると、シグナルペプチドと目的タンパク質とを含む細胞により融合タンパク質が合成される。該細胞内でのこの融合タンパク質のプロセシング中に、該シグナルペプチドは完全又は部分的に除去される。
【0064】
本発明においては、「糖タンパク質」なる語は、ウイルス、ウイルス粒子又はウイルスタンパク質としても定義される。本発明に従い製造されうるウイルス、ウイルス粒子又はウイルスタンパク質には、腸内ウイルス、例えばライノウイルス、ポリオウイルス又はアフタウイルス;ヘルペスウイルス、例えば単純ヘルペスウイルス、仮性狂犬病ウイルス又はウシヘルペスウイルス;オルトミクソウイルス、例えばインフルエンザウイルス;ペスティウイルス;ラブドウイルス;パラミクソウイルス、例えばニューカッスル病ウイルス、呼吸器合胞体ウイルス、ムンプスウイルス又は麻疹ウイルス;レトロウイルス、例えばヒト免疫不全ウイルス、パルボウイルス又はパポバウイルス;ロタウイルス;コロナウイルス;フラビウイルス、例えば黄熱ウイルス又はダニ媒介脳炎ウイルス;肝炎ウイルス、例えばA型肝炎又はB型肝炎ウイルスに由来するものが含まれる。
【0065】
本発明における高活性ウイルス、ウイルス粒子又はウイルスタンパク質は、適当なバイオアッセイ(例えば、ウイルス、ウイルス粒子又はウイルスタンパク質の個々の発現シアル酸付加形態の感染性、ワクチン又はワクチン成分としてのウイルス、ウイルス粒子又はウイルスタンパク質の個々の発現シアル酸付加形態の効力、ウイルス、ウイルス粒子又はウイルスタンパク質の個々の発現シアル酸付加形態の生産性、あるいは当業者に公知の他の有益な医学的又は生物学的特徴の測定)により測定されうるという利点を有する。対応バイオアッセイは、本発明を使用することにより当業者により選択され採用され構築されうる。好ましくは、産生されたウイルスの感染性を、改善されたワクチンが得られるよう示差的シアル酸付加により最適化する。これは該ウイルス、ウイルス粒子又はウイルスタンパク質のシアル酸付加度の最適化を可能にする。
【0066】
好ましくは、ワクチンにおいて使用するための高活性ヒトウイルス、ウイルス粒子又はウイルスタンパク質を産生するヒト細胞系NM−F9又はNM−D4を使用する。該ヒト細胞系は、正常な翻訳後及び翻訳周辺(peri-translational)修飾並びに集合(例えば、グリコシル化)の能力を有し、本明細書に記載されているとおりのシアル酸付加及びシアル酸付加度の最適化を可能にする。
【0067】
ウイルスのシアル酸付加は、しばしば、ウイルスの感染性において重要な役割を果たすことが公知である。シアル酸付加度を最適化することにより、より安全な及び/又はより強力なウイルスワクチンが製造されうる。また、この及び/又はより高い生産性は、改善された、したがってより経済的な製造方法につながりうる。
【0068】
本発明はまた、シアル酸の糖ヌクレオチド生合成経路における少なくとも1つの欠損を含有し、かつ糖タンパク質をコードする核酸でトランスフェクトされた発現細胞系において、ある濃度の少なくとも1つのシアル酸前駆体添加物を添加した培地内で高活性糖タンパク質を発現させることを含む、高活性糖タンパク質の製造方法であって、
該濃度が、
(i)種々の濃度の少なくとも1つのシアル酸前駆体を使用する示差的シアル酸付加により、複数の異なるシアル酸付加形態の糖タンパク質を発現させること、
及び
(ii)適当なバイオアッセイにおいて、参照糖タンパク質と比較して、異なるシアル酸付加形態の活性を測定すること、
及び
(iii)より高い/最高の活性を有するシアル酸付加形態を選択し、該糖タンパク質のより高い/最高の活性レベルと相関する、該シアル酸前駆体添加物の濃度を決定すること
を含む方法により決定される、高活性糖タンパク質の製造方法を提供する。
【0069】
前記方法は、高活性糖タンパク質と決定された糖タンパク質の最適シアル酸付加形態の製造を可能にする。当業者にとっては、本発明の方法を使用することにより、分析的又は予備試験的規模で決定された条件を製造規模へ移すこと、並びに増殖及び培地条件を調節し改善することが可能かつ比較的容易である。他の方法に対する本方法のさらなる利点は、生産性の改善につながりうる規模拡大操作中に、シアル酸前駆体添加物の濃度及び組成の適合化を他の培地成分の最適化に加えて行うことにより、高活性タンパク質の所望のシアル酸付加度を安定化しうることである。
【0070】
本発明における高活性糖タンパク質に関する前記のすべての定義及び説明は、前記及び後記の方法にも有効である。
【0071】
さらに、本発明は、高活性糖タンパク質の同定/決定方法であって、
i)シアル酸の糖ヌクレオチド生合成経路における少なくとも1つの欠損を含有する発現細胞系を、該糖タンパク質をコードする核酸でトランスフェクトすること、
及び
ii)種々の濃度の少なくとも1つのシアル酸前駆体添加物を含有する培地を使用する示差的シアル酸付加により、複数の異なるシアル酸付加形態の糖タンパク質を発現させること、
及び
iii)適当なバイオアッセイにおいて、参照糖タンパク質と比較して、前記の異なるシアル酸付加形態の活性を測定すること、
及び
iv)より高い/最高の活性を有するシアル酸付加形態を選択すること
を含む方法を提供する。
【0072】
原則として、上記方法は、(i)所定の糖タンパク質の活性とそのシアル酸付加度との間に何らかの相関が存在するかどうかを分析するための分析手段、(ii)所定の糖タンパク質のその活性に関する最適シアル酸付加形態が何であるかを決定するための分析手段、及び(iii)最適シアル酸付加形態を与える条件を決定するための分析手段として使用されうる。
【0073】
さらに、本発明は、ヒトグリコシル化、好ましくはシアル酸付加の、生物活性における役割を調べるのに有用である。
【0074】
また、糖タンパク質の示差的シアル酸付加のための方法であって、シアル酸の糖ヌクレオチド生合成経路における少なくとも1つの欠損を含有する発現細胞系を、該糖タンパク質をコードする核酸で形質転換し、複数の異なる濃度の少なくとも1つのシアル酸前駆体添加物を添加した培地内で該細胞を培養し、複数の異なるシアル酸付加形態の糖タンパク質を得ることを特徴とする方法を提供する。
【0075】
本発明はまた、高活性ウイルス、ウイルス粒子又はウイルスタンパク質の同定/決定のための方法、その製造及びその示差的シアル酸付加、並びに本発明により提供される方法から得られる高活性ウイルス、ウイルス粒子又はウイルスタンパク質を提供する。
【0076】
本発明における高活性糖タンパク質に関する前記のすべての定義及び説明は、前記及び後記の方法にも有効である。
【0077】
本発明は、好ましくは、グライコプロファイリング(GlycoProfiling)、グライコエンジニアリング(GlycoEngineering)、グライコアナリティックス(GlycoAnalytics)及びグライコエクスプレス(GlycoExpress)に基づく技術基盤に組み込まれる。
【0078】
・グライコプロファイリング(GlycoProfiling)は、グライコエンジニアリング(GlycoEngineering)のための適当な細胞を選択するために、細胞グリコシル化機構及び該グリコシル化機構により産生された糖(優先的には、細胞表面上に存在するもの)を分析するための或る範囲の技術であると理解される。
【0079】
・グライコエンジニアリング(GlycoEngineering)は、修飾されたグライコプロファイル(GlycoProfile)を有する細胞を与える、細胞のグリコシル化機構を修飾するための或る範囲の技術であると理解される。
【0080】
・グライコアナリティックス(GlycoAnalytics)は、タンパク質のグリコシル化を分子レベルで分析するための或る範囲の技術であると理解される。
【0081】
・グライコエクスプレス(GlycoExpress)は、示差的シアル酸付加、高活性タンパク質の同定及び決定並びに高活性糖タンパク質の製造を含む、本発明の後記の技術及び方法を含む。
【0082】
本発明の糖タンパク質及び方法を含む本明細書に挙げた技術を組合せる利点は、より改善された高活性タンパク質、本発明の意味における、より良好な発現細胞株系、それらの分析及び製造を可能にする。他の利点としては、該方法及び産物の速さ、コスト、安定性及び有効性が挙げられる。
【0083】
前記のすべての定義及び説明は、前記及び後記の糖タンパク質及び方法にも有効である。
【0084】
本発明のもう1つの利点は、糖タンパク質の使用及び機能に関して最適化されたシアル酸付加を伴う高活性糖タンパク質を製造するための、統合された系(統合系)に類似したその方法の組合せである。そのような組合せを以下の好ましい具体例において説明するが、該具体例はこれを限定するものではない。
【0085】
第1ステップにおいては、糖タンパク質、好ましくは分泌性形態の糖タンパク質、好ましくは配列番号1の配列を含む糖タンパク質、より一層好ましくはヒトGM−CSF又はGPAをコードする核酸を、好ましくはエレクトロポレーションにより、シアル酸の糖ヌクレオチド生合成経路における欠損を有する発現細胞系内に好ましくはNM−F9又はNM−D4を使用してトランスフェクトし、好ましくは単一細胞クローニングにより、安定発現クローン(好ましくは無血清培地内、より一層好ましくはタンパク質非含有培地内で増殖するもの)を選択する。
【0086】
第2ステップにおいては、種々の濃度の少なくとも1つのシアル酸前駆体を使用することにより、異なるシアル酸付加形態(好ましくは、一連の少なくとも2、より好ましくは3以上、より好ましくは4以上、より一層好ましくは5以上の異なるシアル酸付加形態)の糖タンパク質を示差的シアル酸付加により製造する。少なくとも1つのバイオアッセイ、例えばヒトGM−CSFに関する細胞増殖in vitroアッセイ、又はGPAに関するある腫瘍細胞に対するより高い特異的免疫原性を示すためのマウス若しくはヒトを使用するin vivo免疫試験により、より高い活性のグライコフォームを決定する。好ましい形態においては、糖タンパク質の用途に応じた最高活性を有する高活性糖タンパク質を決定するために、バイオアッセイの組合せを使用する。本明細書に記載のとおり、選択する高活性糖タンパク質は、すべてのバイオアッセイにおいてより良好である必要はなく、1つのバイオアッセイにおけるより高い活性又は改善された特徴で十分でありうる。しかし、好ましくは、選択する高活性糖タンパク質は、少なくとも2つのバイオアッセイ、例えば1つのin vitroモデル、例えば増殖アッセイにおいて、より良好であり、及び/又は少なくとも1つのin vivoモデルにおいて、より高い活性、及び/又はより高い安定性、及び/又はより長い血清半減期、及び/又はより長いバイオアベイラビリティ、及び/又は免疫原性の改善、及び/又は抗原性の改善を示す。最も好ましいのは、種々の特徴(例えば、受容体媒介機能におけるより高い活性、及びより長い血清半減期を示すことによる改善されたバイオアベイラビリティ)の組合せによるものでありうる、より高いin vivo生物活性をヒトにおいて有するものである。例えば、記載されている条件下の培地内でNM−F9及びシアル酸前駆体添加物としての90mM ManNAcを使用して得られるrhGM−CSFシアル酸付加形態が最高の活性を示したため、これが好ましい高活性糖タンパク質である。
【0087】
もう1つのバイオアッセイにおける更なる試験のために、1つのバイオアッセイにより、より大きなセットから、より小さな群の糖タンパク質シアル酸付加形態を選択することが、速さ及び手段に関しては有利でありうる。GPAの場合には、唯一のシアル酸付加形態、すなわち、最低シアル酸付加形態を作製するので十分であり、したがってこれを無血清培地内でシアル酸前駆体添加物無しで行うので十分である(前記及び後記も参照されたい)。この場合、得られた高活性GPAを、遥かに高い活性(この場合には、動物モデル及びヒトにおいて強力な抗TF免疫応答をもたらす、より高い免疫原性)を示すヒト由来のシアル酸付加GPAと比較することが可能である。しかし、好ましくは、少なくとももう1つのシアル酸付加形態を本発明の方法により作製し、該バイオアッセイにおいて比較する。
【0088】
上記統合系により、高活性糖タンパク質の前記決定の後、該高活性糖タンパク質を製造するためにシアル酸前駆体添加物の対応濃度を使用することにより、糖タンパク質をコードするトランスフェクトされた核酸を含有する同じ発現細胞系を使用することが可能となる。高活性糖タンパク質の製造では、しばしば、製造規模の拡大が望まれる。このためには、シアル酸前駆体添加物の最適条件で十分であるが、所望のシアル酸付加度の高活性タンパク質を安定化するために、例えば他の培地成分を最適化し、異なる反応容器を使用することにより、該シアル酸前駆体添加物の濃度及び組成を調節することが有利又は更には必要かもしれない。これは、本発明を使用することにより当業者により達成されうる。シアル酸前駆体添加物を調節することによりシアル酸付加度を安定化しうることは、先行技術の通常の発現系によっては達成されない該方法の大きな利点である。
【0089】
高活性糖タンパク質の同定及び製造のために実質的に同じ又は類似した方法を使用することを可能にする統合系は、とりわけ、より高い速度及びより低いコストという明らかな利点を有する。
【0090】
上記統合系は、例えばグライコプロファイリング(GlycoProfiling)、グライコエンジニアリング(GlycoEngineering)及びグライコアナリティックス(GlycoAnalytics)として本明細書に記載されている他の製造方法、技術及び方法と組合された場合に特に有利かつ有効である。
【0091】
シアル酸の糖ヌクレオチド生合成経路における復元可能な欠損を有する発現細胞系を作製することは非常に困難である。なぜなら、(i)多数の酵素がシアル酸付加に関与しており、このことは、単一の酵素活性の除去が必ずしも細胞のシアル酸付加能に影響を及ぼさないことを意味し、(ii)該酵素の多くは重複しており、このことは、1つの酵素活性の喪失が、しばしば、他の酵素により補償されることを意味し、(iii)該欠損の復元は、しばしば、可能ではなく、(iv)該欠損の代謝的相補は、しばしば、可能ではないからである。本発明は、好ましくは少なくとも2つの酵素によりシアル酸付加されうる脱シアル酸構造に結合する認識分子で初代細胞又は細胞系から発現細胞系を選択することを含む、シアル酸の糖ヌクレオチド生合成経路における欠損を有する発現細胞系の新規製造方法を提供する。
【0092】
好ましい実施形態においては、選択前に初代細胞又は細胞系からの細胞を突然変異誘発する。突然変異誘発は、化学的若しくは物理的手段によるランダム突然変異誘発又は本明細書に記載の若しくは当業者に公知の部位特異的突然変異誘発により行うことが可能である。
【0093】
本発明の方法とは対照的に、当業者に公知の現在の方法では、特に標的遺伝子が既知でない場合には、シアル酸前駆体経路における遺伝的欠損を有する細胞を作製することは非常に困難であり、時間を浪費し、しばしば可能でない。
【0094】
上記の選択は、当業者に公知の技術(例えば、アフィニティークロマトグラフィー、例えば、磁気細胞選別技術(MACS技術)若しくは免疫沈降若しくはクロマトグラフィー、及び/又はフローサイトメトリーによる細胞選別、及び/又は例えばフローサイトメトリー若しくは免疫細胞化学による少なくとも1つの認識分子への結合特性による細胞の同定、又は分泌性タンパク質の生化学的分析)により、記載されている認識分子の少なくとも1つを使用することによる、該細胞の富化及び/又は単離により行う。
【0095】
選択された細胞を発現細胞系として使用することが可能であり、好ましくは、純粋なクローンを作製するために、公知技術により、例えば単一細胞クローニングにより、更にクローニングする。
【0096】
驚くべきことに、少なくとも2つの酵素によりシアル酸付加されうる脱シアル酸構造を標的化する認識分子の使用は、代謝物質により復元されうる(発現細胞系におけるシアル酸付加度の代謝制御のための前提条件)それらの酵素活性における欠損を有する細胞の選択を可能にし及び/又は該細胞に対して選択を強く偏らせる。
【0097】
本発明の方法はそのような細胞系の作製を可能にし、あるいは速さ、労力の軽減及び/又は成功確率の点でそれらの作製を著しく促進する。これは特にヒト細胞系に当てはまることであるが、ヒト細胞系だけに当てはまるものではない。
【0098】
本発明の方法は、突然変異誘発のための標的遺伝子が既知でない場合に特に適しており、選択はシアル酸付加のダウンレギュレーションの表現型効果に基づいたものである。しかし、それは、部位特異的突然変異誘発を用いる場合、例えばシアル酸付加の表現型ダウンレギュレーションにより安定突然変異誘発クローンの選択を著しく促進することによる、部位特異的組換えによるGlcNAc−エピメラーゼ活性のノックアウトを用いる場合にも有利である。
【0099】
これらの作製される細胞系は、示差的シアル酸付加の目的のために又は本発明における発現細胞系として製造されうるが、それらは、有益又は使用可能である場合には他の目的にも使用されうる。
【0100】
シアル酸付加する能力を欠損した細胞系(その欠損を復元する選択肢を伴っているもの)を作製するためには、原則として、すべての哺乳動物細胞系が適用可能である。製造プロセス(製造方法)には、バイオテクノロジーに適した細胞系、例えばPer.C6、HEK293、K562、CV1、COS−7、ハイブリドーマ細胞、ナマルワ(Namalwa)、BHK及びCHOが好ましい。該欠損を有する新規細胞系を作製するための選抜きの方法は、本発明者らのグライコエンジニアリング(GlycoEngineering)技術に属する前記方法である。
【0101】
上記欠損を有する細胞系を作製するために前記方法を開始する前に、候補細胞系のグリコシル化機構を少なくとも部分的に特徴決定すること(グライコプロファイリング(GlycoProfiling))が非常に有用である。このためには、(i)グリコシル化機構、例えばシアリルトランスフェラーゼ、本明細書中に前記で説明されているシアリルトランスフェラーゼ、トランスポーター(輸送体)、エピメラーゼ、シアリダーゼなどの基質である糖鎖構造に重要な酵素のトランスフェラーゼの関連遺伝子のmRNA発現、(ii)関連酵素、好ましくはシアリルトランスフェラーゼの酵素活性、(iii)シアル酸付加されうる又はシアル酸付加される糖鎖決定基の適当な認識分子による決定、を分析しうる技術の少なくとも1つ(好ましくは、そのような技術の組合せ)を用いるのが好ましい。グライコプロファイリング(GlycoProfiling)は、グリコシル化機構の修飾のための方法(グライコエンジニアリング(GlycoEngineering))に関する決定を可能にする候補細胞系のグリコシル化機構を理解することを助ける。例えば、候補ヒト細胞系、例えばZR75−1、HEK293、NM−wt(K562)、KG1、LS174T、MCF−7、SW480及びT47Dの、グリコシル化に関与する酵素及びタンパク質(例えば、グリコシルトランスフェラーゼ、単糖生合成の酵素及びトランスポーター)のmRNA発現プロフィールをRT−PCRにより分析した。候補細胞系の酵素活性、好ましくはシアリルトランスフェラーゼ活性を分析するためには、バイオアッセイを用いた。バイオアッセイは、糖鎖決定基、好ましくは、少なくとも2つの酵素によりシアル酸付加される又はされうる糖鎖決定基を、適当な方法、例えばフローサイトメトリー及び/又は免疫蛍光顕微鏡検査(好ましくはシアリダーゼと共に、特異的抗体及びレクチンを使用するもの)又は、シアル酸を除去若しくは破壊するための他の方法、例えば穏やかな過ヨウ素酸塩酸化により分析するためにも用いられる。適当なバイオアッセイは公知であり、あるいは当業者により調節又は開発されうる。
【0102】
本発明においては、「初代細胞又は細胞系」なる語は、例えば生物学的試料に由来するもの、例えば血液細胞、組織などからの細胞及び既存の細胞系を含むすべての細胞を包含する。細胞及び細胞系は、公然の(declared)細胞系で時々見られる、単一細胞又は細胞混合物に由来する、特徴決定されたクローンよりなることが可能である。該細胞は既に不死化されていることが可能であり、あるいは本発明の方法の拡張部分として不死化されうる。該不死化は例えば腫瘍由来の細胞の場合のように自発的であってもよいし、あるいは当業者に公知の技術、例えばウイルス形質転換により不死化されてもよい。それにより、当業者は、個々の細胞によって異なりうる、不死化の最良の時点を決定することが可能である。また、該細胞は、他の遺伝子により又は他の遺伝子において、改変されていてもよいし又は改変された状態であってもよい。
【0103】
本発明において、「シアル酸の糖ヌクレオチド生合成経路における欠損」なる表現は、本明細書に記載されているものと同じ欠損を意味する。
【0104】
本発明において、「認識分子」なる語は、部分的に又は好ましくは完全に脱シアル酸されており少なくとも2つの酵素によりシアル酸付加されうる分子に好ましくは及び/又は特異的に結合する分子を意味する。好ましい分子はレクチンであり、より一層好ましいのは、対応する糖鎖構造を認識する抗体である。より一層好ましいのは、TF(トムセン−フリーデンライヒ(Thomsen-Friedenreich)、コア−1)(例えば、Nemod−TF1、Nemod−TF2、A78−G/A7)に結合する抗体、又は末端ガラクトース残基を認識する抗体、又はレクチン、例えばRCA、ECL、PNA、Jacalin、ACA、BPL若しくはAmaranthin、又はシアル酸に結合するもの、例えばSNA及びMAA、又はPankoMabである。
【0105】
より好ましい実施形態においては、該認識分子が結合する脱シアル酸構造はα2−3及びα2−6結合シアル酸によりシアル酸付加されうる。
【0106】
より一層好ましい実施形態においては、該認識分子が結合する構造はO−グリカンである。
【0107】
より一層好ましい実施形態においては、該認識分子が結合する構造は、トムセン−フリーデンライヒ(Thomsen-Friedenreich)構造とも称されるコア−1構造(GalNAcα1−3Gal)である。
【0108】
より一層好ましい実施形態においては、シアル酸の糖ヌクレオチド生合成経路における欠損を有する発現細胞系の作製のための方法は、コア−1構造を認識する抗体で初代細胞又は細胞系から発現細胞系を選択することを含む。
【0109】
より一層好ましい実施形態においては、選択前に該初代細胞又は細胞系に突然変異誘発を行う。
【0110】
一例としては、シアル酸の糖ヌクレオチド生合成経路における欠損を含有する本発明の発現細胞系は、NM−wt細胞を改変することにより作製した。グライコプロファイリング(GlycoProfiling)(前記を参照されたい)により示された多数の候補細胞系のグリコシル化プロフィールに基づき、TF陽性細胞を作製するためにグライコエンジニアリング(GlycoEngineering)のための細胞系を選択した。好ましくは、選択される細胞はC2GNT−L又はMに対して陰性であり、より好ましくは、潜在(cryptic)TFに対して陽性であり、より一層好ましくは、α2,3及びα2,6結合シアル酸により非常に高い度合でシアル酸付加されることが可能であり、最も好ましくは、該細胞はNM−wt細胞である。簡潔に説明すると、該細胞を、当業者に利用可能な任意の方法により、好ましくはフローサイトメトリー又は免疫細胞化学法(実施例に記載されている)により、TF発現に関して分析する。TF陽性細胞は、モノクローナル抗体A78−G/A7又はPankoMabを使用することにより選択される。TF陽性細胞の数が少なすぎる場合には、TF陰性細胞系を突然変異原、好ましくは化学的突然変異原、好ましくはメタンスルホン酸エチル(EMS)で処理する。ついで、前記のとおりにTF陽性細胞を選択する。TF陽性細胞に関する選択は反復する必要があり、ついでTF陽性細胞は、安定にTF陽性である細胞を受容するようクローニングする必要がある。そのようにして、NM−F9及びNM−D4細胞を作製した。
【0111】
同様の適合化された手法により、例えば、特徴決定されたグリコシル化プロフィールを有する糖タンパク質の製造における使用のために、新規グライコプロフィールで、新規細胞系を受容するよう任意の細胞系を糖改変操作することが可能である。そのようなプロフィールは、改善されたシアル酸付加、よりヒト的なグリコシル化、天然糖タンパク質に類似したグリコシル化、又はより低い若しくはより高い免疫原性のグリコシル化でありうる。
【0112】
以下においては、糖タンパク質及び高活性糖タンパク質の医薬組成物又は組成物並びにそれらの用途を更に詳しく説明する。本発明は、本発明の糖タンパク質と希釈剤又は担体とを含む、in vivo又はin vitroでの使用のための本発明の高活性糖タンパク質の組成物又は医薬組成物に関する。本明細書に記載の哺乳動物糖タンパク質及びその組成物は、該糖タンパク質の性質によって定められる広範な用途において使用されうる。これらのすべての用途において、高活性糖タンパク質の使用は常に非常に有益であろう。例えば、本発明の哺乳動物糖タンパク質は、in vitroアッセイ、例えば任意の種類の細胞アッセイ若しくは免疫学的アッセイ、又は分析アッセイ、例えばELISA若しくはRIAにおいて使用されうる。本発明において好ましい糖タンパク質の一例としてGM−CSFを使用する場合には、この増殖因子を、特定の免疫細胞、例えば樹状細胞又はマクロファージの増殖及び/又は分化を誘導するためにin vitroで使用することが可能であり、ついでそれらを、特定の免疫学的アッセイ、例えばELISPOTアッセイ、T細胞アッセイ、細胞傷害性アッセイ、細胞泳動アッセイ、細胞接着アッセイ又は食作用アッセイにおいて使用する。本発明のGM−CSFを細胞増殖アッセイにおいて直接的に使用することも可能である。
【0113】
本明細書に記載の高活性哺乳動物糖タンパク質及びその組成物は診断目的にも有用である。例えば、糖タンパク質の最適シアル酸付加形態を診断目的のための標準として製造することが可能であろう。好ましいのは、本明細書に記載の高活性哺乳動物糖タンパク質及びその組成物のin vivo用途である。本発明の高活性哺乳動物糖タンパク質の広範なin vivo用途、例えば任意の種類の感染性疾患、癌、白血病、造血系の疾患、好中球減少症、血球減少症、骨髄異形成症候群及び自己免疫疾患の予防的及び/又は治療的処置は、具体的な糖タンパク質の性質によって定められ、当業者に公知である。該糖タンパク質がin vivo用途のために適用される哺乳動物種は、該糖タンパク質の起源によって決定される。例えば、ウシGHは、乳生産を増加させるためにウシの治療に使用される。本発明の高活性ウシGHの適用はその治療を改善するであろう。なぜなら、適用の回数及び望ましくない副作用は減少すると予想されるからである。
【0114】
ヒトにおいては、本明細書に記載の糖タンパク質を治療用に使用する。高活性糖タンパク質は常に非常に有益であろう。なぜなら、投与の回数及び望ましくない副作用が軽減されると予想されるからである。患者への投与の場合には、本発明の精製された糖タンパク質を、通常の方法に従い、薬学的に許容される担体又は希釈剤と混合する。治療用製剤は、静脈内注入若しくは皮下注射により、又は当技術分野において公知の任意の他の許容される方法により投与される。該製剤は、所望により、他の治療薬をも含有しうる。投与の量、頻度及び期間は様々であり、使用する特定の化合物の活性、年齢、体重、全身健康状態、性別、投与の食事時間、投与経路、排泄速度、薬物の組合せ及び疾患特異的療法の重篤度を含む種々の要因によって異なるであろう。単一剤形を与える、担体物質と組合される有効成分の量は、治療対象の宿主及び個々の投与方式によって様々である。
【0115】
本発明の好ましい実施形態においては、該医薬組成物はワクチン又はワクチン−アジュバントである。本発明においては、ワクチン組成物なる語は、哺乳動物及びヒトにおいてワクチンとして使用されうる任意の組成物に関するものである。ワクチンは、免疫応答を誘導する治療用又は予防用の医薬組成物を意味する。本発明のワクチン組成物の製造方法の形態は特に限定されるものではなく、所望の形態の組成物は、好ましくは、当技術分野において利用可能な単一の方法、又は適当に組合された方法を適用することにより製造されうる。任意のワクチン及び/又はワクチン−アジュバントの投与は皮下、皮内、静脈内、非経口、筋肉内又は任意の他の許容される方法でありうる。前記剤形により予想される製剤及び医薬組成物は、通常の技術を用いて、通常の薬学的に許容される賦形剤及び添加剤を使用して製造されうる。他のアジュバントは該ワクチン又は該糖タンパク質と共に投与されうる。
【0116】
ヒト腫瘍ワクチンの一例はアシアログリコホリンA(AGPA)である。グリコホリンA(GPA)は、赤血球の膜に存在し幾つかの細胞系にも見出されるシアル酸付加糖タンパク質である。ワクチンの製造のためには、それをヒト血液製剤(ヒト血液調製物)から単離し、酵素により処理して、GPAに結合したすべてのシアル酸を除去する。得られるAGPAは腫瘍特異的糖鎖抗原トムセン−フリーデンライヒ(Thomsen-Friedenreich)(TF)のキャリアであり、したがって、TFに対する持続性免疫反応を誘導し従って患者のTF陽性腫瘍負荷を排除又は少なくとも軽減するためのワクチンとして適用される。本明細書に記載のシアル酸付加欠損細胞の使用は本発明により、酵素処理を要することなく、かつ、製造用にヒト血液製剤を使用することに限定されることなく、AGPAの組換え製造を可能にする。AGPAは、腫瘍及び癌患者の免疫療法のためのワクチンとしての糖タンパク質及びその医薬組成物の使用の一例である。
【0117】
また、本発明は、ワクチン(特に、適当な賦形剤及び/又はアジュバントを含む医薬組成物に配合された場合)において使用するための、本発明により提供される方法により得られた高活性ウイルスタンパク質、ウイルス粒子及び/又はウイルスを提供する。投与量及び投与方法は前臨床及び臨床開発を通じて当業者により決定されうる。
【0118】
ワクチン−アジュバントは、典型的には、弱いとみなされている動物又はヒトワクチンによりもたらされる防御(すなわち、レベル、度合及び/又は持続時間の点で弱い防御)を増強するために使用される。ワクチン−アジュバントとして使用される糖タンパク質の具体例としては、GM−CSF、並びに特定のサイトカイン、例えばIL−2及びIL−7が挙げられる。ワクチン−アジュバントとしての糖タンパク質は、ワクチンとは別々に、あるいはまた、ワクチンと組合せて投与されうる。ワクチン−アジュバントとしての糖タンパク質をワクチンと組合せる場合には、投与する組成物は、与えられた病原体又は抗原に対する特異的応答を惹起するのに有効な免疫原、薬学的に許容されるワクチン担体、及び免疫を増強する量の糖タンパク質を含有する。その概念の一例は、当技術分野で公知の全細胞ワクチンであり、この場合、抗原の担体である細胞によりワクチン−アジュバントが組換え発現される。全細胞ワクチンは、典型的には、細胞死が運命づけられた活性な細胞として適用される。ワクチン接種後、これらの細胞は、ワクチン−アジュバントとして作用する一定量の糖タンパク質、例えばGM−CSF、IL−2又はIL−7を一定時間にわたって産生する。全細胞ワクチンは、ワクチン接種後の該全細胞ワクチンによりワクチン−アジュバントが更に産生されることなく全細胞ワクチンの利点を尚も有するよう、ワクチン接種前に不活化されうる。本発明は該ワクチン−アジュバントを提供するだけでなく、全細胞ワクチンとして使用される細胞系(1若しくは複数)もまた、本発明により提供される方法により作製されうる。全細胞ワクチンに使用される細胞の特徴は、そのような細胞系又は細胞系の組合せが、可能な限り多数の抗原を発現することである。当業者に公知のとおり、これらの細胞又は細胞系の組合せは、当技術分野において公知の方法により、直接的にワクチン接種されたり、あるいは抗原提示細胞、例えば樹状細胞上に負荷(ローディング)されることが可能である。ついで該抗原負荷樹状細胞はワクチン接種に使用され、あるいは当業者に公知の方法により養子T細胞療法のために身体外(ex corpore)でT細胞を刺激するために使用される。本発明においては、該ワクチンは、本明細書に記載の方法により製造された、ワクチン−アジュバントとして本発明の糖タンパク質を発現する細胞系又は細胞系の組合せでありうる。
【0119】
糖タンパク質又はその医薬組成物は、感染性疾患、例えばエイズ、SARS又は或る形態の肝炎の治療のためのワクチン又はワクチン−アジュバントとして使用されうる。哺乳動物糖タンパク質は、ヒトの感染性疾患、例えばHIV、SARS、B型肝炎若しくはA型及びB型インフルエンザ、又は動物の感染性疾患、例えばウシウイルス性下痢、ウマインフルエンザ、ネコ白血病、ネコ呼吸疾患の治療のために感染細胞に対する免疫応答を誘導する本発明に従い製造される弱毒化生又は不活化ウイルス産物又は組換え抗原性ウイルス産物の一部である。
【0120】
本発明は、本発明の糖タンパク質及び/又はその医薬組成物及び/又はその類似体、修飾体、薬理学的に活性な断片並びに場合によっては使用方法に関する情報を含むキットに関する。本発明の好ましい実施形態においては、該キットは、高い/より高い活性の糖タンパク質、例えばGM−CSF、EPO又はFSH及びその薬学的に許容される担体の医薬組成物の容器、並びにワクチン及びその薬学的に許容される担体の医薬組成物の容器を含むワクチンにおける、抗原に対する哺乳動物の免疫原性応答を増強するために使用されうる。
【0121】
本発明の糖タンパク質は、感染性疾患の治療のためのワクチン又はワクチン−アジュバントとして使用されうる。また、本発明の糖タンパク質は、感染性疾患の治療のためのワクチン又はワクチン−アジュバントの製造にも使用されうる。
【0122】
本発明の糖タンパク質は、疾患、例えば白血病、好中球減少症、血球減少症、癌、骨髄移植、造血系の疾患、不妊症及び自己免疫疾患の予防的及び/又は治療的処置に使用されうる。糖タンパク質の、当業者に公知の治療用途の範囲は、非常に広範である。例えば、G−CSFは、白血病性癌患者の化学療法により生じる、生命を脅かす好中球疾患である好中球減少症を治療するための重要な治療剤である。GM−CSFは、好中球減少症からの速い回復を達成するために、化学療法後の比較的高齢のAML患者の治療に特異的に使用される。さらに、GM−CSFは、骨髄移植におけるいくつかの用途及び末梢血幹細胞の動員(mobilization)のための治療剤として承認されている。また、現在研究中である、GM−CSFのいくつかの臨床用途(例えば、HIV及び癌の治療)が存在する。造血系の或る疾患はEPOで治療され、IFN−βは現在、自己免疫疾患である多発性硬化症の治療のための重要な治療剤である。他の一例はFSHであり、これは男性及び女性不妊症の治療に広く使用されている。hCGは不妊症の治療にも適用されるが、女性における無排卵に焦点が合わされている。hGHは、臨床的に証明されている利益(例えば、体脂肪減少及び筋組織増加)を有する。
【0123】
本発明の糖タンパク質は、白血病、好中球減少症、血球減少症、癌、骨髄移植、造血系の疾患、不妊症及び自己免疫疾患を含む群から選ばれる疾患の予防的及び/又は治療的処置のための医薬の製造にも使用されうる。
【0124】
本発明の糖タンパク質、ワクチン又は医薬は、任意の通常の投与経路、例えば非経口、眼内、局所、吸入、経皮、膣内、頬側、経粘膜、経尿道、直腸、鼻腔、経口、肺又は耳経路により使用されうる。
【0125】
本発明の治療用糖タンパク質の他の一例は抗体である。抗体は、高いアフィニティーで特異的標的に結合することにより作用する。抗体は、毒素又は放射性同位体に結合している場合、腫瘍細胞又は感染細胞に対する強力な治療剤である。しかし、「裸」抗体も、細胞傷害性及び/又は食作用活性を有する特異的免疫細胞(ADCC)又は補体(CDC)を標的に導くことにより、特定の又は細胞標的の排除を媒介しうる。特に、抗体のADCC及び/又はCDC活性は、与えられた抗体のグリコシル化における適当な変化により改善されうる。
【0126】
本発明は本明細書に記載の特定の方法、組成物及び細胞系には限定されないと理解されるべきである。なぜなら、そのような方法、組成物及び細胞系は、もちろん、様々でありうるからである。また、本明細書中で用いる用語は、個々の実施形態を説明する目的のものであるに過ぎず、添付の特許請求の範囲のみによって定義される本発明の範囲を限定するものではないと理解されるべきである。
【0127】
特許請求の範囲を含む本明細書において用いる単数形表現は、文脈が例外を明らかに示している場合を除き、それらの対応複数物を含む。したがって、例えば、「生物」に対する言及は1以上の異なる生物を含み、「細胞」に対する言及は1以上のそのような細胞を含み、「方法」又は「製造方法」に対する言及は、当業者に公知の等価なステップ、方法又は製造方法に対する言及を含む、などである。
【0128】
特に示さない限り、本明細書中で用いるすべての科学技術用語は、本発明が属する分野の当業者により一般に理解されているのと同じ意義を有する。本発明の実施又は試験においては、本明細書に記載されているものと類似した又は同等な方法、製造方法及び物質が使用されうるが、適当な方法及び物質は後記で説明されている。
【0129】
以下の表1及び2、並びに図1〜15は、最適化されたシアル酸付加を伴う高活性ヒトタンパク質の製造のための本発明を説明するものである。
【0130】
表1:種々のヒト細胞系におけるグリコシルトランスフェラーゼの発現プロフィール。示されている値はZR75−1に対するパーセントとして表されている。それぞれの場合に、ハウスキーピングβアクチン対照に対するグリコシルトランスフェラーゼmRNA発現の測定のために二重RT−PCRを用いた。
【0131】
表2:モノクローナル抗体及びレクチンによる種々のヒト細胞系における糖鎖決定基の検出。示されている値は、フローサイトメトリー分析により測定した、陰性対照としてのイソタイプ又は二次試薬に対する、抗体/レクチン結合の差引平均である。例えば、モノクローナル抗体A78−G/A7及びレクチンPNA(ピーナツ凝集素)及びアルトカルプス・インテグリフォリア・ジャカリン(Artocarpus integrifolia Jacalin)はTF特異的である。ノイラミニダーゼでの細胞の処理(白色で文字修飾されている)は、アフィニティー結合を可能にするTF露出を引き起こした。
【0132】
図1:NM−wt細胞のグライコエンジニアリング(GlycoEngineering)及び得られたNM−F9細胞の特徴決定。EMS処理によりランダム突然変異誘発を行った。脱シアル酸エピトープの露出によって、EMS処理NM−wt細胞のアフィニティー選択及びそれに続くクローニングが可能となり、TF特異的抗体A78−G/A7を使用した免疫細胞化学法で測定される高露出TF構造を有する細胞クローンが得られた(A,上パネル:EMS処理NM−wt細胞;下パネル:アフィニティー選択されクローニングされたNM−F9細胞)。シアル酸付加度は、(B)TF特異的抗体A78−G/A7、末端βGal認識性レクチンPNA及びシアル酸特異的レクチンSNAをアプライするフローサイトメトリーにより、あるいは(C)チオバルビツール酸法により測定した。
【0133】
図2:NM−wt細胞並びに誘導体NM−F9及びNM−D4によるTF、sTF、Tn、sTn、グリコホリンA(GPA)、アシアログリコホリンA(AGPA)、Lex、sLex及びLeyの発現。細胞をフローサイトメトリーにより分析した。また、細胞をシアリダーゼで処理し、AGPA(A63−C/A9)及びTF(A78−G/A7)の発現を分析した。
【0134】
図3:(A)レクチンPNA、SNA、MAA及びUEAIを使用するフローサイトメトリーによる、あるいは(B)チオバルビツール酸法による、NM−wt細胞並びに誘導体NM−F9及びNM−D4上の膜複合糖質結合シアル酸の測定。
【0135】
図4:(A)mRNA発現分析による、及び(B)酵素活性分析による、NM−F9細胞の遺伝的欠損の分析。mRNA発現分析は、RNAの単離、UDP−GlcNAc−2−エピメラーゼ特異的プライマーを使用するRT−PCR及びゲル電気泳動を含む。対照としてアクチンを泳動させた。酵素活性測定は、14C−UDP−N−アセチルグルコサミンから14C−N−アセチルマンノサミンへのin vitro変換の測定に基づく。UDP−GlcNAc−2−エピメラーゼのmRNAは検出することができなかった。酵素活性の欠如はエピメラーゼの遺伝的欠損を証明した。
【0136】
図5:代謝的相補によるシアル酸付加の復元。NM−F9及びNM−wt細胞を、0又は50mMのそれぞれManNAc又はGlcNAcを含有する血清中で4日間培養した。モノクローナル抗体A78−G/A7(抗TF)及びA63−C/A9(抗AGPA)並びにレクチンSNA、MAA及びPNAをフローサイトメトリー測定において使用した。ManNAcの添加はシアル酸付加の復元及びTFの消失をもたらした。UDP−GlcNAc−2−エピメラーゼ欠損により、GlcNAcは代謝され得なかった。
【0137】
図6:遺伝的相補によるシアル酸付加の復元。NM−F9細胞を、エピメラーゼをコードする構築物pcDNA3.1Zeo(−)/2Epiで安定にトランスフェクトした。モノクローナル抗体A78−G/A7及びA63−C/A9並びにレクチンSNA及びPNAをフローサイトメトリー測定において使用した。シアル酸付加は完全に復元された。
【0138】
図7:NM−F9におけるrhGM−CSFの発現。ヒトGM−CSF発現ベクターで安定にトランスフェクトされたNM−F9細胞をクローニングし、FCSを添加した又は添加していない培地内で培養した。1日後(1d)、2日後(2d)、3日後(3d)及び4日後(4d)に、ELISAにより上清中のGM−CSF濃度を測定した。4日後、NM−F9細胞105個当たり14ng/mlのGM−CSFが分泌された。
【0139】
図8:種々の分泌シグナルペプチドを使用した場合の、NM−F9細胞による、組換え発現された分泌性AGPAの発現。分泌性AGPAの発現のために、GM−CSFのシグナルペプチド(secGM/GPA)、T細胞受容体のシグナルペプチド(secTCR/GPA)、抗体軽鎖κのシグナルペプチド(secAK/GPA)及び内因性シグナルペプチド(secGPA)を含有する4種の発現ベクターを構築した。捕捉体としてのA83−C/B12(抗GPA)及び検出体としてのA63−C/A9を使用するサンドイッチELISAにより細胞上清中で分泌性AGPAを検出した。
【0140】
図9:代謝改変による膜複合糖質の示差的シアル酸付加。NM−F9細胞を無血清培地内(A)又はfcs含有培地内(B)(種々の濃度のManNAcを添加したもの)で4日間培養した。膜複合糖質結合シアル酸の量を細胞膜画分のチオバルビツール酸法により測定した。比較のために、NM−wt細胞も、ManNAc添加を行わないfcs含有培地内で4日間培養した。
【0141】
図10:代謝的相補による細胞表面上の膜タンパク質の示差的シアル酸付加。NM−F9細胞を種々の濃度のManNAcを添加したFCS非含有培地内で4日間培養した。比較のために、NM−wt細胞も、ManNAc添加を行わないFCS非含有培地内で4日間培養した。シアル酸を含まないTFエピトープを認識するレクチンPNAを使用してフローサイトメトリー分析を行った。
【0142】
図11:代謝的相補による分泌性rhGM−CSFの示差的シアル酸付加。(A)種々の濃度のManNAc(0、50、70又は90mM)及び(B)−/+ 1又は3mg/mlのシアロ糖タンパク質フェツインを添加した無血清培地内でrhGM−CSF産生細胞を4日間培養した。シアル酸を含まないrhGM−CSFをELISAにより測定した。まず、上清中のrhGM−CSFの濃度を測定し、ついでNM−F9細胞培養物1ml当たり100ngに調節した。つぎに、シアル酸を含まないrhGM−CSFの検出のために、抗ヒトGM−CSFモノクローナル抗体を使用してrhGM−CSFを捕捉し、シアル酸を含まないN−及びO−グリカンをPNAにより検出した。
【0143】
図12:TF1細胞上の示差的シアル酸付加rhGM−CSFの増殖活性。示差的シアル酸付加のために、90mMまでの漸増濃度のManNAc(A)及び1mg/mlの規定(defined)血清シアロ糖タンパク質と共に90mM ManNAc(B)を添加した無血清培地内でNM−F9細胞を培養した。TF1細胞を、5ng/mlの種々のシアル酸付加rhGM−CSFを含有するNM−F9上清と共に48時間インキュベートした。Leukine(登録商標)(酵母内で産生)及びLeukomax(登録商標)(大腸菌内で産生)を基準として使用した。GM−CSFを含有しないNM−F9上清を陰性対照として使用して、BrdU−増殖アッセイにより細胞増殖を測定した。最高ではないが高いシアル酸付加度を有するrhGM−CSFが、TF1細胞増殖のための最も活性な増殖因子であった。
【0144】
図13:樹状細胞上の示差的シアル酸付加rhGM−CSFの増殖活性。示差的シアル酸付加のために、90mMまでの漸増濃度のManNAcを添加した無血清培地内でNM−F9細胞を培養した。NemodDC細胞を5ng/mlの種々のシアル酸付加rhGM−CSFのNM−F9上清と共に24時間インキュベートした。Leukine(登録商標)を基準として使用した。GM−CSFを含有しないNM−F9上清を陰性対照として使用して、BrdU−増殖アッセイにより細胞増殖を測定した。異なるシアル酸付加度は、選択された糖タンパク質活性に正又は負の影響を及ぼしうる。
【0145】
図14:rhGM−CSF濃度に対する増殖活性の依存性。rhGM−CSFの最高の活性を得るために、90mM ManNAcを添加した無血清培地内でNM−F9細胞を培養した。種々の濃度のLeukomax(登録商標)、又は種々の濃度のrhGM−CSFを含有するそのNM−F9細胞の上清と共に、TF1細胞を48時間培養した。GM−CSFを含有しないNM−F9上清を陰性対照として使用して、BrdU−増殖アッセイにより細胞増殖を測定した。完全ヒトGM−CSFのin vitro活性は、大腸菌(Leukomax)又は酵母(Leukine)内で発現されたhGM−CSFより約500倍高い。
【0146】
図15:rhGM−CSF増殖活性の抑制。NM−F9細胞を無血清培地+/−90mM ManNAc内で培養した。TF1細胞を5ng/mlのシアル酸付加(a)若しくは非シアル酸付加rhGM−CSF(c)又はLeukine(登録商標)(b)のNM−F9上清と共に48時間培養した。rhGM−CSFは種々の濃度の抗ヒトGM−CSF抗体BVD2−23 B6でブロッキングされた。GM−CSFを含有しないNM−F9上清を陰性対照(d)として使用して、BrdU−増殖アッセイにより細胞増殖を測定した。
【0147】
以下の実施例は例示として記載されており、限定的なものではない。実施例においては、コンタミネーション(汚染)活性を伴わない標準的な試薬及びバッファーを使用する。リボヌクレアーゼ及びPCR産物のコンタミネーションを避けるように留意するのが好ましい。
【実施例1】
【0148】
グライコプロファイリング(GlycoProfiling)
種々のヒト細胞系からのグリコシルトランスフェラーゼC1GalT1、C2GNT−L、C2GNT−M、ST6GalNAc−I、ST6GalNAc−II、ST3Gal−I、ST3Gal−II及びST6Gal−Iの発現プロフィールを調べた。細胞系HEK293、LS174T、MCF−7及びSW480をDMEM内で培養し、一方、T47D、ZR75−1、NM−wt及びKG1をRPMI1640内で培養した。どちらの培地にも10%ウシ胎児血清及び2mMグルタミンを添加した。すべての細胞を6%CO2の湿潤雰囲気中、37℃で増殖させた。当業者に公知の標準的な技術を用いて細胞を回収し破壊した。RNAを、グアニジンチオシアネート−フェノール−クロロホルムを使用して抽出し、RNase非含有DNaseで処理した。RNA収量を分光光度法により測定した。
【0149】
mRNA発現分析を以下のとおりに行った。5μgの全細胞RNAをランダムヌクレオチドヘキサマーでモロニーマウス白血病ウイルス逆転写酵素により逆転写した。RT混合物は逆転写酵素、mRNA鋳型、プライマー、バッファー及びdNTPを含む。cDNAの収量及び完全性を測定するために、並びにゲノムDNAでの考えられうるコンタミネーションに関して確認するために、β−アクチンハウスキーピング遺伝子の発現を利用した。この目的には、イントロンにまたがるβ−アクチン特異的プライマーセットを設計した:β−アクチンフォワードプライマー:5'-GGC ATC GTG ATG GAC TCC G-3'及びβ−アクチンリバースプライマー5'-GCT GGA AGG TGG ACA GCG A-3'(アンプリコン長,622bp)。
【0150】
得られたcDNA鎖を直接、増幅のために使用した。前記酵素の1つをコードするcDNAの5’−(フォワード)及び3’−領域(リバース)に特異的にハイブリダイズするフォワードプライマー及びリバースプライマーを設計した(C1GalT1を、フォワードプライマー:GAG ATT CCA GAG ATA CCA TTG及びリバースプライマー:CGT TCA GGT AAG GTA GGT TGを使用して増幅した(アンプリコン長262);C2GNTのフォワード:GTG CTC AGA ATG GGG CAG GAT GTC ACC TGG, リバース:TCA CTA CTA GGA TTC TCC CCA GCA AGC TCC(アンプリコン長360);ST3Gal−Iのフォワード:ATG AGG TGG ACT TGT ACG GC, リバース:AAC GGC TCC AGC AAG ATG(アンプリコン長375);ST3Gal−IIのフォワード:CCC TGC TCT TCA CCT ACT CG, リバース:GCA TCA TCC ACC ACC TCT G(アンプリコン長282);ST6Gal−Iのフォワード:AAA AAC CTT ATC CCT AGG CTG C, リバース:TGG TAG TTT TTG TGC CCA CA(アンプリコン長379);ST6GalNAc−Iのフォワード:ACC ACA GCC AAG ACG CTC, リバース:AAG GGT GGT GCA AAG TGT TC(アンプリコン長407);ST6GalNAc−IIのフォワード:CTG CCA GTA AAT TCA AGC TGC, リバース:TTG CTT GTG ATG AAT CCA TAG G(アンプリコン長184))。PCR反応混合物は、1.5μlのPCRバッファー[100mM Tris−HCl(pH8.3),500mM KCl]、0.8〜1.0μlの25mM MgCl2、5μlのcDNA、1.5μlの2mMデオキシヌクレオチド三リン酸、0.2μlのAmpliTaqポリメラーゼ、0.2μlの0.25μMβ−アクチンプライマー、0.7〜0.8μlの50μM酵素特異的プライマー及びH2O(容量15μlとする)を含有していた。PCRは、変性、アニーリング及び伸長のための複数の温度サイクルで行った。サイクル条件は94℃で1分間、59℃で1分間及び72℃で2分間であった。同じチューブ内で0.2μlのβ−アクチンプライマーでβ−アクチンを同時増幅した。各PCR反応を2回行った。
【0151】
26〜28サイクルの後で得られたPCR反応のアリコートを2%アガロースゲル中で電気泳動し、臭化エチジウム染色の強度により定量した。RT−PCRデータの半定量的分析のために、各cDNA特異的バンドの染色強度を、同時増幅されたβ−アクチンに対応するバンドの染色強度と比較した。比率[(酵素バンドの蛍光単位/β−アクチンの蛍光単位)×100]を計算することにより、各酵素バンドの強度を、適当なβ−アクチンバンドと相関させた。検出限界は二本鎖DNAとして1ngであった。該アッセイは25ngまでは直線関係を示した。
【0152】
グライコプロファイリング(GlycoProfiling)は、NM−wt細胞のみにおいて、C2GNT−L及びC2GNT−Mの欠如により、コア2の合成の完全な欠損を示した。mRNA発現における他の低減(ST6GalNAc−IIは8%へ、ST3Gal−Iは32%へ)は無傷ST6GalNAc−I及びST3Gal−IIにより補償されることが可能であった(表1)。NM−wtのグリコシルトランスフェラーゼプロフィールは、将来のグライコエンジニアリング(GlycoEngineering)及びシアル酸付加研究(TFエピトープを使用することによるもの)のための最良の出発点を表した。
【0153】
特にシアル酸に関する糖鎖決定基は、いくつかのモノクローナル抗体及びレクチンをアプライするフローサイトメトリーにより、前記ヒト細胞系において検出された。
【0154】
細胞を集め、PBSで2回洗浄し、37℃で1時間かけて回復させた。ノイラミニダーゼ処理のために、細胞を、0.1%CaCl2及び0.4%NaClを含有する0.1Mイミダゾールバッファー(pH6.8)中、ビブリオ・コレラ(V. cholerae)ノイラミニダーゼの存在下又は非存在下にて、37℃で30分間インキュベートした。細胞を染色バッファー(4%BSAを含有するHBSS又は10%FCSを含有する培地)に再懸濁させた。細胞表面糖鎖決定基の検査のために、種々の抗体及びレクチンを使用した。例えば、ビオチン標識レクチンピーナツ凝集素(PNA)、アルトカルプス・インテグリフォリア(Artocarpus integrifolia)由来のジャカリン(jacalin)、アマランツス・カウダツス(Amaranthus caudatus)凝集素(アマランチン(Amaranthin)又はACA)及びバウヒニア・プルプレア(Bauhinia purpurea)レクチン(BPL)並びにTF特異的モノクローナル抗体A78−G/A7を使用して、TFの発現を分析した。PankoMabは腫瘍−MUC1を検出する。A83−C/B12は、グリコホリンAをそのグリコシル化には無関係に認識する抗GPA抗体である。細胞をレクチン又は抗体と共に4℃で1時間インキュベートし、染色バッファーで2回洗浄した。該抗体の結合は、二次Cy3−ヤギ抗マウスIgG/IgM抗体により検出した。レクチンの結合は、Cy3結合ストレプトアビジンにより検出した。Coulter Epics XLフローサイトメーター(Beckman Coulter, Germany)を使用してフローサイトメトリー分析を行った。Expo32ソフトウェア(Becton Coulter)を使用して定量分析を行った。
【0155】
ZR75−1の場合を除き、ヒト細胞はTF陰性であった。しかし、シアル酸を切り離すノイラミニダーゼで細胞を処理した後、多数の細胞系上で潜在TFが可視化される。分析した細胞系のうち、NM−wt細胞が潜在TFの主要キャリアであった。本発明においては、多量の元の潜在TF及び遊離シアル酸が好ましい(表2)。
【実施例2】
【0156】
グライコエンジニアリング(GlycoEngineering)及び新規クローンの特徴決定
NM−wt細胞をアルキル化剤メタンスルホン酸エチルで処理することにより、ランダム突然変異誘発を行った。サンプルごとに、NM−wt細胞をPBSで洗浄し、5%CO2、37℃で一晩、EMS(0.1mg/ml、メタンスルホン酸エチル、Sigma-Aldrich)を添加した細胞培養培地1ml当たり106細胞で播いた。細胞を洗浄し、新鮮培地を供給した。2日ごとに、トリパンブルー染色により細胞生存度を測定し、免疫細胞化学的染色により細胞を分析した(図1A)。
【0157】
ついで、高いTF発現の新規表現型を示す細胞を、TF特異的抗体を使用して選択した。NM−wt細胞をB−PBS(PBS中、0.5%BSA)で洗浄し、モノクローナル抗体A78−G/A7又はPankoMabのハイブリドーマ培養の上清50μl及びB−PBSの950μlと共に4℃で30分間インキュベートした。洗浄後、50μlのラット抗マウスIgM抗体又はラット抗マウスIgG抗体(MicroBeads(Miltenyi Biotec, Koln, Germany)に結合しているもの)で操作を繰返した。洗浄後、Miltenyi Biotec(Koln, Germany)により供給された2つの連続的なカラムにより、該製造業者のマニュアルに記載されているとおりに、磁気標識TF陽性細胞を分離した。9日間の培養の後、単離操作を合計3回繰返した。FACS分析(フローサイトメトリー)を抗体染色から始めた。約3×105細胞を一次モノクローナル抗体[A78−G/A7(IgM)、PankoMab(IgG1)のハイブリドーマ培養上清;すべて、細胞培養培地中で1:2希釈されたもの]と共に4℃で1.5時間インキュベートし、ついで、PBSで1:200希釈された二次Cy3結合ヤギ抗マウスIgM/IgG抗体と共に4℃で30分間インキュベートし、再び洗浄した。再懸濁細胞(200μlのPBS)をフローサイトメトリーにより調べた。以下のパラメーターでExpo32ソフトウェア(Becton Coulter)を使用して、定量分析を行った:抗体標識細胞に関するパラメーター:前方散乱(FS):26V,ゲイン1,側方散乱(SS):807V,ゲイン5,FL2:740V,ゲイン1、及びレクチン標識細胞に関するパラメーター:FS:26V,ゲイン1,SS:807V,ゲイン5,FL1:740V,ゲイン1。
【0158】
3回の単離の後、93%TF陽性細胞の細胞集団を得た。しかし、TF陽性細胞の比率は時間経過と共に減少し、単離操作後の14日間の期間中にTF陽性細胞約20%の最低レベルに達した。TF陽性表現型の安定発現のために、細胞を4回目の単離に付し、ついで最後に、単離されたTF陽性細胞を96ウェルプレート(1細胞/100μl)内での限界希釈によりクローニングした。得られた30個の細胞クローンのうち、13個の細胞クローンが多量のTF抗原を発現し、これらのうちの8個の細胞クローンが全細胞集団の細胞表面上で、均一なTF発現を示した(図1A)。現在までに(約30ヶ月間)、これらの細胞クローンはTF抗原を安定に発現している。フローサイトメトリーによる分析は、NM−F9細胞の場合のTF発現レベルの約31倍の増加を示し、このことは、このクローン上のTFの強力な発現を表しており、約35の、FACS分析における差引平均によっても示され、これを更なる特徴決定のために選択した。安定なTF陽性細胞集団を得るためにクローニングが必要なのは、TF陰性細胞の、より高い増殖速度によるものであり、したがってこれは時間経過と共にTF陽性クローンより優勢に増殖し、例えば、NM−F9は、NM−wt細胞より遅い倍加速度を有する。
【0159】
さらに、PankoMab及びラット抗マウスIgG抗体(MicroBeads(Miltenyi Biotec, Koln, Germany)に結合しているもの)を使用した以外は上記TF選択と同じ方法で、MUC1選択を行った。より多くの腫瘍特異的MUC1エピトープTA−MUC1を発現するTF陽性クローンの作製のために、NM−F9細胞を処理し、選択し、単一細胞をクローニングした。これは、選択のためにPankoMabを使用して、前記のとおりに行った。安定なクローンNM−D4は、フローサイトメトリーにおいてPankoMab染色の増強を示したため、該クローンを更なる特徴決定のために選択した。
【0160】
NM−F9及びNM−D4は、より低い連続的な濃度のウシ胎仔血清(FCS)を含有する培地内に継代することにより、無血清培地に適合化することが可能であった。
【0161】
NM−F9及びNM−D4を、それらの糖鎖決定基及びトランスフェラーゼ発現に関して詳細に特徴決定した。評価されうる膜複合糖質のシアル酸付加度を、実施例1に記載のとおりにTF特異的抗体A78−G/A7及びいくつかのレクチン、例えば末端β−Gal認識性PNA、シアル酸特異的サムブクス・ニグラ(Sambucus nigra)レクチン(SNA)、又はその他の例えばマアキア・アムレンシス(Maackia amurensis)レクチンI(MAA)及びウレックス・ユーロパエウス(Ulex europaeus)凝集素I(UEAI)をアプライするフローサイトメトリーにより測定した。PBSで2回及び4%BSA含有HBSS(ハンクス平衡塩)で1回洗浄した後、細胞をHBSS/4%BSA中、FITC結合PNA(1:400)、FITC結合SNA(1:50)、FITC結合MAA(1:50)又はFITC結合UEAIと共に4℃で1時間インキュベートした。洗浄後、分析のために細胞ペレットを200μlのHBSS/4%BSAに再懸濁させた。以下のパラメーターでExpo32ソフトウェア(Becton Coulter)を使用して、定量分析を行った:抗体標識細胞に関するパラメーター:前方散乱(FS):26V,ゲイン1,側方散乱(SS):807V,ゲイン5,FL2:740V,ゲイン1、及びレクチン標識細胞に関するパラメーター:FS:26V,ゲイン1,SS:807V,ゲイン5,FL1:740V,ゲイン1(図2及び3A)。
【0162】
野生型細胞の潜在TFはA78−G/A7及びPNAの結合を妨げたが、誘導体NM−F9及びNM−D4の露出TFは、使用した検出試薬に応じて認識を30〜75倍増強した。この結果はSNAの逆アフィニティーにより定性的に確認された(図1B)。また、膜結合シアル酸の量を定量した。簡潔に説明すると、細胞を集め、PBSで1回洗浄し、針せん断と組み合わせて細胞溶解バッファー(50mM Tris/HCl pH8,150mM NaCl,0,5%NonidetP40,1mM EDTA,200μg/mlフェニルメチルスルホニルフルオリド)中で細胞溶解した。粗膜画分を40.000×gで20分間の遠心分離によりペレット化した。ペレットを水で2回洗浄し、凍結乾燥させた。上清を酵素活性測定に使用した(実施例3を参照されたい)。タンパク質濃度をRoti−Quant−Kit(Roth, Germany)で測定した。該ペレットを2M酢酸で80℃で1時間加水分解することにより、膜複合糖質結合シアル酸の含量を測定し、チオバルビツール酸法(Aminoff, Biochem. J. 81, 384-392, 1961)により遊離シアル酸を測定した。NM−F9及びNM−D4細胞内の膜結合シアル酸は、野生型細胞の場合と比較して、約3分の1に減少している(図1C、3B)。
【実施例3】
【0163】
NM−F9細胞の遺伝的欠損の分析
NM−F9細胞のTF露出は糖ヌクレオチド生合成経路又は輸送体におけるいくつかのシアリルトランスフェラーゼにおける欠損を要すると予想された。実施例1に記載のとおりに種々の酵素のmRNA発現分析を行い、この場合、NM−F9酵素に属するゲルバンドをNM−wtのものと比較し、UDP−GlcNAc−2−エピメラーゼmRNAの全欠如が示された(図4A)。ペーパークロマトグラフィーを使用して14C−UDP−N−アセチルグルコサミンから14C−N−アセチルマンノサミンへの変換を測定することにより、NM−wt及びNM−F9細胞におけるUDP−GlcNAc−2−エピメラーゼの酵素活性を細胞上清において測定した(実施例2を参照されたい)。1Uは37℃で1分当たり1μmolのManNAcの合成に相当する。UDP−GlcNAc−2−エピメラーゼ活性はNM−F9においては、NM−wt細胞における元の300μU/mgから完全に低下した(図4B)。
【0164】
シアル酸付加を復元するNM−F9細胞の能力を調べるために、培地に50mM ManNAcを添加した。4日間の培養後、残存しているシアル酸を含まないグライコトープ(glycotope)を、既に記載されているとおりにモノクローナル抗体A78−G/A7及びA63−C/A9並びにレクチンPNA、SNA及びMAAをアプライするフローサイトメトリーにより検出した。ManNAcを供給することによる代謝的相補は、検出試薬に応じて、潜在膜複合糖質TFの約50%(PNA)〜80%(A78−G/A7)をもたらした。UDP−GlcNAc−2−エピメラーゼ欠損のため、対照GlcNAcはNM−F9細胞により代謝され得なかった(図5)。
【0165】
また、遺伝的相補によるシアル酸付加の復元を調べた。UDP−GlcNAc−2−エピメラーゼcDNAをベクターpcDNA3.1内にクローニングした。SuperFect試薬を使用して、エピメラーゼをコードする構築物pcDNA3.1Zeo(−)/2EpiでNM−F9細胞を安定にトランスフェクトした。細胞を40%コンフルエンスまで増殖させた。5μgのDNAを150mlのDMEMに溶解し、30μlのSuperFectと混合した。室温で10分間のインキュベーションの後、この溶液を細胞に加えた。3時間後、培地を除去し、細胞をPBSで3回洗浄し、新鮮な増殖培地と共にインキュベートした。48時間のインキュベーション時間の後、増殖培地を再び、700μg/ml G418又は750μg/mlゼオシンを含有する培地と交換した。耐性クローンを、400μg/ml G418又は250μg/mlゼオシンを含有する増殖培地内で連続的に維持し、フローサイトメトリーにより分析した。シアル酸を含まないグライコトープを検出するために、モノクローナル抗体A78−G/A7及びA63−C/A9並びにレクチンPNA及びSNAを使用した。シアル酸付加度は完全に復元された(図6)。
【実施例4】
【0166】
NM−F9細胞における分泌性糖タンパク質の組換え発現
組換えヒト増殖因子rhGM−CSFを安定に発現するNM−F9細胞を、エレクトロポレーションを用いて発現ベクターpGT60hGM−CSF(Invivogen, USA)で該細胞をトランスフェクトすることにより作製した。エレクトロポレーションのために、106細胞を集め、洗浄し、400μlの低浸透圧バッファー(Eppendorf, Germany)に再懸濁させ、室温で15分間インキュベートした。ついで細胞懸濁液を8μgのプラスミドDNAと混合し、2mmの間隙を有するエレクトロポレーションキュベット(Eppendorf)内に移した。340Vの電圧及び5μ秒の時間を用いてMultiporator(Eppendorf)においてエレクトロポレーションを行った。培地(10%fcs,RPMI1640中の1%グルタミン,Biochrom)において、エレクトロポレーションからの回復の1〜2日後、安定にトランスフェクトされた細胞を、100μg/mlの抗生物質ヒグロマイシンBを使用することにより選択し、限界希釈法によりクローニングした。約4週間後、GM−CSF含量を定量するために使用しうるGM−CSF特異的ELISA(Becton-Dickenson, USA)を使用することにより、いくつかの細胞クローンの上清を分泌性rhGM−CSFの存在に関して分析した。rhGM−CSFの最高の分泌率を有する1つの細胞クローンを更なる実験のために選択した。第1の実験においては、GM−CSFを安定に発現する約105個のNM−F9細胞を、ウシ胎仔血清(fcs)の存在下及び非存在下にて、培地(前記参照)内で培養した。培地を交換することなく1、2、3及び4日間培養した後、時間経過と共に蓄積した分泌性rhGM−CSFの量を、GM−CSF特異的ELISAを用いることにより測定した。fcsの存在下で培養されたか非存在下で培養されたかには無関係に、4日間で約14ngのGM−CSFが105個の細胞により放出された(図7)。第1日の後の分泌率は、fcsの存在下で培養した場合(約6ng/ml)には、fcsの非存在下で培養した場合(約4ng/ml)と比べて若干高かった。
【0167】
発現ベクター構築物secGPAを使用して、ヒト膜タンパク質グリコホリンA(GPA)をNM−F9細胞内で分泌性アシアログリコホリンAとして組換え発現させた。secGPAは、膜貫通ドメイン及び細胞質尾部を含まないGPAのN末端の91アミノ酸(aa)をコードするcDNAを含有する。このcDNAを真核生物発現ベクターpcDNA5/FRT/V5−His−TOPO及び/又はpcDNA3.1/myc−His C(共にInvitrogen)内にサブクローニングした。さらに、内因性シグナルペプチドが異種ヒトシグナルペプチドにより置換された分泌性AGPAタンパク質の組換え融合タンパク質を発現させるために、NM−F9細胞を使用した。したがって、当業者に公知の分子生物学的方法を用いて、19個のN末端アミノ酸をコードするcDNAの代わりに、GM−CSF由来のシグナルペプチド(MWLQSLLLLGTVACSIS,secGM/GPA)、T細胞受容体由来のシグナルペプチド(MACPGFLWALVISTCLEFSMA,secTCR/GPA)又は抗体κ軽鎖由来のシグナルペプチド(METDTLLLWVLLLWVPPGSTGD,secAK/GPA)をコードする異種cDNAが組込まれたcDNAであるsecGM/GPA、secTCR/GPA及びsecAK/GPAを構築した。後者の2つのシグナルペプチドは、任意の遺伝子産物の分泌性発現を増強するための異種シグナルペプチドとしての使用に関して当業者に既に公知である。secGM/GPAの場合には、該シグナルペプチドとGPAをコードする骨格との間において追加的なアラニンが発現される。それらの3つのcDNA構築物を前記発現ベクター内にサブクローニングし、ヌクレオチド配列を確認した。得られた発現ベクターのそれぞれを、上述した手順、すなわちエレクトロポレーション、ヒグロマイシンBでの選択及び単一細胞クローニングを用いることにより、安定にトランスフェクトされたNM−F9細胞の作製に使用した。発現ベクターpcDNA5/FRT/V5−His−TOPOを使用する場合には、2つのFRTリコンビナーゼ認識部位により挟まれたlacZeo DNAカセット(Invitrogen)を含有する遺伝的に改変されたNM−F9細胞をpcDNA5/FRT/V5−His−TOPO及びリコンビナーゼ発現ベクターpOG44(Invitrogen)で共トランスフェクトした。任意のGPAを細胞培養上清から外へ捕捉するためのA83−CB12抗体及び細胞外ドメイン上に局在するTF抗原を認識するA63−C/A9抗体を使用するサンドイッチELISAにより、細胞培養培地内に放出された分泌性AGPAの分泌に関して、secGM/GPA、secTCR/GPA、secAK/GPA又はsecGPAを発現する細胞クローンをスクリーニングした。図8は、最高量の分泌性AGPA(1ml当たり及び105細胞当たり40ng/mlまで)を産生する細胞クローンがsecGM/GPA構築物で作製されたことを示している。その他の構築物を使用して、1ml当たり及び105細胞当たり最大で13ngの分泌性AGPAを検出することが可能であった。さらに、分泌性AGPAを産生する細胞クローンの数は、secGM/GPA構築物を発現ベクターとして使用した場合には、有意に、より高かった(図8)。
【実施例5】
【0168】
代謝的相補による示差的シアル酸付加
50mM ManNAcでのNM−F9細胞の代謝的相補により得られるシアル酸付加の部分的復元(図5)に関して、シアル酸付加に対する一定量の糖中間体の影響を詳細に調べた。NM−F9細胞を、0〜90mM ManNAcの存在下、fcsを含有する又は含有しない培地内で培養し、該細胞膜のシアル酸付加度をチオバルビツール酸法により分析した(実施例3を参照されたい)。細胞シアル酸付加度はfcsの非存在下又は存在下においてManNAcの量が増加することに伴って代謝的相補により制御されうることを示すことができた(図9)。fcs非含有培地におけるManNAcの添加無しでは、NM−F9細胞の膜画分においてはシアル酸は少量のみしか検出されなかった。fcs非含有培地に加えるManNAcの量を次第に増加させることによりシアル酸付加度を徐々に増加させることが可能であった(50mM ManNAcでプラトーに達した)。30mM ManNAcを添加したfcs含有培地においては、シアル酸付加における更なる増加が達成された。fcs含有培地に90mM ManNAcを添加した場合には、元のNM−wt細胞に匹敵するくらい高い、最高のシアル酸付加度が得られた。種々の濃度のManNAcでの代謝的相補による膜タンパク質の示差的シアル酸付加を、シアル酸を含まないTFエピトープを認識し従ってシアル酸付加度に逆相関するレクチンPNAを使用するフローサイトメトリーにより分析した。細胞膜内のシアル酸の量だけでなく膜内の糖タンパク質上のシアル酸付加度も種々の量のManNAc量での代謝的相補により制御可能であることが示されている(図10)。培地内に存在するManNAcを次第に増加させることにより、次第に増加するシアル酸付加度が観察された。90mM ManNAcを用いた場合、NM−F9細胞における糖タンパク質のシアル酸付加度はNM−wt細胞の場合とほぼ同等に高かった。
【0169】
最後に、分泌性タンパク質(この場合はrhGM−CSF)のシアル酸付加度もManNAc及び血清タンパク質での代謝的相補により制御可能であるかどうかを試験した。rhGM−CSF発現ベクターで安定にトランスフェクトされたNM−F9細胞(実施例4を参照されたい)を再び、漸増量の0〜90mM ManNAcと共に、並びに1及び3mg/mlのシアロ糖タンパク質フェツインの存在下の90mM ManNAcと共に培養した。4日後、得られた細胞培養上清中のrhGM−CSFの濃度をELISAにより測定し(実施例4を参照されたい)、ついで細胞培養培地1ml当たり100ngに調節した。シアル酸を含まないrhGM−CSFの度合を測定するために、サンドイッチELISAを確立した。簡潔に説明すると、GM−CSF特異的ELISA(Becton-Dickenson)の抗ヒトGM−CSFモノクローナル抗体をrhGM−CSFの捕捉に使用し、したがってPBS中1:250の希釈度でマイクロタイタープレートに4℃で一晩コートした。PBS/0.05%Tween20中での洗浄及びPBS中の10%fcsでのブロッキングの後、100μlの該細胞培養上清を該マイクロタイタープレートに加え、室温で2時間インキュベートした。シアル酸を含まないO−グリカンをビオチン化PNA(5〜10μg/ml)及びペルオキシダーゼ結合ストレプトアビジン(1:250希釈物)で検出した。分泌性組換え糖タンパク質、例えばrhGM−CSFも、漸増量のManNAcによるNM−F9細胞の相補により示差的にシアル酸付加されうることが示された(図11)。90mM ManNAcへの1mg/mlフェツインの添加により最高のシアル酸付加度が達成される。90mM ManNAcへの3mg/mlフェツインの添加はシアル酸付加における更なる増強をもたらさない。
【実施例6】
【0170】
種々のシアル酸付加形態の活性の測定による高活性ヒトGM−CSFの同定
種々のシアル酸付加タンパク質の活性を評価するために、GM−CSFの増殖刺激能を採用した。rhGM−CSFを発現するNM−F9細胞(実施例4及び5を参照されたい)を、0、30、50、70及び90mM ManNAcで並びに1mg/mlのフェツインと共に90mM ManNAcを添加した無血清培地を含有する種々の調製物において培養した。実施例5に既に記載されているとおりにシアル酸付加を検出した。TF−1細胞及び単球性樹状細胞系NemodDC(それらの増殖速度は共にGM−CSFに左右される)を、種々のシアル酸付加rhGM−CSFが5ng/mlに調節された細胞上清と共にそれぞれ48時間又は24時間インキュベートした。細胞増殖を、製造業者のプロトコール(Roche Diagnostik GmbH, Mannheim, Germany)に従いBrdU−増殖アッセイにより測定した。固定後、細胞をPOD標識抗BrdU抗体と共にインキュベートした。後続の染色反応を1M硫酸により停止させ、450nmの吸光度(参照690nm)で分光光度法により検出した。GM−CSFを含有しないNM−F9上清を陰性対照として使用した。それぞれ酵母及び大腸菌内で発現された非シアル酸付加rhGM−CSF形態である市販のLeukine(登録商標)(Schering AG)及びLeukomax(登録商標)(Schering-Plough)を基準として使用した。
【0171】
驚くべきことに、rhGM−CSFの細胞増殖刺激活性はrhGM−CSFのシアル酸付加度によって大きく異なり、90mM ManNAcを含有する培養から得られた、最高ではないが高度にシアル酸付加されたrhGM−CSFの1つのシアル酸付加形態が最高活性を示した。このrhGM−CSFは、Leukine(登録商標)(100%)と比較した場合に、TF−1細胞及びNemodDCに関して約178±10%の増殖刺激活性の増加を示した(図12及び13)。市販のrhGM−CSFであるLeukomax(登録商標)及びLeukine(登録商標)の細胞増殖活性は、NM−F9細胞内で発現された最も活性なシアル酸付加形態より低いことが判明した(図12)。5ng/mlのLeukomax(登録商標)又はLeukine(登録商標)で観察された最高の増殖活性は、0.01ng/mlの、NM−F9細胞からの最適シアル酸付加形態により達成される(Leukine(登録商標)又はLeukomax(登録商標)と比較して約500倍低いGM−CSF;図14)。同じ濃度のLeukine(登録商標)、Leukomax(登録商標)又はrhGM−CSFの最適シアル酸付加形態で該活性を分析したところ、活性の増加は5倍までであった(図14)。
【0172】
抗ヒトGM−CSF抗体BVD2−23 B6をTF1細胞増殖アッセイに加えた場合には、GM−CSF活性が特異的にブロッキングされた。5μg/mlの抗体濃度では既に、Leukine(登録商標)又はrhGM−CSFの最適シアル酸付加形態によるTF−1細胞増殖の刺激は約60%ブロッキングされ、40μg/mlの抗体濃度では完全に消失した(図15)。
【0173】
配列表:
配列番号1:メチオニン、トリプトファン、ロイシン、グルタミン、セリン、ロイシン、ロイシン、ロイシン、ロイシン、グリシン、トレオニン、バリン、アラニン、システイン、セリン、イソロイシン、セリン。
【表1】

【表2】
【技術分野】
【0001】
本発明は、最適化されたシアル酸付加度(sialylation degree)を有する高活性糖タンパク質、診断又は治療に用いるための該糖タンパク質を含む医薬組成物、高活性糖タンパク質及びその製造のための条件の決定方法、高活性糖タンパク質の製造方法、並びに分泌性糖タンパク質の示差的シアル酸付加方法に関する。また本発明は、生物学的目的のため、並びに、疾患(特に骨髄移植、好中球減少症、血球減少症、AML及び骨髄異形成症候群、癌、HIV及び/又は造血系疾患)の予防的及び/若しくは治療的処置、又は診断のための、組換え発現された高活性糖タンパク質の使用に関する。
【背景技術】
【0002】
糖タンパク質は、機能及び発生が各糖タンパク質間で大きく異なっている多様な一群である。潜在的治療能を有するタンパク質のうちのほとんどのタンパク質はグリコシル化されており、これは、例えば多数のホルモン(例えば、成長ホルモン、グリカゴン、FSH及びLH)、増殖因子(例えば、GM−CSF、G−CSF、VEGF及びエリスロポエチン)、サイトカイン(例えば、IL−2、IL−7、インターフェロンα及びβ、TNFα)、抗凝固剤(例えば、レピルジン(Lepirudin)、デシルジン(Desirudin))、血液凝固因子(例えば、第VII因子、第VIII因子及び第IX因子)、ワクチン(例えば、B型肝炎抗原)及び抗体で見られる。そのようなタンパク質を製造するための確立された細胞産生系は、元のヒトグリコシル化を有するタンパク質を産生することができない。原核生物(例えば、細菌)及びほとんどの真核細胞系(例えば、酵母、昆虫及び植物細胞)は、グリコシル化を欠くタンパク質を合成し、あるいはヒト糖鎖とは大きく異なるグリカンを含有するタンパク質を合成する。チャイニーズハムスター卵巣(CHO)細胞は、ヒト細胞と同様にタンパク質をグリコシル化しうる一般に使用されている産生系であるが、CHO細胞は、例えば、ガラクトシル化、フコシル化、特に、Nアセチルグリコサミンでのグリコシル化及び特にシアル酸付加の種々の態様において、いくつかの重要な相違を有する。
【0003】
これらの産生系が確立された時点においては、少なくとも或る程度活性である治療用タンパク質が産生されることで十分であった。今日では、(i)治療用タンパク質の適用用量の回数及び濃度を減少させるために、(ii)治療のコストを減少させるために、並びに(iii)副作用を軽減するために、治療用タンパク質の活性を改善することに集中した多大な努力がなされている。
【0004】
1つの方法は、発現系の生産性を増加させるものであるが、ほとんどの場合には、これはグリコシル化を全くもたらさないか非常に異常なグリコシル化しかもたらさず、ほとんどの場合には、それはタンパク質には不適当なものである。他の問題点、例えば分子の安定性及びフォールディングの低下も生じ、このため、しばしば、追加的な製造工程が必要となり、これは時間及びコストを浪費し、無駄、副作用及び最適未満の生物活性を与える。
【0005】
タンパク質の生物活性を改善するための主な方法は、その血清半減期を延長させ、したがってそのバイオアベイラビリティを向上させるものである。これは、PEG付加(PEGylation)と称される方法により行われ、この場合、産生されたタンパク質に或る形態のポリエチレングリコールが化学的に付加される。PEGは分子量を増加させ、したがって血清半減期を延長させる。しかし、この方法にはいくつかの問題が伴う。例えば、ほとんどすべての場合において、PEG付加はその細胞エフェクター機能によりタンパク質の活性を減少させ、ヒトにおける反復投与は、しばしば、中和抗体としての有害な免疫応答を引き起こし、及び/又は、該製造方法は追加的な化学修飾を要し、さらなるコスト、無駄及び時間の浪費を招く多段階の方法となる。
【0006】
糖鎖の修飾は、組換え発現されたタンパク質の血清半減期を改善することに焦点が合わされている。したがって、この技術は、組換え糖タンパク質のシアル酸付加度を最大にすることに焦点が合わされている。シアル酸は、真核細胞の表面上に最もよく見られる末端単糖であり、糖タンパク質のシアル酸付加が多くなればなるほど、循環中のその血清半減期は長くなる、と一般には考えられている。これは、循環非シアル酸付加タンパク質に結合し分解のためにそれを細胞内に導く肝臓内のアシアロタンパク質−受容体としての或る受容体の存在に基づくものである。
【0007】
組換え糖タンパク質のシアル酸付加度を可能な限り高くまで増加させるためのいくつかの方法が提示されており、これらには例えば、(i)ほとんどの末端にシアル酸付加をもたらすのに寄与する特定のシアリルトランスフェラーゼ又はガラクトシルトランスフェラーゼを過剰発現する組換え宿主細胞系を得ること、(ii)糖加水分解酵素を、例えばCuイオンを培地に加えて調節することにより、宿主細胞シアル酸の分解を妨げること、(iii)オリゴ糖産生を増強するための追加的な複合糖質及び他の基質の存在下、組換え糖タンパク質を産生する宿主細胞を培養すること、(iv)培養条件を種々変化させること、(v)追加的な糖鎖を導入するために、したがってより多数のシアル酸を導入するために、該タンパク質を突然変異させること、並びに(vi)該糖タンパク質の産生後、精製されたシアリルトランスフェラーゼでin vitroシアル酸付加を完了させることが含まれる。これらのすべての技術は一連の欠点を伴い、シアル酸付加度を最大にすることにより血清半減期を延長させるよう意図されているに過ぎない。
【0008】
前記のとおり、先行技術は一連の決定的な欠点を有し、生物活性の改善、副作用又は毒性の軽減のためのグリコシル化の可能性のごく一部しか検討しておらず、したがって糖タンパク質を何らかの用途に結びつけるものではない。対応する組換え糖タンパク質に関する適当な試験系及び産生系が全く利用できないため、先行技術は、糖タンパク質の生物活性がシアル酸付加度と相関しているかどうかについての認識をほとんど完全に欠いている。
【発明の概要】
【0009】
本発明は、シアル酸の糖ヌクレオチド生合成経路における少なくとも1つの欠損を含有し、かつ糖タンパク質をコードする核酸でトランスフェクトされた発現細胞系において、ある濃度の少なくとも1つのシアル酸前駆体添加物を添加した培地内で高活性糖タンパク質を発現させることを含む方法により製造される高活性糖タンパク質であって、
該濃度が、
(i)種々の濃度の少なくとも1つのシアル酸前駆体を使用する示差的シアル酸付加により、複数の異なるシアル酸付加形態の糖タンパク質を発現させること、
及び
(ii)適当なバイオアッセイにおいて、参照糖タンパク質と比較して、前記の異なるシアル酸付加形態の活性を測定すること、
及び
(iii)より高い/最高の活性を有するシアル酸付加形態を選択し、該糖タンパク質のより高い/最高の活性レベルと相関する、該シアル酸前駆体添加物の濃度を決定すること
を含む方法により決定される、高活性糖タンパク質を提供することにより、この課題を解決するものである。
【0010】
したがって、また本発明は方法に関する。
【図面の簡単な説明】
【0011】
【図1】NM−wt細胞のグライコエンジニアリング(GlycoEngineering)及び得られたNM−F9細胞の特徴決定を示す。
【図2】NM−wt細胞並びに誘導体NM−F9及びNM−D4によるTF、sTF、Tn、sTn、グリコホリンA(GPA)、アシアログリコホリンA(AGPA)、Lex、sLex及びLeyの発現を示す。
【図3】(A)レクチンPNA、SNA、MAA及びUEAIを使用するフローサイトメトリーによる、あるいは(B)チオバルビツール酸法による、NM−wt細胞並びに誘導体NM−F9及びNM−D4上の膜複合糖質結合シアル酸の測定を示す。
【図4】(A)mRNA発現分析による、及び(B)酵素活性分析による、NM−F9細胞の遺伝的欠損の分析を示す。
【図5】代謝的相補によるシアル酸付加の復元を示す。
【図6】遺伝的相補によるシアル酸付加の復元を示す。
【図7】NM−F9におけるrhGM−CSFの発現を示す。
【図8】種々の分泌シグナルペプチドを使用した場合の、NM−F9細胞による、組換え発現された分泌性AGPAの発現を示す。
【図9】代謝改変による膜複合糖質の示差的シアル酸付加を示す。
【図10】代謝的相補による細胞表面上の膜タンパク質の示差的シアル酸付加を示す。
【図11】代謝的相補による分泌性rhGM−CSFの示差的シアル酸付加を示す。
【図12】TF1細胞上の示差的シアル酸付加rhGM−CSFの増殖活性を示す。
【図13】樹状細胞上の示差的シアル酸付加rhGM−CSFの増殖活性を示す。
【図14】rhGM−CSF濃度に対する増殖活性の依存性を示す。
【図15】rhGM−CSF増殖活性の抑制を示す。
【発明を実施するための形態】
【0012】
本発明において、「参照糖タンパク質」という用語は、本発明の糖タンパク質と同一又は類似のタンパク質配列を有する糖タンパク質、好ましくは、同じタンパク質配列を有する糖タンパク質を意味する。グリコシル化構造を欠くことも可能である。参照糖タンパク質は一定の活性を有し、天然由来又は組換え発現されたものでありうる。好ましい参照体は公知の組換え糖タンパク質である。活性、例えばin vitroにおける受容体媒介活性の改善をもたらす、先行技術の特定のタンパク質配列の突然変異の場合には、本発明のより高い活性の糖タンパク質は、この点に関してより高い活性であることが可能であり、及び/又は、前記活性はより良くなることなく別の生物活性、例えば免疫原性に関してより高い活性でありうる。これらの具体例においては、免疫原性におけるより高い活性は、哺乳動物、好ましくはヒトにおける使用に有利であり、したがって本発明における、より高い活性の糖タンパク質に匹敵する。当業者であれば、前臨床及び/又は臨床試験計画における適当な選択基準を選択することが可能である。
【0013】
本発明において、「発現細胞系」なる語は、当業者に公知の又は本明細書に記載の技術による、標的遺伝子の更なる組換え発現又はウイルス若しくは当業者に公知の適当な物質の感染を伴う又は伴わない、タンパク質、糖タンパク質、ウイルス又は他の生物物質の発現に使用しうる細胞又は細胞系を意味する。
【0014】
本発明において、「シアル酸の糖ヌクレオチド生合成経路における少なくとも1つの欠損を含有する発現細胞系」なる表現は、CMP−シアル酸の合成に関与する酵素に欠損を含有する発現細胞系を意味する。欠損は、対応する酵素活性が減少しているか又は完全に存在しないことを意味し、例えば、酵素の遺伝子、遺伝子の発現、酵素の活性、又はその機能に関連した酵素のバイオアベイラビリティのレベルでの種々の主要欠損によるものでありうる。酵素としては、例えばUDP−N−アセチルグルコサミン−2−エピメラーゼ、キナーゼ(例えば、N−アセチルマンノサミンキナーゼ、N−アセチルグルコサミンキナーゼ)、N−アセチルノイラミン酸−9−P(Neu5Ac−9−P)−シンテターゼ、Neu5Ac−9−P−ホスファターゼ及びCMP−Neu5Acシンテターゼが挙げられる。シアル酸の糖ヌクレオチド生合成経路における、より好ましい欠損は、エピメラーゼの突然変異であり、より一層好ましいのは、UDP−N−アセチルグルコサミン−2−エピメラーゼにおける、そのmRNA発現の欠損を引き起こす欠損である。
【0015】
該欠損は、標準的な条件下及び/又は無血清条件下での細胞のシアル酸付加の軽減又はシアル酸付加のほぼ完全な若しくは完全な欠如をもたらす。このシアル酸の軽減は、シアル酸前駆体添加物の添加により部分的又は完全に復元されうる。この欠損は、CMP−シアル酸の合成に関与する1以上の酵素の欠損に限定されるものではなく、シアル酸前駆体添加物の添加により該効果が部分的又は完全に復元されうるものである限り他の酵素の欠損でありうる。したがって、限定的なものではないが、一例としては、CMP−シアル酸輸送体としての、対応する糖輸送酵素が挙げられる。当業者であれば、どのような単一の欠損又は欠損の組合せが本発明に適しているかを判定することが可能であり、それらのいくつかは以下に更に詳しく説明されている。
【0016】
本発明において、「トランスフェクション」という用語は、ウイルス感染又はトランスフェクション方法(限定的なものではないが、リン酸カルシウム共沈法、エレクトロポレーション、DEAE−デキストラン又はカチオン性脂質試薬との複合体形成及びマイクロインジェクションが含まれる)により組換えタンパク質の発現のために細胞内へ遺伝物質を送達し、それにより発現細胞系の細胞内に核酸を導入するのに用いられうる当業者に公知の方法を意味する。
【0017】
本発明において、「糖タンパク質をコードする核酸」なる表現は、対象の哺乳動物糖タンパク質又はその活性断片及び/若しくは突然変異体(したがって、任意の糖タンパク質、好ましくは、ヒト由来の任意の糖タンパク質が使用されうる)をコードする核酸配列を意味する。哺乳動物糖タンパク質の具体例には、サイトカインのような分子及びそれらの受容体、例えば、腫瘍壊死因子TNF−α及びTNF−β;レニン;ヒト成長ホルモン及びウシ成長ホルモン;成長ホルモン放出因子;副甲状腺ホルモン;甲状腺刺激ホルモン;リポタンパク質;α−1−抗トリプシン;インスリンA鎖及びB鎖;性腺刺激ホルモン、例えば濾胞刺激ホルモン(FSH)、黄体形成ホルモン(LH)、甲状腺刺激ホルモン及びヒト絨毛性性腺刺激ホルモン(hCG);カルシトニン;グルカゴン;凝固因子、例えば第VIIC因子、第IX因子、第VII因子、組織因子及びフォンビルブラント因子;抗凝固因子、例えばプロテインC;心房性ナトリウム利尿因子;肺界面活性物質;プラスミノーゲン活性化因子、例えばウロキナーゼ、ヒト尿及び組織型プラスミノーゲン活性化因子;ボンベシン;トロンビン;造血増殖因子;エンケファリナーゼ;ヒトマクロファージ炎症性タンパク質;血清アルブミン、例えばヒト血清アルブミン;ミュラー抑制性物質;レラキシンA鎖及びB鎖;プロレラキシン;マウス性腺刺激ホルモン関連ペプチド;血管内皮増殖因子;ホルモン又は増殖因子に対する受容体;インテグリン;プロテインA及びD;リウマチ因子;神経栄養因子、例えば骨由来神経栄養因子、ニューロトロフィン3、4、5、6及び神経成長因子β;血小板由来増殖因子;繊維芽細胞増殖因子;表皮増殖因子;トランスフォーミング増殖因子、例えばTGFα及びTGFβ;インスリン様増殖因子I及びII;インスリン様増殖因子結合タンパク質;CDタンパク質、例えばCD3、CD4、CD8及びCD9;エリスロポエチン(EPO);骨誘導因子;イムノトキシン;骨形成タンパク質;インターフェロン、例えばインターフェロンα、β及びγ;コロニー刺激因子(CSF)、例えばM−CSF、GM−CSF及びG−CSF;インターロイキン(IL)、例えばIL−1〜IL−12;スーパーオキシドジスムターゼ;T細胞受容体;表面膜タンパク質;崩壊促進因子;抗体及びイムノアドヘシン;グリコホリンA;MUC1が含まれる。
【0018】
好ましい糖タンパク質は、グリコホリンA、EPO、G−CSF、GM−CSF、FSH、hCG、LH、インターフェロン、インターロイキン、抗体及び/又はそれらのフラグメントを含む群から選ばれる。
【0019】
本発明において、「糖タンパク質」という用語は、ウイルス、ウイルス粒子又はウイルスタンパク質としても定義される。
【0020】
本発明の好ましい実施形態においては、核酸は糖タンパク質の分泌性形態又はその断片をコードしている。好ましい実施形態においては、該分泌性形態は膜貫通ドメインを欠く。別の好ましい実施形態においては、該分泌性形態をコードする核酸は少なくとも1つの分泌シグナルを含む。最も好ましい実施形態においては、該分泌シグナルはGM−CSF由来である。他の実施形態及び詳細は以下で更に詳しく説明されている。
【0021】
本発明において、「シアル酸前駆体添加物」なる語は、糖タンパク質の部分的又は完全なシアル酸付加をもたらす、シアル酸の糖ヌクレオチド生合成経路における欠損を部分的又は完全に復元しうる任意の天然又は合成化合物を意味する。本発明の好ましい実施形態においては、種々の濃度のシアル酸前駆体添加物の添加は、種々のシアル酸付加度を有する糖タンパク質の一連の異なるシアル酸形態を与える。それにより、糖タンパク質のシアル酸付加形態は単一のグライコフォーム又は一群のグライコフォームから構成されうる。これは、単一のシアル酸前駆体添加物を使用することにより、又は2以上の前駆体添加物の組合せを使用することにより達成されうる。本発明の好ましい実施形態においては、シアル酸前駆体添加物として、シアル酸中間体及び/又はシアル酸含有糖タンパク質を使用する。別の好ましい実施形態においては、シアル酸前駆体添加物はManNAc、アセチル化ManNAc、パーアセチル化ManNAc又はフェツインである。別の好ましい実施形態においては、ManNAc、アセチル化ManNAc又はパーアセチル化ManNAcとフェツインとの組合せを使用する。
【0022】
好ましいのは、天然シアル酸修飾を有する糖タンパク質を与えるシアル酸前駆体添加物である。天然シアル酸修飾は、出発物質、中間体グライコフォーム又は最終グライコフォームのような、生物において見出されうる修飾である。さらに、天然糖タンパク質、グリコシル化形態(これらは、生物体内に天然で存在するグライコフォームに可能な限り近い)が好ましい。グライコフォーム、例えば、ヒトにおいて見出される糖鎖又はヒト由来の任意の生物学的物質に類似した、シアル酸を有する糖鎖を有する、ヒト糖タンパク質のグライコフォームが使用されるであろう。それらの好ましい形態は、生物において見出されない非天然側鎖により修飾されたシアル酸を含有しない。例えば、糖タンパク質又は糖脂質内に組込まれるシアル酸は、化学合成により導入されうる、置換された若しくは欠失した若しくは付加された若しくは伸長した若しくは短縮した側鎖、及び/又は生物において見出されない、シアル酸への新規鎖間連結(interlinkage)を含まない。
【0023】
本発明の好ましい形態においては、いずれの細胞生成物のグリコシル化も、非天然シアル酸修飾を与える化学的に操作された前駆体又は代謝産物によって変化させたり又は修飾するのではなく、天然グリコシル化、好ましくはシアル酸付加の度合を最適化することにより変化させたり又は修飾する。化学修飾されたシアル酸代謝産物は、天然グリコシル化に影響を及ぼさない様態でのみ使用されうる。例えば、アセチル化ManNAc又はパーアセチル化ManNAcは天然N−アセチルマンノサミン(ManNAc)より遥かに容易に細胞により取り込まれるため、アセチル化ManNAc又はパーアセチル化ManNAcを添加した細胞培養培地としてManNAcは置換されうる。細胞内で、追加的なアセチル残基が切り離され、CMP−シアル酸の生合成のためにManNAcがプロセシングされる。したがって、アセチル化ManNAcで処理された細胞により産生される糖タンパク質は、合成により修飾された炭水化物(例えば、合成によりアセチル化されたシアル酸)を何ら示さない。
【0024】
本発明において、「異なるシアル酸付加形態」なる表現は、少なくとも1つのシアル酸が別のグライコフォームとは異なる糖タンパク質のグライコフォームを意味する。シアル酸における相違は、シアル酸の量、及び/又は該分子の残部上の少なくとも1つのシアル酸の位置におけるものでありうる。個々のシアル酸付加形態は、糖タンパク質の単一のグライコフォーム又はいくつかのグライコフォームの混合物から構成されることが可能であり、それにより、異なるシアル酸付加形態はお互いに、少なくとも1つのグライコフォームにおいて及び/又はグライコフォームの相対組成において異なる。
【0025】
少なくとも1つのシアル酸付加形態のシアル酸付加度は、当業者に公知の生化学的及び/又は化学的方法により測定することが可能である。例えば、迅速な方法は、シアル酸特異的抗体又はレクチン、例えばSNA及びMAAで、あるいはシアル酸を含まないグリカンを認識する抗体又はレクチン、例えばA78−G/A7及びNemod−TF1又はレクチンPNAで、細胞又は糖タンパク質を染色するものである。染色された細胞はフローサイトメトリーにより分析し、染色された糖タンパク質は、糖タンパク質が精製されている場合には通常のELISAにより、あるいは分析対象の糖タンパク質を捕捉するために糖タンパク質特異的抗体を使用するサンドイッチELISAにより分析することが可能である。当業者に公知のこれらのすべての方法は、細胞又は糖タンパク質の示差的シアル酸付加のモニターを可能にする。したがって、結合したシアル酸特異的抗体又はレクチンの量は、細胞又は糖タンパク質に結合したシアル酸の量と相関し、一方、シアル酸を含まないグリカンを認識する結合抗体又はレクチンの量は、細胞又は糖タンパク質に結合したシアル酸の量と逆相関する。シアル酸含量を定量するためのもう1つの方法は、ノイラミニダーゼを使用することにより酵素的に、あるいは酸加水分解により化学的に、糖タンパク質に結合したすべてのシアル酸を遊離させるものである。ついでサンプル中のシアル酸の濃度を、1,2−ジアミノ−4,5−メチレンジオキシベンゼン(DMB)でのシアル酸の蛍光測定染色、及び蛍光測定検出器を使用するHPLC分析により測定する。あるいは、遊離したシアル酸の量を、チオバルビツレート及びβ−ホルミルピルベートの存在下、549nmで比色検出されうるピンク色色素にシアル酸を変換することにより定量する。N−又はO−グリカンに結合したシアル酸の数に関するデータを得るためには、全N−又はO−グリカンを酵素的又は化学的に遊離させる。アンテナ性(antennarity)に関する遊離グリカン(例えば、HPAEC−PADにより)、結合シアル酸の数(例えば、陰イオン交換クロマトグラフィーと組合されたHPAEC−PADにより)、単糖の組成(例えば、グリカンの酸加水分解の後のHPAEC−PADにより)及び/又は厳密な構造(例えば、質量分析)を分析するためのいくつかの方法が当業者に利用可能である。
【0026】
本発明における定義において、「活性」は、生物学的環境中で分子により発揮される機能又は一連の機能である。一般には、本発明の方法により製造される糖タンパク質は、先行技術により製造される糖タンパク質より活性が高く及び/又はより有効な生物学的産物である。したがって、本発明は、ヒト高活性糖タンパク質、及び該糖タンパク質を含む診断、治療又は他の生物学的系及び試験におけるその使用に関する。本発明における定義においては、「高活性糖タンパク質」なる語、その内容の均等物及びその文法的均等物は、選択された用途に関して、改善された又は最適の活性として理解されるべきであり、したがって、該活性は極限値に近似されることが可能であり、例えば最小化又は最大化されることが可能であり、あるいは、先行技術により製造された対応糖タンパク質及び/又は本発明の示差的シアル酸付加により製造された別のシアル酸付加形態より高い又はより低い活性を表す中間値に設定されることが可能であろう。本発明における改善された又はより高い活性はまた、その生物学的及び/又は薬学的意味において好ましい活性を意味する。例えば、糖タンパク質の生物活性は、有害な生物学的影響を軽減することにより(例えば、有害な免疫作用の刺激の低減又は免疫原性の低減により)或る程度は増強されうるであろう。本発明の方法は、最適化された血清半減期、薬物動態、安定性、抗原性及び/又は免疫原性を有する糖タンパク質を製造するために有効に使用されうる。
【0027】
本発明において、「異なるシアル酸付加形態の活性の測定」なる表現は、糖タンパク質の活性を測定しうる適当なバイオアッセイにおける活性の測定を意味する。当業者であれば、適当なバイオアッセイを決定し、適当なバイオアッセイを構成することが可能である。本発明においては、そのようなバイオアッセイとしては、例えば、生物学的in vitroアッセイ、例えば細胞若しくは分子又は混合アッセイ、例えば増殖アッセイ、アポトーシスアッセイ、細胞接着アッセイ、シグナル伝達アッセイ、遊走アッセイ、細胞毒性アッセイ、食作用アッセイ、細胞溶解アッセイ及び結合アッセイが挙げられる。そのようなバイオアッセイにはまた、動物モデル又はヒトを使用するin vivoアッセイ、例えば生体内分布、薬物動態、薬動力学試験、血清半減期試験、バイオアベイラビリティに関する試験、有効性試験、局在性試験、疾患の治療及び予防試験(臨床研究を含む)が含まれる。そのようなバイオアッセイには、化学的、物理的、物理化学的、生物物理学的及び生化学的試験、例えば温度、せん断応力、圧力、pH、複合体形成(コンジュゲーション)などに対する安定性も含まれる。そのようなバイオアッセイには、糖タンパク質の臨床的使用に関する該糖タンパク質の特性を改善するための免疫原性及び/又は抗原性に関する試験も含まれる。当業者であれば、糖タンパク質及びそのシアル酸付加形態の活性又は記載されている活性の組合せを決定することが可能である。
【0028】
好ましい実施形態においては、糖タンパク質のより高い活性は、少なくとも1つのin vitroモデルにおける、り高い活性、及び/又は少なくとも1つのin vivoモデルにおけるより高い活性、及び/又はより高い安定性、及び/又はより長い血清半減期、及び/又はより長いバイオアベイラビリティ、及び/又は免疫原性の改善、及び/又は抗原性の改善(少なくとも1つのバイオアッセイにより測定される)を特徴とする。全活性における改善(これは本明細書中では、より高い活性とも称される)は、例えば、ヒト又は対応生物において使用される場合の該生成物の、より低い投与量、より長い投与時間間隔、より低い副作用、及び全く無い又はより低い毒性をもたらして、大きく改善された医薬を与えうる。
【0029】
本発明において、「より高い活性を有するシアル酸付加形態の選択」という用語は、バイオアッセイの少なくとも1つにおける比較において、より高い活性を有する糖タンパク質のシアル酸付加形態の選択を意味する。この比較は、本発明の方法により発現される少なくとも1つのシアル酸付加形態を先行技術のグリコシル化形態の糖タンパク質と比較することにより、あるいは本発明の方法により発現される少なくとも2つのシアル酸付加形態を比較することにより行いうる。好ましくは、相当に最適化されたグライコフォームを達成し、同定し、製造するためには、高活性糖タンパク質は3以上の異なるシアル酸付加形態から選ばれる。当業者であれば、どのシアル酸付加形態が、特定のバイオアッセイに関して、より高い活性を有するかを決定することが可能である。好ましい実施形態においては、より高い活性を有するシアル酸付加形態を選択するためには、バイオアッセイの組合せの活性を考慮する。その場合、それぞれのバイオアッセイが、より高い活性を示す必要があるわけではなく、個々の糖タンパク質の用途及び特徴に応じて、いくつかの好ましい生物学的作用が、それほど好ましくはない他のものを補うことが可能であり、本発明においては尚も全体的にはより高い活性のグライコフォームをもたらしうる。例えば、あるグライコフォームは、細胞に対してその受容体に結合することにより、より遥かに高い活性をもたらして、例えば増殖の誘導のような副次的効果を誘導しうるが、若干減少した血清半減期を示す。したがって、組合せとしては、該受容体をより惹起するより高い活性は、全体的な生物活性においてバイオアベイラビリティが短くなることを補う。もう1つの例においては、より短い半減期、及び受容体惹起をもたらす、より高い活性は、どちらも有利である。さらにもう1つの例においては、in vivoでの活性は改善されないが、in vitroでの安定性は糖タンパク質の製造及び貯蔵を改善する。さらにもう1つの例においては、長い半減期の一方で、より低い活性が必要とされる。したがって、実際の有利なシアル酸付加形態を、特定の糖タンパク質に基づいて、その後の使用に関して決定する必要がある。本発明は、この統合された系において、これを初めて可能にするものである。このすべての態様は、決定すべき単一の糖タンパク質に左右され、これらは大きく異なり、予想できず、本発明の後述する方法を必要とする。本発明を使用することにより、当業者は、適当なバイオアッセイ及び/又はバイオアッセイの組合せ、並びに前記の点に関して糖タンパク質の所望の用途のためのより高い活性を有する糖タンパク質のシアル酸付加形態を選択することが可能であり、したがって、標準的な方法により製造されうるグリコシル化形態と比べて糖タンパク質を改善することが可能である。本発明を使用することにより、当業者は、該糖タンパク質のより高い活性レベルと相関する、シアル酸前駆体添加物又はシアル酸前駆体添加物の組合せの濃度を決定することが可能である。
【0030】
本発明において、示差的シアル酸付加により発現させバイオアッセイにおいて比較する複数の異なるシアル酸形態という表現は、バイオアッセイにおいて試験した場合に、先行技術により得られる少なくとも1つの異なるシアル酸付加又はグリコシル化形態と比べて高い活性を示す、本発明の方法により発現される1つのシアル酸形態をも意味する。
【0031】
前記のすべての定義及び説明は、前記及び後記の糖タンパク質及び方法にも有効である。
【0032】
驚くべきことに、本発明の組換え発現された糖タンパク質は、シアル酸前駆体添加物の特定の濃度に対応する特定の度合でシアル酸付加された場合に、最高の生物活性を示すことを見出した。最高の生物活性と相関するこのシアル酸付加度は、分析する各糖タンパク質によって異なり、予測可能ではなく、最高のシアル酸付加度が必ずしも達成可能なわけではない。本発明に従って糖タンパク質のシアル酸付加度を同定するためには、細胞欠損によってシアル酸付加の復元及び種々のシアル酸付加度の達成が可能となるようにシアル酸付加能が欠損した細胞を使用及び/又は作製する。シアル酸付加の復元を達成するための1つの方法は、シアル酸生合成経路のシアル酸前駆体添加物を該細胞の培地に添加することによるものである。該欠損細胞及び少なくとも1つのシアル酸前駆体添加物を使用して、該培地に加えるシアル酸前駆体添加物の量を変化させることにより、該細胞により産生される本発明の任意の糖タンパク質のシアル酸付加度を示差的に制御することが可能である。少なくとも2つの異なるシアル酸付加形態が達成されうる。僅か2つの異なる濃度の前記添加物を使用する場合には、異なるシアル酸付加形態の作製が保証されるよう、濃度勾配は十分大きくなければならない。好ましくは、最高値が方法の特徴(例えば、粘度、溶解度、aw値など)により定められる高濃度及び低濃度が選ばれる。示差的にシアル酸付加された糖タンパク質の少なくとも2つの形態、好ましくは複数の形態(異なるシアル酸形態)を単離し、グリコシル化、好ましくはシアル酸付加、活性、血清半減期、薬物動態、薬動力学、in vivo生物活性、抗原性及び免疫原性のような特性に関して分析することが可能である。そのようにして、該糖タンパク質の、その活性又は任意の他の特性のための最適なシアル酸付加度は、前記の基準の1つ又は組合せにより同定され、その最適シアル酸付加糖タンパク質の製造のための条件は、生産細胞として上記欠損細胞を使用する製造プロセスに移されうる。
【0033】
本発明はまた、高活性糖タンパク質の製造のための特別な方法及び高活性糖タンパク質そのものを教示する。これらの特別な場合には、シアル酸前駆体添加物の最適濃度が他の技術により決定され又は当業者の論理的思考から推定されうる場合には(例えば、後記実施例に記載のとおり)、個々のシアル酸付加形態は、それを、異なる濃度のシアル酸前駆体添加物の対応発現細胞系内での発現により得られる第2の形態と比較することを要することなく、製造されうる。これらの特別な場合も、高活性糖タンパク質及び本発明の方法において、明らかに本発明に包含される。これらの場合、本発明の基本的な発明原理は尚も、本方法及び得られる高活性糖タンパク質の中心的かつ決定的な部分である。これに関しては、前記及び後記の他の説明も参照されたい。一例としては、シアル酸付加を全く又は最小限度しか伴わないGPAが挙げられ、これは後記の実施例の1つにおいて更に詳しく説明されており、この場合の目的は、初めから、最小限度のシアル酸付加及び最大限度のトムセン−フリーデンライヒ(Thomsen-Friedenreich)を伴う形態の作製でありうる。したがって、シアル酸前駆体添加物を加えることなく、好ましくは、培地内にいずれのシアロ糖タンパク質も存在することなく、最低限度のシアル酸付加を伴う形態のみを作製することで十分でありうる。更なる使用のためのこの形態の適合性は、それを、少なくとも1つの他のシアル酸付加形態と比較することにより判定されうるが、この代わりに、ヒト由来のシアル酸付加又は低シアル酸付加GPAとも比較しうる。もう1つの例は、最高シアル酸付加が望ましいことが判明しており、最大限度のシアル酸前駆体添加物を使用することで十分でありうる場合である。しかし、後者の場合には、ほとんどすべての場合において、本明細書中の種々のシアル酸付加形態を作製することによりシアル酸前駆体添加物の組成及び濃度を最適化することにより最高シアル酸付加を最適化することが有利である。この特別な場合には、単一のバイオアッセイとしてのシアル酸度の測定のための技術を用いることで十分かもしれないが、他のバイオアッセイとの組合せが好ましい。
【0034】
新規技術は、新規糖タンパク質の作製及び種々の糖タンパク質の改善(改良バイオジェネリックス(biogenerics)の作製を含む)のための高い潜在性を保有する。
【0035】
以下において、シアル酸の糖ヌクレオチド生合成経路における欠損を更に詳しく説明する。本発明においては、細胞のシアル酸付加能の低下を引き起こす欠損(好ましくはシアル酸付加能の完全な消失)を有し該欠損が復元されうることにより示差的制御シアル酸付加を可能にする欠損を有する発現細胞系により、高活性糖タンパク質を製造する。「シアル酸の糖ヌクレオチド生合成経路における欠損」とも称される該細胞の分子的欠損は、シアル酸の代謝に関与するタンパク質(例えば、糖輸送における例えばCMP−シアル酸輸送体、又はCMP−シアル酸の生合成における例えばキナーゼ、デヒドロゲナーゼ、ホスファターゼ、シンテターゼ、トランスケトラーゼ、トランスアルドラーゼ、イソメラーゼ、トランスフェラーゼ及びエピメラーゼ)の機能喪失(loss-of-function)でありうる。また、シアル酸付加の軽減又は欠如を伴うシアル酸付加不能細胞を与える分子的欠損は、シアル酸の代謝に関与するタンパク質(例えば、炭水化物からシアル酸を遊離させる酵素、例えば、ノイラミニダーゼ)の機能獲得(gain-of-function)でありうる。好ましい標的は、CMP−シアル酸の輸送に寄与するタンパク質、及びシアル酸生合成経路における律速段階を触媒する酵素である。より好ましくは、該欠損はエピメラーゼ、最も好ましくはUDP−GlcNAc−2−エピメラーゼに関するものである。
【0036】
先行技術は、N−アセチルマンノサミン(ManNAc)がCMP−シアル酸の生合成において重要な役割を果たすことを教示している。ManNAcは、シアル酸の細胞生合成における律速段階を代表するUDP−GlcNAc−2−エピメラーゼの作用により、UDP−N−アセチルグルコサミン(UDP−GlcNAc)から形成される。ついでManNAcは特異的キナーゼ(例えば、N−アセチルマンノサミンキナーゼ、N−アセチルグルコサミンキナーゼ)によりリン酸化される。ManNAc−6−PはNeu5Ac−9−P−シンテターゼによりホスホエノールピルビン酸と縮合してN−アセチルノイラミン酸−9−P(NeuAc−9−P)となり、ついで該ホスフェートはNeu5Ac−9−P−ホスファターゼにより遊離され、Neu5AcはCMP−Neu5Acシンテターゼにより活性化される。ついでCMP−Neu5Acは特異的輸送体(例えば、CMP−シアル酸輸送体)により細胞のゴルジ区画内へ輸送され、その部位において、それは、シアル酸を糖タンパク質又は糖脂質の末端ガラクトース(Gal)−、N−アセチルガラクトサミン(GalNAc)−又はN−アセチルグルコサミン(GlcNAc)−残基へ転移させる種々のシアリルトランスフェラーゼ、例えばST6GlcNAcシアリルトランスフェラーゼ、ST6GalNAc−I〜VI、ST3Gal−I〜VI及びST6Gal−Iのための供与体として働く。
【0037】
シアル酸生合成の各酵素、例えばUDP−GlcNAc−2−エピメラーゼ、キナーゼ、Neu5Ac−9−P−シンテターゼ、Neu5Ac−9−P−ホスファターゼ及びCMP−Neu5Acシンテターゼ、又はゴルジ区画内への輸送に必要な各タンパク質は、欠損細胞に欠損酵素の産物を供給することにより該欠損が復元されうるように、細胞のシアル酸付加能を改変するよう、好ましくは軽減するよう、より好ましくは完全に遮断されるよう影響を受けることが可能である。細胞のシアル酸付加能は、シアリルトランスフェラーゼにより触媒される炭水化物へのシアル酸の転移における変化によっても影響されうる。
【0038】
該細胞欠損を生じさせ、遺伝的、転写、翻訳又はタンパク質レベルでタンパク質の合成及び機能を標的化するよう特異的に該細胞欠損を作製するには、多数の選択肢が存在する。
【0039】
特定のタンパク質をコードする遺伝子における突然変異は、しばしば、そのタンパク質の機能喪失を引き起こし、あるいは、時には、機能を獲得するタンパク質を与える。例えば、1個若しくは2個のヌクレオチドの置換により又は(1+n)個若しくは(2+n)個のヌクレオチド(ここで、nは3の倍数である正の整数)の導入若しくは欠失により引き起こされうるフレームシフトにより、遺伝子のコード領域内に停止コドンを与える突然変異が挙げられる。多数の他の「機能喪失」及び「機能獲得」遺伝子突然変異が当業者に公知である。これらの遺伝子突然変異は遺伝子の転写にも影響を及ぼしうる。化学的突然変異原、例えばメタンスルホン酸エチル(EMS)又は物理的突然変異原、例えばUV光により遺伝的突然変異をランダムに誘発するための多数の方法が当業者に公知である。また、現代の分子生物学的技術によって、当業者は突然変異により遺伝子を特異的に標的化することが可能である。これらの技術を用いて、当業者は特定の遺伝子をノックアウトしたり、あるいは新たな遺伝子を導入して機能獲得突然変異体を作製することが可能である。
【0040】
転写レベルでは、特定のタンパク質をコードするmRNAの合成が制御される。遺伝子転写における欠損又は変化は、特定の酵素のmRNA合成のスイッチをオフ又はオンにすることができる。例えば、シアル酸の生合成に関与する酵素をコードするDNAのメチル化、又はDNA結合タンパク質、例えばヒストン、転写因子若しくは転写装置のタンパク質の修飾、例えばアセチル化は転写を妨げたり、あるいは特定の遺伝子の転写のスイッチをオンにすることが可能であろう。前記の及び当業者に公知の遺伝学的及び分子生物学的方法による突然変異の誘発は、任意の遺伝子の転写効率に影響を及ぼすための1つの選択肢である。NM−F9細胞内のUDP−GlcNAc−2−エピメラーゼmRNAの欠如(その欠損はEMS突然変異原での野生型細胞の処理により作製される)は、機能喪失欠損を作製するための転写の影響の一例である。
【0041】
翻訳レベルでは、特定の酵素の、合成されたmRNAを標的化して、好ましくは、リボソームタンパク質の生合成を妨げ又は軽減する。具体例としては、共に特異的mRNA分子を標的化するアンチセンス分子又は小さな干渉性RNA分子を使用する翻訳の阻止及び/又はRNaseによる特定のmRNAの分解が挙げられる。どちらの技術も当業者に公知である。
【0042】
タンパク質レベルでは、酵素活性の欠如は遺伝子突然変異により引き起こされうるであろう。可能な突然変異には、少なくとも1つのアミノ酸の置換、欠失及び/又は修飾が含まれる。該突然変異は、新たな及び非天然構造が基質の結合及び/又はその変換を許容しないタンパク質の構造的再折りたたみによる、あるいは基質との相互作用に寄与する活性中心内の残基を変化させることによる、酵素の触媒特性に関するものでありうる。そのような突然変異を作製するための技術は前記で説明されている。
【0043】
CMP−シアル酸の生合成又は輸送は特定のインヒビターによっても影響されうる。これらのインヒビターは、CMP−シアル酸生合成に関与する酵素若しくはタンパク質の活性を遮断するシアル酸類似体、又はCMP−シアル酸の転移に必要ないずれの輸送体の活性を遮断するシアル酸類似体でありうる。これらのインヒビターは、酵素又はタンパク質の調節中心及び/又は触媒中心に結合して、基質の結合及び/又はその変換若しくは輸送を妨げうる。インヒビターは、シアル酸類似体、小有機分子、ペプチドリガンド、抗体及びそのフラグメント、DNAアプタマー、RNAアプタマー、スピーゲルマー(Spiegelmer)などを含む群から選ばれる高い結合アフィニティーを有する分子である。
【0044】
Nemod Biotherapeutics GmbH & Co. KG, Robert-Rossle-Str. 10, 13125 Berlin(ドイツ)により2003年8月14日付けでBraunschweig(ドイツ)の“DSMZ-Deutsche Sammlung von Mikroorganismen und Zellkulturen GmbH”にDSM ACC2606(NM−F9)及びDSM ACC2605(NM−D4)として寄託されているNM−F9及び/又はNM−D4細胞は、本発明の高活性糖タンパク質の発現のために及び本発明の後記方法における使用のために理想的に適合化された発現細胞系であることが判明している糖改変細胞である。なぜなら、NM−F9及びNM−D4細胞は、代謝的相補により復元されうるようにシアル酸付加能を喪失しているからである。したがって、本発明においては、NM−F9及び/又はNM−D4細胞は、本発明の高活性糖タンパク質及び後記の方法のための、シアル酸の糖ヌクレオチド生合成経路における欠損を有する最も好ましい細胞系である。
【0045】
NM−F9及びNM−D4細胞におけるシアル酸付加の細胞欠損はUDP−GlcNAc−2−エピメラーゼ活性の欠如により引き起こされ、それにより、シアル酸付加は、該エピメラーゼにより触媒される酵素反応の上流のシアル酸生合成の細胞産物での培地補足(代謝的相補とも称される)により復元されうる。そのようなシアル酸中間体はManNAc、ManNAc−6−P、NeuAc−9−P、Neu5Ac及びCMP−Neu5Acでありうる。ManNAcが好ましい。驚くべきことに、細胞シアル酸付加度は、細胞培養培地に加えるManNAcの量により制御することが可能であった。また、細胞シアル酸付加度だけでなく、より一層驚くべきことに、細胞から分泌又は遊離される糖タンパク質の度合も、そのように調節されうる。驚くべきことに、たいていは培地へのウシ胎仔血清(FCS)の添加によりもたらされる、細胞培養培地内のシアル酸付加血清タンパク質の存在は、細胞又は分泌性糖タンパク質のシアル酸付加度に影響を及ぼす。これは妥当だと思われる。なぜなら、該細胞は、CMP−シアル酸を産生するためのそれ自体の生合成経路を要することなく、ある程度は、de novoグリコシル化のために、取り込まれた糖タンパク質からのシアル酸を利用するようであり、あるいは異化経路によりシアル酸をシアル酸の公知又は未知前駆体生合成経路へと導入するためである。したがって、該細胞をFCS非含有培地内で又は更にはシアル酸付加タンパク質非含有培地内で培養する場合には、非常に低い度合のシアル酸付加が得られる。シアル酸付加度は常に、FCSの存在下では、FCSを含有しない同じ細胞培養条件下より高い。したがって、与えられた細胞系において最高のシアル酸付加度を達成するためには、FCS、又はより好ましくはシアル酸付加糖タンパク質、又は最も好ましくはシアル酸付加糖タンパク質フェツインを培地に加える。このようにして、NM−F9細胞又は分泌性糖タンパク質上のシアル酸付加度は、元のNM−wt細胞のシアル酸付加に比べて20%未満、好ましくは10%未満、最も好ましくはほぼ0%のシアル酸付加から、エピメラーゼ欠損を有さない元のNM−wt細胞シアル酸付加と比べてシアル酸付加を完了する(100%)シアル酸付加まで制御されうるであろう。
【0046】
この範囲のなかで、シアル酸付加度を示差的に制御することが可能であり、これは、一定の糖中間体、例えば0〜200mM、好ましくは0〜140mM、より好ましくは0〜90mMの濃度範囲のManNAcを培地に添加することにより行うことが可能である。非常に高いシアル酸付加度を達成するためには、シアロ糖タンパク質を培地に加えることが可能である。異なるシアル酸付加度を有する各糖タンパク質を単離し、異なるシアル酸付加度を有する同じ糖タンパク質とは別々に分析することが可能である。したがって、本明細書に記載のとおり、あるシアル酸付加度は、単一のグライコフォーム又は複数のグライコフォームの組成物を含みうるシアル酸付加形態に対応する。
【0047】
したがって、本発明は、後記で更に詳しく説明するとおり、糖タンパク質の示差的シアル酸付加のための方法又はプロセスを提供する。
【0048】
前記のとおり、異なるシアル酸付加度を有する各糖タンパク質の活性を分析することにより、シアル酸前駆体添加物の最適化濃度を決定することが可能である。分析対象の活性は、目的の糖タンパク質に左右される。例えば、糖タンパク質がサイトカインである場合には、免疫細胞(好ましくは、そのサイトカインにより刺激されることが公知の細胞、より好ましくは、該サイトカインの活性を測定するための標準的なアッセイにおいて使用される細胞)を刺激する該糖タンパク質の能力を分析することになろう。もう1つの例において、糖タンパク質が増殖因子である場合には、細胞増殖(好ましくは、その増殖因子により刺激されることが公知の細胞の細胞増殖、より好ましくは、該増殖因子の増殖活性を測定するための標準的なアッセイにおいて使用される細胞の増殖細胞)を誘導する該糖タンパク質の能力を分析することになろう。もう1つの例においては、血清半減期が対象となる。なぜなら、それは、治療用に適用される糖タンパク質の生物活性に著しく影響を及ぼすからである。本発明は、種々のシアル酸付加度を特徴とする1つの糖タンパク質の種々の形態の発現及び単離を可能にするものであり、該糖タンパク質の任意の活性を分析するために任意のアッセイを適用することにより、該糖タンパク質の活性のための最適なシアル酸付加度を決定することが可能である。
【0049】
以下においては、糖タンパク質及び高活性糖タンパク質を更に詳しく説明する。糖タンパク質の具体例は前記に挙げられている。前記糖タンパク質の多くは「サイトカイン」に属し、サイトカインは本明細書においては、免疫系の細胞において見出されるホルモンの一般的クラスを意味し、リンホカイン及びモノカインの両方並びにその他を含む。この定義は、局所的に作用し血中に循環せず、本発明に従い使用される場合に個体の免疫応答の改変を引き起こすホルモンを含む(これらに限定されるものではない)と意図される。他の適当な免疫調節性サイトカインの具体例には、限定的なものではないがインターフェロン(例えば、IFN−α、IFN−β及びIFN−γ)、インターロイキン(例えば、IL−1、IL−2、IL−3、IL−4、IL−5、IL−6、IL−7、IL−8、IL−9、IL−10及びIL−12)、腫瘍壊死因子(例えば、TNFα及びTNFβ)、エリスロポエチン(EPO)、FLT−3リガンド、マクロファージコロニー刺激因子(M−CSF)、顆粒球コロニー刺激因子(G−CSF)、顆粒球−マクロファージコロニー刺激因子(GM−CSF)、CD2及びICAMが含まれる。エリスロポエチンに関しては、この分子は前駆細胞を赤血球へ成熟させると考えられている。一方、トロンボポエチンは前駆細胞を血小板経路に沿って誘導すると考えられている。CSFは、骨髄内に見出される前駆細胞を成熟血液細胞の特定の型へ分化するように誘導するリンホイシンのファミリーを意味する。前駆細胞から生じる成熟血液細胞の個々の型は存在するCSFの型よって異なる。同様に、顆粒球−マクロファージコロニーの形成はGM−CSFの存在に左右される。また、IL−2、GM−CSF、TNF−αなどのヒト形態に対して実質的な相同性を有する他の哺乳動物のサイトカインは、免疫系に対して同様の活性を示すことが証明されている場合には、本発明において有用であろう。同様に、いずれかの特定のサイトカインに実質的に類似しているがタンパク質配列の比較的わずかな変化を有するタンパク質も本発明において有用であろう。タンパク質分子の機能的能力を妨げることなくタンパク質配列内のいくつかのわずかな変化が可能でありうることはよく知られており、したがって、本発明においてサイトカインとして機能するが現在公知の配列とは若干異なるタンパク質が製造されうる。最後に、本出願における「サイトカイン」なる語の単数形又は複数形の使用は決定的なものではなく、本発明及び特許請求の範囲の解釈を限定するものではない。接着若しくはアクセサリー分子又はそれらの組合せは単独で又はサイトカインと組合せて使用されうる。
【0050】
好ましい糖タンパク質は、グリコホリンA、EPO、G−CSF、GM−CSF、FSH、hCG、LH、インターフェロン、インターロイキン、抗体及び/又はそのフラグメントを含む群から選ばれる。
【0051】
本発明においては、高活性糖タンパク質は内因性タンパク質、好ましくは、発現細胞系において組換え手法により製造されるタンパク質でありうる。したがって、目的の糖タンパク質をコードするcDNA又は遺伝子を分子生物学的方法により発現ベクター内にクローニングし、発現細胞系内で発現させる。この発現は、好ましくは、安定発現である。すべてのその技術は当業者に公知である。多数の発現ベクターが、当業者に公知の哺乳動物細胞における発現に利用可能であり適している。好ましい実施形態においては、糖タンパク質は細胞により分泌される。したがって、発現ベクターは、糖タンパク質の分泌を促進するように構築される。分泌性糖タンパク質は可溶性糖タンパク質、又は膜貫通ドメインが欠失した膜タンパク質でありうる。好ましい実施形態においては、分泌シグナルペプチドをコードする核酸を、糖タンパク質をコードする核酸内に含ませ、これは、該タンパク質の分泌を促進及び/又は増強する。それらのシグナルペプチド及びそれらのコード核酸は当業者に公知である。本発明の好ましい実施形態においては、シグナルペプチドの核酸は配列番号1の配列又は該配列の突然変異、欠失、挿入及び伸長並びに該配列を含有する融合タンパク質を有する。
【0052】
本発明においては、NM−F9細胞内での2つの高活性糖タンパク質、すなわち可溶性タンパク質としての顆粒球マクロファージコロニー刺激因子(GM−CSF)及び膜タンパク質としてのグリコホリンA(GPA)の組換え発現が実施例として記載されている。
【0053】
GM−CSFは、先天性及び適応免疫系の活性化において重要な役割を果たす、骨髄細胞の強力な種特異的増殖因子である。該因子は顆粒球、マクロファージ、好酸球及びそれらの前駆細胞の増殖、発生、分化及び活性化を刺激する。さらに、GM−CSFは、体液性及び細胞性免疫応答にとって中心的な樹状前駆細胞の分化及び成熟に決定的に重要である。赤芽球及び巨核球前駆細胞の増殖におけるエリスロポエチン(EPO)との相乗作用、並びに種々の他のサイトカイン(例えば、IL−1、IL−3、IL−4、G−CSF)との相乗作用が観察される。GM−CSFは好中球、好酸球及びマクロファージの殺微生物活性、酸化的代謝、食作用活性及び細胞傷害性を増強し、好塩基球からのヒスタミン及びロイコトリエンC4の遊離を誘導する。骨髄移植、好中球減少症(特に化学療法及び/又は放射線療法後)並びに化学療法後の感染性疾患及び出血に関する血球減少症の患者を治療する一方で標準的な療法として適用されるヒトGM−CSFのいくつかの臨床的適応が存在する。さらに、それは回収のための幹細胞の末梢プールを増殖させ、急性骨髄球様白血病及び骨髄形成異常症候群を引き起こす白血病細胞の、化学療法中の細胞周期特異的薬に対する感受性を誘導する。組換えヒトGM−CSFは種々の疾患における造血系の再構成を支持する。また、いくつかの実験的療法が実施されており、例えば、照射の致死効果を無効にし造血を回復させる、癌治療におけるGM−CSF/放射線療法の併用、並びに細胞毒性薬(例えば、癌化学療法、HIV)に対する許容度を増強し、より高い投与量及び有意な罹病率減少を可能にするGM−CSF/化学療法の併用が挙げられる。rhGM−CSFは癌及び感染性疾患に対する細胞ワクチン及び癌ワクチンのための有望なアジュバントであり、腫瘍及び感染性疾患に対する、樹状細胞に基づくワクチンの重要成分である。それは養子T細胞移入にも関与している。一般には、rhGM−CSFは先天性免疫系を増強し刺激するであろう。
【0054】
本発明のヒトGM−CSFは、より高い特異的活性を有するため、先行技術により作製されたGM−CSFと比較して有利である。この増強された活性は、用量当たり患者当たりの、より少ない物質の使用、及び/又はより少ない投与、及び/又は望ましくない副作用(例えば、組換えGM−CSFの治療的使用中に観察されている毛細管漏出症候群)の軽減若しくは欠如、及び/又はより低い製造コストを可能にする。
【0055】
本発明においては、rhGM−CSFをUDP−GlcNAc−2−エピメラーゼ欠損細胞、例えばNM−F9において組換え発現させ、ほとんど存在しないシアル酸付加から最大限度のシアル酸付加までの種々のシアル酸付加度を有する種々の形態のrhGM−CSFを、漸増量のシアル酸代謝産物、好ましくはManNAc、例えば0〜90mMのManNAc、90mM ManNAcに加えて1及び3mg/mlのシアロ糖タンパク質フェツインの存在下での代謝的相補により製造した。rhGM−CSFの種々のシアロ形態の活性を分析するために、細胞増殖(好ましくは、TF−1細胞及びヒト単球樹状細胞系NemodDC(www.nemod.com))を刺激する能力を測定する標準的なアッセイを選択した。驚くべきことに、rhGM−CSFの細胞増殖刺激活性はrhGM−CSFのシアル酸付加度に強く左右され、高度にシアル酸付加されているが最大限度にはシアル酸付加されていないrhGM−CSFの1つのシアロ形態が最高活性を示す。商業的に入手可能なrhGM−CSF、好ましくは、細菌及び酵母において発現されるLeukomax(登録商標)及びLeukine(登録商標)の細胞増殖活性を同じアッセイで試験したところ、NM−F9細胞内で発現される最も活性なシアロ形態より低いことが判明した。Leukomax(登録商標)又はLeukine(登録商標)の最高活性を達成するためには、対応in vitro試験において、500倍未満までの、NM−F9細胞からの最適シアロ形態が必要な可能性がある。Leukine(登録商標)、Leukomax(登録商標)、又はrhGM−CSFの最適シアロ形態の同一濃度で活性を分析したところ、活性の増加は1.5倍まで、好ましくは2倍まで、より好ましくは3倍まで、最も好ましくは4倍まで、非常に好ましくは5倍までであった。先行技術は、rhGM−CSFの活性がそのグリコシル化に左右されることは示していなかったため、これらの知見は特に驚くべきものである。
【0056】
rhGM−CSFの最適シアロ形態で見出される、より高い活性に加えて、そのシアロ形態の比較的高いシアル酸付加度は、治療適用における、より長い血清半減期を保証する。有利には、より長い時間間隔の、より低い用量の投与は、免疫原性及び副作用の軽減又は完全な消失を伴う。後者の2つは更に、ヒト関連グリコシル化パターンにより正の様態で影響される。高活性rhGM−CSFの高いシアル酸付加度は薬物動態学的特性を大きく改善する。
【0057】
本発明は、種々の臨床用途だけでなくin vitro用途においても(例えば、分化樹状細胞及び活性化T細胞の作製のために)前記のとおりに有利に使用されうる高活性糖タンパク質分泌性rhGM−CSFを提供する。
【0058】
本発明の新規技術は、他の増殖因子及びホルモン、例えばEPO、FSH及びhCGの改善のために、並びに種々の他の糖タンパク質、例えばサイトカイン及び抗体(前記のとおり)、及び本明細書に記載のその他のものの改善のために非常に有益である。
【0059】
もう1つの例として、アシアログリコホリンA(AGPA)をNM−F9細胞内で組換え発現させた。血液サンプルから単離し次いで酵素により脱シアル酸してトムセン−フリーデンライヒ(Thomsen-Friedenreich)(TF、コア−1)抗原を露出させたグリコホリンA(GPA)に基づき、TF陽性腫瘍の治療のためのワクチンの開発に成功した。
【0060】
GPA及びAGPAは内在性膜タンパク質である。製造方法の確立のためには、任意の細胞系により、好ましくはNM−F9及びNM−D4細胞により細胞培養培地内に分泌されうるGPAの組換え断片を発現させることが非常に有利であろう。したがって、GPAの細胞外グリコシル化ドメインを含有しGPAの膜貫通ドメインと細胞内ドメインとを欠くGPA断片をコードする発現ベクターsecGPAを構築した。最初の実験では、安定にトランスフェクトされたNM−F9細胞の培地においては分泌性AGPAは少量しか検出され得ないことが示された(実施例を参照されたい)。分泌率を上昇させるためには、いくつかの異種シグナルペプチドをGPA断片に融合させて、内因性シグナルペプチドを除去した。驚くべきことに、配列番号1により特徴付けられるアミノ酸配列を含むGM−CSF由来のシグナルペプチドは、GPAの内因性シグナルペプチド並びにT細胞受容体及び抗体κ軽鎖由来のその他の異種シグナルペプチドと比べて最も有効なシグナルペプチドであった。後者の2つのシグナルペプチドのみが分泌性タンパク質の組換え発現における使用に関して利用可能であり当業者に公知である。
【0061】
本発明は、シアル酸付加を欠損した任意の細胞又は細胞系により発現又は産生される高活性糖タンパク質分泌性アシアログリコホリンAを提供し、好ましくは、該細胞はヒト細胞であり、より好ましくは該細胞はNM−F9又はNM−D4細胞であり、それは、シアル酸付加による遮蔽を伴うことなく好ましい密度でTF抗原を含有する。マウスモデルにおいて抗腫瘍ワクチンとして使用される場合には、それは強力な抗TF及び抗腫瘍応答を示し、それはヒトin vitro研究において強力なTF特異的抗腫瘍応答をも示し、これはGPAのシアル酸付加又は他のグリコシル化形態によっては達成されず、このことは、これが本発明における高活性糖タンパク質形態であることを示している。その強力な抗腫瘍応答は、それがTF特異的腫瘍に対する強力な抗腫瘍ワクチン成分として治療又は予防場面において有効に使用されうることを示している。
【0062】
TF糖鎖は、多数の腫瘍組織上には存在するが健康なヒトの正常細胞上には存在しない周知の腫瘍抗原である。グリコホリンA(GPA)は、シアル酸付加により遮蔽されたTF抗原の担体である。血液サンプルから単離し次いで酵素により脱シアル酸してTF抗原を露出させたGPAに基づき、TF陽性腫瘍患者の治療のためのワクチンの開発に成功した。本発明においては、GPAの唯一のグリコシル化部分でもある細胞外ドメインのみを、シアル酸付加を欠損するヒト細胞(好ましくはNM−F9及びNM−D4細胞)内で分泌形態(secGPA、前記を参照されたい)として組換え産生させる。シアル酸付加欠損細胞、例えばNM−F9又はNM−D4細胞を使用する利点は、その細胞内で任意の形態で発現されるGPA上に存在するTF抗原がシアル酸を含有せず、したがって酵素前処理無しで、規定の標準化されうるワクチンとして使用されうることである。
【0063】
本発明はまた、糖タンパク質をコードする核酸配列並びに配列番号1の核酸配列又は該配列の突然変異、欠失、挿入及び伸長並びに該配列を含有する融合タンパク質を含む核酸を提供する。配列番号1又は該配列の突然変異、欠失、挿入及び伸長並びに該配列を含有する融合タンパク質により特徴付けられるGM−CSF由来のシグナルペプチドは、哺乳動物細胞、好ましくはヒト細胞、より好ましくはNM−wt細胞のサブクローン、最も好ましくはNM−F9及びNM−D4細胞内での任意の哺乳動物遺伝子産物(例えば、GPA)の分泌性発現を駆動するために使用されうる。該シグナルペプチドは、当業者に公知の技術、例えば、目的タンパク質をコードするcDNA内へ該コード領域のフレームと合わせて、シグナルペプチドをコードするヌクレオチド配列を分子クローニングすることにより、目的タンパク質内に組込まれる。それにより、該内因性シグナルペプチドは完全又は部分的に除去される。この構築物を、当業者に公知の任意の発現ベクター内にサブクローニングし、当業者に公知の任意の方法、例えば化学的トランスフェクション、リポフェクション、エレクトロポレーション又は感染により細胞内に導入すると、シグナルペプチドと目的タンパク質とを含む細胞により融合タンパク質が合成される。該細胞内でのこの融合タンパク質のプロセシング中に、該シグナルペプチドは完全又は部分的に除去される。
【0064】
本発明においては、「糖タンパク質」なる語は、ウイルス、ウイルス粒子又はウイルスタンパク質としても定義される。本発明に従い製造されうるウイルス、ウイルス粒子又はウイルスタンパク質には、腸内ウイルス、例えばライノウイルス、ポリオウイルス又はアフタウイルス;ヘルペスウイルス、例えば単純ヘルペスウイルス、仮性狂犬病ウイルス又はウシヘルペスウイルス;オルトミクソウイルス、例えばインフルエンザウイルス;ペスティウイルス;ラブドウイルス;パラミクソウイルス、例えばニューカッスル病ウイルス、呼吸器合胞体ウイルス、ムンプスウイルス又は麻疹ウイルス;レトロウイルス、例えばヒト免疫不全ウイルス、パルボウイルス又はパポバウイルス;ロタウイルス;コロナウイルス;フラビウイルス、例えば黄熱ウイルス又はダニ媒介脳炎ウイルス;肝炎ウイルス、例えばA型肝炎又はB型肝炎ウイルスに由来するものが含まれる。
【0065】
本発明における高活性ウイルス、ウイルス粒子又はウイルスタンパク質は、適当なバイオアッセイ(例えば、ウイルス、ウイルス粒子又はウイルスタンパク質の個々の発現シアル酸付加形態の感染性、ワクチン又はワクチン成分としてのウイルス、ウイルス粒子又はウイルスタンパク質の個々の発現シアル酸付加形態の効力、ウイルス、ウイルス粒子又はウイルスタンパク質の個々の発現シアル酸付加形態の生産性、あるいは当業者に公知の他の有益な医学的又は生物学的特徴の測定)により測定されうるという利点を有する。対応バイオアッセイは、本発明を使用することにより当業者により選択され採用され構築されうる。好ましくは、産生されたウイルスの感染性を、改善されたワクチンが得られるよう示差的シアル酸付加により最適化する。これは該ウイルス、ウイルス粒子又はウイルスタンパク質のシアル酸付加度の最適化を可能にする。
【0066】
好ましくは、ワクチンにおいて使用するための高活性ヒトウイルス、ウイルス粒子又はウイルスタンパク質を産生するヒト細胞系NM−F9又はNM−D4を使用する。該ヒト細胞系は、正常な翻訳後及び翻訳周辺(peri-translational)修飾並びに集合(例えば、グリコシル化)の能力を有し、本明細書に記載されているとおりのシアル酸付加及びシアル酸付加度の最適化を可能にする。
【0067】
ウイルスのシアル酸付加は、しばしば、ウイルスの感染性において重要な役割を果たすことが公知である。シアル酸付加度を最適化することにより、より安全な及び/又はより強力なウイルスワクチンが製造されうる。また、この及び/又はより高い生産性は、改善された、したがってより経済的な製造方法につながりうる。
【0068】
本発明はまた、シアル酸の糖ヌクレオチド生合成経路における少なくとも1つの欠損を含有し、かつ糖タンパク質をコードする核酸でトランスフェクトされた発現細胞系において、ある濃度の少なくとも1つのシアル酸前駆体添加物を添加した培地内で高活性糖タンパク質を発現させることを含む、高活性糖タンパク質の製造方法であって、
該濃度が、
(i)種々の濃度の少なくとも1つのシアル酸前駆体を使用する示差的シアル酸付加により、複数の異なるシアル酸付加形態の糖タンパク質を発現させること、
及び
(ii)適当なバイオアッセイにおいて、参照糖タンパク質と比較して、異なるシアル酸付加形態の活性を測定すること、
及び
(iii)より高い/最高の活性を有するシアル酸付加形態を選択し、該糖タンパク質のより高い/最高の活性レベルと相関する、該シアル酸前駆体添加物の濃度を決定すること
を含む方法により決定される、高活性糖タンパク質の製造方法を提供する。
【0069】
前記方法は、高活性糖タンパク質と決定された糖タンパク質の最適シアル酸付加形態の製造を可能にする。当業者にとっては、本発明の方法を使用することにより、分析的又は予備試験的規模で決定された条件を製造規模へ移すこと、並びに増殖及び培地条件を調節し改善することが可能かつ比較的容易である。他の方法に対する本方法のさらなる利点は、生産性の改善につながりうる規模拡大操作中に、シアル酸前駆体添加物の濃度及び組成の適合化を他の培地成分の最適化に加えて行うことにより、高活性タンパク質の所望のシアル酸付加度を安定化しうることである。
【0070】
本発明における高活性糖タンパク質に関する前記のすべての定義及び説明は、前記及び後記の方法にも有効である。
【0071】
さらに、本発明は、高活性糖タンパク質の同定/決定方法であって、
i)シアル酸の糖ヌクレオチド生合成経路における少なくとも1つの欠損を含有する発現細胞系を、該糖タンパク質をコードする核酸でトランスフェクトすること、
及び
ii)種々の濃度の少なくとも1つのシアル酸前駆体添加物を含有する培地を使用する示差的シアル酸付加により、複数の異なるシアル酸付加形態の糖タンパク質を発現させること、
及び
iii)適当なバイオアッセイにおいて、参照糖タンパク質と比較して、前記の異なるシアル酸付加形態の活性を測定すること、
及び
iv)より高い/最高の活性を有するシアル酸付加形態を選択すること
を含む方法を提供する。
【0072】
原則として、上記方法は、(i)所定の糖タンパク質の活性とそのシアル酸付加度との間に何らかの相関が存在するかどうかを分析するための分析手段、(ii)所定の糖タンパク質のその活性に関する最適シアル酸付加形態が何であるかを決定するための分析手段、及び(iii)最適シアル酸付加形態を与える条件を決定するための分析手段として使用されうる。
【0073】
さらに、本発明は、ヒトグリコシル化、好ましくはシアル酸付加の、生物活性における役割を調べるのに有用である。
【0074】
また、糖タンパク質の示差的シアル酸付加のための方法であって、シアル酸の糖ヌクレオチド生合成経路における少なくとも1つの欠損を含有する発現細胞系を、該糖タンパク質をコードする核酸で形質転換し、複数の異なる濃度の少なくとも1つのシアル酸前駆体添加物を添加した培地内で該細胞を培養し、複数の異なるシアル酸付加形態の糖タンパク質を得ることを特徴とする方法を提供する。
【0075】
本発明はまた、高活性ウイルス、ウイルス粒子又はウイルスタンパク質の同定/決定のための方法、その製造及びその示差的シアル酸付加、並びに本発明により提供される方法から得られる高活性ウイルス、ウイルス粒子又はウイルスタンパク質を提供する。
【0076】
本発明における高活性糖タンパク質に関する前記のすべての定義及び説明は、前記及び後記の方法にも有効である。
【0077】
本発明は、好ましくは、グライコプロファイリング(GlycoProfiling)、グライコエンジニアリング(GlycoEngineering)、グライコアナリティックス(GlycoAnalytics)及びグライコエクスプレス(GlycoExpress)に基づく技術基盤に組み込まれる。
【0078】
・グライコプロファイリング(GlycoProfiling)は、グライコエンジニアリング(GlycoEngineering)のための適当な細胞を選択するために、細胞グリコシル化機構及び該グリコシル化機構により産生された糖(優先的には、細胞表面上に存在するもの)を分析するための或る範囲の技術であると理解される。
【0079】
・グライコエンジニアリング(GlycoEngineering)は、修飾されたグライコプロファイル(GlycoProfile)を有する細胞を与える、細胞のグリコシル化機構を修飾するための或る範囲の技術であると理解される。
【0080】
・グライコアナリティックス(GlycoAnalytics)は、タンパク質のグリコシル化を分子レベルで分析するための或る範囲の技術であると理解される。
【0081】
・グライコエクスプレス(GlycoExpress)は、示差的シアル酸付加、高活性タンパク質の同定及び決定並びに高活性糖タンパク質の製造を含む、本発明の後記の技術及び方法を含む。
【0082】
本発明の糖タンパク質及び方法を含む本明細書に挙げた技術を組合せる利点は、より改善された高活性タンパク質、本発明の意味における、より良好な発現細胞株系、それらの分析及び製造を可能にする。他の利点としては、該方法及び産物の速さ、コスト、安定性及び有効性が挙げられる。
【0083】
前記のすべての定義及び説明は、前記及び後記の糖タンパク質及び方法にも有効である。
【0084】
本発明のもう1つの利点は、糖タンパク質の使用及び機能に関して最適化されたシアル酸付加を伴う高活性糖タンパク質を製造するための、統合された系(統合系)に類似したその方法の組合せである。そのような組合せを以下の好ましい具体例において説明するが、該具体例はこれを限定するものではない。
【0085】
第1ステップにおいては、糖タンパク質、好ましくは分泌性形態の糖タンパク質、好ましくは配列番号1の配列を含む糖タンパク質、より一層好ましくはヒトGM−CSF又はGPAをコードする核酸を、好ましくはエレクトロポレーションにより、シアル酸の糖ヌクレオチド生合成経路における欠損を有する発現細胞系内に好ましくはNM−F9又はNM−D4を使用してトランスフェクトし、好ましくは単一細胞クローニングにより、安定発現クローン(好ましくは無血清培地内、より一層好ましくはタンパク質非含有培地内で増殖するもの)を選択する。
【0086】
第2ステップにおいては、種々の濃度の少なくとも1つのシアル酸前駆体を使用することにより、異なるシアル酸付加形態(好ましくは、一連の少なくとも2、より好ましくは3以上、より好ましくは4以上、より一層好ましくは5以上の異なるシアル酸付加形態)の糖タンパク質を示差的シアル酸付加により製造する。少なくとも1つのバイオアッセイ、例えばヒトGM−CSFに関する細胞増殖in vitroアッセイ、又はGPAに関するある腫瘍細胞に対するより高い特異的免疫原性を示すためのマウス若しくはヒトを使用するin vivo免疫試験により、より高い活性のグライコフォームを決定する。好ましい形態においては、糖タンパク質の用途に応じた最高活性を有する高活性糖タンパク質を決定するために、バイオアッセイの組合せを使用する。本明細書に記載のとおり、選択する高活性糖タンパク質は、すべてのバイオアッセイにおいてより良好である必要はなく、1つのバイオアッセイにおけるより高い活性又は改善された特徴で十分でありうる。しかし、好ましくは、選択する高活性糖タンパク質は、少なくとも2つのバイオアッセイ、例えば1つのin vitroモデル、例えば増殖アッセイにおいて、より良好であり、及び/又は少なくとも1つのin vivoモデルにおいて、より高い活性、及び/又はより高い安定性、及び/又はより長い血清半減期、及び/又はより長いバイオアベイラビリティ、及び/又は免疫原性の改善、及び/又は抗原性の改善を示す。最も好ましいのは、種々の特徴(例えば、受容体媒介機能におけるより高い活性、及びより長い血清半減期を示すことによる改善されたバイオアベイラビリティ)の組合せによるものでありうる、より高いin vivo生物活性をヒトにおいて有するものである。例えば、記載されている条件下の培地内でNM−F9及びシアル酸前駆体添加物としての90mM ManNAcを使用して得られるrhGM−CSFシアル酸付加形態が最高の活性を示したため、これが好ましい高活性糖タンパク質である。
【0087】
もう1つのバイオアッセイにおける更なる試験のために、1つのバイオアッセイにより、より大きなセットから、より小さな群の糖タンパク質シアル酸付加形態を選択することが、速さ及び手段に関しては有利でありうる。GPAの場合には、唯一のシアル酸付加形態、すなわち、最低シアル酸付加形態を作製するので十分であり、したがってこれを無血清培地内でシアル酸前駆体添加物無しで行うので十分である(前記及び後記も参照されたい)。この場合、得られた高活性GPAを、遥かに高い活性(この場合には、動物モデル及びヒトにおいて強力な抗TF免疫応答をもたらす、より高い免疫原性)を示すヒト由来のシアル酸付加GPAと比較することが可能である。しかし、好ましくは、少なくとももう1つのシアル酸付加形態を本発明の方法により作製し、該バイオアッセイにおいて比較する。
【0088】
上記統合系により、高活性糖タンパク質の前記決定の後、該高活性糖タンパク質を製造するためにシアル酸前駆体添加物の対応濃度を使用することにより、糖タンパク質をコードするトランスフェクトされた核酸を含有する同じ発現細胞系を使用することが可能となる。高活性糖タンパク質の製造では、しばしば、製造規模の拡大が望まれる。このためには、シアル酸前駆体添加物の最適条件で十分であるが、所望のシアル酸付加度の高活性タンパク質を安定化するために、例えば他の培地成分を最適化し、異なる反応容器を使用することにより、該シアル酸前駆体添加物の濃度及び組成を調節することが有利又は更には必要かもしれない。これは、本発明を使用することにより当業者により達成されうる。シアル酸前駆体添加物を調節することによりシアル酸付加度を安定化しうることは、先行技術の通常の発現系によっては達成されない該方法の大きな利点である。
【0089】
高活性糖タンパク質の同定及び製造のために実質的に同じ又は類似した方法を使用することを可能にする統合系は、とりわけ、より高い速度及びより低いコストという明らかな利点を有する。
【0090】
上記統合系は、例えばグライコプロファイリング(GlycoProfiling)、グライコエンジニアリング(GlycoEngineering)及びグライコアナリティックス(GlycoAnalytics)として本明細書に記載されている他の製造方法、技術及び方法と組合された場合に特に有利かつ有効である。
【0091】
シアル酸の糖ヌクレオチド生合成経路における復元可能な欠損を有する発現細胞系を作製することは非常に困難である。なぜなら、(i)多数の酵素がシアル酸付加に関与しており、このことは、単一の酵素活性の除去が必ずしも細胞のシアル酸付加能に影響を及ぼさないことを意味し、(ii)該酵素の多くは重複しており、このことは、1つの酵素活性の喪失が、しばしば、他の酵素により補償されることを意味し、(iii)該欠損の復元は、しばしば、可能ではなく、(iv)該欠損の代謝的相補は、しばしば、可能ではないからである。本発明は、好ましくは少なくとも2つの酵素によりシアル酸付加されうる脱シアル酸構造に結合する認識分子で初代細胞又は細胞系から発現細胞系を選択することを含む、シアル酸の糖ヌクレオチド生合成経路における欠損を有する発現細胞系の新規製造方法を提供する。
【0092】
好ましい実施形態においては、選択前に初代細胞又は細胞系からの細胞を突然変異誘発する。突然変異誘発は、化学的若しくは物理的手段によるランダム突然変異誘発又は本明細書に記載の若しくは当業者に公知の部位特異的突然変異誘発により行うことが可能である。
【0093】
本発明の方法とは対照的に、当業者に公知の現在の方法では、特に標的遺伝子が既知でない場合には、シアル酸前駆体経路における遺伝的欠損を有する細胞を作製することは非常に困難であり、時間を浪費し、しばしば可能でない。
【0094】
上記の選択は、当業者に公知の技術(例えば、アフィニティークロマトグラフィー、例えば、磁気細胞選別技術(MACS技術)若しくは免疫沈降若しくはクロマトグラフィー、及び/又はフローサイトメトリーによる細胞選別、及び/又は例えばフローサイトメトリー若しくは免疫細胞化学による少なくとも1つの認識分子への結合特性による細胞の同定、又は分泌性タンパク質の生化学的分析)により、記載されている認識分子の少なくとも1つを使用することによる、該細胞の富化及び/又は単離により行う。
【0095】
選択された細胞を発現細胞系として使用することが可能であり、好ましくは、純粋なクローンを作製するために、公知技術により、例えば単一細胞クローニングにより、更にクローニングする。
【0096】
驚くべきことに、少なくとも2つの酵素によりシアル酸付加されうる脱シアル酸構造を標的化する認識分子の使用は、代謝物質により復元されうる(発現細胞系におけるシアル酸付加度の代謝制御のための前提条件)それらの酵素活性における欠損を有する細胞の選択を可能にし及び/又は該細胞に対して選択を強く偏らせる。
【0097】
本発明の方法はそのような細胞系の作製を可能にし、あるいは速さ、労力の軽減及び/又は成功確率の点でそれらの作製を著しく促進する。これは特にヒト細胞系に当てはまることであるが、ヒト細胞系だけに当てはまるものではない。
【0098】
本発明の方法は、突然変異誘発のための標的遺伝子が既知でない場合に特に適しており、選択はシアル酸付加のダウンレギュレーションの表現型効果に基づいたものである。しかし、それは、部位特異的突然変異誘発を用いる場合、例えばシアル酸付加の表現型ダウンレギュレーションにより安定突然変異誘発クローンの選択を著しく促進することによる、部位特異的組換えによるGlcNAc−エピメラーゼ活性のノックアウトを用いる場合にも有利である。
【0099】
これらの作製される細胞系は、示差的シアル酸付加の目的のために又は本発明における発現細胞系として製造されうるが、それらは、有益又は使用可能である場合には他の目的にも使用されうる。
【0100】
シアル酸付加する能力を欠損した細胞系(その欠損を復元する選択肢を伴っているもの)を作製するためには、原則として、すべての哺乳動物細胞系が適用可能である。製造プロセス(製造方法)には、バイオテクノロジーに適した細胞系、例えばPer.C6、HEK293、K562、CV1、COS−7、ハイブリドーマ細胞、ナマルワ(Namalwa)、BHK及びCHOが好ましい。該欠損を有する新規細胞系を作製するための選抜きの方法は、本発明者らのグライコエンジニアリング(GlycoEngineering)技術に属する前記方法である。
【0101】
上記欠損を有する細胞系を作製するために前記方法を開始する前に、候補細胞系のグリコシル化機構を少なくとも部分的に特徴決定すること(グライコプロファイリング(GlycoProfiling))が非常に有用である。このためには、(i)グリコシル化機構、例えばシアリルトランスフェラーゼ、本明細書中に前記で説明されているシアリルトランスフェラーゼ、トランスポーター(輸送体)、エピメラーゼ、シアリダーゼなどの基質である糖鎖構造に重要な酵素のトランスフェラーゼの関連遺伝子のmRNA発現、(ii)関連酵素、好ましくはシアリルトランスフェラーゼの酵素活性、(iii)シアル酸付加されうる又はシアル酸付加される糖鎖決定基の適当な認識分子による決定、を分析しうる技術の少なくとも1つ(好ましくは、そのような技術の組合せ)を用いるのが好ましい。グライコプロファイリング(GlycoProfiling)は、グリコシル化機構の修飾のための方法(グライコエンジニアリング(GlycoEngineering))に関する決定を可能にする候補細胞系のグリコシル化機構を理解することを助ける。例えば、候補ヒト細胞系、例えばZR75−1、HEK293、NM−wt(K562)、KG1、LS174T、MCF−7、SW480及びT47Dの、グリコシル化に関与する酵素及びタンパク質(例えば、グリコシルトランスフェラーゼ、単糖生合成の酵素及びトランスポーター)のmRNA発現プロフィールをRT−PCRにより分析した。候補細胞系の酵素活性、好ましくはシアリルトランスフェラーゼ活性を分析するためには、バイオアッセイを用いた。バイオアッセイは、糖鎖決定基、好ましくは、少なくとも2つの酵素によりシアル酸付加される又はされうる糖鎖決定基を、適当な方法、例えばフローサイトメトリー及び/又は免疫蛍光顕微鏡検査(好ましくはシアリダーゼと共に、特異的抗体及びレクチンを使用するもの)又は、シアル酸を除去若しくは破壊するための他の方法、例えば穏やかな過ヨウ素酸塩酸化により分析するためにも用いられる。適当なバイオアッセイは公知であり、あるいは当業者により調節又は開発されうる。
【0102】
本発明においては、「初代細胞又は細胞系」なる語は、例えば生物学的試料に由来するもの、例えば血液細胞、組織などからの細胞及び既存の細胞系を含むすべての細胞を包含する。細胞及び細胞系は、公然の(declared)細胞系で時々見られる、単一細胞又は細胞混合物に由来する、特徴決定されたクローンよりなることが可能である。該細胞は既に不死化されていることが可能であり、あるいは本発明の方法の拡張部分として不死化されうる。該不死化は例えば腫瘍由来の細胞の場合のように自発的であってもよいし、あるいは当業者に公知の技術、例えばウイルス形質転換により不死化されてもよい。それにより、当業者は、個々の細胞によって異なりうる、不死化の最良の時点を決定することが可能である。また、該細胞は、他の遺伝子により又は他の遺伝子において、改変されていてもよいし又は改変された状態であってもよい。
【0103】
本発明において、「シアル酸の糖ヌクレオチド生合成経路における欠損」なる表現は、本明細書に記載されているものと同じ欠損を意味する。
【0104】
本発明において、「認識分子」なる語は、部分的に又は好ましくは完全に脱シアル酸されており少なくとも2つの酵素によりシアル酸付加されうる分子に好ましくは及び/又は特異的に結合する分子を意味する。好ましい分子はレクチンであり、より一層好ましいのは、対応する糖鎖構造を認識する抗体である。より一層好ましいのは、TF(トムセン−フリーデンライヒ(Thomsen-Friedenreich)、コア−1)(例えば、Nemod−TF1、Nemod−TF2、A78−G/A7)に結合する抗体、又は末端ガラクトース残基を認識する抗体、又はレクチン、例えばRCA、ECL、PNA、Jacalin、ACA、BPL若しくはAmaranthin、又はシアル酸に結合するもの、例えばSNA及びMAA、又はPankoMabである。
【0105】
より好ましい実施形態においては、該認識分子が結合する脱シアル酸構造はα2−3及びα2−6結合シアル酸によりシアル酸付加されうる。
【0106】
より一層好ましい実施形態においては、該認識分子が結合する構造はO−グリカンである。
【0107】
より一層好ましい実施形態においては、該認識分子が結合する構造は、トムセン−フリーデンライヒ(Thomsen-Friedenreich)構造とも称されるコア−1構造(GalNAcα1−3Gal)である。
【0108】
より一層好ましい実施形態においては、シアル酸の糖ヌクレオチド生合成経路における欠損を有する発現細胞系の作製のための方法は、コア−1構造を認識する抗体で初代細胞又は細胞系から発現細胞系を選択することを含む。
【0109】
より一層好ましい実施形態においては、選択前に該初代細胞又は細胞系に突然変異誘発を行う。
【0110】
一例としては、シアル酸の糖ヌクレオチド生合成経路における欠損を含有する本発明の発現細胞系は、NM−wt細胞を改変することにより作製した。グライコプロファイリング(GlycoProfiling)(前記を参照されたい)により示された多数の候補細胞系のグリコシル化プロフィールに基づき、TF陽性細胞を作製するためにグライコエンジニアリング(GlycoEngineering)のための細胞系を選択した。好ましくは、選択される細胞はC2GNT−L又はMに対して陰性であり、より好ましくは、潜在(cryptic)TFに対して陽性であり、より一層好ましくは、α2,3及びα2,6結合シアル酸により非常に高い度合でシアル酸付加されることが可能であり、最も好ましくは、該細胞はNM−wt細胞である。簡潔に説明すると、該細胞を、当業者に利用可能な任意の方法により、好ましくはフローサイトメトリー又は免疫細胞化学法(実施例に記載されている)により、TF発現に関して分析する。TF陽性細胞は、モノクローナル抗体A78−G/A7又はPankoMabを使用することにより選択される。TF陽性細胞の数が少なすぎる場合には、TF陰性細胞系を突然変異原、好ましくは化学的突然変異原、好ましくはメタンスルホン酸エチル(EMS)で処理する。ついで、前記のとおりにTF陽性細胞を選択する。TF陽性細胞に関する選択は反復する必要があり、ついでTF陽性細胞は、安定にTF陽性である細胞を受容するようクローニングする必要がある。そのようにして、NM−F9及びNM−D4細胞を作製した。
【0111】
同様の適合化された手法により、例えば、特徴決定されたグリコシル化プロフィールを有する糖タンパク質の製造における使用のために、新規グライコプロフィールで、新規細胞系を受容するよう任意の細胞系を糖改変操作することが可能である。そのようなプロフィールは、改善されたシアル酸付加、よりヒト的なグリコシル化、天然糖タンパク質に類似したグリコシル化、又はより低い若しくはより高い免疫原性のグリコシル化でありうる。
【0112】
以下においては、糖タンパク質及び高活性糖タンパク質の医薬組成物又は組成物並びにそれらの用途を更に詳しく説明する。本発明は、本発明の糖タンパク質と希釈剤又は担体とを含む、in vivo又はin vitroでの使用のための本発明の高活性糖タンパク質の組成物又は医薬組成物に関する。本明細書に記載の哺乳動物糖タンパク質及びその組成物は、該糖タンパク質の性質によって定められる広範な用途において使用されうる。これらのすべての用途において、高活性糖タンパク質の使用は常に非常に有益であろう。例えば、本発明の哺乳動物糖タンパク質は、in vitroアッセイ、例えば任意の種類の細胞アッセイ若しくは免疫学的アッセイ、又は分析アッセイ、例えばELISA若しくはRIAにおいて使用されうる。本発明において好ましい糖タンパク質の一例としてGM−CSFを使用する場合には、この増殖因子を、特定の免疫細胞、例えば樹状細胞又はマクロファージの増殖及び/又は分化を誘導するためにin vitroで使用することが可能であり、ついでそれらを、特定の免疫学的アッセイ、例えばELISPOTアッセイ、T細胞アッセイ、細胞傷害性アッセイ、細胞泳動アッセイ、細胞接着アッセイ又は食作用アッセイにおいて使用する。本発明のGM−CSFを細胞増殖アッセイにおいて直接的に使用することも可能である。
【0113】
本明細書に記載の高活性哺乳動物糖タンパク質及びその組成物は診断目的にも有用である。例えば、糖タンパク質の最適シアル酸付加形態を診断目的のための標準として製造することが可能であろう。好ましいのは、本明細書に記載の高活性哺乳動物糖タンパク質及びその組成物のin vivo用途である。本発明の高活性哺乳動物糖タンパク質の広範なin vivo用途、例えば任意の種類の感染性疾患、癌、白血病、造血系の疾患、好中球減少症、血球減少症、骨髄異形成症候群及び自己免疫疾患の予防的及び/又は治療的処置は、具体的な糖タンパク質の性質によって定められ、当業者に公知である。該糖タンパク質がin vivo用途のために適用される哺乳動物種は、該糖タンパク質の起源によって決定される。例えば、ウシGHは、乳生産を増加させるためにウシの治療に使用される。本発明の高活性ウシGHの適用はその治療を改善するであろう。なぜなら、適用の回数及び望ましくない副作用は減少すると予想されるからである。
【0114】
ヒトにおいては、本明細書に記載の糖タンパク質を治療用に使用する。高活性糖タンパク質は常に非常に有益であろう。なぜなら、投与の回数及び望ましくない副作用が軽減されると予想されるからである。患者への投与の場合には、本発明の精製された糖タンパク質を、通常の方法に従い、薬学的に許容される担体又は希釈剤と混合する。治療用製剤は、静脈内注入若しくは皮下注射により、又は当技術分野において公知の任意の他の許容される方法により投与される。該製剤は、所望により、他の治療薬をも含有しうる。投与の量、頻度及び期間は様々であり、使用する特定の化合物の活性、年齢、体重、全身健康状態、性別、投与の食事時間、投与経路、排泄速度、薬物の組合せ及び疾患特異的療法の重篤度を含む種々の要因によって異なるであろう。単一剤形を与える、担体物質と組合される有効成分の量は、治療対象の宿主及び個々の投与方式によって様々である。
【0115】
本発明の好ましい実施形態においては、該医薬組成物はワクチン又はワクチン−アジュバントである。本発明においては、ワクチン組成物なる語は、哺乳動物及びヒトにおいてワクチンとして使用されうる任意の組成物に関するものである。ワクチンは、免疫応答を誘導する治療用又は予防用の医薬組成物を意味する。本発明のワクチン組成物の製造方法の形態は特に限定されるものではなく、所望の形態の組成物は、好ましくは、当技術分野において利用可能な単一の方法、又は適当に組合された方法を適用することにより製造されうる。任意のワクチン及び/又はワクチン−アジュバントの投与は皮下、皮内、静脈内、非経口、筋肉内又は任意の他の許容される方法でありうる。前記剤形により予想される製剤及び医薬組成物は、通常の技術を用いて、通常の薬学的に許容される賦形剤及び添加剤を使用して製造されうる。他のアジュバントは該ワクチン又は該糖タンパク質と共に投与されうる。
【0116】
ヒト腫瘍ワクチンの一例はアシアログリコホリンA(AGPA)である。グリコホリンA(GPA)は、赤血球の膜に存在し幾つかの細胞系にも見出されるシアル酸付加糖タンパク質である。ワクチンの製造のためには、それをヒト血液製剤(ヒト血液調製物)から単離し、酵素により処理して、GPAに結合したすべてのシアル酸を除去する。得られるAGPAは腫瘍特異的糖鎖抗原トムセン−フリーデンライヒ(Thomsen-Friedenreich)(TF)のキャリアであり、したがって、TFに対する持続性免疫反応を誘導し従って患者のTF陽性腫瘍負荷を排除又は少なくとも軽減するためのワクチンとして適用される。本明細書に記載のシアル酸付加欠損細胞の使用は本発明により、酵素処理を要することなく、かつ、製造用にヒト血液製剤を使用することに限定されることなく、AGPAの組換え製造を可能にする。AGPAは、腫瘍及び癌患者の免疫療法のためのワクチンとしての糖タンパク質及びその医薬組成物の使用の一例である。
【0117】
また、本発明は、ワクチン(特に、適当な賦形剤及び/又はアジュバントを含む医薬組成物に配合された場合)において使用するための、本発明により提供される方法により得られた高活性ウイルスタンパク質、ウイルス粒子及び/又はウイルスを提供する。投与量及び投与方法は前臨床及び臨床開発を通じて当業者により決定されうる。
【0118】
ワクチン−アジュバントは、典型的には、弱いとみなされている動物又はヒトワクチンによりもたらされる防御(すなわち、レベル、度合及び/又は持続時間の点で弱い防御)を増強するために使用される。ワクチン−アジュバントとして使用される糖タンパク質の具体例としては、GM−CSF、並びに特定のサイトカイン、例えばIL−2及びIL−7が挙げられる。ワクチン−アジュバントとしての糖タンパク質は、ワクチンとは別々に、あるいはまた、ワクチンと組合せて投与されうる。ワクチン−アジュバントとしての糖タンパク質をワクチンと組合せる場合には、投与する組成物は、与えられた病原体又は抗原に対する特異的応答を惹起するのに有効な免疫原、薬学的に許容されるワクチン担体、及び免疫を増強する量の糖タンパク質を含有する。その概念の一例は、当技術分野で公知の全細胞ワクチンであり、この場合、抗原の担体である細胞によりワクチン−アジュバントが組換え発現される。全細胞ワクチンは、典型的には、細胞死が運命づけられた活性な細胞として適用される。ワクチン接種後、これらの細胞は、ワクチン−アジュバントとして作用する一定量の糖タンパク質、例えばGM−CSF、IL−2又はIL−7を一定時間にわたって産生する。全細胞ワクチンは、ワクチン接種後の該全細胞ワクチンによりワクチン−アジュバントが更に産生されることなく全細胞ワクチンの利点を尚も有するよう、ワクチン接種前に不活化されうる。本発明は該ワクチン−アジュバントを提供するだけでなく、全細胞ワクチンとして使用される細胞系(1若しくは複数)もまた、本発明により提供される方法により作製されうる。全細胞ワクチンに使用される細胞の特徴は、そのような細胞系又は細胞系の組合せが、可能な限り多数の抗原を発現することである。当業者に公知のとおり、これらの細胞又は細胞系の組合せは、当技術分野において公知の方法により、直接的にワクチン接種されたり、あるいは抗原提示細胞、例えば樹状細胞上に負荷(ローディング)されることが可能である。ついで該抗原負荷樹状細胞はワクチン接種に使用され、あるいは当業者に公知の方法により養子T細胞療法のために身体外(ex corpore)でT細胞を刺激するために使用される。本発明においては、該ワクチンは、本明細書に記載の方法により製造された、ワクチン−アジュバントとして本発明の糖タンパク質を発現する細胞系又は細胞系の組合せでありうる。
【0119】
糖タンパク質又はその医薬組成物は、感染性疾患、例えばエイズ、SARS又は或る形態の肝炎の治療のためのワクチン又はワクチン−アジュバントとして使用されうる。哺乳動物糖タンパク質は、ヒトの感染性疾患、例えばHIV、SARS、B型肝炎若しくはA型及びB型インフルエンザ、又は動物の感染性疾患、例えばウシウイルス性下痢、ウマインフルエンザ、ネコ白血病、ネコ呼吸疾患の治療のために感染細胞に対する免疫応答を誘導する本発明に従い製造される弱毒化生又は不活化ウイルス産物又は組換え抗原性ウイルス産物の一部である。
【0120】
本発明は、本発明の糖タンパク質及び/又はその医薬組成物及び/又はその類似体、修飾体、薬理学的に活性な断片並びに場合によっては使用方法に関する情報を含むキットに関する。本発明の好ましい実施形態においては、該キットは、高い/より高い活性の糖タンパク質、例えばGM−CSF、EPO又はFSH及びその薬学的に許容される担体の医薬組成物の容器、並びにワクチン及びその薬学的に許容される担体の医薬組成物の容器を含むワクチンにおける、抗原に対する哺乳動物の免疫原性応答を増強するために使用されうる。
【0121】
本発明の糖タンパク質は、感染性疾患の治療のためのワクチン又はワクチン−アジュバントとして使用されうる。また、本発明の糖タンパク質は、感染性疾患の治療のためのワクチン又はワクチン−アジュバントの製造にも使用されうる。
【0122】
本発明の糖タンパク質は、疾患、例えば白血病、好中球減少症、血球減少症、癌、骨髄移植、造血系の疾患、不妊症及び自己免疫疾患の予防的及び/又は治療的処置に使用されうる。糖タンパク質の、当業者に公知の治療用途の範囲は、非常に広範である。例えば、G−CSFは、白血病性癌患者の化学療法により生じる、生命を脅かす好中球疾患である好中球減少症を治療するための重要な治療剤である。GM−CSFは、好中球減少症からの速い回復を達成するために、化学療法後の比較的高齢のAML患者の治療に特異的に使用される。さらに、GM−CSFは、骨髄移植におけるいくつかの用途及び末梢血幹細胞の動員(mobilization)のための治療剤として承認されている。また、現在研究中である、GM−CSFのいくつかの臨床用途(例えば、HIV及び癌の治療)が存在する。造血系の或る疾患はEPOで治療され、IFN−βは現在、自己免疫疾患である多発性硬化症の治療のための重要な治療剤である。他の一例はFSHであり、これは男性及び女性不妊症の治療に広く使用されている。hCGは不妊症の治療にも適用されるが、女性における無排卵に焦点が合わされている。hGHは、臨床的に証明されている利益(例えば、体脂肪減少及び筋組織増加)を有する。
【0123】
本発明の糖タンパク質は、白血病、好中球減少症、血球減少症、癌、骨髄移植、造血系の疾患、不妊症及び自己免疫疾患を含む群から選ばれる疾患の予防的及び/又は治療的処置のための医薬の製造にも使用されうる。
【0124】
本発明の糖タンパク質、ワクチン又は医薬は、任意の通常の投与経路、例えば非経口、眼内、局所、吸入、経皮、膣内、頬側、経粘膜、経尿道、直腸、鼻腔、経口、肺又は耳経路により使用されうる。
【0125】
本発明の治療用糖タンパク質の他の一例は抗体である。抗体は、高いアフィニティーで特異的標的に結合することにより作用する。抗体は、毒素又は放射性同位体に結合している場合、腫瘍細胞又は感染細胞に対する強力な治療剤である。しかし、「裸」抗体も、細胞傷害性及び/又は食作用活性を有する特異的免疫細胞(ADCC)又は補体(CDC)を標的に導くことにより、特定の又は細胞標的の排除を媒介しうる。特に、抗体のADCC及び/又はCDC活性は、与えられた抗体のグリコシル化における適当な変化により改善されうる。
【0126】
本発明は本明細書に記載の特定の方法、組成物及び細胞系には限定されないと理解されるべきである。なぜなら、そのような方法、組成物及び細胞系は、もちろん、様々でありうるからである。また、本明細書中で用いる用語は、個々の実施形態を説明する目的のものであるに過ぎず、添付の特許請求の範囲のみによって定義される本発明の範囲を限定するものではないと理解されるべきである。
【0127】
特許請求の範囲を含む本明細書において用いる単数形表現は、文脈が例外を明らかに示している場合を除き、それらの対応複数物を含む。したがって、例えば、「生物」に対する言及は1以上の異なる生物を含み、「細胞」に対する言及は1以上のそのような細胞を含み、「方法」又は「製造方法」に対する言及は、当業者に公知の等価なステップ、方法又は製造方法に対する言及を含む、などである。
【0128】
特に示さない限り、本明細書中で用いるすべての科学技術用語は、本発明が属する分野の当業者により一般に理解されているのと同じ意義を有する。本発明の実施又は試験においては、本明細書に記載されているものと類似した又は同等な方法、製造方法及び物質が使用されうるが、適当な方法及び物質は後記で説明されている。
【0129】
以下の表1及び2、並びに図1〜15は、最適化されたシアル酸付加を伴う高活性ヒトタンパク質の製造のための本発明を説明するものである。
【0130】
表1:種々のヒト細胞系におけるグリコシルトランスフェラーゼの発現プロフィール。示されている値はZR75−1に対するパーセントとして表されている。それぞれの場合に、ハウスキーピングβアクチン対照に対するグリコシルトランスフェラーゼmRNA発現の測定のために二重RT−PCRを用いた。
【0131】
表2:モノクローナル抗体及びレクチンによる種々のヒト細胞系における糖鎖決定基の検出。示されている値は、フローサイトメトリー分析により測定した、陰性対照としてのイソタイプ又は二次試薬に対する、抗体/レクチン結合の差引平均である。例えば、モノクローナル抗体A78−G/A7及びレクチンPNA(ピーナツ凝集素)及びアルトカルプス・インテグリフォリア・ジャカリン(Artocarpus integrifolia Jacalin)はTF特異的である。ノイラミニダーゼでの細胞の処理(白色で文字修飾されている)は、アフィニティー結合を可能にするTF露出を引き起こした。
【0132】
図1:NM−wt細胞のグライコエンジニアリング(GlycoEngineering)及び得られたNM−F9細胞の特徴決定。EMS処理によりランダム突然変異誘発を行った。脱シアル酸エピトープの露出によって、EMS処理NM−wt細胞のアフィニティー選択及びそれに続くクローニングが可能となり、TF特異的抗体A78−G/A7を使用した免疫細胞化学法で測定される高露出TF構造を有する細胞クローンが得られた(A,上パネル:EMS処理NM−wt細胞;下パネル:アフィニティー選択されクローニングされたNM−F9細胞)。シアル酸付加度は、(B)TF特異的抗体A78−G/A7、末端βGal認識性レクチンPNA及びシアル酸特異的レクチンSNAをアプライするフローサイトメトリーにより、あるいは(C)チオバルビツール酸法により測定した。
【0133】
図2:NM−wt細胞並びに誘導体NM−F9及びNM−D4によるTF、sTF、Tn、sTn、グリコホリンA(GPA)、アシアログリコホリンA(AGPA)、Lex、sLex及びLeyの発現。細胞をフローサイトメトリーにより分析した。また、細胞をシアリダーゼで処理し、AGPA(A63−C/A9)及びTF(A78−G/A7)の発現を分析した。
【0134】
図3:(A)レクチンPNA、SNA、MAA及びUEAIを使用するフローサイトメトリーによる、あるいは(B)チオバルビツール酸法による、NM−wt細胞並びに誘導体NM−F9及びNM−D4上の膜複合糖質結合シアル酸の測定。
【0135】
図4:(A)mRNA発現分析による、及び(B)酵素活性分析による、NM−F9細胞の遺伝的欠損の分析。mRNA発現分析は、RNAの単離、UDP−GlcNAc−2−エピメラーゼ特異的プライマーを使用するRT−PCR及びゲル電気泳動を含む。対照としてアクチンを泳動させた。酵素活性測定は、14C−UDP−N−アセチルグルコサミンから14C−N−アセチルマンノサミンへのin vitro変換の測定に基づく。UDP−GlcNAc−2−エピメラーゼのmRNAは検出することができなかった。酵素活性の欠如はエピメラーゼの遺伝的欠損を証明した。
【0136】
図5:代謝的相補によるシアル酸付加の復元。NM−F9及びNM−wt細胞を、0又は50mMのそれぞれManNAc又はGlcNAcを含有する血清中で4日間培養した。モノクローナル抗体A78−G/A7(抗TF)及びA63−C/A9(抗AGPA)並びにレクチンSNA、MAA及びPNAをフローサイトメトリー測定において使用した。ManNAcの添加はシアル酸付加の復元及びTFの消失をもたらした。UDP−GlcNAc−2−エピメラーゼ欠損により、GlcNAcは代謝され得なかった。
【0137】
図6:遺伝的相補によるシアル酸付加の復元。NM−F9細胞を、エピメラーゼをコードする構築物pcDNA3.1Zeo(−)/2Epiで安定にトランスフェクトした。モノクローナル抗体A78−G/A7及びA63−C/A9並びにレクチンSNA及びPNAをフローサイトメトリー測定において使用した。シアル酸付加は完全に復元された。
【0138】
図7:NM−F9におけるrhGM−CSFの発現。ヒトGM−CSF発現ベクターで安定にトランスフェクトされたNM−F9細胞をクローニングし、FCSを添加した又は添加していない培地内で培養した。1日後(1d)、2日後(2d)、3日後(3d)及び4日後(4d)に、ELISAにより上清中のGM−CSF濃度を測定した。4日後、NM−F9細胞105個当たり14ng/mlのGM−CSFが分泌された。
【0139】
図8:種々の分泌シグナルペプチドを使用した場合の、NM−F9細胞による、組換え発現された分泌性AGPAの発現。分泌性AGPAの発現のために、GM−CSFのシグナルペプチド(secGM/GPA)、T細胞受容体のシグナルペプチド(secTCR/GPA)、抗体軽鎖κのシグナルペプチド(secAK/GPA)及び内因性シグナルペプチド(secGPA)を含有する4種の発現ベクターを構築した。捕捉体としてのA83−C/B12(抗GPA)及び検出体としてのA63−C/A9を使用するサンドイッチELISAにより細胞上清中で分泌性AGPAを検出した。
【0140】
図9:代謝改変による膜複合糖質の示差的シアル酸付加。NM−F9細胞を無血清培地内(A)又はfcs含有培地内(B)(種々の濃度のManNAcを添加したもの)で4日間培養した。膜複合糖質結合シアル酸の量を細胞膜画分のチオバルビツール酸法により測定した。比較のために、NM−wt細胞も、ManNAc添加を行わないfcs含有培地内で4日間培養した。
【0141】
図10:代謝的相補による細胞表面上の膜タンパク質の示差的シアル酸付加。NM−F9細胞を種々の濃度のManNAcを添加したFCS非含有培地内で4日間培養した。比較のために、NM−wt細胞も、ManNAc添加を行わないFCS非含有培地内で4日間培養した。シアル酸を含まないTFエピトープを認識するレクチンPNAを使用してフローサイトメトリー分析を行った。
【0142】
図11:代謝的相補による分泌性rhGM−CSFの示差的シアル酸付加。(A)種々の濃度のManNAc(0、50、70又は90mM)及び(B)−/+ 1又は3mg/mlのシアロ糖タンパク質フェツインを添加した無血清培地内でrhGM−CSF産生細胞を4日間培養した。シアル酸を含まないrhGM−CSFをELISAにより測定した。まず、上清中のrhGM−CSFの濃度を測定し、ついでNM−F9細胞培養物1ml当たり100ngに調節した。つぎに、シアル酸を含まないrhGM−CSFの検出のために、抗ヒトGM−CSFモノクローナル抗体を使用してrhGM−CSFを捕捉し、シアル酸を含まないN−及びO−グリカンをPNAにより検出した。
【0143】
図12:TF1細胞上の示差的シアル酸付加rhGM−CSFの増殖活性。示差的シアル酸付加のために、90mMまでの漸増濃度のManNAc(A)及び1mg/mlの規定(defined)血清シアロ糖タンパク質と共に90mM ManNAc(B)を添加した無血清培地内でNM−F9細胞を培養した。TF1細胞を、5ng/mlの種々のシアル酸付加rhGM−CSFを含有するNM−F9上清と共に48時間インキュベートした。Leukine(登録商標)(酵母内で産生)及びLeukomax(登録商標)(大腸菌内で産生)を基準として使用した。GM−CSFを含有しないNM−F9上清を陰性対照として使用して、BrdU−増殖アッセイにより細胞増殖を測定した。最高ではないが高いシアル酸付加度を有するrhGM−CSFが、TF1細胞増殖のための最も活性な増殖因子であった。
【0144】
図13:樹状細胞上の示差的シアル酸付加rhGM−CSFの増殖活性。示差的シアル酸付加のために、90mMまでの漸増濃度のManNAcを添加した無血清培地内でNM−F9細胞を培養した。NemodDC細胞を5ng/mlの種々のシアル酸付加rhGM−CSFのNM−F9上清と共に24時間インキュベートした。Leukine(登録商標)を基準として使用した。GM−CSFを含有しないNM−F9上清を陰性対照として使用して、BrdU−増殖アッセイにより細胞増殖を測定した。異なるシアル酸付加度は、選択された糖タンパク質活性に正又は負の影響を及ぼしうる。
【0145】
図14:rhGM−CSF濃度に対する増殖活性の依存性。rhGM−CSFの最高の活性を得るために、90mM ManNAcを添加した無血清培地内でNM−F9細胞を培養した。種々の濃度のLeukomax(登録商標)、又は種々の濃度のrhGM−CSFを含有するそのNM−F9細胞の上清と共に、TF1細胞を48時間培養した。GM−CSFを含有しないNM−F9上清を陰性対照として使用して、BrdU−増殖アッセイにより細胞増殖を測定した。完全ヒトGM−CSFのin vitro活性は、大腸菌(Leukomax)又は酵母(Leukine)内で発現されたhGM−CSFより約500倍高い。
【0146】
図15:rhGM−CSF増殖活性の抑制。NM−F9細胞を無血清培地+/−90mM ManNAc内で培養した。TF1細胞を5ng/mlのシアル酸付加(a)若しくは非シアル酸付加rhGM−CSF(c)又はLeukine(登録商標)(b)のNM−F9上清と共に48時間培養した。rhGM−CSFは種々の濃度の抗ヒトGM−CSF抗体BVD2−23 B6でブロッキングされた。GM−CSFを含有しないNM−F9上清を陰性対照(d)として使用して、BrdU−増殖アッセイにより細胞増殖を測定した。
【0147】
以下の実施例は例示として記載されており、限定的なものではない。実施例においては、コンタミネーション(汚染)活性を伴わない標準的な試薬及びバッファーを使用する。リボヌクレアーゼ及びPCR産物のコンタミネーションを避けるように留意するのが好ましい。
【実施例1】
【0148】
グライコプロファイリング(GlycoProfiling)
種々のヒト細胞系からのグリコシルトランスフェラーゼC1GalT1、C2GNT−L、C2GNT−M、ST6GalNAc−I、ST6GalNAc−II、ST3Gal−I、ST3Gal−II及びST6Gal−Iの発現プロフィールを調べた。細胞系HEK293、LS174T、MCF−7及びSW480をDMEM内で培養し、一方、T47D、ZR75−1、NM−wt及びKG1をRPMI1640内で培養した。どちらの培地にも10%ウシ胎児血清及び2mMグルタミンを添加した。すべての細胞を6%CO2の湿潤雰囲気中、37℃で増殖させた。当業者に公知の標準的な技術を用いて細胞を回収し破壊した。RNAを、グアニジンチオシアネート−フェノール−クロロホルムを使用して抽出し、RNase非含有DNaseで処理した。RNA収量を分光光度法により測定した。
【0149】
mRNA発現分析を以下のとおりに行った。5μgの全細胞RNAをランダムヌクレオチドヘキサマーでモロニーマウス白血病ウイルス逆転写酵素により逆転写した。RT混合物は逆転写酵素、mRNA鋳型、プライマー、バッファー及びdNTPを含む。cDNAの収量及び完全性を測定するために、並びにゲノムDNAでの考えられうるコンタミネーションに関して確認するために、β−アクチンハウスキーピング遺伝子の発現を利用した。この目的には、イントロンにまたがるβ−アクチン特異的プライマーセットを設計した:β−アクチンフォワードプライマー:5'-GGC ATC GTG ATG GAC TCC G-3'及びβ−アクチンリバースプライマー5'-GCT GGA AGG TGG ACA GCG A-3'(アンプリコン長,622bp)。
【0150】
得られたcDNA鎖を直接、増幅のために使用した。前記酵素の1つをコードするcDNAの5’−(フォワード)及び3’−領域(リバース)に特異的にハイブリダイズするフォワードプライマー及びリバースプライマーを設計した(C1GalT1を、フォワードプライマー:GAG ATT CCA GAG ATA CCA TTG及びリバースプライマー:CGT TCA GGT AAG GTA GGT TGを使用して増幅した(アンプリコン長262);C2GNTのフォワード:GTG CTC AGA ATG GGG CAG GAT GTC ACC TGG, リバース:TCA CTA CTA GGA TTC TCC CCA GCA AGC TCC(アンプリコン長360);ST3Gal−Iのフォワード:ATG AGG TGG ACT TGT ACG GC, リバース:AAC GGC TCC AGC AAG ATG(アンプリコン長375);ST3Gal−IIのフォワード:CCC TGC TCT TCA CCT ACT CG, リバース:GCA TCA TCC ACC ACC TCT G(アンプリコン長282);ST6Gal−Iのフォワード:AAA AAC CTT ATC CCT AGG CTG C, リバース:TGG TAG TTT TTG TGC CCA CA(アンプリコン長379);ST6GalNAc−Iのフォワード:ACC ACA GCC AAG ACG CTC, リバース:AAG GGT GGT GCA AAG TGT TC(アンプリコン長407);ST6GalNAc−IIのフォワード:CTG CCA GTA AAT TCA AGC TGC, リバース:TTG CTT GTG ATG AAT CCA TAG G(アンプリコン長184))。PCR反応混合物は、1.5μlのPCRバッファー[100mM Tris−HCl(pH8.3),500mM KCl]、0.8〜1.0μlの25mM MgCl2、5μlのcDNA、1.5μlの2mMデオキシヌクレオチド三リン酸、0.2μlのAmpliTaqポリメラーゼ、0.2μlの0.25μMβ−アクチンプライマー、0.7〜0.8μlの50μM酵素特異的プライマー及びH2O(容量15μlとする)を含有していた。PCRは、変性、アニーリング及び伸長のための複数の温度サイクルで行った。サイクル条件は94℃で1分間、59℃で1分間及び72℃で2分間であった。同じチューブ内で0.2μlのβ−アクチンプライマーでβ−アクチンを同時増幅した。各PCR反応を2回行った。
【0151】
26〜28サイクルの後で得られたPCR反応のアリコートを2%アガロースゲル中で電気泳動し、臭化エチジウム染色の強度により定量した。RT−PCRデータの半定量的分析のために、各cDNA特異的バンドの染色強度を、同時増幅されたβ−アクチンに対応するバンドの染色強度と比較した。比率[(酵素バンドの蛍光単位/β−アクチンの蛍光単位)×100]を計算することにより、各酵素バンドの強度を、適当なβ−アクチンバンドと相関させた。検出限界は二本鎖DNAとして1ngであった。該アッセイは25ngまでは直線関係を示した。
【0152】
グライコプロファイリング(GlycoProfiling)は、NM−wt細胞のみにおいて、C2GNT−L及びC2GNT−Mの欠如により、コア2の合成の完全な欠損を示した。mRNA発現における他の低減(ST6GalNAc−IIは8%へ、ST3Gal−Iは32%へ)は無傷ST6GalNAc−I及びST3Gal−IIにより補償されることが可能であった(表1)。NM−wtのグリコシルトランスフェラーゼプロフィールは、将来のグライコエンジニアリング(GlycoEngineering)及びシアル酸付加研究(TFエピトープを使用することによるもの)のための最良の出発点を表した。
【0153】
特にシアル酸に関する糖鎖決定基は、いくつかのモノクローナル抗体及びレクチンをアプライするフローサイトメトリーにより、前記ヒト細胞系において検出された。
【0154】
細胞を集め、PBSで2回洗浄し、37℃で1時間かけて回復させた。ノイラミニダーゼ処理のために、細胞を、0.1%CaCl2及び0.4%NaClを含有する0.1Mイミダゾールバッファー(pH6.8)中、ビブリオ・コレラ(V. cholerae)ノイラミニダーゼの存在下又は非存在下にて、37℃で30分間インキュベートした。細胞を染色バッファー(4%BSAを含有するHBSS又は10%FCSを含有する培地)に再懸濁させた。細胞表面糖鎖決定基の検査のために、種々の抗体及びレクチンを使用した。例えば、ビオチン標識レクチンピーナツ凝集素(PNA)、アルトカルプス・インテグリフォリア(Artocarpus integrifolia)由来のジャカリン(jacalin)、アマランツス・カウダツス(Amaranthus caudatus)凝集素(アマランチン(Amaranthin)又はACA)及びバウヒニア・プルプレア(Bauhinia purpurea)レクチン(BPL)並びにTF特異的モノクローナル抗体A78−G/A7を使用して、TFの発現を分析した。PankoMabは腫瘍−MUC1を検出する。A83−C/B12は、グリコホリンAをそのグリコシル化には無関係に認識する抗GPA抗体である。細胞をレクチン又は抗体と共に4℃で1時間インキュベートし、染色バッファーで2回洗浄した。該抗体の結合は、二次Cy3−ヤギ抗マウスIgG/IgM抗体により検出した。レクチンの結合は、Cy3結合ストレプトアビジンにより検出した。Coulter Epics XLフローサイトメーター(Beckman Coulter, Germany)を使用してフローサイトメトリー分析を行った。Expo32ソフトウェア(Becton Coulter)を使用して定量分析を行った。
【0155】
ZR75−1の場合を除き、ヒト細胞はTF陰性であった。しかし、シアル酸を切り離すノイラミニダーゼで細胞を処理した後、多数の細胞系上で潜在TFが可視化される。分析した細胞系のうち、NM−wt細胞が潜在TFの主要キャリアであった。本発明においては、多量の元の潜在TF及び遊離シアル酸が好ましい(表2)。
【実施例2】
【0156】
グライコエンジニアリング(GlycoEngineering)及び新規クローンの特徴決定
NM−wt細胞をアルキル化剤メタンスルホン酸エチルで処理することにより、ランダム突然変異誘発を行った。サンプルごとに、NM−wt細胞をPBSで洗浄し、5%CO2、37℃で一晩、EMS(0.1mg/ml、メタンスルホン酸エチル、Sigma-Aldrich)を添加した細胞培養培地1ml当たり106細胞で播いた。細胞を洗浄し、新鮮培地を供給した。2日ごとに、トリパンブルー染色により細胞生存度を測定し、免疫細胞化学的染色により細胞を分析した(図1A)。
【0157】
ついで、高いTF発現の新規表現型を示す細胞を、TF特異的抗体を使用して選択した。NM−wt細胞をB−PBS(PBS中、0.5%BSA)で洗浄し、モノクローナル抗体A78−G/A7又はPankoMabのハイブリドーマ培養の上清50μl及びB−PBSの950μlと共に4℃で30分間インキュベートした。洗浄後、50μlのラット抗マウスIgM抗体又はラット抗マウスIgG抗体(MicroBeads(Miltenyi Biotec, Koln, Germany)に結合しているもの)で操作を繰返した。洗浄後、Miltenyi Biotec(Koln, Germany)により供給された2つの連続的なカラムにより、該製造業者のマニュアルに記載されているとおりに、磁気標識TF陽性細胞を分離した。9日間の培養の後、単離操作を合計3回繰返した。FACS分析(フローサイトメトリー)を抗体染色から始めた。約3×105細胞を一次モノクローナル抗体[A78−G/A7(IgM)、PankoMab(IgG1)のハイブリドーマ培養上清;すべて、細胞培養培地中で1:2希釈されたもの]と共に4℃で1.5時間インキュベートし、ついで、PBSで1:200希釈された二次Cy3結合ヤギ抗マウスIgM/IgG抗体と共に4℃で30分間インキュベートし、再び洗浄した。再懸濁細胞(200μlのPBS)をフローサイトメトリーにより調べた。以下のパラメーターでExpo32ソフトウェア(Becton Coulter)を使用して、定量分析を行った:抗体標識細胞に関するパラメーター:前方散乱(FS):26V,ゲイン1,側方散乱(SS):807V,ゲイン5,FL2:740V,ゲイン1、及びレクチン標識細胞に関するパラメーター:FS:26V,ゲイン1,SS:807V,ゲイン5,FL1:740V,ゲイン1。
【0158】
3回の単離の後、93%TF陽性細胞の細胞集団を得た。しかし、TF陽性細胞の比率は時間経過と共に減少し、単離操作後の14日間の期間中にTF陽性細胞約20%の最低レベルに達した。TF陽性表現型の安定発現のために、細胞を4回目の単離に付し、ついで最後に、単離されたTF陽性細胞を96ウェルプレート(1細胞/100μl)内での限界希釈によりクローニングした。得られた30個の細胞クローンのうち、13個の細胞クローンが多量のTF抗原を発現し、これらのうちの8個の細胞クローンが全細胞集団の細胞表面上で、均一なTF発現を示した(図1A)。現在までに(約30ヶ月間)、これらの細胞クローンはTF抗原を安定に発現している。フローサイトメトリーによる分析は、NM−F9細胞の場合のTF発現レベルの約31倍の増加を示し、このことは、このクローン上のTFの強力な発現を表しており、約35の、FACS分析における差引平均によっても示され、これを更なる特徴決定のために選択した。安定なTF陽性細胞集団を得るためにクローニングが必要なのは、TF陰性細胞の、より高い増殖速度によるものであり、したがってこれは時間経過と共にTF陽性クローンより優勢に増殖し、例えば、NM−F9は、NM−wt細胞より遅い倍加速度を有する。
【0159】
さらに、PankoMab及びラット抗マウスIgG抗体(MicroBeads(Miltenyi Biotec, Koln, Germany)に結合しているもの)を使用した以外は上記TF選択と同じ方法で、MUC1選択を行った。より多くの腫瘍特異的MUC1エピトープTA−MUC1を発現するTF陽性クローンの作製のために、NM−F9細胞を処理し、選択し、単一細胞をクローニングした。これは、選択のためにPankoMabを使用して、前記のとおりに行った。安定なクローンNM−D4は、フローサイトメトリーにおいてPankoMab染色の増強を示したため、該クローンを更なる特徴決定のために選択した。
【0160】
NM−F9及びNM−D4は、より低い連続的な濃度のウシ胎仔血清(FCS)を含有する培地内に継代することにより、無血清培地に適合化することが可能であった。
【0161】
NM−F9及びNM−D4を、それらの糖鎖決定基及びトランスフェラーゼ発現に関して詳細に特徴決定した。評価されうる膜複合糖質のシアル酸付加度を、実施例1に記載のとおりにTF特異的抗体A78−G/A7及びいくつかのレクチン、例えば末端β−Gal認識性PNA、シアル酸特異的サムブクス・ニグラ(Sambucus nigra)レクチン(SNA)、又はその他の例えばマアキア・アムレンシス(Maackia amurensis)レクチンI(MAA)及びウレックス・ユーロパエウス(Ulex europaeus)凝集素I(UEAI)をアプライするフローサイトメトリーにより測定した。PBSで2回及び4%BSA含有HBSS(ハンクス平衡塩)で1回洗浄した後、細胞をHBSS/4%BSA中、FITC結合PNA(1:400)、FITC結合SNA(1:50)、FITC結合MAA(1:50)又はFITC結合UEAIと共に4℃で1時間インキュベートした。洗浄後、分析のために細胞ペレットを200μlのHBSS/4%BSAに再懸濁させた。以下のパラメーターでExpo32ソフトウェア(Becton Coulter)を使用して、定量分析を行った:抗体標識細胞に関するパラメーター:前方散乱(FS):26V,ゲイン1,側方散乱(SS):807V,ゲイン5,FL2:740V,ゲイン1、及びレクチン標識細胞に関するパラメーター:FS:26V,ゲイン1,SS:807V,ゲイン5,FL1:740V,ゲイン1(図2及び3A)。
【0162】
野生型細胞の潜在TFはA78−G/A7及びPNAの結合を妨げたが、誘導体NM−F9及びNM−D4の露出TFは、使用した検出試薬に応じて認識を30〜75倍増強した。この結果はSNAの逆アフィニティーにより定性的に確認された(図1B)。また、膜結合シアル酸の量を定量した。簡潔に説明すると、細胞を集め、PBSで1回洗浄し、針せん断と組み合わせて細胞溶解バッファー(50mM Tris/HCl pH8,150mM NaCl,0,5%NonidetP40,1mM EDTA,200μg/mlフェニルメチルスルホニルフルオリド)中で細胞溶解した。粗膜画分を40.000×gで20分間の遠心分離によりペレット化した。ペレットを水で2回洗浄し、凍結乾燥させた。上清を酵素活性測定に使用した(実施例3を参照されたい)。タンパク質濃度をRoti−Quant−Kit(Roth, Germany)で測定した。該ペレットを2M酢酸で80℃で1時間加水分解することにより、膜複合糖質結合シアル酸の含量を測定し、チオバルビツール酸法(Aminoff, Biochem. J. 81, 384-392, 1961)により遊離シアル酸を測定した。NM−F9及びNM−D4細胞内の膜結合シアル酸は、野生型細胞の場合と比較して、約3分の1に減少している(図1C、3B)。
【実施例3】
【0163】
NM−F9細胞の遺伝的欠損の分析
NM−F9細胞のTF露出は糖ヌクレオチド生合成経路又は輸送体におけるいくつかのシアリルトランスフェラーゼにおける欠損を要すると予想された。実施例1に記載のとおりに種々の酵素のmRNA発現分析を行い、この場合、NM−F9酵素に属するゲルバンドをNM−wtのものと比較し、UDP−GlcNAc−2−エピメラーゼmRNAの全欠如が示された(図4A)。ペーパークロマトグラフィーを使用して14C−UDP−N−アセチルグルコサミンから14C−N−アセチルマンノサミンへの変換を測定することにより、NM−wt及びNM−F9細胞におけるUDP−GlcNAc−2−エピメラーゼの酵素活性を細胞上清において測定した(実施例2を参照されたい)。1Uは37℃で1分当たり1μmolのManNAcの合成に相当する。UDP−GlcNAc−2−エピメラーゼ活性はNM−F9においては、NM−wt細胞における元の300μU/mgから完全に低下した(図4B)。
【0164】
シアル酸付加を復元するNM−F9細胞の能力を調べるために、培地に50mM ManNAcを添加した。4日間の培養後、残存しているシアル酸を含まないグライコトープ(glycotope)を、既に記載されているとおりにモノクローナル抗体A78−G/A7及びA63−C/A9並びにレクチンPNA、SNA及びMAAをアプライするフローサイトメトリーにより検出した。ManNAcを供給することによる代謝的相補は、検出試薬に応じて、潜在膜複合糖質TFの約50%(PNA)〜80%(A78−G/A7)をもたらした。UDP−GlcNAc−2−エピメラーゼ欠損のため、対照GlcNAcはNM−F9細胞により代謝され得なかった(図5)。
【0165】
また、遺伝的相補によるシアル酸付加の復元を調べた。UDP−GlcNAc−2−エピメラーゼcDNAをベクターpcDNA3.1内にクローニングした。SuperFect試薬を使用して、エピメラーゼをコードする構築物pcDNA3.1Zeo(−)/2EpiでNM−F9細胞を安定にトランスフェクトした。細胞を40%コンフルエンスまで増殖させた。5μgのDNAを150mlのDMEMに溶解し、30μlのSuperFectと混合した。室温で10分間のインキュベーションの後、この溶液を細胞に加えた。3時間後、培地を除去し、細胞をPBSで3回洗浄し、新鮮な増殖培地と共にインキュベートした。48時間のインキュベーション時間の後、増殖培地を再び、700μg/ml G418又は750μg/mlゼオシンを含有する培地と交換した。耐性クローンを、400μg/ml G418又は250μg/mlゼオシンを含有する増殖培地内で連続的に維持し、フローサイトメトリーにより分析した。シアル酸を含まないグライコトープを検出するために、モノクローナル抗体A78−G/A7及びA63−C/A9並びにレクチンPNA及びSNAを使用した。シアル酸付加度は完全に復元された(図6)。
【実施例4】
【0166】
NM−F9細胞における分泌性糖タンパク質の組換え発現
組換えヒト増殖因子rhGM−CSFを安定に発現するNM−F9細胞を、エレクトロポレーションを用いて発現ベクターpGT60hGM−CSF(Invivogen, USA)で該細胞をトランスフェクトすることにより作製した。エレクトロポレーションのために、106細胞を集め、洗浄し、400μlの低浸透圧バッファー(Eppendorf, Germany)に再懸濁させ、室温で15分間インキュベートした。ついで細胞懸濁液を8μgのプラスミドDNAと混合し、2mmの間隙を有するエレクトロポレーションキュベット(Eppendorf)内に移した。340Vの電圧及び5μ秒の時間を用いてMultiporator(Eppendorf)においてエレクトロポレーションを行った。培地(10%fcs,RPMI1640中の1%グルタミン,Biochrom)において、エレクトロポレーションからの回復の1〜2日後、安定にトランスフェクトされた細胞を、100μg/mlの抗生物質ヒグロマイシンBを使用することにより選択し、限界希釈法によりクローニングした。約4週間後、GM−CSF含量を定量するために使用しうるGM−CSF特異的ELISA(Becton-Dickenson, USA)を使用することにより、いくつかの細胞クローンの上清を分泌性rhGM−CSFの存在に関して分析した。rhGM−CSFの最高の分泌率を有する1つの細胞クローンを更なる実験のために選択した。第1の実験においては、GM−CSFを安定に発現する約105個のNM−F9細胞を、ウシ胎仔血清(fcs)の存在下及び非存在下にて、培地(前記参照)内で培養した。培地を交換することなく1、2、3及び4日間培養した後、時間経過と共に蓄積した分泌性rhGM−CSFの量を、GM−CSF特異的ELISAを用いることにより測定した。fcsの存在下で培養されたか非存在下で培養されたかには無関係に、4日間で約14ngのGM−CSFが105個の細胞により放出された(図7)。第1日の後の分泌率は、fcsの存在下で培養した場合(約6ng/ml)には、fcsの非存在下で培養した場合(約4ng/ml)と比べて若干高かった。
【0167】
発現ベクター構築物secGPAを使用して、ヒト膜タンパク質グリコホリンA(GPA)をNM−F9細胞内で分泌性アシアログリコホリンAとして組換え発現させた。secGPAは、膜貫通ドメイン及び細胞質尾部を含まないGPAのN末端の91アミノ酸(aa)をコードするcDNAを含有する。このcDNAを真核生物発現ベクターpcDNA5/FRT/V5−His−TOPO及び/又はpcDNA3.1/myc−His C(共にInvitrogen)内にサブクローニングした。さらに、内因性シグナルペプチドが異種ヒトシグナルペプチドにより置換された分泌性AGPAタンパク質の組換え融合タンパク質を発現させるために、NM−F9細胞を使用した。したがって、当業者に公知の分子生物学的方法を用いて、19個のN末端アミノ酸をコードするcDNAの代わりに、GM−CSF由来のシグナルペプチド(MWLQSLLLLGTVACSIS,secGM/GPA)、T細胞受容体由来のシグナルペプチド(MACPGFLWALVISTCLEFSMA,secTCR/GPA)又は抗体κ軽鎖由来のシグナルペプチド(METDTLLLWVLLLWVPPGSTGD,secAK/GPA)をコードする異種cDNAが組込まれたcDNAであるsecGM/GPA、secTCR/GPA及びsecAK/GPAを構築した。後者の2つのシグナルペプチドは、任意の遺伝子産物の分泌性発現を増強するための異種シグナルペプチドとしての使用に関して当業者に既に公知である。secGM/GPAの場合には、該シグナルペプチドとGPAをコードする骨格との間において追加的なアラニンが発現される。それらの3つのcDNA構築物を前記発現ベクター内にサブクローニングし、ヌクレオチド配列を確認した。得られた発現ベクターのそれぞれを、上述した手順、すなわちエレクトロポレーション、ヒグロマイシンBでの選択及び単一細胞クローニングを用いることにより、安定にトランスフェクトされたNM−F9細胞の作製に使用した。発現ベクターpcDNA5/FRT/V5−His−TOPOを使用する場合には、2つのFRTリコンビナーゼ認識部位により挟まれたlacZeo DNAカセット(Invitrogen)を含有する遺伝的に改変されたNM−F9細胞をpcDNA5/FRT/V5−His−TOPO及びリコンビナーゼ発現ベクターpOG44(Invitrogen)で共トランスフェクトした。任意のGPAを細胞培養上清から外へ捕捉するためのA83−CB12抗体及び細胞外ドメイン上に局在するTF抗原を認識するA63−C/A9抗体を使用するサンドイッチELISAにより、細胞培養培地内に放出された分泌性AGPAの分泌に関して、secGM/GPA、secTCR/GPA、secAK/GPA又はsecGPAを発現する細胞クローンをスクリーニングした。図8は、最高量の分泌性AGPA(1ml当たり及び105細胞当たり40ng/mlまで)を産生する細胞クローンがsecGM/GPA構築物で作製されたことを示している。その他の構築物を使用して、1ml当たり及び105細胞当たり最大で13ngの分泌性AGPAを検出することが可能であった。さらに、分泌性AGPAを産生する細胞クローンの数は、secGM/GPA構築物を発現ベクターとして使用した場合には、有意に、より高かった(図8)。
【実施例5】
【0168】
代謝的相補による示差的シアル酸付加
50mM ManNAcでのNM−F9細胞の代謝的相補により得られるシアル酸付加の部分的復元(図5)に関して、シアル酸付加に対する一定量の糖中間体の影響を詳細に調べた。NM−F9細胞を、0〜90mM ManNAcの存在下、fcsを含有する又は含有しない培地内で培養し、該細胞膜のシアル酸付加度をチオバルビツール酸法により分析した(実施例3を参照されたい)。細胞シアル酸付加度はfcsの非存在下又は存在下においてManNAcの量が増加することに伴って代謝的相補により制御されうることを示すことができた(図9)。fcs非含有培地におけるManNAcの添加無しでは、NM−F9細胞の膜画分においてはシアル酸は少量のみしか検出されなかった。fcs非含有培地に加えるManNAcの量を次第に増加させることによりシアル酸付加度を徐々に増加させることが可能であった(50mM ManNAcでプラトーに達した)。30mM ManNAcを添加したfcs含有培地においては、シアル酸付加における更なる増加が達成された。fcs含有培地に90mM ManNAcを添加した場合には、元のNM−wt細胞に匹敵するくらい高い、最高のシアル酸付加度が得られた。種々の濃度のManNAcでの代謝的相補による膜タンパク質の示差的シアル酸付加を、シアル酸を含まないTFエピトープを認識し従ってシアル酸付加度に逆相関するレクチンPNAを使用するフローサイトメトリーにより分析した。細胞膜内のシアル酸の量だけでなく膜内の糖タンパク質上のシアル酸付加度も種々の量のManNAc量での代謝的相補により制御可能であることが示されている(図10)。培地内に存在するManNAcを次第に増加させることにより、次第に増加するシアル酸付加度が観察された。90mM ManNAcを用いた場合、NM−F9細胞における糖タンパク質のシアル酸付加度はNM−wt細胞の場合とほぼ同等に高かった。
【0169】
最後に、分泌性タンパク質(この場合はrhGM−CSF)のシアル酸付加度もManNAc及び血清タンパク質での代謝的相補により制御可能であるかどうかを試験した。rhGM−CSF発現ベクターで安定にトランスフェクトされたNM−F9細胞(実施例4を参照されたい)を再び、漸増量の0〜90mM ManNAcと共に、並びに1及び3mg/mlのシアロ糖タンパク質フェツインの存在下の90mM ManNAcと共に培養した。4日後、得られた細胞培養上清中のrhGM−CSFの濃度をELISAにより測定し(実施例4を参照されたい)、ついで細胞培養培地1ml当たり100ngに調節した。シアル酸を含まないrhGM−CSFの度合を測定するために、サンドイッチELISAを確立した。簡潔に説明すると、GM−CSF特異的ELISA(Becton-Dickenson)の抗ヒトGM−CSFモノクローナル抗体をrhGM−CSFの捕捉に使用し、したがってPBS中1:250の希釈度でマイクロタイタープレートに4℃で一晩コートした。PBS/0.05%Tween20中での洗浄及びPBS中の10%fcsでのブロッキングの後、100μlの該細胞培養上清を該マイクロタイタープレートに加え、室温で2時間インキュベートした。シアル酸を含まないO−グリカンをビオチン化PNA(5〜10μg/ml)及びペルオキシダーゼ結合ストレプトアビジン(1:250希釈物)で検出した。分泌性組換え糖タンパク質、例えばrhGM−CSFも、漸増量のManNAcによるNM−F9細胞の相補により示差的にシアル酸付加されうることが示された(図11)。90mM ManNAcへの1mg/mlフェツインの添加により最高のシアル酸付加度が達成される。90mM ManNAcへの3mg/mlフェツインの添加はシアル酸付加における更なる増強をもたらさない。
【実施例6】
【0170】
種々のシアル酸付加形態の活性の測定による高活性ヒトGM−CSFの同定
種々のシアル酸付加タンパク質の活性を評価するために、GM−CSFの増殖刺激能を採用した。rhGM−CSFを発現するNM−F9細胞(実施例4及び5を参照されたい)を、0、30、50、70及び90mM ManNAcで並びに1mg/mlのフェツインと共に90mM ManNAcを添加した無血清培地を含有する種々の調製物において培養した。実施例5に既に記載されているとおりにシアル酸付加を検出した。TF−1細胞及び単球性樹状細胞系NemodDC(それらの増殖速度は共にGM−CSFに左右される)を、種々のシアル酸付加rhGM−CSFが5ng/mlに調節された細胞上清と共にそれぞれ48時間又は24時間インキュベートした。細胞増殖を、製造業者のプロトコール(Roche Diagnostik GmbH, Mannheim, Germany)に従いBrdU−増殖アッセイにより測定した。固定後、細胞をPOD標識抗BrdU抗体と共にインキュベートした。後続の染色反応を1M硫酸により停止させ、450nmの吸光度(参照690nm)で分光光度法により検出した。GM−CSFを含有しないNM−F9上清を陰性対照として使用した。それぞれ酵母及び大腸菌内で発現された非シアル酸付加rhGM−CSF形態である市販のLeukine(登録商標)(Schering AG)及びLeukomax(登録商標)(Schering-Plough)を基準として使用した。
【0171】
驚くべきことに、rhGM−CSFの細胞増殖刺激活性はrhGM−CSFのシアル酸付加度によって大きく異なり、90mM ManNAcを含有する培養から得られた、最高ではないが高度にシアル酸付加されたrhGM−CSFの1つのシアル酸付加形態が最高活性を示した。このrhGM−CSFは、Leukine(登録商標)(100%)と比較した場合に、TF−1細胞及びNemodDCに関して約178±10%の増殖刺激活性の増加を示した(図12及び13)。市販のrhGM−CSFであるLeukomax(登録商標)及びLeukine(登録商標)の細胞増殖活性は、NM−F9細胞内で発現された最も活性なシアル酸付加形態より低いことが判明した(図12)。5ng/mlのLeukomax(登録商標)又はLeukine(登録商標)で観察された最高の増殖活性は、0.01ng/mlの、NM−F9細胞からの最適シアル酸付加形態により達成される(Leukine(登録商標)又はLeukomax(登録商標)と比較して約500倍低いGM−CSF;図14)。同じ濃度のLeukine(登録商標)、Leukomax(登録商標)又はrhGM−CSFの最適シアル酸付加形態で該活性を分析したところ、活性の増加は5倍までであった(図14)。
【0172】
抗ヒトGM−CSF抗体BVD2−23 B6をTF1細胞増殖アッセイに加えた場合には、GM−CSF活性が特異的にブロッキングされた。5μg/mlの抗体濃度では既に、Leukine(登録商標)又はrhGM−CSFの最適シアル酸付加形態によるTF−1細胞増殖の刺激は約60%ブロッキングされ、40μg/mlの抗体濃度では完全に消失した(図15)。
【0173】
配列表:
配列番号1:メチオニン、トリプトファン、ロイシン、グルタミン、セリン、ロイシン、ロイシン、ロイシン、ロイシン、グリシン、トレオニン、バリン、アラニン、システイン、セリン、イソロイシン、セリン。
【表1】
【表2】
【特許請求の範囲】
【請求項1】
シアル酸の糖ヌクレオチド生合成経路における、細胞のシアル酸付加能の低下を引き起こす少なくとも1つの欠損を含有し、かつ糖タンパク質をコードする核酸でトランスフェクトされた発現細胞系において、所望の濃度の少なくとも1つのシアル酸前駆体添加物を添加した培地内で糖タンパク質を発現させるための、少なくとも1つのシアル酸前駆体添加物の所望の濃度を決定する方法であって、該シアル酸前駆体添加物が該欠損を復元して該糖タンパク質のシアル酸付加をもたらす天然又は合成化合物であり、
(i)種々の濃度の少なくとも1つのシアル酸前駆体を使用する示差的シアル酸付加により、複数の異なるシアル酸付加形態の糖タンパク質を発現させること、
及び
(ii)適当なバイオアッセイにおいて、参照糖タンパク質と比較して、前記の異なるシアル酸付加形態の活性を測定すること、
及び
(iii)より高い/最高の活性を有するシアル酸付加形態を選択し、該糖タンパク質のより高い/最高の活性レベルと相関する、該シアル酸前駆体添加物の濃度を決定すること
を含む方法。
【請求項2】
ただ1つのシアル酸付加形態がステップ(i)において発現される、請求項1に記載の方法。
【請求項3】
該糖タンパク質のより高い活性が、少なくとも1つのin vitroモデルにおけるより高い活性、及び/又は少なくとも1つのin vivoモデルにおけるより高い活性、及び/又はより高い安定性、及び/又はより長い血清半減期、及び/又はより長いバイオアベイラビリティ、及び/又は免疫原性の改善、及び/又は抗原性の改善(少なくとも1つのバイオアッセイにより測定される)を特徴とする、請求項1又は2に記載の方法。
【請求項4】
糖タンパク質の製造方法であって、
シアル酸の糖ヌクレオチド生合成経路における、細胞のシアル酸付加能の低下を引き起こす少なくとも1つの欠損を含有し、かつ糖タンパク質をコードする核酸でトランスフェクトされた発現細胞系において、請求項1〜3のいずれか1項に記載の方法により決定された濃度の少なくとも1つのシアル酸前駆体添加物を添加した培地内で糖タンパク質を発現させることを含み、該シアル酸前駆体添加物が該欠損を復元して該糖タンパク質のシアル酸付加をもたらす天然又は合成化合物である、前記方法。
【請求項5】
部分的にシアル酸付加された糖タンパク質が生成される、請求項4に記載の方法。
【請求項6】
糖タンパク質が発現細胞系の細胞により分泌される、請求項1〜5のいずれか1項に記載の方法。
【請求項7】
シアル酸の生合成経路における欠損が、CMP−シアル酸輸送体を含む、糖輸送に関与するタンパク質、又は、キナーゼ、デヒドロゲナーゼ、ホスファターゼ、シンテターゼ、トランスケトラーゼ、トランスアルドラーゼ、イソメラーゼ、トランスフェラーゼ及びエピメラーゼを含む、CMP−シアル酸の生合成に関与するタンパク質、の機能喪失である、請求項1〜6のいずれか1項に記載の方法。
【請求項8】
シアル酸の生合成経路における欠損がエピメラーゼの突然変異である、請求項1〜7のいずれか1項に記載の方法。
【請求項9】
発現細胞系がNM−F9(受託番号DSM ACC2606)又はNM−D4(受託番号DSM ACC2605)である、請求項1〜8のいずれか1項に記載の方法。
【請求項10】
糖タンパク質が、グリコホリンA、EPO、G−CSF、GM−CSF、FSH、hCG、LH、インターフェロン、インターロイキン、抗体及び/又はそれらのフラグメントを含む群より選ばれるものである、請求項1〜9のいずれか1項に記載の方法。
【請求項11】
少なくとも1つのシアル酸前駆体添加物がManNAc、アセチル化ManNAc、パーアセチル化ManNAc又はフェツインである、請求項1〜10のいずれか1項に記載の方法。
【請求項12】
天然シアル酸修飾を有する糖タンパク質を生じるシアル酸前駆体添加物を使用する、請求項1〜11のいずれか1項に記載の方法。
【請求項13】
欠損が、UDP−N−アセチルグルコサミン−2−エピメラーゼの酵素活性の減少又は不在を引き起こす、請求項1〜12のいずれか1項に記載の方法。
【請求項14】
活性糖タンパク質の同定/決定方法であって、
i)シアル酸の糖ヌクレオチド生合成経路における、細胞のシアル酸付加能の低下を引き起こす少なくとも1つの欠損を含有する発現細胞系を、該糖タンパク質をコードする核酸でトランスフェクトすること、
及び
ii)種々の濃度の少なくとも1つのシアル酸前駆体添加物を含有する培地を使用する示差的シアル酸付加により、複数の異なるシアル酸付加形態の糖タンパク質を発現させることであって、該シアル酸前駆体添加物が該欠損を復元して該糖タンパク質のシアル酸付加をもたらす天然又は合成化合物であり、
及び
iii)適当なバイオアッセイにおいて、参照糖タンパク質と比較して、前記の異なるシアル酸付加形態の活性を測定すること、
及び
iv)より高い/最高の活性を有するシアル酸付加形態を選択すること
を含む方法。
【請求項15】
α2−3及びα2−6結合シアル酸によりシアル酸付加されうる脱シアル酸構造に結合する認識分子を用いて初代細胞又は細胞系から発現細胞系を選択することを含む、シアル酸の糖ヌクレオチド生合成経路における、細胞のシアル酸付加能の低下を引き起こす欠損を有する発現細胞系の作製方法。
【請求項16】
選択前に初代細胞又は細胞系からの細胞に突然変異誘発を行う、請求項15に記載の方法。
【請求項17】
構造がO−グリカンである、請求項15又は16に記載の方法。
【請求項18】
認識分子がレクチン又は糖鎖特異的抗体である、請求項15〜17のいずれか1項に記載の方法。
【請求項19】
認識分子がレクチン又はコア−1構造を認識する糖鎖特異的抗体である、請求項15〜18のいずれか1項に記載の方法。
【請求項20】
発現細胞系が、Per.C6、HEK293、K562、CV1、COS−7、ハイブリドーマ細胞、ナマルワ(Namalwa)、BHK及びCHOを含む群から誘導される、請求項15〜19のいずれか1項に記載の方法。
【請求項21】
ヒト細胞系NM−F9(DSM ACC2606)又はNM−D4(DSM ACC2605)。
【請求項22】
細胞系NM−F9(DSM ACC2606)又はNM−D4(DSM ACC2605)の細胞において糖タンパク質を発現させることを含む、糖タンパク質の発現方法。
【請求項23】
細胞系NM−F9(DSM ACC2606)の細胞において90mM ManNAcを添加した培地内でGM−CSFを発現させることを含む方法により生成されるGM−CSF糖タンパク質。
【請求項1】
シアル酸の糖ヌクレオチド生合成経路における、細胞のシアル酸付加能の低下を引き起こす少なくとも1つの欠損を含有し、かつ糖タンパク質をコードする核酸でトランスフェクトされた発現細胞系において、所望の濃度の少なくとも1つのシアル酸前駆体添加物を添加した培地内で糖タンパク質を発現させるための、少なくとも1つのシアル酸前駆体添加物の所望の濃度を決定する方法であって、該シアル酸前駆体添加物が該欠損を復元して該糖タンパク質のシアル酸付加をもたらす天然又は合成化合物であり、
(i)種々の濃度の少なくとも1つのシアル酸前駆体を使用する示差的シアル酸付加により、複数の異なるシアル酸付加形態の糖タンパク質を発現させること、
及び
(ii)適当なバイオアッセイにおいて、参照糖タンパク質と比較して、前記の異なるシアル酸付加形態の活性を測定すること、
及び
(iii)より高い/最高の活性を有するシアル酸付加形態を選択し、該糖タンパク質のより高い/最高の活性レベルと相関する、該シアル酸前駆体添加物の濃度を決定すること
を含む方法。
【請求項2】
ただ1つのシアル酸付加形態がステップ(i)において発現される、請求項1に記載の方法。
【請求項3】
該糖タンパク質のより高い活性が、少なくとも1つのin vitroモデルにおけるより高い活性、及び/又は少なくとも1つのin vivoモデルにおけるより高い活性、及び/又はより高い安定性、及び/又はより長い血清半減期、及び/又はより長いバイオアベイラビリティ、及び/又は免疫原性の改善、及び/又は抗原性の改善(少なくとも1つのバイオアッセイにより測定される)を特徴とする、請求項1又は2に記載の方法。
【請求項4】
糖タンパク質の製造方法であって、
シアル酸の糖ヌクレオチド生合成経路における、細胞のシアル酸付加能の低下を引き起こす少なくとも1つの欠損を含有し、かつ糖タンパク質をコードする核酸でトランスフェクトされた発現細胞系において、請求項1〜3のいずれか1項に記載の方法により決定された濃度の少なくとも1つのシアル酸前駆体添加物を添加した培地内で糖タンパク質を発現させることを含み、該シアル酸前駆体添加物が該欠損を復元して該糖タンパク質のシアル酸付加をもたらす天然又は合成化合物である、前記方法。
【請求項5】
部分的にシアル酸付加された糖タンパク質が生成される、請求項4に記載の方法。
【請求項6】
糖タンパク質が発現細胞系の細胞により分泌される、請求項1〜5のいずれか1項に記載の方法。
【請求項7】
シアル酸の生合成経路における欠損が、CMP−シアル酸輸送体を含む、糖輸送に関与するタンパク質、又は、キナーゼ、デヒドロゲナーゼ、ホスファターゼ、シンテターゼ、トランスケトラーゼ、トランスアルドラーゼ、イソメラーゼ、トランスフェラーゼ及びエピメラーゼを含む、CMP−シアル酸の生合成に関与するタンパク質、の機能喪失である、請求項1〜6のいずれか1項に記載の方法。
【請求項8】
シアル酸の生合成経路における欠損がエピメラーゼの突然変異である、請求項1〜7のいずれか1項に記載の方法。
【請求項9】
発現細胞系がNM−F9(受託番号DSM ACC2606)又はNM−D4(受託番号DSM ACC2605)である、請求項1〜8のいずれか1項に記載の方法。
【請求項10】
糖タンパク質が、グリコホリンA、EPO、G−CSF、GM−CSF、FSH、hCG、LH、インターフェロン、インターロイキン、抗体及び/又はそれらのフラグメントを含む群より選ばれるものである、請求項1〜9のいずれか1項に記載の方法。
【請求項11】
少なくとも1つのシアル酸前駆体添加物がManNAc、アセチル化ManNAc、パーアセチル化ManNAc又はフェツインである、請求項1〜10のいずれか1項に記載の方法。
【請求項12】
天然シアル酸修飾を有する糖タンパク質を生じるシアル酸前駆体添加物を使用する、請求項1〜11のいずれか1項に記載の方法。
【請求項13】
欠損が、UDP−N−アセチルグルコサミン−2−エピメラーゼの酵素活性の減少又は不在を引き起こす、請求項1〜12のいずれか1項に記載の方法。
【請求項14】
活性糖タンパク質の同定/決定方法であって、
i)シアル酸の糖ヌクレオチド生合成経路における、細胞のシアル酸付加能の低下を引き起こす少なくとも1つの欠損を含有する発現細胞系を、該糖タンパク質をコードする核酸でトランスフェクトすること、
及び
ii)種々の濃度の少なくとも1つのシアル酸前駆体添加物を含有する培地を使用する示差的シアル酸付加により、複数の異なるシアル酸付加形態の糖タンパク質を発現させることであって、該シアル酸前駆体添加物が該欠損を復元して該糖タンパク質のシアル酸付加をもたらす天然又は合成化合物であり、
及び
iii)適当なバイオアッセイにおいて、参照糖タンパク質と比較して、前記の異なるシアル酸付加形態の活性を測定すること、
及び
iv)より高い/最高の活性を有するシアル酸付加形態を選択すること
を含む方法。
【請求項15】
α2−3及びα2−6結合シアル酸によりシアル酸付加されうる脱シアル酸構造に結合する認識分子を用いて初代細胞又は細胞系から発現細胞系を選択することを含む、シアル酸の糖ヌクレオチド生合成経路における、細胞のシアル酸付加能の低下を引き起こす欠損を有する発現細胞系の作製方法。
【請求項16】
選択前に初代細胞又は細胞系からの細胞に突然変異誘発を行う、請求項15に記載の方法。
【請求項17】
構造がO−グリカンである、請求項15又は16に記載の方法。
【請求項18】
認識分子がレクチン又は糖鎖特異的抗体である、請求項15〜17のいずれか1項に記載の方法。
【請求項19】
認識分子がレクチン又はコア−1構造を認識する糖鎖特異的抗体である、請求項15〜18のいずれか1項に記載の方法。
【請求項20】
発現細胞系が、Per.C6、HEK293、K562、CV1、COS−7、ハイブリドーマ細胞、ナマルワ(Namalwa)、BHK及びCHOを含む群から誘導される、請求項15〜19のいずれか1項に記載の方法。
【請求項21】
ヒト細胞系NM−F9(DSM ACC2606)又はNM−D4(DSM ACC2605)。
【請求項22】
細胞系NM−F9(DSM ACC2606)又はNM−D4(DSM ACC2605)の細胞において糖タンパク質を発現させることを含む、糖タンパク質の発現方法。
【請求項23】
細胞系NM−F9(DSM ACC2606)の細胞において90mM ManNAcを添加した培地内でGM−CSFを発現させることを含む方法により生成されるGM−CSF糖タンパク質。
【図1】

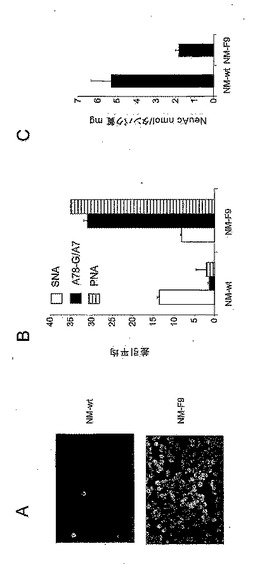
【図2】

【図3】

【図4】

【図5】
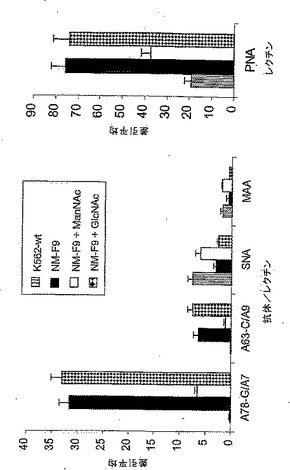
【図6】
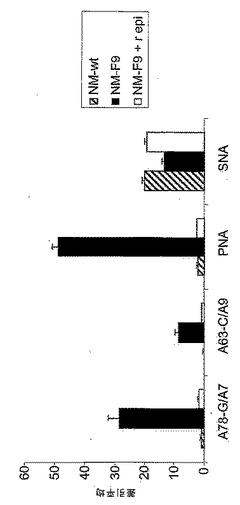
【図7】

【図8】

【図9】

【図10】
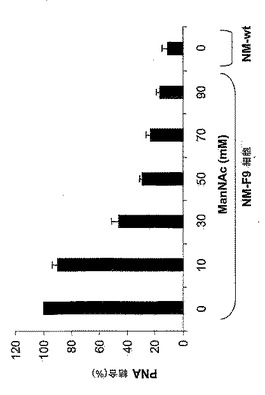
【図11】

【図12】
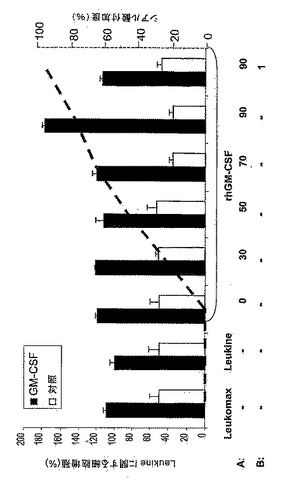
【図13】
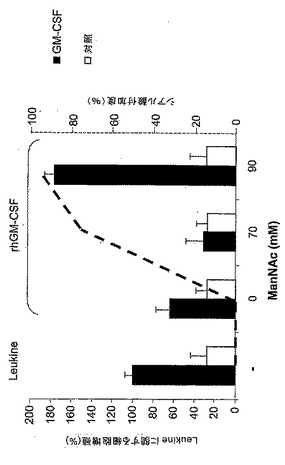
【図14】
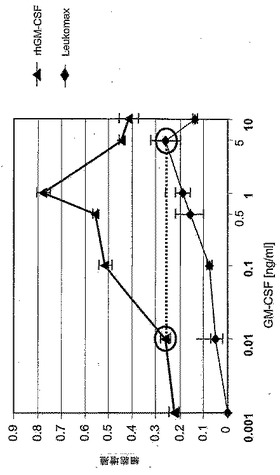
【図15】
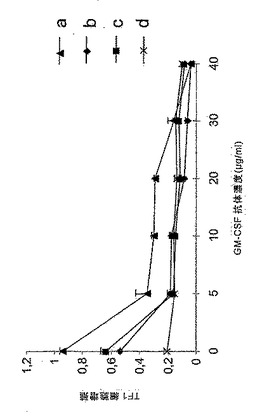
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【公開番号】特開2012−87123(P2012−87123A)
【公開日】平成24年5月10日(2012.5.10)
【国際特許分類】
【出願番号】特願2011−229367(P2011−229367)
【出願日】平成23年10月19日(2011.10.19)
【分割の表示】特願2006−552577(P2006−552577)の分割
【原出願日】平成17年2月14日(2005.2.14)
【出願人】(506274888)グリコトープ ゲーエムベーハー (2)
【Fターム(参考)】
【公開日】平成24年5月10日(2012.5.10)
【国際特許分類】
【出願日】平成23年10月19日(2011.10.19)
【分割の表示】特願2006−552577(P2006−552577)の分割
【原出願日】平成17年2月14日(2005.2.14)
【出願人】(506274888)グリコトープ ゲーエムベーハー (2)
【Fターム(参考)】
[ Back to top ]