
Blimp1およびレポーター分子を共発現する改変細胞およびその使用方法
【課題】発現されるときにレポーター分子と共発現されるポリペプチドを産生する改変遺伝物質を含む、遺伝子改変細胞またはそのような細胞を含む非ヒト生物を提供する。
【解決手段】ここでポリペプチドは造血細胞の最終分化に関連する。遺伝物質遺伝子は、Blimp対立遺伝子またはその一部、フラグメントもしくは機能的な形態である。さらに、B細胞系列細胞におけるレポーター分子の同定は、そのような細胞がASCへの分化に決定付けされている、またはASCに分化したことを示す。または、T細胞系列の細胞におけるレポーター分子活性は、これらの細胞が活性化されていることを示す。従って、リンパ球におけるBlimpの存在は、細胞が最終分化する、または最終分化に決定付けられていることを示す。例示的なT細胞としては、CD4+T細胞およびCD8+T細胞が含まれ、かつ例示的なB細胞は、ASCである。
【解決手段】ここでポリペプチドは造血細胞の最終分化に関連する。遺伝物質遺伝子は、Blimp対立遺伝子またはその一部、フラグメントもしくは機能的な形態である。さらに、B細胞系列細胞におけるレポーター分子の同定は、そのような細胞がASCへの分化に決定付けされている、またはASCに分化したことを示す。または、T細胞系列の細胞におけるレポーター分子活性は、これらの細胞が活性化されていることを示す。従って、リンパ球におけるBlimpの存在は、細胞が最終分化する、または最終分化に決定付けられていることを示す。例示的なT細胞としては、CD4+T細胞およびCD8+T細胞が含まれ、かつ例示的なB細胞は、ASCである。
【発明の詳細な説明】
【技術分野】
【0001】
発明の背景
発明の分野
本発明は、一般に、特定の系列の造血細胞およびそれらの分化段階を同定するモデルシステムに関する。さらに詳細には、本発明は、一般に、同定可能なシグナルを惹起することができるレポーター分子を共産生する(co-produce)ように改変された最終分化の遺伝子マーカーを有する、遺伝子改変細胞およびそのような細胞を含む非ヒト動物、ならびに、限定されるわけではないが、発達中の胚細胞、異常分化を伴う細胞(例えば、癌細胞)および造血細胞系列の細胞(例えば、Bおよび/またはT細胞など)などの細胞の分化またはトランスフォーメーション状態を調節することができる分子を同定する際のそれらの使用を提供する。同定された分子は、治療および予防用の薬学的組成物の基礎となる。
【背景技術】
【0002】
先行技術の説明
本明細書において参照する文献の詳細は、本明細書の末尾にも記載する。
【0003】
本明細書中のいずれの先行技術への参照も、この先行技術が、任意の国における共通一般知識の一部をなすことの承認またはいずれの形態の提案とも解釈されず、また解釈すべきでない。
【0004】
細胞寿命は、細胞分裂、分化、形態発生およびアポトーシスの変更を指示する無数の選択肢としての、および高度に調節された生化学経路を含む。分裂および/または分化する細胞の潜在能力は、多種多様である。例えば、胚は、任意の細胞タイプに分化する能力を保持する全能細胞を含む。幹細胞を含む他の細胞タイプは、多能性であり、全てではないが一定範囲の細胞表現型に最終的に分化することができる。一つの最終形態に決定付けられることとなる細胞もあり、これらは、最終分化する。
【0005】
最終分化細胞への細胞の正常な突然変異を阻害するまたはアポトーシスを防止する変化は、分化または細胞死を伴わない無制御細胞分裂を特徴とする腫瘍発現の引き金の役割を果たす。従って、分化および正常なアポトーシスを促進する薬剤により、腫瘍発現のスイッチを切ることができる。
【0006】
特定の細胞タイプの最終分化期間の間に発現される分子は、熱心に研究されてきた。しかし、分子レベルでのこの期間の間の事象の順序を理解するには、この発達段階に発現される分子の時間的および空間的発現パターンを理解する必要がある。
【0007】
Bリンパ球誘導成熟因子(Blimp)は、98kDa転写因子であり、これは、当初、B細胞リンパ腫細胞系の分化の間に誘導される因子として同定された(Turner et al., Cell 77: 297,1994(非特許文献1))。ヒト細胞由来の対応する因子は、PRDM-1と呼ばれる。Blimp-1は、B細胞最終分化の調節に顕著な役割を有することが報告されている。具体的には、Blimp-1は、ヒトおよびマウスからの抗体分泌細胞(ASC)において発現されるが、記憶細胞においては発現されない(Angelin-Duclos et al., J Immunol 165 : 5462, 2000(非特許文献2))。Blimp-1の異所発現は、リンパ腫および初代B細胞をASC細胞に最終分化させるために十分である(Turner et al.,(前記)(非特許文献1), Schliephake et al., Eur J Immunol 26 : 268, 1996(非特許文献3); Messika et al., J Exp Med 188 : 515, 1998(非特許文献4) ; Knodel et al., Eur J Immunol 31:1972, 2001(非特許文献5))。アンチセンスまたは優性干渉アプローチによるBlimp-1の発現の阻害により、完全ASC分化には不可欠であると考えられる細胞周期退出が抑制される(Soro et al., J Immunol 163 : 611, 1999(非特許文献6); Angelin-Duclos et al., J Immunol 165:5462, 2000(非特許文献7); Johnson et al., Eur J Immunol 32:3765, 2002(非特許文献8))。また、B細胞にBlimp-1がないマウスは、免疫グロブリンをほとんど産生せず、著しく低減されたASC区画を有する。(Shapiro-Shelef et al., Immunity 19 :607, 2003.(非特許文献9))。
【0008】
Blimp-1は、B細胞系列の細胞でしか産生されないと当初報告されたが、Blimp-1が骨髄系分化の間にも産生されることは、今では明らかである(Keller et al., Genes Dev 5:868, 1991(非特許文献10), Chang et al., Nat Immunol 1: 169, 2000(非特許文献11))。Blimp-1は、骨髄分化に関与するc-mycの抑制に必要とされる(Chang et al.,(前記), 2000(非特許文献11); Marcu et al., Annu Rev Biochem 61:809, 1992(非特許文献12))。U937細胞におけるBlimp-1の過剰産生は、例えば、マクロファージ分化の誘導に十分である(Chang et al., (前記), 2000(非特許文献11))。従って、マクロファージおよびB細胞におけるBlimp-1によるc-mycの抑制が、これら二系列における最終分化の特徴である。Blimp-1は、マウスおよびアフリカツメガエルの胚発達の間にも広範に産生される(de Souza et al., Embo J 18:6062, 1999(非特許文献13); Rosenbaum et al., Embo J, 9:897, 1990(非特許文献14))。
【0009】
Bリンパ球は、中でも最も集中的に研究された胚細胞型であるが、B細胞発達の初期段階は、比較的よく特性付けされている一方で、ASCへのBリンパ球の最終分化を制御するプロセスについてはほとんど判っていない。ASC(形質細胞)は、体液免疫応答の直接的伝達因子である。これらは、防御免疫に不可欠な血清免疫グロブリンを大量に分泌する。従って、ASCへのBリンパ球の最終分化は、強い治療的関心の対象となっている。例えば、ASCへの最終分化は、有効なワクチン接種戦略における非常に重要な要素である。さらに、多発性骨髄腫は、ASCの分化経路完了の失敗により生じる。
【0010】
しかし、ASCは、主として骨髄および脾臓にある非常に限定された細胞の非常に珍しい集団の代表である。マウスおよびヒトにおけるASC集団は、不均一な寿命および細胞表面表現型の細胞を含み、そのため、純粋なASCの決定的で見込みのある単離を行うことができない(Fong et al., Proc Natl Acad Sci U S A 11 : 11, 2003(非特許文献15); Medina et al., Blood 99. 2154, 2002(非特許文献16); O'Connor et al., J.Exp Med 195:737, 2002(非特許文献17); Manz et al., Curr Opin Immunol 14 : 517, 2002(非特許文献18); Underhill et al., Blood 24:24, 2003(非特許文献19))。
【0011】
T細胞最終分化プログラムは、ほとんど理解されていない(Sprent et al., Immunol Lett 85 : 145-149, 2003(非特許文献20))。感染に反応して、抗原特異的T細胞は、エフェクター細胞に分化し、大規模にクローン増幅される。T細胞数の恒常性は、その後の収縮段階により維持され、この段階では、エフェクター細胞の90%より多くが排除され、小率が記憶T細胞になる(Sprent et al., Annu Rev Immunol 20 : 551-579,2002(非特許文献21))。収縮は、感染の量または継続段階間に依存するので、このプロセスは、遺伝的制御下にあると提案された(Badovinac et al., Nat Immunol 5 : 809-817, 2004(非特許文献22); Badovinac et al., Nat Immunol 3: 619-626, 2002(非特許文献23))。T調節細胞の欠如のため(Khattri et al., Nat Immunol 4: 337-342,2003(非特許文献24); Hori et al., Science 299 : 1057-1061, 2003(非特許文献25); Fontenot et al., Nat Immunol 4 : 330-336,2003(非特許文献26))、下方調節性分子CTLA-4の喪失のため(Chambers et al., Immunity 7 : 885-895, 1997(非特許文献27))または自己免疫を生じさせる非肥満糖尿病(NOD)マウスでは遺伝子欠失のために増幅が増進した場合、T細胞数を制御する能力は不可欠である。
【0012】
広範な状況下および様々な刺激下でASC、T細胞および造血系の他の細胞の最終分化をモニタリングすることができることは、異常分化を随伴する一定範囲の状態(例えば、癌、自己免疫疾患)の治療および/または予防に使用するための戦略および試薬、または適切な免疫応答の発生の場合など正常な発生プログラムを利用する戦略および試薬を開発する際に極めて価値があるだろう。
【先行技術文献】
【非特許文献】
【0013】
【非特許文献1】Turner et al., Cell 77: 297,1994
【非特許文献2】Angelin-Duclos et al., J Immunol 165 : 5462, 2000
【非特許文献3】Schliephake et al., Eur J Immunol 26 : 268, 1996
【非特許文献4】Messika et al., J Exp Med 188 : 515, 1998
【非特許文献5】Knodel et al., Eur J Immunol 31:1972, 2001
【非特許文献6】Soro et al.,J Immunol 163 : 611, 1999
【非特許文献7】Angelin-Duclos et al., J Immunol 165:5462, 2000
【非特許文献8】Johnson et al., Eur J Immunol 32:3765, 2002
【非特許文献9】Shapiro-Shelef et al., Immunity 19 :607, 2003.
【非特許文献10】Keller et al., Genes Dev 5:868, 1991
【非特許文献11】Chang et al., Nat Immunol 1: 169, 2000
【非特許文献12】Marcu et al., Annu Rev Biochem 61:809, 1992
【非特許文献13】de Souza et al., Embo J 18:6062, 1999
【非特許文献14】Rosenbaum et al., Embo J, 9:897, 1990
【非特許文献15】Fong et al.,Proc Natl Acad Sci U S A 11 : 11, 2003
【非特許文献16】Medina et al., Blood 99. 2154, 2002
【非特許文献17】O'Connor et al.,J.Exp Med 195:737, 2002
【非特許文献18】Manz et al.,Curr Opin Immunol 14 : 517, 2002
【非特許文献19】Underhill et al., Blood 24:24, 2003
【非特許文献20】Sprent et al., Immunol Lett 85 : 145-149, 2003
【非特許文献21】Sprent et al., Annu Rev Immunol 20 : 551-579,2002
【非特許文献22】Badovinac et al., Nat Immunol 5 : 809-817, 2004
【非特許文献23】Badovinac et al., Nat Immunol 3: 619-626, 2002
【非特許文献24】Khattri et al., Nat Immunol 4: 337-342,2003
【非特許文献25】Hori et al., Science 299 : 1057-1061, 2003
【非特許文献26】Fontenot et al., Nat Immunol 4 : 330-336,2003
【非特許文献27】Chambers et al., Immunity 7 : 885-895,1997
【発明の概要】
【0014】
本明細書を通して、その文脈が別様に求めていない限り、語「含む(comprise)」、または「含む(comprises)」もしくは「含んでいる(comprising)」などの語尾変化したものは、示されている要素もしくは整数または要素もしくは整数の群の包含を意味するが、任意の他の要素もしくは整数または要素もしくは整数の群の排除は意味しないことは理解されるだろう。
【0015】
ヌクレオチドおよびアミノ酸配列は、配列識別子番号(配列番号:)によって参照する。配列番号:は、<400>1(配列番号:1)、<400>2(配列番号:2)など、配列識別子に数値対応している。配列識別子の一覧を表1に提供する。配列表は、添付されている。
【0016】
遺伝子および他の遺伝物質(例えば、mRNA、構築物など)は、イタリック体で表しており、それらのタンパク質性発現産物は、イタリック体でない形で表す。従って、転写因子Blimpは、Blimpの発現産物である。用語「Blimp」または「Blimp」は、ヒト相同体を含む任意の動物または哺乳類に由来する全ての相同体または変異体分子を示すために用いている。従って、ヒトPRDM-1およびその産物、PRDM-1は、用語BlimpまたはBlimpに包含される。別様に述べられていない限り、Blimpへの言及は、そのポリペプチドの機能的な形態への言及であり、改変Blimpへの言及は、Blimpの機能的な形態をコードする遺伝子または対立遺伝子配列への言及である。
【0017】
本発明は、造血および胚細胞の分化におけるBlimpの役割の同定に、一部、基づいている。Blimpの存在またはBlimpのレベルについてスクリーニングすることにより、細胞の最終分化の段階について判定を下すことができる。Blimpの役割の同定により、ASC(形質細胞)などであるが、これに限定されるわけではない、造血細胞の実質的に均質な集団を同定することがさらに可能となる。
【0018】
より詳細には、本発明は、Blimpおよびレポーター分子をコードする遺伝物質を共発現する細胞を含む、遺伝子改変細胞またはインビボもしくはインビトロ系を提供する。リンパ球系列などであるが、これに限定されるわけではない、造血系列の細胞におけるレポーター活性の検出は、レポーター活性を有するおよび機能的Blimpを産生する細胞が、最終分化に決定付けられていることの指標となる。従って、機能的Blimpを産生するレポーター活性B細胞の検出は、これらの細胞が、抗体分泌細胞(ASC)への分化に決定付けられていることの指標となる。また、本明細書において記載のように、機能的Blimpを発現するT細胞におけるレポーター活性の検出は、これらの細胞が、活性化CD4+ T細胞またはエフェクターCD8+ T細胞などの活性化/記憶T細胞であることの指標となる。従って、本発明は、様々な条件下または様々な刺激もしくは薬剤の存在下での特定の細胞の分化またはトランスフォーメーション状態のモニタリングを助長する、遺伝子改変細胞またはそのような細胞を含む非ヒト動物を提供する。本発明は、発達中の幹細胞を含む胚細胞、異常分化を伴う細胞(例えば、癌細胞)、および、例えばBおよび/またはT細胞など造血細胞系列の細胞であるが、これらに限定されない、細胞の分化またはトランスフォーメーション状態を調節することができる分子を同定するための、ハイスループットスクリーニング法を含むスクリーニング法をさらに提供する。
【0019】
具体的には、遺伝子改変細胞またはそのような細胞を含む非ヒト生物を本発明によって提供する。一つの態様において、本細胞は、レポーター分子をコードするように改変されたmRNAから翻訳されたBlimpを産生する。好ましくは、そのレポーター分子コード配列が、Blimp対立遺伝子のイントロンに挿入される。この改変Blimp対立遺伝子が、ヘテロ接合体の形態で存在するとき、他方の対立遺伝子は、機能的Blimpを発現する。一部の態様において、改変対立遺伝子は、機能的Blimpポリペプチドまたはその機能的な形態を発現し得る。他の態様において、改変対立遺伝子は、非機能的Blimpポリペプチドを発現した。一つの態様において、改変細胞は、なかでもASCを同定および単離するのためのインビトロまたはインビボ細胞モデル系において有用である。もう一つの態様において、該改変細胞は、広範なアッセイにおけるT細胞および/またはB細胞などの造血細胞の分化状態のモニタリングに有用である。
【0020】
一つの局面において、本発明は、発現されるとき、レポーター分子と共発現される、ポリペプチドを産生する改変遺伝子を含み、ならびにポリペプチドが造血細胞の最終分化に関連している、遺伝子改変細胞またはそのような細胞を含む非ヒト生物を提供する。好ましくは、この遺伝物質は、Blimp遺伝子、またはその一部、フラグメント、相同体、誘導体もしくは機能的な形態である。さらに、B細胞系列細胞におけるレポーター分子の同定は、そのような細胞が、分化に決定付けられているか、ASCに分化したことの指標となる。または、T細胞系列の細胞におけるレポーター分子の活性は、これらの細胞が活性化されていることの指標となる。従って、本明細書において記載のように、リンパ球におけるBlimpの存在は、その細胞が最終分化するか、最終分化に決定付けられていることの指標となる。例示的なT細胞としては、CD4+ T細胞およびCD8+ T細胞が挙げられ、例示的なB細胞は、ASCである。非機能的Blimpポリペプチドが産生される場合、レポーター活性細胞の検出は、それらの細胞が、それらが機能的Blimpポリペプチドを産生することができた場合にそれらを最終分化させるために十分な条件に暴露されていることを示す。
【0021】
遺伝子改変非ヒト生物は、移植のために生殖体、胚またはES細胞の形態で提供することができる。胚は、好ましくは、凍結状態で維持され、任意で使用説明書と共に販売される。ターゲティング構築物および遺伝子改変細胞は、好ましくは凍結状態でも維持され、任意で使用説明書と共に販売される。本明細書においては、全てのそのような細胞をインビトロまたはインビボ細胞モデル系と呼ぶ。
【0022】
本発明は、単細胞、組織および生体レベルで、インビボおよびインビトロで、細胞における遺伝子発現および分化の運命をモニターするシステムを提供する。従って、レポーター活性は、生細胞でモニターすることができ、遺伝子発現は、固定された組織においてモニターすることができる。好ましくは、このレポーター発現カセットは、蛍光または他の発光部分をコードする。例えば特定の系列または細胞の最終分化についてのマーカーとしてBlimp-1の発現をレポートする生物および細胞が利用できることは、広範な用途において極めて有用なツールとなる。B細胞系列の細胞に関して、この系は、ASCの研究、単離およびモニタリングに幅広く使用することができる。前に述べたように、ASCは、これらの細胞が有効な抗体応答に非常に重要であるとはいえ、これまで研究に利用することができなかった。さらに、ASCにおける異常分化は、ヒトでは多発性骨髄腫の原因となり、その結果、ASCは、この理由についての研究のための重要な細胞タイプとなっている。
【0023】
関連態様において、本発明は、造血系の細胞を表現型解析および/またはモニターする方法を提供し、この方法は、発現されるとき、Blimpあるいはその一部、フラグメント、変異体、相同体または機能的もしくは非機能的な形態およびレポーター分子を共発現する、Blimpタンパク質をコードする改変Blimp遺伝子を含む、遺伝子改変細胞またはそのような細胞を含む非ヒト動物をスクリーニングする段階を含み、レポーター活性の検出は、細胞表現型および/または最終分化への細胞の決定付け(commitment)の指標または予測となる。造血細胞としては、B細胞、T細胞、樹状細胞、マクロファージ、ナチュラルキラー細胞、顆粒球、赤血球、好酸球、巨核球、骨髄細胞、脾細胞、皮膚細胞もしくは間質細胞またはこれらの誘導体が挙げられるが、それらに限定されるわけではない。一つの特定の態様において、造血細胞は、Bおよび/またはT細胞などのリンパ球である。
【0024】
さらなる態様において、レポーター活性またはレポーター活性の変化を示す細胞が、レポーター活性を示さない細胞の中から単離または選択される。レポーター活性細胞の単離は、任意の便利な方法によるものであり得る。例えば、フローサイトメトリー、レーザー走査サイトメトリー、クロマトグラフィーおよび/または他の同等の手法が便利に利用される。フローサイトメトリー法が特に好ましい。加えて、例えば薬物選択マーカーなどのさらなる選択マーカーを使用して、本発明の改変細胞を単離または選択することができる。
【0025】
本発明は、Blimp-1発現またはBlimp-1活性の拮抗薬(antagonist)および作動薬(agonist)も提供する。Blimp-1発現の作動薬の一例は、IL-21などであるが、これに限定されるわけではない、サイトカインである。本作動薬および拮抗薬の組換え、合成または単離形ならびに1つまたは複数の薬学的に許容される担体、希釈剤または賦形剤を含む薬学的組成物が、さらに考えられる。Blimp-1発現またはBlimp-1タンパク質の産生への言及は、単細胞におけるものまたは細胞集団内でのものを含む。
【0026】
(表1)配列識別子の一覧

【図面の簡単な説明】
【0027】
【図1】Blimp-1遺伝子座およびターゲティング戦略の図である。A)Blimp-1タンパク質の構造ドメイン。エキソン7〜8によりコードされているタンパク質のセグメントを示す。Acidic、NおよびC末端酸性領域;PR、網膜芽腫相互作用性ジンクフィンガータンパク質RIZに対するホモロジー領域;Pro、プロリン・リッチ領域;Zn、5つのジンクフィンガー。B)Blimp-1のゲノム遺伝子座、8つのエキソンをボックスとして示し、イントロンを黒線として示す。コード領域は、灰色であり、非翻訳領域は、白色である。5’および3’プローブとともに、サザン・ハイブリダイゼーションに使用した制限酵素に印を付けてある。相同組換え事象およびその後の操作から誘導された標的化対立遺伝子を示す。C)ターゲティング事象の期待産物を明らかにするためにBlimp-1遺伝子座の5’および3’末端を使用する標的化および対照ES細胞DNAに対するサザン・ハイブリダイゼーション(4.8kb 5’アームおよび4.5kb 3’アーム)。IL-15+/-IL-21においてエクスビボで0〜3日間培養したblimpgfp/+LPS刺激B細胞におけるBlimp-1の発現。Blimp-1発現は、マウスBlimp-1に対するモノクローナル抗体を使用して検出し、α-アクチンに対するヤギポリクローナル抗体を負荷対照として使用した。+/+、野生型C57B1/6マウス;+/T blimpgfp/+マウス。
【図2】インビボでのB細胞におけるBlimpgfp発現のFACS分析の結果を示すグラフ図である。A)Blimpgfp/+マウス(上のパネル)および対照(下のパネル)におけるリンパ節、脾臓および骨髄でのシンデカン-1およびBlimpgfpの発現。B)B220陽性B細胞におけるBlimpgfpの発現。
【図3】Blimpgfp選別細胞のELIspot分析の結果を示すグラブ図および写真図である。Gfp陽性細胞を未治療Blimpgfp/+マウスの骨髄(BM)および脾臓から選別し、Elispotアッセイにおいて分析した。アイソタイプ特異的抗体または抗κ抗体を使用して、Elispotプレートを被覆し、分泌された免疫グロブリンを検出した。A)200個のgfp陽性選別細胞におけるアイソタイプ特異的免疫グロブリンの分布(3回の実験のうちの1回の代表実験)。B)ELIspotプレートの単一の代表ウエルにおけるκ鎖の検出(サンプル:選別された骨髄細胞)。左、200個のgfp陽性細胞をインプット;中央、100000個のgfp陰性細胞をインプット;右、100000個の未選別細胞をインプット。
【図4A】インビボでのLPSでの抗体分泌性細胞の誘導を示すFACS分析の結果のグラフ図である。Blimpgfp/+マウスに2ugの大腸菌(E.coli)LPSを静脈内注射した。これらのマウスの脾臓を、LPS治療後の示されている時点で分析した。LPSは、ASCの形成を誘導し、その結果、脾臓において3日目には約0.5%から約5%に頻度が増加する。上のパネル、シンデカン-1およびBlimpgfpについてのFACSスキャン。中央のパネル、gfp陽性ゲート細胞内のシンデカン-1およびB220。下のパネル、示されている時点でのGFP陽性細胞におけるシンデカン-1およびB220発現についてのヒストグラム。
【図4B】インビボでのLPSでの抗体分泌性細胞の誘導を示すFACS分析の結果のグラフ図である。Blimpgfp/+マウスに2ugの大腸菌(E.coli)LPSを静脈内注射した。これらのマウスの骨髄を、LPS治療後の示されている時点で分析した。LPSは、ASCの形成を誘導し、その結果、骨髄において4日目には約0.05%から約0.25%に頻度が増加する。上のパネル、シンデカン-1およびBlimpgfpについてのFACSスキャン。中央のパネル、gfp陽性ゲート細胞内のシンデカン-1およびB220。下のパネル、示されている時点でのGFP陽性細胞におけるシンデカン-1およびB220発現についてのヒストグラム。
【図5】図5Aは、Blimpgfp発現の動態のグラフ図である。Blimpgfp/+マウス(赤線)および野生型C57B1/6マウス(青線)からの刺激後B細胞によるBlimpgfp発現のフローサイトメトリーヒストグラムを示す。ヒストグラムゲートは、Blimpgfp陽性集団のパーセンテージを示す。高度に精製された小さな休止B細胞を、組換えCD40L、IL-4およびIL-5(最上パネル)またはLPS(20ug/mL)(最下パネル)で刺激した。異なる培養日数で細胞を回収し、フローサイトメトリーで分析した。LPSで刺激した細胞は2日目にBlimpの発現を開始し、一方、CD40LおよびIL-4/IL-5に応じてBlimp発現が3日目に顕性になった。図5Bは、Blimpgfp陽性細胞が抗体を分泌することを示すグラフ図である。Blimpgfp/+B細胞をLPSで4日間刺激した。細胞を回収し、シンデカン-1(Synd-1)特異的抗体で染色し、自動細胞堆積ユニットを使用して、GFP発現領域(左のパネル、A〜C)およびGFP非発現領域(左のパネル、D)を、様々なアイソタイプ特異的抗体で被覆されたElispotプレートに直接分別した。分別された細胞を標準的なElispot法に従って処理した。右のパネルは、選別された領域におけるIg分泌性細胞の数を示す。大部分のBlimpgfp細胞は、Igを分泌するが、全てのBlimpgfp陰性細胞が、検査したいずれのIgアイソタイプも分泌しない。図5Cは、様々な刺激に応答しての種々のBlimpgfp発現を示すグラフ図である。高度に精製された小さな休止B細胞を、i)組換えCD40LおよびIL-4;ii)CD40L、IL-4およびIL-5;iii)LPS;iv)LPSおよびIL-4;v)LPSおよび抗IgDモノクローナル抗体で刺激した。4日培養した後、細胞を回収し、Synd-1特異的抗体で染色し、フローサイトメトリーで分析した。フローサイトメトリー分析の2パラメータープロットをここに示す。
【図6】活性化B細胞を移植したマウスの分析結果を示すグラフ図である。Blimpgfp/+マウスの精製された休止脾臓B細胞を20ug/mL LPSの存在下で3日間活性化した。3×106の細胞(約2×106のgfp陽性細胞を含有、すなわち、抗体分泌性細胞、A)をLPSで3回洗浄し、WT受容個体に静脈内注射により移植した。7日後、それらの受容マウスをドナーASCの存在について分析した(B)。
【図7】図7Aは、Blimpgfp/+×blimpgfp/+交雑からのマウス骨の遺伝子型解析結果を表にまとめたものである。図7Bは、離乳したマウス(左)またはE9.5日目の胚(右)の遺伝子型解析の代表的PCR結果の写真図である。
【図8】20ug/mLのLPSの存在下で培養し、3日目にGFP陽性、すなわち抗体分泌性細胞、の存在について分析した、Blimpgfp/gftおよびBlimpgfp/+再構成マウスの脾細胞の写真図およびグラフ図である(A)。その後、両方の培養物のGFP陽性細胞を選別し(B、ゲートR1)、ELIspotアッセイにおいて分析した。Blimpgfp/+細胞は、60〜70%の抗体分泌性細胞を生じ(B、下のパネル左)、Blimpgfp/gftは、5〜7%しか抗体分泌性細胞をもたらさず、ヘテロ接合細胞によって生じたスポットと比較して小さなELIspotしか生じなかった(B、下のパネル、右)。ELISPOTプレートの単一代表ウエルにおけるIgMおよびκ鎖の検出(200個のgfp陽性細胞をインプット)。
【図9】骨髄由来マクロファージ(BMM)および血液単球のFACS分析の結果のグラフ図である。骨髄細胞を10ng/mL rMCSFの存在下で7日間培養し、培地を交換し、非付着細胞を培養3日目および5日目に除去した。付着細胞(BMM)をBlimpgfp発現について分析した(左パネル)。さらに、MacI/Gr1二重陽性血球細胞をFACSで分析した(右パネル)(黒線-野生型、赤線-Blimpgfp/+)。
【図10】インビトロで産生させた樹状細胞(DC)のFACS分析を示すグラフ図である。骨髄細胞を100ng/mL Flt3リガンドの存在下で8日間培養した。その後、細胞を、さらに24時間培養(左カラム)、またはCpG(1.5uM)、GMCSF(50ng/mL)、gIFN(20ng/mL)およびIL-4(20ng/mL)で刺激(中央カラム)もしくは1ug/mLのLPSで刺激(右カラム)した。形質細胞性DCおよび従来型DCについてのBlimpgfp発現をヒストグラムで示す(実線-野生型、点線-Blimpgfp/+)。
【図11】インビボおよびインビトロでのT細胞のFACS分析を示すグラフ図である。Blimpgfp/+マウスの胸腺T細胞(左)およびリンパ節T細胞(中央)およびインビトロ活性化CD4+/CD8+精製リンパ節細胞(右)をFACSにおいて分析した。ゲートされたT細胞集団のBlimpgfp発現レベルをヒストグラムで示す(下のパネル;黒線-野生型;赤線-Blimpgfp/+)。
【図12】Blimpgfp/+レポーターマウスにおいて検出することができ、成熟刺激により誘導することができるNK系列におけるBlimp-1発現を示すグラフ図である。A)インビボ脾臓NK細胞は、GFP+である。B)Blimpgfp/+脾臓から選別されたNK細胞を4日間、IL-15中で、その後、2日間、示されているサイトカイン中で培養した。mfiは、Blimpgfpの蛍光係数の平均である。C)IL-15+/-IL-21においてエクスビボで7日間培養した、+/+NK細胞におけるBlimp-1の発現。マウスBlimp-1に対するモノクローナル抗体を使用してBlimp-1発現を検出し、α-アクチンに対するヤギポリクローナル抗体を負荷対照として使用した。
【図13−1】マウスBlimp-1/PRDM-1のcDNAおよび予測アミノ酸配列を示す図である。コード配列を大文字で示す。
【図13−2】図13−1の続きを示す図である。
【図13−3】図13−2の続きを示す図である。
【図13−4】図13−3の続きを示す図である。
【図14】図13におけるヌクレオチド配列から誘導されたマウスBlimp-1/PRDM-1のアミノ酸配列(大文字)を示す図である。
【図15−1】ヒトBlimp-1/PRDM-1のcDNAおよび予測アミノ酸配列を示す図である。コード配列を大文字で示す。
【図15−2】図15−1の続きを示す図である。
【図16】図15におけるヌクレオチド配列から誘導されたヒトBlimp-1/PRDM-1のアミノ酸配列(大文字)を示す図である。
【図17−1】マウスBlimp-1のゲノムヌクレオチド配列を示す図である。このゲノム遺伝子座は、太字大文字の8エキソンを含む。ATGおよび停止コドンに下線を引いてある。
【図17−2】図17−1の続きを示す図である。
【図17−3】図17−2の続きを示す図である。
【図17−4】図17−3の続きを示す図である。
【図17−5】図17−4の続きを示す図である。
【図17−6】図17−5の続きを示す図である。
【図18−1】ヒトBlimp-1のゲノムヌクレオチド配列を示す図である。このゲノム遺伝子座は、大文字、太字の8エキソンを含む。ATGおよび停止コドンに下線を引いてある。
【図18−2】図18−1の続きを示す図である。
【図18−3】図18−2の続きを示す図である。
【図18−4】図18−3の続きを示す図である。
【図18−5】図18−4の続きを示す図である。
【図18−6】図18−5の続きを示す図である。
【図18−7】図18−6の続きを示す図である。
【図19A】Blimp-1が、活性化/記憶T細胞において発現されることを示すグラフ図である。Blimp-1欠失T細胞は、活性化/記憶表現型を有する。A)示されている表現型の、ゲートされたCD4+またはCD8+脾臓T細胞を、GFP蛍光および活性化状態について検査した。Blimpgfp/+マウス由来のCD62LlowCD4+ T細胞およびCD44highCD8+ T細胞の大部分は、GFPが低く、少数のCD4+ T細胞(低または高CD62L)のみが、高GFPである。Blimp GFPは、Blimpgfp.gfp T細胞の同じ集団内で強く発現される(ドットブロット);Blimp-1欠失CD4+およびCD8+脾臓T細胞の表現型(ヒストグラム、実線blimp-1+/+、点線blimp-1gfp/gfp)。
【図19B】Blimp-1が、活性化/記憶T細胞において発現されることを示すグラフ図である。Blimp-1欠失T細胞は、活性化/記憶表現型を有する。B)インビトロ培養によりBlimp-1発現が誘導される。ナイーブ(naive)CD4+ T細胞をTh1/Th2分極条件下で2ラウンド成長させた。
【図19C】Blimp-1が、活性化/記憶T細胞において発現されることを示すグラフ図である。Blimp-1欠失T細胞は、活性化/記憶表現型を有する。C)Th1またはTh2条件下で上と同じように成長させたCD4+細胞における野生型Blimp-1タンパク質についてのウエスタンブロット。抗Zap-70を負荷対照として使用した。形質細胞分化を誘導するようにLPSで4日間刺激したB細胞を陽性対照として使用した。
【図19D】Blimp-1が、活性化/記憶T細胞において発現されることを示すグラフ図である。Blimp-1欠失T細胞は、活性化/記憶表現型を有する。D)示されている遺伝子型で再構成されたマウスを、HSV感染後、gB特異的四量体を使用して優性gB498-505エピトープに特異的なCD8+ T細胞の出現について分析した。HSV感染データは、各々の遺伝子型の少なくとも3匹のマウスの代表である。
【図19E】Blimp-1が、活性化/記憶T細胞において発現されることを示すグラフ図である。Blimp-1欠失T細胞は、活性化/記憶表現型を有する。E)インビトロ培養gB特異的CTLは、正常な細胞毒性機能を示す。
【図20A】Blimp-1陽性CD4+ T細胞集団の分子分析の図である。A)Blimp-1は、CD25+サプレッサーT細胞において発現される。CD25+およびCD25-CD4+ T細胞を選別し(左のパネル)、ナイーブCD4+をTh1およびTh2分極条件下で分化させ(右のパネル)、全ての細胞集団をRT-PCR分析に供した。
【図20B】Blimp-1陽性CD4+ T細胞集団の分子分析の図である。B)Blimp-1欠失CD4+細胞は、大量のIFNgを分泌し、IL-10分泌欠損を示した。CD4+細胞は、脾臓からエクスビボで選別し、抗CD3およびCD28結合プレートで24時間再刺激したか、またはTh1もしくはTh2細胞にインビトロで分化させ、再刺激に供した。上清中のIL-10およびIFNgは、ELISAで検出した。
【図21】Blimp-1欠失マウスが、致死性リンパ球増殖亢進症候群を発現することを示す図である。A)対照細胞またはBlimpgfp/gfp胎児肝臓由来幹細胞で再構成したRag1-/-マウスの組織学的検査。Blimpgfp/gfp再構築マウスは、瀕死の場合、犠牲にした。Blimpgfp/+マウスからの正常な組織学的外観の器官を、Blimpgfp/gfpマウスで観察されたリンパ球浸潤と対比させる。B)Blimpgfp/+再構成マウスではなく、Blimpgfp/gfp再構成マウスにおける病的状態の急速な発症。検査した各遺伝子型の動物の数を示す。
【図22A】Blimp-1が、T細胞の恒常性増殖を調節することを示すデータのグラフ図である。A)示されている遺伝子型由来の3×106のナイーブCD4+またはCD8+脾臓T細胞を非放射線照射Rag2-/-受容個体に養子的に移入した。受容個体を体重減少および窮迫の徴候についてモニターした。グラフは、3〜4週間にわたる体重の変化率を示す。4〜6匹のマウスを各遺伝子型の細胞で再構成した。
【図22B】Blimp-1が、T細胞の恒常性増殖を調節することを示すデータのグラフ図である。B)体重が10%より多く減少したマウスを犠牲にし、脾臓T細胞数を判定した。
【図22C】Blimp-1が、T細胞の恒常性増殖を調節することを示すデータのグラフ図である。C)Blimpgfp/gfp CD8+ T細胞の移入後の代表Rag2-/-マウスの脾腫。
【図22D】Blimp-1が、T細胞の恒常性増殖を調節することを示すデータのグラフ図である。D)移入の3週間後のドナーT細胞におけるBlimpgfp発現のフローサイトメトリー分析。
【図23】Blimp-1が、二次刺激後に誘導され、サイトカイン応答性を調節することを示すデータの図である。A)示されている遺伝子型のナイーブCD8+ T細胞を抗CD3/CD28およびIL-2の存在下で5日間培養し、その後、5日間、IL-21を含有する二次培養物に再接種した。B)上と同じ細胞培養物を、示されているサイトカインの組み合わせと共にまたはそれらを伴わずに抗CD3/CD28で刺激した。一次または二次培養物において、示されている時点で、全生細胞数を判定した。CD44high活性化/記憶CD8+細胞は、一次培養のみに供した。
【発明を実施するための形態】
【0028】
好ましい態様の詳細な説明
本発明は、造血細胞系の細胞もしくは胚性細胞を同定および単離するための、および/または造血細胞もしくは胚性細胞の分化をモニターするための方法の開発に、一部基づいており、この方法は、(レポーターによる)ポリペプチドの存在の検出または定量を含み、その存在は、細胞の最終分化に関連している。
【0029】
特に好ましい態様において、ポリペプチドは、Blimpまたはその一部、フラグメントもしくは機能的な形態であり、これらは、レポーター分子と共発現される。
【0030】
従って、本発明の一つの局面は、発現されるとき、レポーター分子と共発現されるポリペプチドを産生する、ポリペプチドをコードする遺伝物質を含み、ポリペプチドが、細胞における最終分化への決定付けを含む細胞表現型に関連している、遺伝子改変細胞またはそのような細胞を含む非ヒト生物を提供する。
【0031】
さらなる局面において、本発明は、発現されるとき、レポーター分子と共発現されるBlimpまたはその一部、フラグメントもしくは機能的な形態を産生する、Blimpポリペプチドをコードする改変Blimp遺伝子を含み、Blimpの存在が、細胞における最終分化への決定付けを含む細胞表現型に関連している、遺伝子改変細胞またはそのような細胞を含む非ヒト生物を提供する。
【0032】
さらに好ましい局面において、本発明は、Blimpコード配列またはその一部、フラグメントもしくは機能的な形態およびレポーター分子コード配列を含む、Blimp mRNA転写産物をコードする改変Blimp遺伝子を含み、Blimpの存在が、細胞における最終分化への決定付けを含む細胞表現型に関連している、遺伝子改変細胞またはそのような細胞を含む非ヒト生物を提供する。
【0033】
好ましくは、レポーター分子コード配列は、Blimp対立遺伝子のイントロンに挿入される。このようにして、改変Blimp対立遺伝子は、内因性Blimp調節要素の制御下で2シストロンのRNAからレポーターを共産生する。
【0034】
用語「共発現」および「共産生」は、同時または実質的に同時の2つ以上の核酸領域の転写(1つまたは複数のRNAとして発現される)および同時または実質的に同時な、それらのその後の翻訳(1つまたは複数のポリペプチドとして産生される)を指すように本明細書においては広義に用いている。好ましくは、Blimpまたはその一部、フラグメントもしくは機能的な形態とレポーター分子の両方をコードする1つの転写産物が発現される。各場合、レポーターの発現は、レポートされる分子の発現に作動可能に連結している。
【0035】
本明細書において「細胞表現型」への言及は、細胞の分子特性または機能特性を包含する。例えば、ASC細胞は、Blimp-1(分子マーカー)を発現し、ならびになかでも、高いIg分泌率、MHCクラスII分子の不在および低い表面Igレベルを示すことにより、他のB細胞とは機能的に区別される。本明細書において用いる場合、この用語は、調節されたレベルのBlimp-1発現の分子特性に加えて、全範囲の分子もしくは機能特性、または任意の特定の分子もしくは機能特性への言及である。
【0036】
本遺伝子改変細胞またはそのような細胞を含む非ヒト生物は、ヒト、非ヒト霊長類、家畜、愛玩生物、実験検査生物、爬虫類または両生類などであるが、これらに限定されない、任意の生物由来の細胞または遺伝物質を含むことができる。好ましくは、本遺伝子改変生物は、マウスまたは他の実験検査動物、例えばラット、モルモット、ブタ、ウサギもしくはヒツジである。
【0037】
本明細書において用いる場合、単数形「ある(a、an)」および「その(the)」は、その文脈が明確に別様に指示していない限り、複数の局面を含む。従って、例えば、「細胞」への言及は、単細胞のみでなく2個以上の細胞を含み;「遺伝子」への言及は、1つの遺伝子のみでなく2つ以上の遺伝子を含むなどである。
【0038】
本発明の改変遺伝子は、B細胞系列細胞などの造血系の細胞における最終分化についてのマーカーである。
【0039】
「遺伝子改変細胞」への言及は、その改変前の形態を基準にして変更されているゲノムおよびその後代のコード領域または非コード領域からのヌクレオチドの配列を含むように操作された任意の細胞への言及である。特に、細胞は、最終分化の遺伝子マーカーおよびレポーター分子コード配列を共発現するように遺伝子改変される。好ましくは、細胞は、Blimpまたはその一部、フラグメントもしくは機能的な部分およびレポーター分子を共発現するように遺伝子改変される。レポーター分子は、同定可能なシグナルを直接または間接的に生じることができる任意の分子であり得る。蛍光または他の発光レポーター分子が、特に好ましい。
【0040】
便利には、最初にターゲティング構築物を用いて、細胞または生物体内で改変遺伝子配列を発生させる。一般に、しかし排他的でなく、ターゲティング構築物は、相同組換えにより標的配列を改変する。または、改変された遺伝子配列を人工染色体を使用して導入することができる。ターゲティングまたは他の構築物は、例えば、Sambrook, Molecular Cloning: A Laboratoy Manual, 3rd Edition, CSHLP, CSH, NY, 2001 ; Ausubel (Ed) Current Protocols in Molecular Biology, 5th Edition, John Wiley & Sons, Inc, NY, 2002などの、分子生物学実験室マニュアルに記載されている当技術分野において周知の方法を用いて、産生させ、標的細胞に導入することができる。ターゲティング構築物は、エレクトロポレーション、ウイルス媒介移入またはマイクロインジェクションなどの任意の方法により細胞に導入することができる。一般には選択マーカーを利用して、ターゲティング構築物が首尾よく組みこまれた細胞を最初に同定する。当業者には理解されるように、本改変生物は、一定の細胞においてのみBlimp対立遺伝子およびレポーター分子を発現するように遺伝子改変することができる。
【0041】
一つの特定の態様において、本発明は、ターゲティング構築物としての使用に適する核酸構築物を提供し、構築物は、Blimp-1の対立遺伝子の全てまたは一部分およびレポーター構築物を含む。構築物は、機能的に活性なBlimp-1ポリペプチドまたは機能的に不活性なBlimp-1ポリペプチドをコードする遺伝物質を含む。特定の態様において、構築物は、DNA結合モチーフを含むジンクフィンガードメインを欠く部分的Blimp-1ポリペプチドをコードする。特に好ましい態様において、構築物は、核酸構築物のリコンビナーゼ媒介欠失および標的遺伝子配列への核酸構築物の相同組換えを助長する部位に隣接している。または、構築物は、それがエピソーム的に複製する宿主細胞に導入することができる。
【0042】
遺伝子改変生物は、Hogan et al., Manipulating the Mouse Embryo : A Laboratory Manual, Cold Spring Harbour Laboratory Press, CSH NY, 1986 ; Mansour et al., Nature 336 :348-352,1988 ; Pickert, Transgenic Animal Technology: A Laboratory Handbook, Academic Press, San Diago, CA, 1994に記載されているような当技術分野において周知の技法を用いて産生させる。胚性幹細胞(ES細胞)を含む幹細胞を発生の胚盤胞期に受容生物の胚に導入する。そこではそれらを内細胞塊に組み込むことができ、この内細胞塊で、それらは、受容生物の生殖細胞系に発達し寄与する。ES細胞は、インビトロで維持されている移植前の胚から便利に得られる(Robertson et al., Nature 322 : 445-448, 1986)。正確なターゲティングが確認されたら、改変細胞を胚盤胞期または桑実期または他の適する発達段階に注入して、キメラ生物を発生させる。または改変細胞を解離胚細胞と凝集させて、凝集キメラを形成することができる。その後、そのキメラ生物を、適する雌里親生物に移植し、胚を出産日まで発達させることができる。キメラ後代を繁殖させて、ターゲティング構築物によりもたらされたヌクレオチド配列を各細胞のゲノムが含有する子孫を得る。遺伝子改変生物は、ヘテロ接合改変を含むことがあり、または両方の対立遺伝子が影響を受けることがある。
【0043】
本発明によると、驚くべきことに、Blimp-1は、ASC経路に沿って分化する決定付けではなく、ASCによる抗体の産生に不可欠であると判定される。従って、B細胞系列細胞におけるBlimpの(例えば、レポーター分子との共発現による)同定は、細胞が、ASCに分化するように決定付けられているか、ASCに分化したことを示す。
【0044】
さらに、本明細書において開示するように、Blimpは、T細胞恒常性を含むリンパ球恒常性および最終分化するようになるT細胞の能力に不可欠である。生体哺乳動物におけるBlimpの不在は、進行性多臓器リンパ球増殖性疾患を導く。
【0045】
従って、本発明のもう一つの局面は、発現されるとき、レポーター分子と共発現されるポリペプチドを産生するポリペプチドをコードする遺伝物質を含む、遺伝子改変細胞またはそのような細胞を含む非ヒト生物を提供し、レポーター分子の検出は、細胞表現型および/または最終分化への細胞の決定付けの指標となる。
【0046】
さらなる態様において、本発明は、発現されるとき、レポーター分子と共発現されるBlimpまたはその一部、フラグメントもしくは機能的な形態を産生する、Blimpポリペプチドをコードする改変Blimp遺伝子を含む、遺伝子改変細胞またはそのような細胞を含む非ヒト生物を提供し、レポーター分子の検出は、細胞表現型および/または最終分化への細胞の決定付けの指標となる。
【0047】
さらに好ましい局面において、本発明は、Blimpコード配列またはその一部、フラグメントもしくは機能的な形態およびレポーター分子コード配列を含む、Blimp mRNA転写産物をコードする改変Blimp遺伝子を含む、遺伝子改変細胞またはそのような細胞を含む非ヒト生物を提供し、この場合、レポーター分子の検出は、細胞表現型および/または最終分化への細胞の決定付けの指標となる。
【0048】
好ましくは、レポーター分子コード配列は、Blimp対立遺伝子のイントロンに挿入される。このようにして、改変Blimp対立遺伝子は、内因性Blimp調節要素の制御下で2シストロンのRNAからレポーターを共産生する。
【0049】
従って、本発明のもう一つの局面は、発現されるとき、レポーター分子と共発現されるポリペプチドを産生する、ポリペプチドをコードする遺伝物質を含む、遺伝子改変細胞またはそのような細胞を含む非ヒト生物を提供し、この場合、造血系の細胞におけるレポーター分子の検出は、細胞表現型および/または最終分化への細胞の決定付けの指標となる。
【0050】
さらなる局面において、本発明は、発現されるとき、レポーター分子と共発現されるBlimpまたはその一部、フラグメントもしくは機能的な形態を産生する、Blimpポリペプチドをコードする改変Blimp遺伝子を含む、遺伝子改変細胞またはそのような細胞を含む非ヒト生物を提供し、B細胞におけるレポーター分子の検出は、レポーター分子活性を有する細胞がASCへの分化に決定付けられていることの指標となる。
【0051】
さらに好ましい局面において、本発明は、Blimpコード配列またはその一部、フラグメントもしくは機能的な形態およびレポーター分子コード配列を含む、Blimp mRNA転写産物をコードする改変Blimp遺伝子を含む、遺伝子改変細胞またはそのような細胞を含む非ヒト生物を提供し、この場合、T細胞におけるレポーター分子の検出は、レポーター分子活性を有する細胞が活性化T細胞であることの指標となる。
【0052】
好ましくは、レポーター分子コード配列は、Blimp対立遺伝子のイントロンに挿入される。このようにして、改変Blimp対立遺伝子は、内因性Blimp調節要素の制御下で2シストロンのRNAからレポーターを共産生する。
【0053】
本明細書においてBlimp-1遺伝子またはその核酸発現産物(RNA)への言及は、その相同体、部分、フラグメント、機能的な形態(低ストリンジェンシー条件下でそれにハイブリダイズするか、最適なアラインメント後に全部分または機能的な部分に対する有意な配列類似性、例えば少なくとも約60%の配列類似性を含む機能的変異体または誘導体を含む)を含む。Blimp-1ポリペプチドまたはタンパク質への言及は、その全ての相同体、部分、フラグメントまたは機能的な形態(最適なアラインメント後に少なくとも60%のアミノ酸配列類似性を有する機能的変異体またな誘導体を含む)を含むように広義で用いられている。
【0054】
本分子の機能的な部分としては、その特定の機能、例えば基質結合、三次コンフォメーションまたは転写活性に重要である完全長分子の部分が挙げられる。転写開始部位は、容易にマッピングされ、プロモーター活性を付与する部位は容易に同定される(例えば、Tunyaplin et al., Nucleic Acid Research 28 (24):4846-4855, 2000参照)。機能的な部分は、分子の発現および活性の調節に重要である。機能的変異体または誘導体は、参照分子の発現および活性の調節に重要な機能活性の少なくとも1つを保持する。Blimp-1に関しては、その発現が、最終分化、ASCによるIg分泌の誘導、およびT細胞の活性化に関連している。
【0055】
改変Blimp遺伝子は、機能的に活性なBlimpポリペプチド、機能的に不活性なBlimpポリペプチド、および/または部分的Blimpポリペプチド、例えばDNA結合モチーフを含むジンクフィンガードメインを欠く、ポリペプチドもしくはペプチドなどをコードすることができる。用語「ポリペプチド」および「タンパク質」は、本発明では交換可能に用いている。
【0056】
ペプチド形の「部分」は、5個未満のアミノ酸を含むエピトープほどの小ささであってもよいし、または数百キロダルトンほどの大きさであってもよい。相同体と比較されるポリペプチド配列の長さは、一般に、少なくとも約16のアミノ酸、通常は少なくとも約20残基、さらに通常は少なくとも約24残基、典型的には少なくとも約28残基および好ましくは約35より多い残基であろう。核酸分子の「一部」は、少なくとも約10ヌクレオチド、または好ましくは約13ヌクレオチド、またはさらに好ましくは少なくとも約20ヌクレオチドの最小サイズを有すると定義し、少なくとも約35ヌクレオチドの最小サイズを有することもある。この定義は、10〜35の範囲内の全てのサイズのヌクレオチド、ならびに50、100、300、500、600ヌクレオチドを含む35より多くのヌクレオチド、またはこれらの値の範囲内の任意の数のヌクレオチドを有する核酸分子を含む。
【0057】
本発明は、変異体Blimpポリペプチドをコードする改変Blimp対立遺伝子も企図する。「変異体」ポリペプチドとしては、天然タンパク質のN末端および/もしくはC末端での1つまたは複数のアミノ酸の欠失(いわゆるトランケーション)もしくは付加;天然タンパク質における1つまたは複数の部位での1つまたは複数のアミノ酸の欠失または付加;または天然タンパク質における1つまたは複数の部位での1つまたは複数のアミノ酸の置換により、天然タンパク質から誘導されるタンパク質が挙げられる。本発明に包含される変異体タンパク質は、生物活性であり、すなわち、天然タンパク質の望ましい生物活性を保持し続けている(すなわち、それらは、例えばc-mycおよび/またはCIITAの転写リプレッサーである)。または、変異体Blimpポリペプチドは、非機能的である。このような変異体は、例えば、遺伝的多形の結果または人間が操作した結果として生じ得る。天然Blimpポリペプチドの生物活性変異体は、デフォルトパラメータを使用する本明細書の他の箇所で説明する配列アラインメントプログラムによって判定して、天然タンパク質についてのアミノ酸配列と少なくとも40%、50%、60%、70%、一般には少なくとも75%、80%、85%、好ましくは約90%から95%またはそれより多く、さらに好ましくは約98%以上の配列類似性を有するであろう。Blimpポリペプチドの生物活性変異体は、ポリペプチドとは、一般に100、50もしくは20個という多さのアミノ酸残基、または好適には1〜15個という少なさのアミノ酸残基、1〜10個という少なさ、例えば6〜10個、5個という少なさ、4、3、2個という少なさ、またはさらには1個のアミノ酸残基が異なり得る。
【0058】
Blimpポリペプチドは、アミノ酸置換、欠失、トランケーションおよび挿入を含む様々な方法で変更することができる。このような操作のための方法は、当技術分野において一般に公知である。例えば、Blimpポリペプチドのアミノ酸配列変異体は、コードされている核酸配列の突然変異により作製することができる。突然変異誘発およびヌクレオチド配列変更のための方法は、当技術分野において周知である。例えば、Kunkel (Proc. Natl. Acad. Sci. USA 82 : 488-492,1985), Kunkel et al.,(Methods in Enzymol. 154 : 367-382,1987),米国特許第4,873,192号, Watson et al. ("Molecular Biology of the Gene", Fourth Edition, Benjamin/Cummings, Menlo Park, Calif., 1987)およびこれらに引用されている参考文献を参照されたい。対象となるタンパク質の生物活性に影響を及ぼす、または及ぼさない適切なアミノ酸置換基についてのガイダンスは、Dayhoffらのモデル(Natl. Biomed. Res. Found 5 : 345-358, 1978)において見出すことができる。例えば、DNA結合モチーフを含有するジンクフィンガードメインの全てまたは一部の欠失は、機能的に不活性であるBlimp変異体を生じさせるであろう。一部の態様において、動物モデルは、一部または全ての組織において遺伝子改変非機能的Blimp対立遺伝子がヘテロ接合性であるが、他の対立遺伝子は、機能的Blimpポリペプチドを発現することができる機能的Blimp対立遺伝子を含む。他の態様において、特定の細胞または組織ではBlimpを発現しないヘテロ接合性動物を産生させる。または1つまたは2つの改変Blimp対立遺伝子により細胞、組織または非ヒト生物において機能的Blimpを産生させることができる。点突然変異またはトランケーションにより作製されたコンビナトリアルライブラリの遺伝子産物をスクリーングするための方法、および選択された特性を有する遺伝子産物についてcDNAライブラリをスクリーニングするための方法は、当技術分野において公知である。このような方法は、Blimpポリペプチドのコンビナトリアル突然変異誘発により作製された遺伝子ライブラリの迅速なスクリーニングに適応できる。ライブラリの中の機能的突然変異体の頻度を増す技法である、再帰的集合型突然変異誘発(Recursive ensemble mutagenesis)(REM)をスクリーニングアッセイと併用して、Blimpポリペプチド変異体を同定することができる(Arkin et al., Proc. Natl. Acad. Sci, USA 89 : 7811-7815, 1992; Delgrave et al. Protein Engineering 6:327-331, 1993)。より詳細に下で論じるような、保存的置換、例えば、1つのアミノ酸と類似した特性を有する別のアミノ酸との交換が望ましいこともある。
【0059】
親Blimpアミノ酸配列と比較すると、変異体Blimpポリペプチドは、配列に沿った様々な位置での保存的アミノ酸配列置換を含有し得る。「保存的アミノ酸置換」は、アミノ酸残基が類似した側鎖を有するアミノ酸残基で置換されることである。類似した側鎖を有するアミノ酸残基のファミリーは、当技術分野において定義されており、それらは、一般に、次のように下位分類することができる:
【0060】
酸性:この残基は、生理pHでHイオンを喪失するため負の電荷を有し、およびこの残基は、水溶液に誘引されて、ペプチドの配座の表面位置を求め、ペプチドが生理pHの水性媒体中にあるときにはそこに収容される。酸性側鎖を有するアミノ酸としては、グルタミン酸およびアスパラギン酸が挙げられる。
【0061】
塩基性:この残基は、生理pHでまたはその1または2pH単位(例えば、ヒスチジン)内でHイオンと会合するため正の電荷を有し、およびこの残基は、水溶液に誘引されて、ペプチドの配座の表面位置を求め、ペプチドが生理pHの水性媒体中にあるときにはそこに収容される。塩基性側鎖を有するアミノ酸としては、アルギニン、リジンおよびヒスチジンが挙げられる。
【0062】
荷電:これらの残基は、生理pHで帯電しており、従って、酸性側鎖または塩基性側鎖を有するアミノ酸(すなわち、グルタミン酸、アスパラギン酸、アルギニン、リジンおよびヒスチジン)を含む。
【0063】
疎水性:これらの残基は、生理pHで帯電しておらず、およびその残基は、水溶液に反発して、ペプチドの配座の内部位置を求め、ペプチドが水性媒体中にあるときにはそこに収容される。疎水性側鎖を有するアミノ酸としては、チロシン、バリン、イソロイシン、ロイシン、メチオニン、フェニルアラニンおよびトリプトファンが挙げられる。
【0064】
中性/極性:これらの残基は、生理pHで帯電していないが、その残基は、ペプチドの配座の内部位置を求めてペプチドが水性媒体中にあるときにそこに収容されることになるほどには水溶液と反発しない。中性/極性側鎖を有するアミノ酸としては、アスパラギン酸、グルタミン酸、システイン、ヒスチジン、セリンおよびトレオニンが挙げられる。
【0065】
本明細書においては、一定のアミノ酸を、それらの側鎖がたとえ極性基を欠く場合であっても疎水性をもたらすほどには大きくないことから、「小さい」とも特性付けしている。プロリンを除き、「小さい」アミノ酸は、少なくとも1つの極性基が1つの側鎖上にあるときには4つ以下の炭素原子を有するアミノ酸、およびそうでないときには3つ以下の炭素原子を有するアミノ酸である。小さい側鎖を有するアミノ酸としては、グリシン、セリン、アラニンおよびトレオニンが挙げられる。遺伝子にコードされている第二アミノ酸プロリンは、ペプチド鎖の二次コンフォメーションに対するその公知の作用のため、特別な場合である。プロリンの構造は、その側鎖がαアミノ基の窒素ならびにα炭素に結合している点で、他の全ての天然由来アミノ酸とは異なる。しかし、幾つかのアミノ酸類似性マトリックス(例えば、Dayhoff et al., 1978 (前記);および Gonnet et al., Science 256 (5062): 1443-1445,1992に例えば開示されているようなPAM120マトリックスおよびPAM250マトリックス)は、グリシン、セリン、アラニンおよびトレオニンと同じグループにプロリンを含む。従って、本発明の目的のために、プロリンは、「小さい」アミノ酸と分類する。
【0066】
極性または非極性と分類するために必要とされる誘引度または反発度は、自由裁量であり、従って、本発明が特に企図するアミノ酸は、様々に分類されている。特に名を挙げていない大部分のアミノ酸は、公知の性質を基に分類することができる。
【0067】
アミノ酸残基は、環状または非環状;芳香族性または非芳香族性;残基の側鎖置換基を基準にした自明の分類;および小さいまたは大きいとさらに下位に分類することができる。残基は、追加の極性置換基が存在するという条件ではカルボキシル炭素を含め合計4個以下の炭素原子を含有する場合;そうでなければ3個以下の場合、小さいと見なす。勿論、小さい残基は、必ず非芳香族である。それらの構造特性に依存して、アミノ酸残基は、2つ以上のクラスに分類することができる。天然由来タンパク質アミノ酸について、この理論体系による下位分類法を表Aに提示する。
【0068】
(表A)アミノ酸下位分類法
【0069】
保存的アミノ酸置換も、側鎖に基づくグループ分けを含む。例えば、脂肪族側鎖を有するアミノ酸のグループは、グリシン、アラニン、バリン、ロイシンおよびイソロイシンであり;脂肪族-ヒドロキシル側鎖を有するアミノ酸のグループは、セリンおよびトレオニンであり;アミド含有側鎖を有するアミノ酸のグループは、アスパラギンおよびグルタミンであり;芳香族側鎖を有するアミノ酸のグループは、フェニルアラニン、チロシンおよびトリプトファンであり;塩基性側鎖を有するアミノ酸のグループは、リジン、アルギニンおよびヒスチジンであり;ならびに硫黄含有側鎖を有するアミノ酸のグループは、システインおよびメチオニンである。例えば、イソロイシンもしくはバリンでのロイシンの置換、グルタミン酸のアスパラギン酸の置換、セリンでのトレオニンの置換、または構造的に関連したアミノ酸でのアミノ酸の同様の置換は、結果として生じる変異体ポリペプチドの特性に重要な影響を及ぼさないであろうと予測するのが妥当である。アミノ酸変更が、結果として機能的Blimpポリペプチドを生じさせるかどうかは、その活性をアッセイすることにより容易に判定することができる。保存的置換を、下記表B中、例示的な置換の見出しの下に示す。さらに好ましい置換を、好ましい置換の見出しの下に示す。本発明の範囲に入るアミノ酸置換は、一般に、(a)その置換領域内のペプチド骨格の構造、(b)標的部位における分子の電荷もしくは疎水性、または(c)側鎖の嵩、を維持することに対するそれらの影響が有意に異ならない置換を選択することにより、達成することができる。置換導入後、変異体を生物活性についてスクリーニングする。
【0070】
(表B)例示的なおよび好ましいアミノ酸置換
【0071】
または、保存的置換を行うために類似したアミノ酸をその側鎖の同一性に基づき3つのカテゴリーにグループ分けすることができる。Zubay, G., Biochemistry, third edition, Wm. C. Brown Publishers (1993)に記載されているように、第一のグループは、グルタミン酸、アスパラギン酸、アルギニン、リジン、ヒスチジンを含み、これらは全て、荷電側鎖を有し;第二のグループは、グリシン、セリン、トレオニン、システイン、チロシン、グルタミン、アスパラギンを含み、および第三のグループは、ロイシン、イソロイシン、バリン、アラニン、プロリン、フェニルアラニン、トリプトファン、メチオニンを含む。
【0072】
従って、Blimpポリペプチド中の予想される非必須なアミノ酸は、典型的に、同じ側鎖ファミリーからの別のアミノ酸残基で置換される。または飽和突然変異誘発などによりBlimpポリヌクレオチドコード配列の全てまたは一部に沿ってランダムに突然変異を導入することができ、結果として生じた突然変異体を親ポリペプチドの活性についてスクリーニングして、その活性を保持する突然変異体を同定することができる。コード配列の突然変異誘発後、コードされたペプチドを組換え発現させることができ、そのポリペプチドの活性を判定することができる。
【0073】
従って、本発明は、天然由来Blimpポリペプチド配列の変異体またはそれらの生物活性フラグメントも企図し、この場合の変異体は、1つまたは複数のアミノ酸残基の付加、欠失または置換により天然由来配列とは区別される。一般に、変異体は、例えば配列番号:2および4のいずれか一方に記載の親Blimpポリペプチド配列との少なくとも約30、40、50、55、60、65、70、75、80、85、90、91、92、93、94、95、96、97、98、99%の類似性を示すであろう。望ましくは、変異体は、例えば配列番号:2および4のいずれか一方に記載の参照Blimpポリペプチド配列との少なくとも30、40、50、55、60、65、70、75、80、85、90、91、92、93、94、95、96、97、98、99%の配列同一性を有するであろう。さらに、1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、30、40、50、60、70、80、90、100以上のアミノ酸の付加、欠失または置換により天然または親配列とは異なるが、親Blimpポリペプチドの特性を保持する配列が考えられる。Blimpポリペプチドは、本明細書において定義するようなストリンジェンシー条件、特に高ストリンジェンシー条件下で、Blimpポリヌクレオチド配列にハイブリダイズするポリヌクレオチドまたはその非コード鎖によりコードされるポリペプチドも含む。
【0074】
一部の態様において、変異体ポリペプチドは、少なくとも1個、しかし50、40、30、20、15、10、8、6、5、4、3または2個未満のアミノ酸残基がBlimp配列と異なる。別の態様において、変異体ポリペプチドは、少なくとも1%、しかし20%、15%、10%または5%未満の残基が配列番号:2および4のいずれか一方における対応する配列と異なる。(この比較にアラインメントが必要な場合、最大類似性のために配列をアラインしなければならない。欠失もしくは挿入またはミスマッチからの「ループド」アウト("Looped" out)配列の差異を考慮に入れる)。好適には、差異は、非必須残基における差異もしくは変更または保存的置換である。
【0075】
「非必須」アミノ酸残基は、態様ポリペプチドの野生型配列から変更することができるが、その活性の1つまたは複数を撤廃しない、または実質的に変更しない残基である。好適には、変更は、これらの活性の1つを実質的に変更せず、例えば、活性は、野生型の少なくとも20%、40%、60%、70%または80%である。「必須」アミノ酸残基は、本発明のBlimpポリペプチドの野生型配列から変更されたとき、結果的に親分子の活性が撤廃される、例えば、野生型活性の20%未満が存在する、残基である。
【0076】
他の態様において、変異体ポリペプチドは、例えば配列番号:2および4のいずれか一方に記載のBlimpポリペプチドの対応する配列との、少なくとも約50%、55%、60%、65%、70%、75%、80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%またはそれより高い類似性を有するアミノ酸配列を含む。
【0077】
本発明は、任意の哺乳動物または動物(鳥類を含む)被験者からの、例えばヒト、非ヒト霊長類、家畜、実験動物、愛玩動物または野生動物からのBlimpを包含する。「Blimp」への言及は、上記のいずれかの種由来のBlimpまたはBlimpならびにその構造的もしくは進化的等価物または相同体を含む。例えば、本発明は、配列番号:2もしくは4との少なくとも約60%の類似性、または配列番号:1、3、5もしくは6との少なくとも約60%の同一性を実質的に有するアミノ酸配列を有するBlimpまたはBlimpを包含する。少なくとも約60%への言及は、60、61、62、63、64%およびそれに続く100%までの級数のうちの連続的な数字を含む。
【0078】
核酸形での分子の機能誘導体は、低ストリンジェンシー条件下で分子またはその相補形にハイブリダイズできるヌクレオチド配列を含む核酸分子を含む。
【0079】
本明細書において用いる用語「類似性」または「同一性」は、ヌクレオチドまたはアミノ酸レベルでの比較される配列間の正確な同一性を含む。ヌクレオチドレベルで同一性がない場合、「類似性」は、それにもかかわらず構造的、機能的、生化学的および/または配座レベルで互いに関連している異なるアミノ酸を結果的に生じる配列間の差異を含む。アミノ酸レベルで同一性がない場合、「類似性」は、それにもかかわらず構造的、機能的、生化学的および/または配座レベルで互いに関連しているアミノ酸を含む。特に好ましい態様において、ヌクレオチドおよびアミノ酸配列比較は、類似性ではなく同一性のレベルで行われる。
【0080】
2つ以上のポリヌクレオチドまたはポリペプチド間の配列の関係を記述するために使用する用語としては、「参照配列」、「比較ウインドウ」、「配列類似性」「配列同一性」、「配列類似率」、「配列同一率」、「実質的に類似した」および「実質的同一性」が挙げられる。「参照配列」は、長さで、少なくとも12、しかししばしば15〜18、多くの場合、少なくとも25以上、例えば30個のモノマー単位(ヌクレオチドおよびアミノ酸残基を含む)である。2つのポリヌクレオチドは、各々、(1)その2つのヌクレオチドの間で類似している配列(すなわち、完全ポリヌクレオチド配列の一部だけ)、および(2)その2つのポリヌクレオチドの間で互いに異なる配列を含み得るので、典型的に、2つの(またはそれより多い)ポリヌクレオチドの間の配列比較は、局所的な配列類似性領域を同定および比較するように「比較ウインドウ」全体にわたってその2つポリヌクレオチドの配列を比較することによって行われる。「比較ウインドウ」は、参照配列と比較される典型的には12個の連続する残基の概念セグメントを指す。2つの配列の最適なアラインメントのために、比較ウインドウは、(付加または欠失を含まない)参照配列と比較して約20%以下の付加または欠失(すなわち、ギャップ)を含み得る。比較ウインドウをアラインするための配列の最適なアラインメントは、コンピュータによるアルゴリズムの実現(米国、ウィスコンシン州、Madison、575 Science Drive、Genetics Computer Group、Wisconsin Genetics Software Package Release 7.0におけるGAP、BESTFIT、FASTA、およびTFASTA)によって、または検査、および選択された任意の様々な方法により生成される最良アラインメント(すなわち、比較ウインドウ全体にわたって最高の相同率を結果として生じるもの)によって行うことができる。例えば、Altschul et al., Nucl. Acids Res. 25. 3389, 1997により開示されているようなプログラムのBLASTファミリーを参照することもできる。配列分析の詳細な論考は、Ausubel et al., Current Protocols in Molecular Biology John Wiley & Sons Inc, 1994-1998, Chapter 15)のUnit 19.3において見出すことができる。
【0081】
本明細書において用いる用語「配列類似性」および「配列同一性」は、配列が、比較ウインドウ全体にわたってヌクレオチドごとのベースまたはアミノ酸ごとのベースで同一であるか、機能的にもしくは構造的に類似していることの程度を指す。従って、例えば「配列同一率」は、比較ウインドウ全体にわたって最適にアラインされた2つの配列を比較し、両方の配列において同一の核酸塩基(例えば、A、T、C、G、I)または同一のアミノ酸残基(例えば、Ala、Pro、Ser、Thr、Gly、Val、Leu、Ile、Phe、Tyr、Trp、Lys、Arg、His、Asp、Glu、Asn、Gln、CysおよびMet)が発生する位置の数を決定してマッチした位置の数を得、そのマッチした位置の数を比較ウインドウ内の全位置数(すなわち、そのウインドウのサイズ)で割り、その結果に100を掛けて配列同一率を得ることによって計算する。本発明の目的のために、「配列同一性」は、DNASISコンピュータプログラム(米国、カリフォルニア州、South San FranciscoのHitachi Software engineering Co.,Ltd.から入手できる、Windows用Version 2.5)により、そのソフトウェアに添付されている参照マニュアルにおいて使用されているような標準デフォルトを使用して計算される「マッチ率」を意味すると理解されよう。同様のコメントが配列類似性に関連して適用される。
【0082】
さらに、Blimp相同体または誘導体は、低ストリンジェンシー条件下で、配列番号:1、3、5もしくは6、またはそれらからの相補形にハイブリダイズすることができると定義することができる。
【0083】
本明細書において低ストリンジェンシーへの言及は、ハイブリダイゼーションのために少なくとも約0から少なくとも約15% v/vのホルムアミドおよび少なくとも約1Mから少なくとも約2Mの塩、ならびに洗浄条件のために少なくとも約1Mから少なくとも約2Mの塩を含み、包含する。一般に、低ストリンジェンシーは、約25〜30℃から約42℃である。この温度は、変更することができ、使用される温度が高いほど、ホルムアルデヒドが置換され、および/または代替ストリンジェンシー条件がもたらされる。必要な場合には、ハイブリダイゼーションのために少なくとも約16% v/vから少なくとも約30% v/vのホルムアミドおよび少なくとも約0.5Mから少なくとも約0.9Mの塩、ならびに洗浄条件のために少なくとも約0.5Mから少なくとも約0.9Mの塩を含み、包含する中ストリンジェンシー、またはハイブリダイゼーションのために少なくとも約31% v/vから少なくとも約50% v/vのホルムアミドおよび少なくとも約0.01Mから少なくとも約0.15Mの塩、ならびに洗浄条件のために少なくとも約0.01Mから少なくとも約0.15Mの塩を含み、包含する、高ストリンジェンシーなどの代替ストリンジェンシー条件を適用することができる。一般に、洗浄は、Tm=69.3+0.41(G+C)%(Marmur et al., J. Mol. Biol. 5: 109, 1962)で行われる。しかし、2本鎖DNAのTmは、ミスマッチ塩基対の数が1%増加するごとに1℃減少する(Bonner et al., Eur. J. Biochem. 46: 83, 1974)。ホルムアミドは、これらのハイブリダイゼーション条件では任意である。従って、特に好ましいストリンジェンシーレベルは、次のように定義される:低ストリンジェンシーは、25〜42℃で、6×SSC緩衝液、0.1% w/v SDSであり;中ストリンジェンシーは、20℃から65℃の範囲の温度で、2×SSC緩衝液、0.1% w/v SDSであり;高ストリンジェンシーは、少なくとも65℃の温度で、0.1×SSC緩衝液、0.1% w/v SDSである。
【0084】
好ましくは、改変Blimp遺伝子は、Blimpの対立遺伝子の全てまたは一部分を含む核酸構築物を使用し、それにレポーター分子をコードする核酸配列を挿入することにより改変される。
【0085】
レポーター分子は、便利には、レポーター発現カセットまたはレポーター構築物によりコードされる。レポーター構築物は、Blimp-1調節要素の制御下で導入され得、および細胞、組織または生物におけるBlimp-1発現パターンを忠実にレポートすることができる。
【0086】
「レポーター」とは、レポーター遺伝子により一般にコードされ、レポーターアッセイで測定される任意の分子、タンパク質またはポリペプチドを意味する。レポーターは、遺伝子配列の活性の理解を可能にする検出可能なシグナルを提供する。それらは、活性を直接レポートすることができ、または下流の標的の活性をモニターすることにより間接的にモニターすることができる。レポータータンパク質は、他のタンパク質と区別可能でなければならず、理想的には容易に定量されなけらばならない。それによって決定されたエピトープと抗体の間の反応性は、任意で二次またはさらに進んだ抗体と共に、容易に利用することができる。一般的なレポータータンパク質としては、ルシフェラーゼ、クロラムフェニコールトランスフェラーゼ(CAT)、β-ガラクトシダーゼ(B-gal)、または蛍光タンパク質、例えば緑色蛍光タンパク質(GFP)が挙げられる。本明細書においてGFPへの言及は、任意の蛍光または発光タンパク質(クラゲまたは他の生物に由来するものを含む)、およびDSRed、HcRed(Clontech)またはhrGFP(Stratagene)などの色変異体を含む全ての相同体、誘導体、類似体を包含することを意味する。好ましくは、レポーター発現カセットは、蛍光または他の発光GFPをコードする。GFPレポーターは、生細胞において容易に検出することができ、細胞選別用途に特に有用であり、好ましい。
【0087】
蛍光または発光マーカーの例は、次の表2に含まれているものであるが、これらに限定されない中から選択することができる。
【0088】
(表2)
1 Ex:ピーク励起波長(nm)
2 Em:ピーク発光波長(nm)
【0089】
蛍光発光を分析する任意の好適な方法は、本発明に包含される。これに関して、本発明は、例えばLakowicz et al., Biophys. J. 72: 567, 1997により開示されているような、二光子および三光子時間分解蛍光顕微鏡;例えば参照として本明細書に組み入れられるEriksson et al., Biophys. J. 2: 64, 1993に開示されているような、蛍光寿命イメージング;および例えばYouvan et al., Biotechnology et Elia 3: 1-18, 1997により開示されているような、蛍光共鳴エネルギー移動を含むが、それらに限定されない技法を企図する。
【0090】
本発明に従って使用することができる例示的な蛍光体としては、Dowerら(国際特許公開公報第93/06121号)により開示されたものが挙げられる。好ましくは、蛍光染料を利用する。蛍光色素を本レポーター分子に組み込むために、任意の適する蛍光染料を使用することができる。例えば、非常に多数の蛍光色素を記載している米国特許第5,573,909号(Singerら)および同第5,326,692号(Brinkleyら)を参照することができる。米国特許第5,227,487号、同第5,274,113号、同第5,405,975号、同第5,433,986号、同第5,442,045号、同第5,451,663号、同第5,453,517号、同第5,459,276号、同第5,515,864号、同第5,648,270号および同第5,723,218号に記載されている蛍光色素も参照することができる。
【0091】
最新のフローサイトメータは、これらのタスクを1秒あたり100,000細胞/粒子まで行うことができる。フィルターおよびダイクロイックミラーの光学アレイの使用により、異なる蛍光波長を同時に分離、検出することができる。加えて、異なる励起波長を有する多数のレーザーを使用することができる。それ故、様々な蛍光体を使用して、ターゲティングし、例えば個々の細胞の細胞内および細胞外特性を検査することができる。散乱光測定により、個々の細胞のサイズ、形状、粒度および/または複雑性ならびに、それ故、対象となる特定の集団への所属を分類することもできる(Shapiro, Practical flow cytometry, 3rd Ed., Brisbane, Wiley-Liss, 1995)。
【0092】
本発明の方法において使用することができる好適なフローサイトメトリーとしては、単一励起レーザー、一般には488nmのスペクトル線を用いて15mWで動作するアルゴンイオン空気冷却レーザー、を使用して、5〜9の光学パラメータを測定するもの(表3参照)が挙げられる。より最新式のフローサイトメータは、アルゴンイオンレーザー(488または514nm)に加えてHeNeレーザー(633nm)またはHeCdレーザー(325nm)などの多重励起レーザーを使用することができる。担体についての、異なる光学検出可能/定量可能な属性に対応する光学パラメータを、フローサイトメータによって測定して、その担体のコード(または多次元空間でのアドレス指定能力)をもたらす質的および/または量的情報のマトリックスを生じることができる。
【0093】
例えば、Biggsら(Cytometry 36: 36-45, 1999)は、細胞を免疫表現型解析する(すなわち、分類する)の目的のために、3つの励起レーザーを使用する11パラメータのフローサイトメータを構築し、前方および側方散乱測定に加えて9つの区別可能なフルオロフォア(fluorophore)の使用を実証した。現在市販されている最大パラメータ数は、17:前方散乱、側方散乱、および各々が5つの蛍光検出器を有する3つの励起レーザー、である。全てのパラメータを適切に使用することができるかどうかは、吸光係数、量子収率、および全ての蛍光体の間のスペクトルのオーバーラップ量に依存する(Malemed et al., "Flow cytometry and sorting" ,2nd Ed., New York, Wiley-Liss, 1990)。しかし、本発明は、いずれの特定のフローサイトメータにも、いずれの特定のパラメータセットにも限定されないことは理解されるだろう。これに関して、本発明は、従来のフローサイトメータの代わりに、例えばFu et al., Nature Biotechnology 17: 1109-1111, 1999に開示されているような微細加工フローサイトメータの使用も企図する。
【0094】
(表3)フローサイトメータにより測定することができる例示的な光学パラメータ
* 488nm励起レーザーを使用
† バンドパス(bandpass)フィルタの幅
# ロングパス(longpass)フィルタ
【0095】
この選別能力を有するフローサイトメータは、「蛍光活性化細胞選別装置」(FACS)として公知である。従って、検出可能にユニークな担体の集団を得る本発明の方法における選別段階は、蛍光活性化細胞選別(FACS)などによるフローサイトメトリー技法によって行うことができる。とはいえ、本発明に関して、FACSは、より正確には「蛍光活性化担体または固体支持体選別」である(例えば、"Methods in Cell Biology" Vol. 33, Darzynkiewica, Z. and Crissman, H. A. , eds. , Academic Press参照)。
【0096】
さらなる実施形態において、本発明は、造血細胞系の細胞を表現型解析および/またはモニターするための方法を提供し、この方法は、改変Blimp遺伝子を含む遺伝子改変細胞またはそのような細胞を含む非ヒト生物のスクリーニングを含み、この場合、遺伝子の発現は、細胞表現型および/または最終分化への細胞の決定付けの指標となる。造血細胞としては、B細胞、T細胞、樹状細胞、マクロファージおよびナチュラルキラー細胞、顆粒球、好酸球、赤血球、巨核球、骨髄、間質、脾細胞前駆細胞ならびにこれらの誘導体が挙げられるが、それらに限定されるわけではない。
【0097】
好ましくは、改変Blimp遺伝子は、発現されるとき、レポーター分子と共発現される、Blimpまたはその一部、フラグメントもしくは機能的な形態を産生する、Blimpコード配列またはその一部、フラグメントもしくは機能形態およびレポーター分子コード配列を含むBlimp mRNA転写産物をコードし、この場合、レポーター分子の検出は、細胞表現型および/または最終分化への細胞の決定付けの指標となる。
【0098】
さらなる態様において、レポーター活性またはレポーター活性の変更を示す細胞を、レポーター活性を示さない細胞の中から単離または選択する。レポーター活性細胞の単離は、フローサイトメトリー、レーザー走査サイトメトリー、クロマトグラフィーおよび/または他の同等の手順によるものであり得る。加えて、さらなる選択マーカーを使用して、本発明の改変細胞を単離または選択することができる。フローサイトメトリー単離が特に好ましい。
【0099】
好ましくは、細胞は、B細胞系列の細胞集団において同定または単離されたASCである。
【0100】
従って、本発明は、実質的にB細胞の集団から実質的に精製されたASC集団を単離する方法を提供し、該方法は、改変Blimp遺伝子を含む遺伝子改変細胞またはそのような細胞を含む非ヒト生物を、ASCへの分化を誘導することができる薬剤または組成物と接触させることを含み、該遺伝子の発現または活性は、レポーター構築物によりレポートされ、ならびに該レポーター活性の検出は、レポーター分子活性を有する細胞がASCであることの指標となり、必要な場合には、該生物からB細胞を単離し、レポーター分子の活性を基にASCを単離する。
【0101】
好ましくは、改変細胞は、発現されるとき、レポーター分子と共発現されるBlimpまたはその一部、フラグメントもしくは機能的な形態を産生する、Blimpコード配列またはその一部、フラグメントもしくは機能的な形態およびレポーター分子コード配列を含むBlimp mRNA転写産物をコードする改変Blimp遺伝子を含み、レポーター活性は、レポーター分子活性を有する細胞がASCであることの指標となる。
【0102】
好ましくは、細胞のスクリーニングは、蛍光レポーター分子のフローサイトメトリー分析により達成される。
【0103】
B細胞は、例えば密度勾配遠心分離、フローサイトメトリー、または磁性ビーズの使用により、生物またはサンプルから便利に単離される。B細胞を選択的に、クローン的にもしくは多クローン的にまたは別様に有効に活性化し、それらのASCへの分化を誘導する任意の薬剤または組成物を包含する。多クローン性活性化因子の一例は、LPSである。
【0104】
一つの態様において、レポーターはGFPであり、該ASCは、フローサイトメトリーにより単離される。
【0105】
「実質的に精製された」とは、ASCが、細胞の少なくとも約60から95%、好ましくは少なくとも約97%、さらに好ましくは少なくとも約99%を含み、例えば、少なくとも約60、61、62、63、64%およびそれに続く100%までの級数における連続的な数字である。または、未選別細胞より約100,000倍の濃縮が企図される。
【0106】
本発明は、ワクチンの抗原性、またはASCによる抗体産生を強化または抑制する薬剤の能力を検査するための方法も提供し、低減されたレポーター活性は、抗体応答およびレポーター活性を下方調節もしくは阻害する薬剤の指標となり、または対照を基準にして強化されたレポーター活性は、抗体応答の陽性調節因子である薬剤の指標となる。この局面によると、この方法は以下の段階を含む:
(i)遺伝子改変細胞またはそのような細胞を含む非ヒト動物に薬剤またはワクチンを投与する段階であって、細胞または生物は、発現されるとき、レポーター分子と共発現される、Blimpまたはその一部、フラグメントもしくは機能的な形態を産生する、Blimpポリペプチドをコードする改変Blimp-1遺伝子を含む、段階;
(ii)その存在が、細胞表現型およびASCによる抗体産生を調節する該薬剤またはワクチンの能力の指標となる、レポーター分子の存在について細胞または生物を検査する段階。
【0107】
もう一つの態様において、本発明は、遺伝子組成物またはタンパク質性組成物を含むワクチンの抗原性または免疫原性を検査するための方法を提供し、この方法は以下の段階を含む:
(i)遺伝子改変細胞またはそのような細胞を含む非ヒト動物にワクチンを投与する段階であって、細胞または生物は、発現されるとき、レポーター分子と共発現される、Blimpまたはその一部、フラグメントもしくは機能的な形態を産生する、Blimpポリペプチドをコードする改変Blimp-1遺伝子を含む、段階;ならびに
(ii)その存在が、T細胞および/またはB細胞の活性化を調節するワクチンの能力の指標となる、レポーター分子の存在について細胞または生物を検査する段階。
【0108】
一部の態様において、Blimp遺伝子は、Blimpコード配列またはその一部、フラグメントもしくは機能的な形態およびレポーター分子コード配列を含む、Blimp mRNA転写産物をコードする。他の態様において、レポーター分子コード配列は、Blimp対立遺伝子のイントロン内に挿入される。さらなる態様において、改変Blimp対立遺伝子は、ホモ接合またはヘテロ接合の形態で存在する。アッセイの目的に依存して、一部の態様において、改変Blimp対立遺伝子は、機能的Blimp転写因子またはその機能的な部分、形態、相同体もしくは変異体をコードする。他の態様において、改変Blimp対立遺伝子は、非機能的Blimp転写因子またはその非機能的な部分、形態、相同体もしくは変異体をコードする。実例となる態様において、細胞または遺伝物質は、ヒト、非ヒト霊長類、家畜、愛玩生物、実験検査生物、爬虫類または両生類に由来する。実験検査動物の例としては、齧歯動物(マウスを含む)、モルモット、ブタ、カモ、ウサギまたはヒツジが挙げられる。
【0109】
本明細書において開示する方法の幾つかの実例となる態様において、細胞は、造血細胞または胚細胞である。本明細書において開示する場合のBlimpは、B細胞とT細胞両方の最終分化に不可欠であり、従って、好ましい細胞系列は、リンパ球細胞である。特定の態様において、リンパ球細胞タイプは、B細胞およびT細胞から選択される。B細胞の場合、その最終分化形態は、ASCであり、これらの細胞は、本明細書において開示する方法を使用して、さらに、実質的に精製することができる。細胞がB細胞である場合、最終分化T細胞としては、CD4+ T細胞およびCD8+ T細胞が挙げられるが、これらに限定されるわけではない。便利には、レポーター分子の検出は、細胞表現型および/または特定の条件下または試験薬剤の存在下での最終分化への細胞の決定付けの指標または予測となる。さらにいっそいう便利には、レポーター分子は、蛍光または発光レポーター分子である。
【0110】
本発明は、細胞分化を調節する際に使用するための、本明細書において記載の方法により同定される、Blimp-1発現またはBlimp-1活性の拮抗薬および作動薬を含む、ASCなどであるが、これに限定されない、細胞の最終分化の拮抗薬または作動薬にも関する。この調節成分、作動薬または拮抗薬が向けられる分子を、本明細書においては総称して「標的」または「標的分子」と呼ぶ。
【0111】
従って、もう一つの態様では、本発明は、造血細胞における最終分化の作動薬または拮抗薬についてのインビボまたはインビトロスクリーニング方法を提供し、この方法は、以下の段階を含む:遺伝子改変細胞またはそのような細胞を含む非ヒト動物に1つまたは複数の薬剤を暴露する段階であって、細胞または生物は、発現されるとき、レポーター分子と共発現されるBlimpまたはその一部、フラグメントもしくは機能的な形態を産生する、Blimpポリペプチドをコードする改変Blimp-1遺伝子を含む、段階;およびレポーター分子の存在またはレポーター分子のレベルの変化について細胞または生物を検査する段階であって、レポーター分子の存在は、最終分化を作動し、またはそれに拮抗する1つまたは複数の薬剤の能力の指標となる、段階。Blimpの作動薬は、造血細胞の最終分化を直接または間接的に誘導し、ならびに例えば癌および/または自己免疫疾患の治療または予防の際に、ならびに病的感染への適切な免疫応答を促進する際に有用である。最終分化細胞の発生を阻害する分子は、狼瘡患者などの自己免疫患者において、またはアレルギーの場合などの免疫機能不全を治療する際に有用である。
【0112】
好ましくは、改変細胞は、発現されるとき、レポーター分子と共発現されるBlimpまたはその一部、フラグメント、相同体、変異体、誘導体、機能的な形態もしくは非機能的な形態を産生する、Blimpコード配列またはその一部、フラグメントもしくは機能的な形態およびレポーター分子コード配列を含むBlimp mRNA転写産物をコードする改変Blimp遺伝子を含む造血細胞であり、ここで、レポーター活性は、レポーター分子活性を有する細胞が最終分化するかまたは最終分化に決定付けられていることの指標となる。さらに好ましくは、造血細胞は、リンパ球系列の細胞である。一部の態様において、最終分化細胞はASCであり、他の態様において、最終分化細胞は、CD4 T細胞および/またはCD8 T細胞である。改変Blimp対立遺伝子は、細胞、組織または非ヒト生物中にホモ接合またはヘテロ接合の形態で存在する。さらに、特定の用途によっては、Blimp対立遺伝子は、転写(機能)活性Blimpポリペプチドを発現する。従って、あるアッセイでは、細胞における最終分化を調節または誘導するために機能的Blimpポリペプチドを有することが有用であろう。他の形態では、改変Blimp対立遺伝子は、機能的Blimpを発現せず、Blimp対立遺伝子が発現されたかどうか、またはBlimp発現のレベルが機能Blimpポリペプチドを産生することができる細胞において調節されたかどうかを、レポーター活性の検出によって判定することで十分であろう。
【0113】
細胞(インビトロ)アッセイは、特に便利であり、細胞においてその活性を容易に検出することができるレポーター分子とカップリングさせたときのアッセイは、ハイスループットスクリーニングに理想的に適する。当業者には公知であるように、多数の異なる形式が利用可能である。一つの有用な例は、Ulleras et al., Toxicology 206 (2) : 245-256, 2005に記載されている。
【0114】
分子または分化状態の「調節」は、その機能活性または分化の全てまたは一部を完全にまたは部分的に阻害または低減または下方調節すること、およびその機能活性または分化の全てまたは一部を強化または上方調節することを含む。分子が、遺伝子配列である場合、その機能活性は、例えば、その結合能力または転写もしくは翻訳活性、またはその半減期を調節することにより、調節することができる。分子が、コードされたポリペプチドである場合、その機能活性は、例えば、その結合能力、その半減期、細胞もしくは膜における位置、またはその酵素活性を調節することにより、調節することができる。調節成分は、調節を達成する作動薬または拮抗薬である。強化された分化は、低減された細胞分裂の指標にもなる。
【0115】
拮抗薬または作動薬の一例は、タンパク質、ポリペプチドまたはペプチドである。これらの用語は、交換可能に用いることができる。これらの用語は、アミノ酸のポリマーおよびその同等物を指し、特定長の産物を指さず、従って、ポリペプチド、ペプチド、オリゴペプチドおよびタンパク質は、ポリペプチドの一定義に包含される。これらの用語は、ポリペプチドの改変、例えば、グリコシル化、アセチル化、リン酸化なども除外しない。この定義に包含されるものは、例えば、表4に与えるものなどの、例えば非天然アミノ酸を含むアミノ酸の1つまたは複数の類似体を含有するポリペプチド、または置換されている結合を有するポリペプチドである。このようなポリペプチドは、細胞に進入することができるために必要であり得る。アミノ酸の化学的類似体を有するポリペプチドは、プロテアーゼ媒介消化に対して、より耐性であり得る。拮抗薬または作動薬の一例は、Blimpの化学的類似体である。拮抗薬および作動薬は、例えばc-myc発現がBlimp-1により抑制されるなど、Blimpが相互作用する分子に作用することができる。
【0116】
遺伝分子は、作動薬および拮抗薬調節成分へも発達させられる。用語「遺伝分子」、「核酸」、「ヌクレオチド」および「ポリヌクレオチド」は、当業者には容易に理解されるように、RNA、cDNA、ゲノムDNA、合成形および混合型ポリマー、センスとアンチセンス鎖の両方を含み、化学的にもしくは生化学的に改変されていてもよいし、または非天然もしくは誘導体化ヌクレオチド塩基を含有していてもよい。このような改変としては、例えば、ラベル;メチル化;1つまたは複数の天然由来ヌクレオチドの類似体(例えば、モルホリン環)での置換;ヌクレオチド間調節、例えば非荷電結合(例えば、メチルホスホネート、ホスホトリエステル、ホスホアミデート、カルバメートなど)、荷電結合(例えば、ホスホロチオエート、ホスホロジチオエートなど)、ペンデント部分(pendent moiety)(例えば、ポリペプチド)、インターカレーター(intercalator)(例えば、アクリジン、ソラレンなど)、キレート化剤、アルキル化剤および改変結合(例えば、α-アノマー核酸など)が挙げられる。水素結合および他の化学的相互作用により指定配列に結合する能力に関してポリヌクレオチドを模倣する合成分子も含まれる。このような分子は当技術分野において公知であり、このような分子としては、例えば、分子骨格内でペプチド結合がリン酸エステル結合と置き換わっているものが挙げられる。アンチセンス分子の改変は周知であり、Kurrek, Eur. J. Biochem. 270:1628-1644, 2003に要約されている。
【0117】
例えばアンチセンスポリヌクレオチド配列は、転写産物のサイレンシングに有用である。さらに、Blimp遺伝子座の全てまたは一部を含有するポリヌクレオチドベクターを、プロモータの制御下、アンチセンス配向に配置し、細胞に導入することができる。細胞内でのこのようなアンチセンス構築物の発現は、標的転写および/または翻訳に干渉するであろう。そのような分子は、自己免疫状態における免疫応答の抑制に特に有用であり得る。さらに、共抑制、およびRNAiまたはsiRNAを誘導するメカニズムを利用することもできる。またはアンチセンスもしくはセンス分子を直接投与することができる。後者の形態では、アンチセンスまたはセンス分子を組成物に調合し、その後、任意の数の手段により標的細胞に投与することができる。
【0118】
アンチセンスおよびセンス分子に関する変形は、モルホリンヌクレオチド誘導体およびホスホロジアミデート結合からなるオリゴヌクレオチドであるモルホリノの使用を伴う(例えば、Summerton and Weller, Antisense and Nucleic Acid Drug Development 7: 187-195,1997)。このような化合物を胚に注入し、mRNAでの干渉の影響を観察する。
【0119】
一つの態様において、本発明は、Blimpをコードする核酸分子の機能または作用を調節する際に使用するためにオリゴヌクレオチドおよび類似の化学種などの化合物を利用する。すなわち、オリゴヌクレオチドが、転写または翻訳後遺伝子サイレンシングを誘導する。これは、内因性リガンドをコードする1つまたは複数の核酸分子と特異的にハイブリダイズするオリゴヌクレオチドを提供することにより達成される。オリゴヌクレオチドは、細胞に直接提供してもよいし、または細胞内で発生させてもよい。本明細書において用いる場合、用語「標的核酸」および「阻害剤をコードする核酸分子」は、阻害剤をコードするDNA、そのようなDNAから転写されたRNA(プレmRNAおよびmRNAまたはそれらの一部を含む)、ならびにまたそのようなRNAから誘導されたcDNAを包含するように、便宜上、用いている。本発明の化合物とその標的核酸のハイブリダイゼーションを、一般に、「アンチセンス」と呼ぶ。従って、本発明の一部の好ましい態様の実施に含まれると考えられる好ましいメカニズムを、本明細書においては「アンチセンス阻害」と呼ぶ。典型的に、このようなアンチセンス阻害は、少なくとも1つの鎖またはセグメントが切断、分解または別様に作動不能にされるような、オリゴヌクレオチド鎖またはセグメントの水素結合に基づくハイブリダイゼーションに基づく。これに関して、そのようなアンチセンス阻害は、特定の核酸分子およびそれらの機能をターゲットにするのが、本発明では好ましい。
【0120】
干渉されるDNAの機能としては、複製および転写を挙げることができる。例えば、複製および転写は、内因性細胞テンプレート、ベクター、プラスミド構築物またはその他由来であり得る。干渉されるRNAの機能としては、タンパク質翻訳部位へのRNAの転移、RNA合成部位から遠い細胞内の部位へのRNAの転移、RNAからのタンパク質の翻訳、1つまたは複数のRNA種を生じさせるようなRNAのスプライシング、およびRNA内に束縛されるかRNAにより助長され得るRNAの関与する触媒活性または複合体形成、といった機能を挙げることができる。一例では、標的機能へのこのような干渉の結果、Blimpレベルが低下する。本発明の状況において、「調節」および「発現の調節」は、遺伝子をコードする核酸分子、例えばDNAまたはRNAの量またはレベルの増加(刺激)または低下(阻害)のいずれかを意味する。阻害は、多くの場合、発現の調節の好ましい形態であり、mRNAは、多くの場合、好ましい標的核酸である。
【0121】
アンチセンス化合物は、標的核酸への化合物の結合が標的核酸の正常な機能に干渉して活性を喪失させるとき、ならびに特異的結合が望まれる条件下、すなわちインビボアッセイまたは治療的処置の場合には生理条件下、およびインビトロアッセイの場合にはアッセイを行う条件下で、非標的核酸配列へのアンチセンス化合物の非特異的結合を回避するために十分な程度の相補性があるとき、特異的にハイブリダイズすることができる。
【0122】
本明細書において用いる場合、「相補的な」は、オリゴマー化合物の2つの核酸塩基間の正確な対合能力を指す。例えば、オリゴヌクレオチド(オリゴヌクレオチド化合物)の一定の位置の核酸塩基が、標的核酸の一定の位置の核酸塩基と水素結合することができる場合(標的核酸は、DNA、RNAまたはオリゴヌクレオチド分子である)には、そのオリゴヌクレオチドと標的核酸間の水素結合の位置は、相補的位置と考える。そのオリゴヌクレオチドとさらなるDNA、RNAまたはオリゴヌクレオチド分子は、各分子中の十分な数の相補的位置が、互いに水素結合することができる核酸塩基によって占有されているとき、互いに相補的である。従って、「特異的にハイブリダイズ可能な」および「相補的な」は、安定で特異的な結合が、オリゴヌクレオチドと標的核酸の間に発生するような十分な数の核酸塩基に関する正確な対合または相補性の十分な程度を示すために使用する用語である。
【0123】
本発明によると、化合物としては、アンチセンスオリゴマー化合物、アンチセンスオリゴヌクレオチド、リボザイム、外部ガイド配列(EGS)オリゴヌクレオチド、可変スプライサー(alternate splicer)、プライマー、プローブ、および標的核酸の少なくとも一部分にハイブリダイズする他のオリゴマー化合物が挙げられる。従って、これらの化合物は、1本鎖、2本鎖、環状またはヘアピンオリゴマー化合物の形態で導入することができ、ならびに内部または末端バルジ(bulge)またはループなどの構造要素を含有することができる。系に導入されると、本発明の化合物は、標的核酸の改変を果たすように、1つまたは複数の酵素または構造タンパク質の作用を惹起する。このような酵素の1つの非限定的な例は、RNアーゼH(RNA:DNA二重鎖のRNA鎖を切断する細胞エンドヌクレアーゼ)である。「DNA様」である1本鎖アンチセンス化合物が、RNアーゼHを惹起することは、当技術分野においては公知である。従って、RNアーゼHの活性化は、結果としてRNA標的の切断を生じさせ、それにより遺伝子発現のオリゴヌクレオチド媒介阻害の効率が大きく上昇する。同様の役割が、酵素のRNアーゼIIIおよびリボヌクレアーゼLファミリーのものなどの他のリボヌクレアーゼにおいて仮定されている。
【0124】
アンチセンス化合物の好ましい形態は、1本鎖アンチセンスオリゴヌクレオチドであるが、多くの種において、2本鎖構造、例えば2本鎖RNA(dsRNA)分子の導入は、遺伝子またはその関連遺伝子産物の機能の、強力で特異的なアンチセンス媒介低下を誘導することが証明されている。この現象は、植物においても動物においても発生する。
【0125】
本発明の状況において、用語「オリゴマー化合物」は、複数のモノマー単位を含むポリマーまたはオリゴマーを指す。本発明の状況において、用語「オリゴヌクレオチド」は、リボ核酸(RNA)もしくはデオキシリボ核酸(DNA)またはその模倣体、キメラ、類似体および相同体のオリゴマーまたはポリマーを指す。この用語は、天然由来核酸塩基、糖および共有結合性ヌクレオシド間(骨格)結合からなるオリゴヌクレオチド、ならびに同様に機能する天然由来タンパク質を有するオリゴヌクレオチドを包含する。このような改変または置換オリゴヌクレオチドは、例えば細胞取り込み向上、標的核酸に対する親和性向上およびヌクレアーゼの存在下での安定性上昇などの望ましい特性のため、多くの場合、天然形態より好ましい。
【0126】
オリゴヌクレオチドは、本発明の化合物の好ましい形態であるが、本発明は、本明細書において記載するものなどのオリゴヌクレオチド、類似体および模倣体を含むが、これらに限定されるわけではない、化合物の他のファミリーも企図する。
【0127】
翻訳開始コドンと翻訳終結コドンの間の領域を指すことが当技術分野において公知である、オープンリーディングフレーム(ORF)または「コード領域」は、有効にターゲティングすることができる領域である。本発明の状況の中で、1つの領域は、遺伝子のオープンリーディングフレーム(ORF)の翻訳開始または終結コドンを包含する遺伝子内領域である。
【0128】
他の標的領域としては、翻訳開始コドンから5’方向にあるmRNAの一部を指すことが当技術分野においては公知であり、従って、mRNA(またはその遺伝子の対応するヌクレオチド)の5’キャップ部位と翻訳開始コドンの間のヌクレオチドを含む、5’非翻訳領域(5’UTR);ならびに翻訳終結コドンから3’方向にあるmRNAの一部を指すことが当技術分野においては公知であり、従って、mRNA(またはその遺伝子の対応するヌクレオチド)の翻訳終結コドンと3’末端の間のヌクレオチドを含む、3’非翻訳領域(3’UTR)が挙げられる。mRNAの5’キャップ部位は、5’-5’三リン酸結合によりmRNAの5’の大部分の残基に連結しているN7-メチル化グアノシン残基を含む。mRNAの5’キャップ領域は、5’キャップ構造それ自体、ならびにそのキャップ部位に隣接する最初の50のヌクレオチドを含むと考えられる。5’キャップ領域のターゲティングも好ましい。
【0129】
当技術分野において公知であるように、ヌクレオシドは、塩基と糖の組み合わせである。ヌクレオシドの塩基部分は、一般には複素環式塩基である。そのような複素環式塩基の2つの最も一般的なクラスは、プリンおよびピリミジンである。ヌクレオチドは、そのヌクレオシドの糖部分に共有結合で連結されたリン酸基をさらに含むヌクレオシドである。ペントフラノシル糖を含むヌクレオシドについては、リン酸基をその糖の2’、3’または5’ヒドロキシル部分のいずれかに連結させることができる。オリゴヌクレオチドを形成する場合、リン酸基は、隣接するヌクレオシドと互いに共有結合で連結して、線状ポリマー化合物を形成する。そして次に、この線状ポリマー化合物のそれぞれの末端をさらに連結させて、環状化合物を形成することができるが、線状化合物が一般に好ましい。加えて、線状化合物は、内部核酸塩基相補性を有することがあり、従って、完全または部分的2本鎖化合物を生じさせるような様式でフォールディングすることがある。オリゴヌクレオチド内のリン酸基は、そのオリゴヌクレオチドのヌクレオシド間骨格を形成すると一般に言及されている。RNAおよびDNAの正常な結合または骨格は、3’-5’ホスホジエステル結合である。
【0130】
本発明において有用な好ましいアンチセンス化合物の具体的な例としては、改変された骨格または非天然ヌクレオシド間結合を含有するオリゴヌクレオチドが挙げられる。本明細書において定義するように、改変された骨格を有するオリゴヌクレオチドは、その骨格内にリン原子を保持するもの、およびその骨格内にリン原子を有さないものを含む。本明細書の目的のために、および当技術分野において時として参照されるように、ヌクレオシド間骨格内にリン原子を有さない改変オリゴヌクレオチドもオリゴヌクレオシドとみなすことができる。
【0131】
中にリン原子を含有する好ましい改変オリゴヌクレオチド骨格としては、例えば、正常な3’-5’結合を有する、ホスホロチオエート、キラルホスホロチオエート、ホスホロジチオエート、ホスホトリエステル、アミノアルキルホスホトリエステル、メチルおよび他のアルキルホスホネート(3’-アルキレンホスホネート、5’-アルキレンホスホネートを含む)およびキラルホスホネート、ホスフィネート、ホスホルアミデート(3’-アミノホスホルアミデートを含む)およびアミノアルキルホスホルアミデート、チオノホスホルアミデート、チオノアルキルホスホネート、チオノアルキルホスホトリエステル、セレノホスフェートおよびボラノホスフェート、および、これらの2’-5’連結型類似体、ならびに1つまたは複数のヌクレオチド間結合が3’-3’、5’-5’、または2’-2’結合である、逆極性を有するものが挙げられる。逆極性を有する好ましいオリゴヌクレオチドは、3’の大部分のヌクレオチド間結合に単一3’-3’結合、すなわち、無塩基であり得る(核酸塩基が欠失しているか、その位置にヒドロキシル基を有する)単一の逆ヌクレオシド残基、を含む。様々な塩、混合塩および遊離酸形態も含まれる。
【0132】
本発明の単離または組換え作動薬および拮抗薬は、直接使用されるか、薬学的試薬または他の試薬としてのそれらの有効性を向上させるために、当技術分野において周知の方法によりさらに改変することができる。活性化合物についての重要な考慮事項としては、調合物および送達方法が挙げられる。
【0133】
作動薬または拮抗薬は、遺伝子形またはタンパク質形の標的の全てまたは一部により決定される分子、例えば、抗体、模倣体またはアンチセンス分子を含む。
【0134】
この点に関しては、抗イディオタイプ抗体、キメラ抗体およびヒト化抗体を含む抗体が有用であり、現在、それらの作製は、当業者には日常的である。ペプチドまたは非ペプチド模倣体は、機能のために重要である標的分子の残基を同定することにより、標的の作動薬として開発することができる。標的分子と相互作用し、改善された薬理学的特性を有する分子を設計するために、モデリングを使用することができる。全てのそのような分子は、細胞に進入できるように改変する必要があるだろう。
【0135】
合理的な創薬により、対象となる生物活性ポリペプチドのまたはそれらが相互作用する小分子(例えば、作動薬、拮抗薬、阻害剤またはエンハンサー)の構造類似体を産生させて、例えばポリペプチドのさらに活性もしくは安定な形態である薬物、または例えばポリペプチドの機能を強化するか、ポリペプチドの機能に干渉する薬物を形成することができる。例えば、Hodgson(Bio/Technology 9: 19-21, 1991)を参照されたい。一つのアプローチでは、X線結晶学により、コンピュータモデリングにより、または最も典型的にはアプローチの併用により、対象となるタンパク質の三次元構造を、先ず、決定する。ポリペプチドの構造に関する有用な情報は、相同タンパク質の構造に基づくモデリングによっても得ることができる。合理的な創薬の一例は、HIVプロテアーゼ阻害剤の開発である(Erickson et al., Science 249 : 527-533,1990)。加えて、アラニンスキャンによって標的分子を分析することができる(Wells, Methods Enzymol. 202 : 2699-2705, 1991)。この技法では、アミノ酸残基をAlaにより置換し、ペプチド活性に対するその影響を判定する。そのペプチドの各アミノ酸残基をこの要領で分析して、そのペプチドの重要な領域を決定する。
【0136】
機能的アッセイにより選択した標的特異的抗体を単離すること、およびその後、その結晶構造を解明することも可能である。原則として、このアプローチは、その後の創薬の基礎となり得るファーマコア(Pharmacore)を生じさせる。機能的薬理活性抗体に対する抗イディオタイプ抗体(抗id)を作製することにより、タンパク質結晶学を全て回避することができる。鏡像の鏡像として、抗idの結合部位は、元の受容体の類似体であると予想されよう。その後、その抗idを使用して、化学的または生物学的に産生させたペプチドバンクのバンクからペプチドを同定および単離することができる。その後、選択されたペプチドはファーマコアとしての機能を果たすであろう。
【0137】
本明細書において考えられる類似体は、側鎖の改変;ペプチド、ポリペプチドまたはタンパク質合成中の非天然アミノ酸および/またはそれらの誘導体の組み込み;架橋剤の使用;ならびにタンパク質性分子またはそれらの類似体に配座的拘束を負わせる他の方法を含むが、これらに限定されるわけではない。
【0138】
本発明により考えられる側鎖改変の例としては、アルデヒドとの反応による還元アルキル化、その後のNaBH4での還元;メチルアセトイミデートでのアミジン化;無水酢酸でのアシル化;シアネートでのアミノ基のカルバモイル化;2,4,6-トリニトロベンゼンスルホン酸(TNBS)でのアミノ基のトリニトロベンジル化;無水コハク酸および無水テトラヒドロフタル酸でのアミノ基のアシル化;ならびにピリドキサル-5-リン酸でのリジンのピリドキシル化、その後のNaBH4での還元などによる、アミノ基の改変が挙げられる。
【0139】
アルギニン残基のグアニジン基は、2,3-ブタンジオン、フェニルグリオキサールおよびグリオキサールなどの試薬での複素環式縮合生成物の形成によって改変することができる。
【0140】
カルボキシル基は、O-アシルイソウレア形成によるカルボジイミド活性化、その後の、例えば対応するアミドへの、二次的誘導体化によって改変することができる。
【0141】
スルフヒドリル基は、ヨード酢酸またはヨードアセトアミドでのカルボキシメチル化;システイン酸への過ギ酸酸化;他のチオール化合物との混合ジスルフィドの形成;マレイミド、無水マレイン酸または他の置換マレイミドとの反応;4-クロロメルクリベンゾエート、4-クロロメルクリフェニルスルホン酸、フェニルメルクリクロライド、2-クロロメルクリ-4-ニトロフェノールおよび他の水銀剤を使用する水銀性誘導体の形成;アルカリ性pHでのシアネートでのカルバモイル化などの方法によって改変することができる。
【0142】
トリプトファン残基は、例えば、N-ブロモスクシンイミドでの酸化または2-ヒドロキシ-5-ニトロベンジルブロマイドもしくはスルフェニルハライドでのインドール環のアルキル化によって改変することができる。一方、チロシン残基は、テトラニトロメタンでニトロ化して3-ニトロチロシン誘導体を形成することによって変性することができる。
【0143】
ヒスチジン残基のイミダゾール環の改変は、ヨード酢酸誘導体でのアルキル化またはジエチルピロカルボネートでのN-カルボエトキシル化によって達成することができる。
【0144】
ペプチド合成中に非天然アミノ酸および誘導体を組み込む例としては、ノルロイシン、4-アミノ酪酸、4-アミノ-3-ヒドロキシ-5-フェニルペンタン酸、6-アミノへキサン酸、t-ブチルグリシン、ノルバリン、フェニルグリシン、オルニチン、サルコシン、4-アミノ-3-ヒドロキシ-6-メチルヘプタン酸、2-チエニルアラニンおよび/またはアミノ酸のD異性体の使用が挙げられるが、これらに限定されるわけではない。本明細書で考えられる非天然アミノ酸のリストを表4に示す。
【0145】
(表4)非典型アミノ酸についてのコード
【0146】
例えば3D配座を安定させるために、架橋剤を使用することができ、例えば、n=1からn=6の(CH2)nスペーサー基を有する二官能性イミドエステル、グルタルアルデヒド、N-ヒドロキシスクシンイミドエステルなどの二官能性架橋剤、ならびにアミノ反応性部分(例えば、N-ヒドロキシスクシンイミド)および別の基に特異的な反応性部分(例えば、マレイミド、またはジチオ部分(SH)またはカルボジイミド(COOH))を通常含有するヘテロ二官能性試薬を使用する。加えて、例えば、CαおよびNα-メチルアミノ酸の組み込み、およびアミノ酸のCα原子とCβ原子の間への二重結合の導入により、ペプチドを配座的に拘束することができる。
【0147】
天然産物、コンビナトリアルまたはファージ提示法は、全て、調節成分をスクリーニングするために利用することができる。ハイスループットスクリーニング法の大量選択を利用することができ、これに本発明の細胞を利用できるようにすることができる。
【0148】
Blimp-1で作動する遺伝子ネットワークの他のメンバーを同定する際、ツーハイブリッドスクリーニングも有用である。標的相互作用および調節成分についてのスクリーニングは、2つの物理的に分けることができる機能的ドメインからなる転写因子を利用する酵母ツーハイブリッド系を使用して行うことができる。最も一般的に使用されるのは、DNA結合ドメインおよび転写活性化ドメインからなる酵母GAL4転写活性化因子である。2つの異なるクローニングベクターを使用して、潜在的結合性タンパク質をコードする遺伝子へのGAL4ドメインの別個の融合体を作製する。これらの融合タンパク質は共発現され、核を標的にし、ならびに相互作用が発生する場合にはレポーター遺伝子(例えば、lacZ)の活性化によって検出可能な表現型が生じる。この場合、例えば、S.セレビジエ(S.cerevisiae)を、cDNA GAL4活性化ドメイン融合体を発現するライブラリまたはベクター、およびGAL4に融合したMyb経路構成要素を発現するベクターで共形質転換する。lacZをレポーター遺伝子として使用する場合、融合タンパク質の共発現により青色が生じるであろう。標的と相互作用する小分子または他の候補化合物は、結果として細胞の色の喪失をもたらすであろう。Munderら(Appl. Microbiol. Biotechnol. 52(3): 311-320, 1999)およびYoungら(Nat. Biotechnol. 16(10): 946-950, 1998)が開示したような酵母ツーハイブリッド系を参照することができる。この系によってこのように同定された分子を、その後、本発明の遺伝子改変生物または遺伝子改変細胞において再検査する。
【0149】
組換え、合成または単離された形態の本作動薬および拮抗薬ならびに1つまたは複数の薬学的に許容される担体、希釈剤または賦形剤を含む治療用薬学的組成物が、さらに企図される。癌の治療または免疫応答の調節が特に企図される。
【0150】
治療法という用語は、ある状態または疾病の治療または予防への言及と解釈するものとする。用語「治療する」および「改善する」は、交換可能に用いている。
【0151】
用語「組成物」または「薬剤」または「薬物」は、所望の薬理および/または生理作用を誘導する化学的化合物を指す。この用語は、塩、エステル、アミド、プロドラッグ、活性代謝産物、類似体などを含むが、これらに限定されるわけではない、本明細書において特に述べる化合物の薬学的に許容されるおよび薬理学的に活性な成分も含む。上の用語を用いるときには、それが、活性薬剤それ自体ならびに薬学的に許容される、薬理学的に活性な塩、エステル、アミド、プロドラッグ、代謝産物、類似体などを含むと理解しなければならない。用語「化合物」は、狭く解釈するべきではなく、ペプチド、ポリペプチドおよびタンパク質ならびに遺伝分子、例えばRNA、DNA、およびその模倣体および化学的類似体まで拡大する。
【0152】
フレーズ「疾病もしくは状態を改善する」または「治療」または「治療的」は、最も広い状況で用いており、疾病もしくは状態、または疾病もしくは状態の1つまたは複数の症状、または疾病もしくは状態の症状の頻度におけるあらゆる測定可能なまたは実質的に有意な改善、ならびにその疾病からの完全回復または状態、その症状もしくはその基礎原因の除去を含む。本発明は、疾病または状態の大きな範囲に応用でき、担当技術者は、症例ごとベースで表現型の評価の正確なパラメータを決定しなければならない。状態は、1つまたは複数の疾病に随伴することもあり、またはそのように関連していないこともある。状態の改善は、あらゆる所望の生理学的または心理学的変化を包含する。
【0153】
本組成物の有効量は、当業者により最良に確立される。本明細書において用いる場合の化合物の「有効量」という用語は、所望の治療または生理効果を生じさせるために十分な薬剤の量を意味する。時には望ましくない効果、例えば副作用が、所望の治療効果とともに現れる。それ故、担当者は、何が適切な「有効量」であるかを判定する際、潜在的利益と潜在的リスクとのバランスをとる。必要とされる正確な量は、被験者の種、年齢および全身の健康状態、投与方式などに依存して、被験者ごとに変わるであろう。従って、正確な「有効量」を特定することができないこともある。しかし、いずれの個々の症例においても、適切な「有効量」は、当業者が日常的な経験のみを利用して決定することができる。
【0154】
組換え、合成または単離形態の本発明の作動薬および拮抗薬、ならびに1つまたは複数の薬学的に許容される担体、希釈剤または賦形剤を含む治療用薬学的組成物が、さらに企図される。癌の治療または免疫応答の調節が、特に企図される。
【0155】
治療法という用語は、ある状態または疾病の治療または予防への言及と解釈するものとする。用語「治療する」および「改善する」は、交換可能に用いている。
【0156】
用語「組成物」または「薬剤」または「薬物」は、所望の薬理および/または生理作用を誘導する化学的化合物を指す。この用語は、塩、エステル、アミド、プロドラッグ、活性代謝産物、類似体などを含むが、これらに限定されるわけではない、本明細書において特に述べる化合物の薬学的に許容されるおよび薬理学的に活性な成分も包含する。上記の用語を用いるときには、それが、活性薬剤それ自体ならびに薬学的に許容される、薬理学的に活性な塩、エステル、アミド、プロドラッグ、代謝産物、類似体などを含むと理解しなければならない。用語「化合物」は、狭く解釈するべきではなく、ペプチド、ポリペプチドおよびタンパク質ならびに遺伝分子、例えばRNA、DNA、およびその模倣体および化学的類似体まで拡大する。
【0157】
フレーズ「疾病もしくは状態を改善する」または「治療」または「治療的」は、最も広い状況で用いており、疾病もしくは状態または疾病もしくは状態の1つまたは複数の症状または疾病もしくは状態の症状の頻度におけるあらゆる測定可能なまたは実質的に有意な改善、ならびにその疾病からの完全回復または状態、その症状もしくはその基礎原因の除去を含む。本発明は、疾病または状態の大きな範囲に応用でき、担当技術者は、症例ごとベースで表現型の評価の正確なパラメータを決定しなければならない。状態は、1つまたは複数の疾病に随伴することもあり、またはそのように関連していいこともある。状態の改善は、あらゆる所望の生理学的または心理学的変化を包含する。
【0158】
本組成物の有効量は、当業者により最良に確立される。本明細書において用いる場合の化合物の「有効量」という用語は、所望の治療または生理効果を生じさせるために十分な薬剤の量を意味する。時には望ましくない効果、例えば副作用が、所望の治療効果とともに現れる。それ故、担当者は、何が適切な「有効量」であるかを判定する際、潜在的利益と潜在的リスクのバランスをとる。必要とされる正確な量は、被験者の種、年齢および全身の健康状態、投与方式などに依存して、被験者ごとに変わるであろう。従って、正確な「有効量」を特定することができないこともある。しかし、いずれの個々の症例においても、適切な「有効量」は、当業者が日常的な経験のみを利用して決定することができる。
【0159】
本発明のポリペプチド、核酸、抗体、ペプチド、化学的類似体、作動薬、拮抗薬または模倣体は、薬学的組成物に調合することができ、薬学的組成物は、従来の薬学的調剤法に従って調製される。例えば、Remington's Pharmaceutical Sciences, 18th Ed. (1990, Mack Publishing, Company, Easton, PA, U.S.A.を)参照されたい。組成物は、活性薬剤または活性薬剤の薬学的に許容される塩を含有することができる。これらの組成物は、活性物質の1つに加えて、薬学的に許容される賦形剤、担体、緩衝液、安定剤または当技術分野において周知の他の材料を含むことができる。そのような材料は、非毒性でなければならず、活性成分の有効度に干渉してはならない。担体は、投与、例えば、静脈内投与、経口投与、髄腔内投与または非経口投与、に望ましい製剤の形態に依存して多種多様な形態を取ることができる。
【0160】
経口投与用の化合物は、固体または液体製剤、例えばカプセル、ピル、錠剤、ロゼンジ、粉末、懸濁液または乳剤に調合することができる。経口剤形の組成物を調製する場合、通常の製薬用媒体、例えば、経口液体製剤(例えば、懸濁液、エリキシルおよび溶液など)の場合には、例えば、水、グリコール、油、アルコール、着香剤、保存料、着色剤、懸濁化剤など;または経口固体製剤(例えば、粉末、カプセルおよび錠剤など)の場合には、デンプン、糖、希釈剤、顆粒化剤、滑沢剤、結合剤、崩壊剤など、のいずれを利用してもよい。投与の容易さのため、錠剤およびカプセルが、最も有利な経口単位剤形の代表であり、この場合、固体の製薬用担体が、明らかに利用される。所望される場合、標準的な技法により錠剤に糖衣または腸溶コーチングを施すことができる。活性薬剤をカプセル化して、それを安定化して胃腸管を通過させ、一方、それと同時に、血液脳関門を通過できるようにすることができる。例えば、国際特許公開公報第96/11698号を参照されたい。
【0161】
非経口投与用の化合物は、製薬用担体に溶解し、溶液または懸濁液のいずれかとして投与することができる。実例となる適する担体は、水、食塩水、デキストロース溶液、フルクトース溶液、エタノール、または動物、植物もしくは合成起源の油である。担体は、他の成分、例えば、保存料、懸濁化剤、可溶化剤、緩衝液なども含有することができる。化合物を髄腔内投与する場合、それらを髄液に溶解することもできる。
【0162】
活性薬剤は、好ましくは、治療有効量で投与される。実際の投与量ならびに投与の速度および時間経過は、治療する状態の性質および重症度に依存するであろう。治療の処方、例えば、投薬量の決定、タイミングなどは、一般担当者または専門家の責任の範囲内であり、典型的には治療すべき疾患、個々の患者の状態、送達部位、投与方法および担当者には公知の他の因子を考慮に入れる。技法およびプロトコルの例は、Remington's Pharmaceutical Sciences,(前記)において見出すことができる。
【0163】
または、抗体もしくは細胞特異的リガンドなどのターゲティング系の使用によるターゲティング療法を用いて、一定の細胞タイプにさらに特異的に活性薬剤を送達することができる。ターゲティングは、様々な理由のため、例えば、その薬剤が許容できないほど毒性である場合、またはそれがあまりにも高い投薬量を別様に必要とする場合、またはそれが別様に標的細胞に侵入することができない場合、望ましいであろう。
【0164】
これらの薬剤を直接投与する代わりに、標的細胞において、例えば、上記のものなどのウイルスベクターにおいて、または米国特許第5,550,050号ならびに国際特許公開公報第92/19195号、同第94/25503号、同第95/01203号、同第95/05452号、同第96/02286号、同第96/02646号、同第96/40871号、同第96/40959号および同第97/12635号に記載されているものなどの細胞ベースの送達系において産生させることができる。ベクターを標的細胞にターゲティングし、発現産物の発現を特定の細胞、発達段階または細胞周期段階に限定することができるだろう。細胞ベースの送達系は、患者体内の所望の標的部位に移植するために設計され、ならびに標的薬剤のコード配列を含有する。または、治療すべき細胞において産生されたまたは治療すべき細胞にターゲティングされた活性化薬剤により活性形態に転化させるために、薬剤を前駆体形で投与することもできる。例えば、欧州特許出願第0 425 731A号および国際特許公開公報第90/07936号を参照されたい。
【0165】
以下の非限定的実施例により、本発明をさらに説明する。
【実施例】
【0166】
実施例1
Blimp-1突然変異対立遺伝子(Blimpgfp)の作製
イントロン3’からエキソン6に、スプライス受容体、3つ全てのリーディングフレーム内の停止コドン、内因性リボソーム認識部位(IRES)、eGFPをコードするcDNA、および転写を停止させるSV40ポリアデニル化シグナルからなるeGFP発現カセットが挿入されている、Blimp-1ターゲティング構築物を産生させた。組み込まれたターゲティングベクターでの胚性幹(ES)細胞の選択を可能にするために、PGK-Neoγ遺伝子もそのイントロンに挿入されている。それらのeGFPカセットおよびNeoγカセットをFrt部位に隣接させて、挿入されたDNAのflpリコンビナーゼ媒介欠失を可能にする。Blimp-1ターゲティング構築物、G418により選択される耐性クローンを用いてC57BL/6 ES細胞をエレクトロポレーションし、5’および3’ゲノムDNAプローブへのサザンハイブリダイゼーションによりスクリーニングした(図1C)。Blimpgfp対立遺伝子を有する4つの正確にターゲティングされたクローン(図1C)を、300個のスクリーニングされたクローンから同定した。これらをBALB/c胚盤胞に注入して、キメラ創始者を得た。これらのキメラを増殖させ、生殖系伝達を1つのクローン(4F3)で達成した。
【0167】
実施例2
Blimp-1の発現パターンの記述を可能にするGFPレポーター
当初、Blimp-1は、ASC分化を受けるように誘導されたBリンパ球においてのみ発現されると報告された(Turner et al., (前記))。しかし、その後の研究により、胚形成中(Chang et al., Mech Dev 117:305, 2002)、および骨髄細胞における(Chang et al., (前記), 2000)Blimp-1のより幅広い発現パターンが明らかになった。Blimpgfp対立遺伝子により、造血系列の中でも、より広くは生物においても、Blimp-1の発現パターンをより完全に定義することができる。上で概説したターゲティング戦略は、結果としてBlimpgfp対立遺伝子を生じさせ、それが、内因性Blimp-1調節要素の制御下で2シストロンのmRNAからGFPを発現し、従って、完全Blimp-1発現パターンを反復すると予測される。加えて、この戦略は、Blimp-1 mRNA転写産物を分断して、DNA結合モチーフを含有するジンクフィンガードメインを欠くトランケートされたバージョンのBlimp-1タンパク質(エキソン1〜6)を産生させる。これに従って、インビトロでLPSを用いて分化するように誘導されたBlimpgfp/+ B細胞のウエスタンブロッティングは、野生型Blimp-1タンパク質バンドおよびトランケート型Blimp-1タンパク質バンドの両方を示した(図1D)。生細胞におけるGFP発現および固定された組織におけるBlimp-1タンパク質のモニタリングにより、インビボおよびインビトロでのBリンパ球の遺伝子活性および分化の運命を単細胞レベルでモニターすることができる。
【0168】
実施例3
ASCにおけるBlimp-1のインビボ発現
Blimpgfp/+マウスにおけるリンパ組織の検査は、高Blimp-1の小集団が、骨髄(0.1〜0.2%)、脾臓(0.4〜0.6%)およびリンパ節(0.1%)において細胞を発現していることを示した(図2)。さらに、GFP+細胞の表現型解析は、それらが、以前に定義された高Synd-1/低B220のASC集団、ならびに以前に十分に特性付けされていなかった低Synd-1〜Synd-1陰性の表現型を表すことを示した(図2、(Underhill et al., (前記))。これらの細胞がASCであることを確認するために、Blimpgfp/+骨髄および脾臓からGFP+細胞を選別し、Ig産生についてのELISpot分析に供した。図3からわかるように、細胞の75〜100%がIg分泌性細胞であった(3回の独立した実験の代表)。さらに、GFP陰性画分の選別は、0.001%のASCの頻度(細胞100,000個あたり1個未満)、これに対して、未選別骨髄におけるこれらの細胞の頻度は、0.05〜0.09%であった(100,000個あたり50〜90個)。従って、Blimpgfp発現性ASCの単離は、未分別細胞の100,000倍の濃縮をもたらし、これらの希少細胞を単離するための実質的に決定的な方法をもたらす。加えて、GFP+ ASC集団における全てのIgアイソタイプを表した(図3)。
【0169】
GFPレポーターマウスを使用してBlimpgfp/+マウスにおけるASCの産生をさらに検査するために、2μgのリポ多糖類(LPS)を静脈内注射し、注射後1〜7日のGFP発現について分析した(LPS注射は、結果として成熟B細胞の多クローン性活性化を生じさせる)。LPS注射は、結果として、注射後3日目にピークに達する脾臓GFP+細胞の数の劇的な増加を、その後、7日周辺に定常状態レベルへの低下を生じさせる(図4A)。ゲートされたGFP+細胞の分析は、ASC分化が、Synd-1+/B220+細胞の出現、続いてSind-1+/B220-の出現に伴い同時波で発生したこと、および最終的に、ASCの画分が、Synd-1-/B220-になることを示した。この分化は、骨髄においても反映され、この場合、GFP+細胞は、Synd-1+/-/B220-定常状態集団を迅速に生じさせるSynd-1+/B220+細胞として、注射後4日目に出現する(図4B)。要約すると、LPS治療は、Blimpgfp/+マウス株を使用して初めて表現型的に定義することができるASC分化の波を誘導する。
【0170】
実施例4
インビトロで誘導されるASCにおけるBlimp-1の発現
ASC系列への決定付けおよびASC系列までの進行に影響を及ぼすパラメータをインビトロで定量分析する方法論を開発した。このシステムは、パーコール(Percoll)勾配遠心分離および磁性ビーズ濃縮により精製し、B細胞増殖およびASCへの分化を誘導する様々な刺激の存在下で培養する、小さな休止B細胞の単離を含む。これらの条件としては、IL-4および抗CD40を使用するT依存性応答の模倣またはLPSを使用するT非依存性反応の模倣が挙げられる。加えて、IL-5を濃度調節しながらこれらの培養物に滴下して分化速度を加速し、および抗IgD(1.19)架橋を行って、抗原特異的応答を活性化することができる。1〜5日目にフローサイトメトリーにより培養物を分析して、BlimpgfpおよびSynd-1+発現性細胞の頻度を測定した。培養物中のASCの数は、ELIspotにより判定した。
【0171】
CD40L/IL-4/IL-5またはLPSを使用するBlimpgfp誘導の時間経過の分析(図5A)は、最初のGFP+細胞が、2日後にLPS培養物において観察されることを示した。その後、4日目における約50%GFP+細胞のピークまで、陽性細胞の数は増加する。対照的に、CD40L/IL-4/IL-5治療は、結果として、より少数のGFP発現性細胞の遅発性誘導を生じさせる。興味深いことに、CD40L/IL-4/IL-5誘導GFP発現性細胞の大部分が、Synd-1+でもあるのに対し、LPSは、Synd-1+とSynd-1-両方のGFP発現性細胞を誘導する(図5B)。全てのGFP+細胞が、実際にASCであるかを判定するために、LPS治療BlimpgfpB細胞からの4つの画分を選別した(図5C)。IgM、IgG3およびIgG2bについてのELIspotアッセイは、培養物中の全てのASCが、GFP+画分(A、B、C)によって表され、Synd-1発現レベルと相関しないことを明確に示す。さらに、ASCの頻度は、GFP+集団間で変わらなかった(図5C)。従って、これらのデータは、Blimpgfpが、ASCの運命のマーカーである一方で、Synd-1+が、インビボと同様にインビトロでもASC活性の亜集団の単なる指標であることを明確に示している(図2)。BlimpgfpおよびSynd-1発現の調節を、上で概説した様々な刺激を使用して検査した。興味深いことに、図5Cに概略を示すように、GFP+/Synd-1+およびGFP+/Synd-1- ASC形成の頻度は、異なる刺激により調節され得る。従って、Blimpgfp発現に続いて、インビトロおよびインビボでのASC誘導の調節を同定するための簡単で決定的な方法論をもたらす。
【0172】
最後に、インビトロ由来Blimpgfp陽性ASCを、静脈内注射の7日後に放射線未照射宿主の骨髄および脾臓において検出することができる(図6)。従って、Blimpgfp対立遺伝子は、インビボでの生存、移動および機能特性に対するASCのインビトロ治療の効果を検査する方法をもたらす。
【0173】
実施例5
Blimp-1は胚形成に必要とされる
ホモ接合のBlimpgfp/gfp動物を産生するために、Blimpgfp/+個体を交雑させた。Blimp-1野生型およびBlimpgfp特異的PCRプライマーを使用して、これらの交雑からの子孫を生後21日目に遺伝子型解析した。Blimpgfp/+マウスが、生存しており健康であったのに対し、Blimpgfp/gfp個体は、同定されなかった。これは、Blimp-1欠失が、結果として胚性致死または早期産後致死を生じさせることを示している(図7)。Blimpgfp/gfp動物が死亡する段階により近い段階を検査するために、Blimpgfp/+マウスの時限交雑から生じた胚を検査した。これらのデータは、Blimpgfp/gfp胚が、胚性段階E15.5ほどもの後期に生存していることを示す。しかし、生存能力がある、より高い齢の個体が、文献に記載されたことはない。Blimp-1が胚形成中に広範に発現されることは公知であり、この発見は、Blimpgfpマウスを使用する分析により支持されている。
【0174】
実施例6
Blimp-1は、抗体産生に不可欠である
Blimpgfp/gfp動物の胚性致死を回避するために、および抗体産生におけるBlimp-1の重要性を直接検査するために、致死量の放射線を照射した同系マウスの胎仔肝臓幹細胞再構成を用いて、造血形全体にわたって機能的なBlimp-1タンパク質を欠く成体マウスを産生させた。これらのBlimpgfp/gfpキメラ動物は、健康であり、検査した全ての造血細胞系列を比較的正常な数、含有している。LPSまたはCD40L/IL-4およびIL-5、いずれかでの刺激後のこれらのマウスにおけるASC集団のインビトロ分析は、主としてsynd-1+であるGFP+ Blimp欠失細胞の存在を示した(図8A)。重要なことに、ELIspotアッセイにより評価したところ、これらの細胞は、抗体を分泌できなかった(図8B)。従って、Blimpgfp、ここに記載するマウスモデルは、ASCを単離するための決定的ツールをもたらすばかりでなく、ホモ接合突然変異Blimpgfp/gfp脾細胞からのBlimp-1発現性細胞の集団の同定を可能にし、それにより、Blimp-1欠失の表現型の根底にあるメカニズムの分析を大いに助長する。
【0175】
実施例7
他の造血系列におけるBlimp-1の発現
Blimpgfpレポーター系により、初めて、造血におけるBlimp-1の発現パターンも定義することができた。上で述べたように、Blimpgfpマウスのリンパ器官の分析は、GFP高産生性集団が、ほぼ排他的にASCであることを示した。しかし、低レベルのGFP産生性細胞も出現した。
【0176】
Blimp-1は、ヒトおよびマウスマクロファージならびに顆粒球により発現されることが報告されている。MCSF-1の存在下での血液単球細胞および骨髄由来マクロファージ培養物のフローサイトメトリー分析は、これらの細胞タイプにおける明確なBlimpgfp発現を示した(図9)。しかし、GFP蛍光は、顆粒球では観察されなかった。対照的に、インビボで単離された樹状細胞は、Blimp-1 mRNA発現を欠く。同様に、flt3Lを有する骨髄細胞の培養から誘導された形質細胞性および従来の樹状細胞は、Blimpgfp蛍光を欠く。しかし、選別された樹状細胞のエクスビボ活性化、またはCpG DNAによるflt3L培養物のインビトロ活性化は、結果として、Blimp-1発現、主として従来の樹状細胞を生じさせた(図10)
【0177】
Blimpgfp/+マウスからの胸腺および休止脾臓の分析は、Blimp-1が、T細胞発達中に発現されないことを実証した。しかし、Blimpgfp発現性T細胞の小集団が、リンパ節に存在した。これらの細胞は、存在する活性化T細胞の小集団を提示する可能性があったため、本発明者らは、コンカナバリンA(T細胞を強力に活性化することが公知である条件)の存在下または不在下、インビトロで、リンパ節T細胞を抗CD3モノクローナル抗体で刺激した。インビトロ活性化T細胞は、Blimpgfpを発現し、インビボ分析を支持した(図12C)。
【0178】
BlimpgfpマウスにおけるNK系列の検査は、検査した他の造血系列とは異なり、NK細胞が、Blimp-1を構成的に発現することを示した。NK細胞は、血液、脾臓および骨髄からNK1.1+/CD122+細胞として同定され、均一にGFP+であることを示した(図12A)。この発現は、IL-15の存在下で培養された成熟NK細胞がGFP+であるためインビトロで維持され、かつ、NK細胞の最終分化を誘導するサイトカイン、例えばIL-21またはIL-12/IL-18によってさらに誘導することができる(図12B)。NK細胞におけるBlimp-1の発現は、Blimp-1特異的モノクローナル抗体でのウエスタンブロッティングによっても確認した。
【0179】
要約すると、Blimpgfpレポーターマウスは、様々な造血系列の後期においてBlimp-1を誘導することが明らかになり、その結果、これらの細胞タイプの成熟の調節因子を同定する方法が得られた。重要なことに、非Bリンパ系細胞タイプにおける比較的低い産生レベルのGFPは、均一ASC集団の単離に干渉しない。
【0180】
実施例8
Blimpgfpマウスを使用する、Blimp-1の癌における役割の検査
ASC分化の検査におけるその有用性に加えて、Blimpgfpレポーターマウスは、この細胞タイプの悪性トランスフォーメーションを検査するために使用することができる。マウスでは形質細胞種およびヒトでは多発性骨髄腫と呼ばれるASCの腫瘍は、IgHイントロン性エンハンサーの制御下、B細胞系列においてv-abl癌遺伝子を発現するEμ-v-ablトランスジェニックマウス(Rosenbaum et al., (前記))において、特異的におよび頻繁に惹起される。これらのマウスをBlimpgfp突然変異マウスと交雑させて、腫瘍の潜在性および発病率に対するBlimp-1遺伝子の一方または両方のコピーの欠失の影響を判定した。2つの結果が考えられる:Blimp-1は、形質細胞分化プログラムを誘導することにより、v-ablトランスフォーメーションの機会に関するウインドウを開くために必要となり得る。このトランスジーンは、初期B細胞における発現にもかかわらず、形質細胞種しか誘導しない(Rosenbaum et al., (前記))。従って、機能的Blimp-1対立遺伝子の欠失により、腫瘍発病率の低下または潜在性の増加が予測されよう。または、大きな比率のv-abl誘導形質細胞種が、再配列されたおよび活性化したc-myc遺伝子も有するので、Mycは、トランスフォーメーションにおける本質的な共同作用性活性であり得る(Rosenbaum et al., (前記))。Blimp-1は、最終ASC分化の間のc-myc発現を抑制すると一般に考えられている(Lin et al., Science 276:596, 1997)。このシナリオでは、機能的Blimp-1の欠失が、持続性c-myc発現を可能にすることとなり、それにより、形質細胞種の発現が加速し得る。
【0181】
Blimp-1が、実際、ASC腫瘍形成において一定の役割を果たすならば、それ故、Blimpgfpレポーター株は、腫瘍進行に対するBlimp-1阻害/誘導の効果を判定するための有用な動物モデルをもたらす。
【0182】
実施例9
T細胞における最終分化の調節におけるBlimpの役割を評価する方法
マウス
Blimpgfpマウス(Kallies et al., J Exp Med 200:967-977, 2004)、Rag1-/-マウスおよびRag2-/-マウスを、C57BL/6バックグラウンドで保持した。Blimpgfp遺伝子型解析および胎仔肝臓キメラを、記載されている(Kallies et al., 2004 (前記))とおり作製した。
【0183】
フローサイトメトリーおよびELISA
CD4(GK1.5)、CD8(53.6.7)、TCRβ(H57-597)、Ly5.2(ALI-4A2)に対するモノクローナル抗体(mAb)を、プロテインG-セファロースカラム(Amersham Pharmacia Biotech)でハイブリドーマ上清から精製し、ビオチン(Pierce Chemical Company)、アロフィコシアニン(APC)およびフィコエリトリン(PE)(ProZyme)に、推奨どおりにコンジュゲートさせた。ビオチン化抗CD25(7D4)およびCD122(Tm-β1)ならびにPEコンジュゲート型抗CD44(IM7)、CD62L(MEL-14)、IFNγ.□□□〜1□.、IL-4(11B11)は、PharMingenから入手した。ビオチン化mAbは、ストレプトアビジン-PEまたはCy5と共に発現された(Southern Biotechnologies Inc.)。LSRサイトメーター(BD Bioscience)で細胞を分析し、高速フローサイトメーター(Moflo cytomation and BD Biosciences)で細胞選別を行った。サイトカインについての細胞内染色は、当技術分野において公知の標準的な手順に従って行った。IFNγおよびIL-10産生についてのELISAは、記載されている(Brady et al., J Immunol 172: 2048-2058, 2004)とおり行った。IL-4 ELISAは、1つのモノクローナル抗体を捕捉試薬として使用し、第二のモノクローナル抗体を検出に使用した。ELISAは、三回重複で行い、組換えタンパク質標準物質を使用して定量した。
【0184】
ウエスタンブロッティング
例えば、Rosenbauerら(Embo J 21:211-20, 2002)が記載したように、全タンパク質抽出物を相当数の細胞から産生させ、ウエスタンブロッティングを行った。抗Blimp-1 mAbは、以前に記載されている(Kallies et al., 2004 (前記))。抗Vav1を使用して、等量のタンパク質負荷を確認した。
【0185】
HSV感染
20μLのPBSで希釈した4×105単純疱疹ウイルス(HSV-1 KOS株)にマウスを感染させた。後ろ足の足蹠と踵の間への皮下注射によって投与した。感染したマウスの脾臓および膝窩リンパ節を、その後、分析のために回収した。
【0186】
インビトロ細胞毒性
gB特異的細胞毒性T細胞リンパ球(CTL)を常用の手順[Belz, 2001]により作製した。感染したマウスから脾臓を除去し、108個の1000Gy照射gB498-505被覆C57BL/6脾細胞を5日間単細胞培養した。細胞毒性は、従来の51Cr放出アッセイで評価した。EL4(H-2b)標的細胞を、Na51Crで1時間標識し、gBペプチドを60分間適用し、2回洗浄し、5,000標的/ウエルでプレーティングした。その後、それらをエフェクター集団と共に5時間インキュベートし、その後、γ計数用に上清を回収した。二倍リンパ球希釈物を三回重複でアッセイし、一方、未処理およびTriton X-100破壊対照は、四回重複で測定した。特異的溶解率は、100×(エフェクターを伴う標的からの51Cr放出-標的のみからの51Cr放出)/(Triton X-100を伴う標的からの51Cr放出)として計算した。T細胞不在下でインキュベートした標的からの51Cr放出レベルは、全Triton X-100媒介51Cr放出の10%未満であった。
【0187】
gB特異的CD8+ T細胞の四量体染色
H-2Kb糖タンパク質と単純疱疹ウイルスの糖タンパクB(gB498-505)由来のペプチド(SSIEFARL)とのMHCクラスI四量体型複合体[Altman, 1996 #102; Allan, 2003 #100]を使用して、ウイルス特異的CD8+ T細胞を同定した。カルボキシ末端膜貫通ドメインがbirAビオチン化モチーフで置換されている組換えH-2Kb分子を、ヒトβ2-ミクログロブリンおよびウイルスペプチドでリフォールディングし、birAでビオチン化し、4:1のモル比でニュートラビジン(neutravidin)-PE(オレゴン州、Eugene、Molecular Probes)と複合させた。室温で、60分間、リンパ球をPBS/BSA/アジド中の四量体型複合体について染色し、その後、抗CD8αAPCで染色し、2回洗浄し、フローサイトメトリーによって分析した。
【0188】
組織学
器官を10%緩衝ホルマリン中で固定し、パラフィンに包埋し、薄片に切り、ヘマトキシリン/エオシンで染色した。
【0189】
実施例10
Blimp発現は、IL-21により誘導することができ、ナイーブCD4+およびCD8+ T細胞の活性化エフェクター細胞への成熟に関する重要な成分であり、正常なリンパ球恒常性に不可欠である
【0190】
Blimpgfp/+マウスからのリンパ器官の検査は、形質細胞において高レベルのGFPを示した。さらに、T細胞集団は、低レベルのGFPを発現した(図19A)。機能的Blimp対立遺伝子を発現しない、Blimpgfp/gfp細胞で再構成されたRag1-/-マウスのさらなる分析は、この同じ集団の顕著な増幅およびGFP蛍光の有意な増加を示した(図20)。これらのマウスのT細胞区画のより広範なフローサイトメトリー分析は、胸腺細胞(データは示していない)およびナイーブT細胞にはGFP蛍光がないことを示した(図20)。
【0191】
対照的に、および本明細書において示すように、Blimp-1は、活性化/記憶タイプCD4+(TCRβ+CD62Llow)およびCD8+(TCRβ+CD44high)T細胞において有意に発現される(図20)。より詳細なFACS分析では、代替マーカーを使用して、Blimp-1欠失マウスにおいて、増幅されたT細胞プールの性質を判定した。野生型胎仔肝臓で再構成されたマウスとは対照的に、Blimpgfp/gfp細胞で再構成されたRag1-/-マウスは、低CD62L、GFP陽性のT細胞集団の強い増幅を示した。さらに、これらのマウスは、低CD27〜CD27陰性のT細胞の数の強い上昇、CD25陽性CD4細胞の増加ならびに明瞭な高CD122集団の欠如を有した。代替マーカー、例えばCD69を使用する分析により、GFP+細胞の活性化状態を確認した(データは示していない)。Blimpgfp/gfpで再構成されたRag1-/-マウスにおけるエフェクターCD4 T細胞数の増加を、単離脾臓CD4 T細胞のエクスビボ刺激により確認し、結果として、8〜10倍高いIFN分泌が生じたが、IL-10の分泌は減少した。
【0192】
抗CD3/CD28およびIL-2(CD8+細胞について)またはTh1/Th2分極条件(CD4+細胞について)の存在下でのナイーブT細胞のインビトロ培養は、一次刺激培養物において低レベルのGFPを示した。しかし、再び刺激すると、Blimpgfp/+およびBlimpgfp/gfpマウスからのCD4およびCD8 T細胞は、迅速にGFP+になった(図19B)。重要なことに、野生型CD4+エフェクターT細胞におけるBlimp-1発現が、ウエスタンブロッティングにより確認された(図19C)。これらのデータは、Blimp-1発現の誘導が、ナイーブCD4+およびCD8+のエフェクター細胞への成熟の一成分であることを証明している。
【0193】
特異的抗原刺激後のBlimp-1発現をさらに評価するために、感染性疾患モデルを使用した。マウスを単純疱疹ウイルス(HSV)に感染させ、感染後、様々な時点で、優性エピトープに特異的な四量体および標準的な細胞毒性機能アッセイを使用してウイルス特異的CD8+ T細胞の一次および記憶応答についてモニターした。
【0194】
これらの実験により抗原特異的T細胞におけるBlimp-1の発現が確認された。また、これらの実験は、Blimpgfp/gfp細胞が、抗原特異的標的細胞を溶解する能力に関して十分機能的であることを示している。
【0195】
Blimpgfp/gfpで再構成されたマウスにおいて観察された活性化T細胞の増幅プールは、Blimp-1不在下での異常応答性または脱調節恒常性を示唆していた。この結論に合わせて、Blimpgfp/gfpで再構成されたマウスは、顕著な体重減少、外皮の皺および下痢を示し、再構成後6週間目から犠牲にしなければならなかった。これらのマウスの組織分析は、広範なリンパ球浸潤、ならびに肺、肝臓および胃腸管を含む様々な器官の炎症を示した(図21A)。組織損傷は、肺および腸において最も顕著であり、このプロセスが、これらのマウスにおいて観察された体重減少、下痢および死亡に関連していた。再構成されたマウスのリンパ器官および肝臓のフローサイトメトリー分析は、全ての器官における完全に活性化したCD4+およびCD8+ T細胞の大きな増幅を示した(図2B)。CD4+細胞は、主としてTh1に偏っていた。さらにBlimpgfp/gfp Rag2-/-胎仔肝臓を有するRag1-/-マウスは、結果として一切死なず、観察された病状にはT細胞が強く関与していた(図21)。
【0196】
無制御T細胞増幅が、T細胞に固有のものであるのかを判定するために、選別されたT細胞のRag1-/-への養子移入実験を行った。これらのアッセイにおいて、野生型T細胞は、恒常性増幅を制限されることによるリンパ球減少性環境に応答する(図22A)。T細胞、すなわち3×106個のCD4+またはCD8+ C57BL/6またはBlimpgfp/gfp、をRag1-/-受容個体に注射し、移入の3週間後に脾臓T細胞の数を評価した。図22Aに示すように、両方の系列のBlimpgfp/gfp T細胞が、劇的に増幅された恒常性増幅能力を有した。さらに、結果として生じたT細胞全てが、GFPを発現し、野生型対照物は、活性化された表現型を提示した(図22)。Blimpgfp/gfp T細胞を受けた受容個体は、図21に記載されているような、体重減少、巨脾腫(図22)および類似した範囲のリンパ球浸潤表現型を急速に発現した。これらのデータは、Blimp-1欠失T細胞が、多臓器浸潤および死を結果として生じさせる異常調節増幅を提示することを実証している。
【0197】
自己免疫性は、機能的CD4+CD25+サプレッサーT細胞の喪失に関連付けられている。Blimp+/gfpおよびBlimpgfp/gfp CD4+ T細胞におけるこの集団の分析は、CD25+ CD4 T細胞におけるGFPの中等度〜強度の発現を示した。これは、調節T細胞集団におけるBlimp-1についての機能を示唆している。インビトロおよびインビボでの機能分析により、Tregについての重要な因子FoxP3、およびTregに関連した他の遺伝子の不変の発現が観察された。Blimp欠失細胞CD4細胞は、高レベルのIFNを分泌し、IL-10分泌不良を示す。
【0198】
インビボでBlimp-1欠失T細胞において見られる異常調節恒常性が、それ自体を強化増幅としてインビトロでも表すかを判定するために、培養ナイーブCD8+ T細胞を抗CD3/CD28ならびに細胞増殖および恒常性を調節することが公知であるサイトカイン(IL-2、IL-15およびIL-21を含む)の存在下でインキュベートした。上の条件下で7日間成長させたナイーブCD8+細胞は、GFP発現、および遺伝子型の間で類似した増殖プロフィールをほとんど示さなかった(図23A)。ナイーブCD4+細胞、およびPMA/イオノマイシンの組み合わせにより刺激したものについて、同様の結果が観察された(データは示していない)。対照的に、IL-2、IL-15またはIL-21の存在下でのCD8+細胞の二次刺激は、結果として、Blimp-1発現の強い誘導を生じさせた(図23A)。この発現プロフィールと一致して、Blimp-1が十分な細胞と比較して、蓄積細胞数の有意な増加が、Blimpgfp/gfp細胞の増殖応答において観察された。予測されたとおり、活性化/記憶CD44high細胞を一次応答のために使用したとき、増殖能に同様の強い差異が観察された。この増加は、全てのサイトカイン条件下で観察されたが、IL-21の存在下で最も顕著であった(図23B)。IL-21は、NODマウスにおける糖尿病感受性遺伝子候補であることが最近明らかになった(King, et al. Cell 117:265-277, 2004)Tヘルパーサイトカインである。このモデルでは、IL-21が、エフェクターT細胞の代謝回転を増加させ、その結果としてリンパ球減少誘導恒常性増殖および糖尿病が生じた(King, et al., 2004 (前記))。興味深いことに、IL-21は、B細胞最終分化におけるBlimp-1の強力な刺激因子であり、またCD8+ T細胞におけるGFPの最も効率的な誘導因子であった。このことは、リンパ球分化におけるこのサイトカインの共通の役割を示唆している。
【0199】
分子およびインビトロ試験によって、Blimp-1は、ヒストンメチルトランスフェラーゼ(Gyory et al., Nat Immunol 5: 299-308, 2004)、デアセチラーゼ(Yu et al., Mol Cell Biol 20:2592-3603, 2000)および標的をサイレンシングさせるためのGrouchoファミリーのコリプレッサー(Ren et al., Genes Dev 13:125-137, 1999)を補充することができる強力な転写リプレッサーであることが示された。プロモータおよびマイクロアレイの研究により、B細胞系列において多数のBlimp-1標的が同定された(Shaffer et al, Immunity 17:51-62, 2002; Shaffer et al., Immunity 21:81-93, 2004)。これらのうち、c-myc(Lin et al., Science 276:596, 1997)、CIITA(Piskurich et al., Nat Immunol 1:526-532, 2000)を含む多数が、T細胞系列で発現される。
【0200】
実施例11
Blimpは、全てのリンパ球における保存的な最終分化プログラムの主調節因子である
要約すると、本明細書において記載のデータは、Blimp-1が、活性化された従来のT細胞において様々な状況下で発現されることを実証している。Blimp-1欠失T細胞を注射した、または突然変異幹細胞で再構成されたマウスが、進行性多器官リンパ球増殖性疾患の結果として死亡するので、Blimp発現は、正常なリンパ球恒常性には不可欠である。加えて、T細胞分化の恒常性を調節することが公知であるIL-21などのサイトカインは、Blimp-1発現の強力な誘導因子であり、Blimp-1不在下、インビトロで強化増殖を支持した。従って、Blimp-1発現は、初期刺激によってではなく、免疫応答の完了に向けて、エフェクターT細胞の分化を誘導し得る。従って、Blimp-1は、T細胞収縮の遺伝プログラムおよび/または免疫恒常性に不可欠である記憶形成を調節するように機能する、同定された最初の転写因子である。
【0201】
BおよびTリンパ球は、それらの発達中に多数の細胞的および遺伝子的類似性、例えば、共通の前駆細胞、定序逐次VDJ組換えおよび類似した発達チェックポイントを共有するが、形質細胞へのB細胞の最終分化が、明確な機能的終点である(Calame et al., Annu Rev Immunol 8:8, 2003)一方で、T細胞発生の最終段階は、ほとんど定義されていない。BおよびT細胞系列内のBlimp-1の類似した機能および発現プロフィールは、その外見的に異なる外観にもかかわらず、Blimp-1が、全てのリンパ球における保存的最終分化プログラムの主調節因子であるという興味深い可能性を提起する。
【0202】
Blimp欠失は、過形成および無制御増殖の原因となり、その一方で、Blimpの発現が、リンパ球恒常性ならびにASC、T細胞およびB細胞を含む造血細胞の最終分化を可能にする。従って、サイトカインおよび他の免疫調節成分、化学物質、ペプチドまたは小もしくは中分子剤は、本明細書において記載のインビトロおよびインビボ細胞モデル系においてスクリーニングして、治療薬または予防薬としての可能性を判定することができる。
【0203】
従って、本モデルレポーター系は、T細胞およびB細胞などの免疫系の細胞における最終分化を調節する薬剤の能力を評価する際に有用である。
【0204】
本明細書において記載の本発明には、明記している以外の様々な変化および変更の余地があることは、当業者には理解されるだろう。本発明が、全てのそのような変化および変更を包含することは、理解されるはずである。本発明は、個々にまたは集合的に、本明細書に引用している、または本明細書において指摘している段階、特徴、組成物および化合物の全て、ならびに段階または特徴のいずれか2つ以上の任意のおよび全ての組み合わせも包含する。
【0205】
参考文献一覧
【技術分野】
【0001】
発明の背景
発明の分野
本発明は、一般に、特定の系列の造血細胞およびそれらの分化段階を同定するモデルシステムに関する。さらに詳細には、本発明は、一般に、同定可能なシグナルを惹起することができるレポーター分子を共産生する(co-produce)ように改変された最終分化の遺伝子マーカーを有する、遺伝子改変細胞およびそのような細胞を含む非ヒト動物、ならびに、限定されるわけではないが、発達中の胚細胞、異常分化を伴う細胞(例えば、癌細胞)および造血細胞系列の細胞(例えば、Bおよび/またはT細胞など)などの細胞の分化またはトランスフォーメーション状態を調節することができる分子を同定する際のそれらの使用を提供する。同定された分子は、治療および予防用の薬学的組成物の基礎となる。
【背景技術】
【0002】
先行技術の説明
本明細書において参照する文献の詳細は、本明細書の末尾にも記載する。
【0003】
本明細書中のいずれの先行技術への参照も、この先行技術が、任意の国における共通一般知識の一部をなすことの承認またはいずれの形態の提案とも解釈されず、また解釈すべきでない。
【0004】
細胞寿命は、細胞分裂、分化、形態発生およびアポトーシスの変更を指示する無数の選択肢としての、および高度に調節された生化学経路を含む。分裂および/または分化する細胞の潜在能力は、多種多様である。例えば、胚は、任意の細胞タイプに分化する能力を保持する全能細胞を含む。幹細胞を含む他の細胞タイプは、多能性であり、全てではないが一定範囲の細胞表現型に最終的に分化することができる。一つの最終形態に決定付けられることとなる細胞もあり、これらは、最終分化する。
【0005】
最終分化細胞への細胞の正常な突然変異を阻害するまたはアポトーシスを防止する変化は、分化または細胞死を伴わない無制御細胞分裂を特徴とする腫瘍発現の引き金の役割を果たす。従って、分化および正常なアポトーシスを促進する薬剤により、腫瘍発現のスイッチを切ることができる。
【0006】
特定の細胞タイプの最終分化期間の間に発現される分子は、熱心に研究されてきた。しかし、分子レベルでのこの期間の間の事象の順序を理解するには、この発達段階に発現される分子の時間的および空間的発現パターンを理解する必要がある。
【0007】
Bリンパ球誘導成熟因子(Blimp)は、98kDa転写因子であり、これは、当初、B細胞リンパ腫細胞系の分化の間に誘導される因子として同定された(Turner et al., Cell 77: 297,1994(非特許文献1))。ヒト細胞由来の対応する因子は、PRDM-1と呼ばれる。Blimp-1は、B細胞最終分化の調節に顕著な役割を有することが報告されている。具体的には、Blimp-1は、ヒトおよびマウスからの抗体分泌細胞(ASC)において発現されるが、記憶細胞においては発現されない(Angelin-Duclos et al., J Immunol 165 : 5462, 2000(非特許文献2))。Blimp-1の異所発現は、リンパ腫および初代B細胞をASC細胞に最終分化させるために十分である(Turner et al.,(前記)(非特許文献1), Schliephake et al., Eur J Immunol 26 : 268, 1996(非特許文献3); Messika et al., J Exp Med 188 : 515, 1998(非特許文献4) ; Knodel et al., Eur J Immunol 31:1972, 2001(非特許文献5))。アンチセンスまたは優性干渉アプローチによるBlimp-1の発現の阻害により、完全ASC分化には不可欠であると考えられる細胞周期退出が抑制される(Soro et al., J Immunol 163 : 611, 1999(非特許文献6); Angelin-Duclos et al., J Immunol 165:5462, 2000(非特許文献7); Johnson et al., Eur J Immunol 32:3765, 2002(非特許文献8))。また、B細胞にBlimp-1がないマウスは、免疫グロブリンをほとんど産生せず、著しく低減されたASC区画を有する。(Shapiro-Shelef et al., Immunity 19 :607, 2003.(非特許文献9))。
【0008】
Blimp-1は、B細胞系列の細胞でしか産生されないと当初報告されたが、Blimp-1が骨髄系分化の間にも産生されることは、今では明らかである(Keller et al., Genes Dev 5:868, 1991(非特許文献10), Chang et al., Nat Immunol 1: 169, 2000(非特許文献11))。Blimp-1は、骨髄分化に関与するc-mycの抑制に必要とされる(Chang et al.,(前記), 2000(非特許文献11); Marcu et al., Annu Rev Biochem 61:809, 1992(非特許文献12))。U937細胞におけるBlimp-1の過剰産生は、例えば、マクロファージ分化の誘導に十分である(Chang et al., (前記), 2000(非特許文献11))。従って、マクロファージおよびB細胞におけるBlimp-1によるc-mycの抑制が、これら二系列における最終分化の特徴である。Blimp-1は、マウスおよびアフリカツメガエルの胚発達の間にも広範に産生される(de Souza et al., Embo J 18:6062, 1999(非特許文献13); Rosenbaum et al., Embo J, 9:897, 1990(非特許文献14))。
【0009】
Bリンパ球は、中でも最も集中的に研究された胚細胞型であるが、B細胞発達の初期段階は、比較的よく特性付けされている一方で、ASCへのBリンパ球の最終分化を制御するプロセスについてはほとんど判っていない。ASC(形質細胞)は、体液免疫応答の直接的伝達因子である。これらは、防御免疫に不可欠な血清免疫グロブリンを大量に分泌する。従って、ASCへのBリンパ球の最終分化は、強い治療的関心の対象となっている。例えば、ASCへの最終分化は、有効なワクチン接種戦略における非常に重要な要素である。さらに、多発性骨髄腫は、ASCの分化経路完了の失敗により生じる。
【0010】
しかし、ASCは、主として骨髄および脾臓にある非常に限定された細胞の非常に珍しい集団の代表である。マウスおよびヒトにおけるASC集団は、不均一な寿命および細胞表面表現型の細胞を含み、そのため、純粋なASCの決定的で見込みのある単離を行うことができない(Fong et al., Proc Natl Acad Sci U S A 11 : 11, 2003(非特許文献15); Medina et al., Blood 99. 2154, 2002(非特許文献16); O'Connor et al., J.Exp Med 195:737, 2002(非特許文献17); Manz et al., Curr Opin Immunol 14 : 517, 2002(非特許文献18); Underhill et al., Blood 24:24, 2003(非特許文献19))。
【0011】
T細胞最終分化プログラムは、ほとんど理解されていない(Sprent et al., Immunol Lett 85 : 145-149, 2003(非特許文献20))。感染に反応して、抗原特異的T細胞は、エフェクター細胞に分化し、大規模にクローン増幅される。T細胞数の恒常性は、その後の収縮段階により維持され、この段階では、エフェクター細胞の90%より多くが排除され、小率が記憶T細胞になる(Sprent et al., Annu Rev Immunol 20 : 551-579,2002(非特許文献21))。収縮は、感染の量または継続段階間に依存するので、このプロセスは、遺伝的制御下にあると提案された(Badovinac et al., Nat Immunol 5 : 809-817, 2004(非特許文献22); Badovinac et al., Nat Immunol 3: 619-626, 2002(非特許文献23))。T調節細胞の欠如のため(Khattri et al., Nat Immunol 4: 337-342,2003(非特許文献24); Hori et al., Science 299 : 1057-1061, 2003(非特許文献25); Fontenot et al., Nat Immunol 4 : 330-336,2003(非特許文献26))、下方調節性分子CTLA-4の喪失のため(Chambers et al., Immunity 7 : 885-895, 1997(非特許文献27))または自己免疫を生じさせる非肥満糖尿病(NOD)マウスでは遺伝子欠失のために増幅が増進した場合、T細胞数を制御する能力は不可欠である。
【0012】
広範な状況下および様々な刺激下でASC、T細胞および造血系の他の細胞の最終分化をモニタリングすることができることは、異常分化を随伴する一定範囲の状態(例えば、癌、自己免疫疾患)の治療および/または予防に使用するための戦略および試薬、または適切な免疫応答の発生の場合など正常な発生プログラムを利用する戦略および試薬を開発する際に極めて価値があるだろう。
【先行技術文献】
【非特許文献】
【0013】
【非特許文献1】Turner et al., Cell 77: 297,1994
【非特許文献2】Angelin-Duclos et al., J Immunol 165 : 5462, 2000
【非特許文献3】Schliephake et al., Eur J Immunol 26 : 268, 1996
【非特許文献4】Messika et al., J Exp Med 188 : 515, 1998
【非特許文献5】Knodel et al., Eur J Immunol 31:1972, 2001
【非特許文献6】Soro et al.,J Immunol 163 : 611, 1999
【非特許文献7】Angelin-Duclos et al., J Immunol 165:5462, 2000
【非特許文献8】Johnson et al., Eur J Immunol 32:3765, 2002
【非特許文献9】Shapiro-Shelef et al., Immunity 19 :607, 2003.
【非特許文献10】Keller et al., Genes Dev 5:868, 1991
【非特許文献11】Chang et al., Nat Immunol 1: 169, 2000
【非特許文献12】Marcu et al., Annu Rev Biochem 61:809, 1992
【非特許文献13】de Souza et al., Embo J 18:6062, 1999
【非特許文献14】Rosenbaum et al., Embo J, 9:897, 1990
【非特許文献15】Fong et al.,Proc Natl Acad Sci U S A 11 : 11, 2003
【非特許文献16】Medina et al., Blood 99. 2154, 2002
【非特許文献17】O'Connor et al.,J.Exp Med 195:737, 2002
【非特許文献18】Manz et al.,Curr Opin Immunol 14 : 517, 2002
【非特許文献19】Underhill et al., Blood 24:24, 2003
【非特許文献20】Sprent et al., Immunol Lett 85 : 145-149, 2003
【非特許文献21】Sprent et al., Annu Rev Immunol 20 : 551-579,2002
【非特許文献22】Badovinac et al., Nat Immunol 5 : 809-817, 2004
【非特許文献23】Badovinac et al., Nat Immunol 3: 619-626, 2002
【非特許文献24】Khattri et al., Nat Immunol 4: 337-342,2003
【非特許文献25】Hori et al., Science 299 : 1057-1061, 2003
【非特許文献26】Fontenot et al., Nat Immunol 4 : 330-336,2003
【非特許文献27】Chambers et al., Immunity 7 : 885-895,1997
【発明の概要】
【0014】
本明細書を通して、その文脈が別様に求めていない限り、語「含む(comprise)」、または「含む(comprises)」もしくは「含んでいる(comprising)」などの語尾変化したものは、示されている要素もしくは整数または要素もしくは整数の群の包含を意味するが、任意の他の要素もしくは整数または要素もしくは整数の群の排除は意味しないことは理解されるだろう。
【0015】
ヌクレオチドおよびアミノ酸配列は、配列識別子番号(配列番号:)によって参照する。配列番号:は、<400>1(配列番号:1)、<400>2(配列番号:2)など、配列識別子に数値対応している。配列識別子の一覧を表1に提供する。配列表は、添付されている。
【0016】
遺伝子および他の遺伝物質(例えば、mRNA、構築物など)は、イタリック体で表しており、それらのタンパク質性発現産物は、イタリック体でない形で表す。従って、転写因子Blimpは、Blimpの発現産物である。用語「Blimp」または「Blimp」は、ヒト相同体を含む任意の動物または哺乳類に由来する全ての相同体または変異体分子を示すために用いている。従って、ヒトPRDM-1およびその産物、PRDM-1は、用語BlimpまたはBlimpに包含される。別様に述べられていない限り、Blimpへの言及は、そのポリペプチドの機能的な形態への言及であり、改変Blimpへの言及は、Blimpの機能的な形態をコードする遺伝子または対立遺伝子配列への言及である。
【0017】
本発明は、造血および胚細胞の分化におけるBlimpの役割の同定に、一部、基づいている。Blimpの存在またはBlimpのレベルについてスクリーニングすることにより、細胞の最終分化の段階について判定を下すことができる。Blimpの役割の同定により、ASC(形質細胞)などであるが、これに限定されるわけではない、造血細胞の実質的に均質な集団を同定することがさらに可能となる。
【0018】
より詳細には、本発明は、Blimpおよびレポーター分子をコードする遺伝物質を共発現する細胞を含む、遺伝子改変細胞またはインビボもしくはインビトロ系を提供する。リンパ球系列などであるが、これに限定されるわけではない、造血系列の細胞におけるレポーター活性の検出は、レポーター活性を有するおよび機能的Blimpを産生する細胞が、最終分化に決定付けられていることの指標となる。従って、機能的Blimpを産生するレポーター活性B細胞の検出は、これらの細胞が、抗体分泌細胞(ASC)への分化に決定付けられていることの指標となる。また、本明細書において記載のように、機能的Blimpを発現するT細胞におけるレポーター活性の検出は、これらの細胞が、活性化CD4+ T細胞またはエフェクターCD8+ T細胞などの活性化/記憶T細胞であることの指標となる。従って、本発明は、様々な条件下または様々な刺激もしくは薬剤の存在下での特定の細胞の分化またはトランスフォーメーション状態のモニタリングを助長する、遺伝子改変細胞またはそのような細胞を含む非ヒト動物を提供する。本発明は、発達中の幹細胞を含む胚細胞、異常分化を伴う細胞(例えば、癌細胞)、および、例えばBおよび/またはT細胞など造血細胞系列の細胞であるが、これらに限定されない、細胞の分化またはトランスフォーメーション状態を調節することができる分子を同定するための、ハイスループットスクリーニング法を含むスクリーニング法をさらに提供する。
【0019】
具体的には、遺伝子改変細胞またはそのような細胞を含む非ヒト生物を本発明によって提供する。一つの態様において、本細胞は、レポーター分子をコードするように改変されたmRNAから翻訳されたBlimpを産生する。好ましくは、そのレポーター分子コード配列が、Blimp対立遺伝子のイントロンに挿入される。この改変Blimp対立遺伝子が、ヘテロ接合体の形態で存在するとき、他方の対立遺伝子は、機能的Blimpを発現する。一部の態様において、改変対立遺伝子は、機能的Blimpポリペプチドまたはその機能的な形態を発現し得る。他の態様において、改変対立遺伝子は、非機能的Blimpポリペプチドを発現した。一つの態様において、改変細胞は、なかでもASCを同定および単離するのためのインビトロまたはインビボ細胞モデル系において有用である。もう一つの態様において、該改変細胞は、広範なアッセイにおけるT細胞および/またはB細胞などの造血細胞の分化状態のモニタリングに有用である。
【0020】
一つの局面において、本発明は、発現されるとき、レポーター分子と共発現される、ポリペプチドを産生する改変遺伝子を含み、ならびにポリペプチドが造血細胞の最終分化に関連している、遺伝子改変細胞またはそのような細胞を含む非ヒト生物を提供する。好ましくは、この遺伝物質は、Blimp遺伝子、またはその一部、フラグメント、相同体、誘導体もしくは機能的な形態である。さらに、B細胞系列細胞におけるレポーター分子の同定は、そのような細胞が、分化に決定付けられているか、ASCに分化したことの指標となる。または、T細胞系列の細胞におけるレポーター分子の活性は、これらの細胞が活性化されていることの指標となる。従って、本明細書において記載のように、リンパ球におけるBlimpの存在は、その細胞が最終分化するか、最終分化に決定付けられていることの指標となる。例示的なT細胞としては、CD4+ T細胞およびCD8+ T細胞が挙げられ、例示的なB細胞は、ASCである。非機能的Blimpポリペプチドが産生される場合、レポーター活性細胞の検出は、それらの細胞が、それらが機能的Blimpポリペプチドを産生することができた場合にそれらを最終分化させるために十分な条件に暴露されていることを示す。
【0021】
遺伝子改変非ヒト生物は、移植のために生殖体、胚またはES細胞の形態で提供することができる。胚は、好ましくは、凍結状態で維持され、任意で使用説明書と共に販売される。ターゲティング構築物および遺伝子改変細胞は、好ましくは凍結状態でも維持され、任意で使用説明書と共に販売される。本明細書においては、全てのそのような細胞をインビトロまたはインビボ細胞モデル系と呼ぶ。
【0022】
本発明は、単細胞、組織および生体レベルで、インビボおよびインビトロで、細胞における遺伝子発現および分化の運命をモニターするシステムを提供する。従って、レポーター活性は、生細胞でモニターすることができ、遺伝子発現は、固定された組織においてモニターすることができる。好ましくは、このレポーター発現カセットは、蛍光または他の発光部分をコードする。例えば特定の系列または細胞の最終分化についてのマーカーとしてBlimp-1の発現をレポートする生物および細胞が利用できることは、広範な用途において極めて有用なツールとなる。B細胞系列の細胞に関して、この系は、ASCの研究、単離およびモニタリングに幅広く使用することができる。前に述べたように、ASCは、これらの細胞が有効な抗体応答に非常に重要であるとはいえ、これまで研究に利用することができなかった。さらに、ASCにおける異常分化は、ヒトでは多発性骨髄腫の原因となり、その結果、ASCは、この理由についての研究のための重要な細胞タイプとなっている。
【0023】
関連態様において、本発明は、造血系の細胞を表現型解析および/またはモニターする方法を提供し、この方法は、発現されるとき、Blimpあるいはその一部、フラグメント、変異体、相同体または機能的もしくは非機能的な形態およびレポーター分子を共発現する、Blimpタンパク質をコードする改変Blimp遺伝子を含む、遺伝子改変細胞またはそのような細胞を含む非ヒト動物をスクリーニングする段階を含み、レポーター活性の検出は、細胞表現型および/または最終分化への細胞の決定付け(commitment)の指標または予測となる。造血細胞としては、B細胞、T細胞、樹状細胞、マクロファージ、ナチュラルキラー細胞、顆粒球、赤血球、好酸球、巨核球、骨髄細胞、脾細胞、皮膚細胞もしくは間質細胞またはこれらの誘導体が挙げられるが、それらに限定されるわけではない。一つの特定の態様において、造血細胞は、Bおよび/またはT細胞などのリンパ球である。
【0024】
さらなる態様において、レポーター活性またはレポーター活性の変化を示す細胞が、レポーター活性を示さない細胞の中から単離または選択される。レポーター活性細胞の単離は、任意の便利な方法によるものであり得る。例えば、フローサイトメトリー、レーザー走査サイトメトリー、クロマトグラフィーおよび/または他の同等の手法が便利に利用される。フローサイトメトリー法が特に好ましい。加えて、例えば薬物選択マーカーなどのさらなる選択マーカーを使用して、本発明の改変細胞を単離または選択することができる。
【0025】
本発明は、Blimp-1発現またはBlimp-1活性の拮抗薬(antagonist)および作動薬(agonist)も提供する。Blimp-1発現の作動薬の一例は、IL-21などであるが、これに限定されるわけではない、サイトカインである。本作動薬および拮抗薬の組換え、合成または単離形ならびに1つまたは複数の薬学的に許容される担体、希釈剤または賦形剤を含む薬学的組成物が、さらに考えられる。Blimp-1発現またはBlimp-1タンパク質の産生への言及は、単細胞におけるものまたは細胞集団内でのものを含む。
【0026】
(表1)配列識別子の一覧
【図面の簡単な説明】
【0027】
【図1】Blimp-1遺伝子座およびターゲティング戦略の図である。A)Blimp-1タンパク質の構造ドメイン。エキソン7〜8によりコードされているタンパク質のセグメントを示す。Acidic、NおよびC末端酸性領域;PR、網膜芽腫相互作用性ジンクフィンガータンパク質RIZに対するホモロジー領域;Pro、プロリン・リッチ領域;Zn、5つのジンクフィンガー。B)Blimp-1のゲノム遺伝子座、8つのエキソンをボックスとして示し、イントロンを黒線として示す。コード領域は、灰色であり、非翻訳領域は、白色である。5’および3’プローブとともに、サザン・ハイブリダイゼーションに使用した制限酵素に印を付けてある。相同組換え事象およびその後の操作から誘導された標的化対立遺伝子を示す。C)ターゲティング事象の期待産物を明らかにするためにBlimp-1遺伝子座の5’および3’末端を使用する標的化および対照ES細胞DNAに対するサザン・ハイブリダイゼーション(4.8kb 5’アームおよび4.5kb 3’アーム)。IL-15+/-IL-21においてエクスビボで0〜3日間培養したblimpgfp/+LPS刺激B細胞におけるBlimp-1の発現。Blimp-1発現は、マウスBlimp-1に対するモノクローナル抗体を使用して検出し、α-アクチンに対するヤギポリクローナル抗体を負荷対照として使用した。+/+、野生型C57B1/6マウス;+/T blimpgfp/+マウス。
【図2】インビボでのB細胞におけるBlimpgfp発現のFACS分析の結果を示すグラフ図である。A)Blimpgfp/+マウス(上のパネル)および対照(下のパネル)におけるリンパ節、脾臓および骨髄でのシンデカン-1およびBlimpgfpの発現。B)B220陽性B細胞におけるBlimpgfpの発現。
【図3】Blimpgfp選別細胞のELIspot分析の結果を示すグラブ図および写真図である。Gfp陽性細胞を未治療Blimpgfp/+マウスの骨髄(BM)および脾臓から選別し、Elispotアッセイにおいて分析した。アイソタイプ特異的抗体または抗κ抗体を使用して、Elispotプレートを被覆し、分泌された免疫グロブリンを検出した。A)200個のgfp陽性選別細胞におけるアイソタイプ特異的免疫グロブリンの分布(3回の実験のうちの1回の代表実験)。B)ELIspotプレートの単一の代表ウエルにおけるκ鎖の検出(サンプル:選別された骨髄細胞)。左、200個のgfp陽性細胞をインプット;中央、100000個のgfp陰性細胞をインプット;右、100000個の未選別細胞をインプット。
【図4A】インビボでのLPSでの抗体分泌性細胞の誘導を示すFACS分析の結果のグラフ図である。Blimpgfp/+マウスに2ugの大腸菌(E.coli)LPSを静脈内注射した。これらのマウスの脾臓を、LPS治療後の示されている時点で分析した。LPSは、ASCの形成を誘導し、その結果、脾臓において3日目には約0.5%から約5%に頻度が増加する。上のパネル、シンデカン-1およびBlimpgfpについてのFACSスキャン。中央のパネル、gfp陽性ゲート細胞内のシンデカン-1およびB220。下のパネル、示されている時点でのGFP陽性細胞におけるシンデカン-1およびB220発現についてのヒストグラム。
【図4B】インビボでのLPSでの抗体分泌性細胞の誘導を示すFACS分析の結果のグラフ図である。Blimpgfp/+マウスに2ugの大腸菌(E.coli)LPSを静脈内注射した。これらのマウスの骨髄を、LPS治療後の示されている時点で分析した。LPSは、ASCの形成を誘導し、その結果、骨髄において4日目には約0.05%から約0.25%に頻度が増加する。上のパネル、シンデカン-1およびBlimpgfpについてのFACSスキャン。中央のパネル、gfp陽性ゲート細胞内のシンデカン-1およびB220。下のパネル、示されている時点でのGFP陽性細胞におけるシンデカン-1およびB220発現についてのヒストグラム。
【図5】図5Aは、Blimpgfp発現の動態のグラフ図である。Blimpgfp/+マウス(赤線)および野生型C57B1/6マウス(青線)からの刺激後B細胞によるBlimpgfp発現のフローサイトメトリーヒストグラムを示す。ヒストグラムゲートは、Blimpgfp陽性集団のパーセンテージを示す。高度に精製された小さな休止B細胞を、組換えCD40L、IL-4およびIL-5(最上パネル)またはLPS(20ug/mL)(最下パネル)で刺激した。異なる培養日数で細胞を回収し、フローサイトメトリーで分析した。LPSで刺激した細胞は2日目にBlimpの発現を開始し、一方、CD40LおよびIL-4/IL-5に応じてBlimp発現が3日目に顕性になった。図5Bは、Blimpgfp陽性細胞が抗体を分泌することを示すグラフ図である。Blimpgfp/+B細胞をLPSで4日間刺激した。細胞を回収し、シンデカン-1(Synd-1)特異的抗体で染色し、自動細胞堆積ユニットを使用して、GFP発現領域(左のパネル、A〜C)およびGFP非発現領域(左のパネル、D)を、様々なアイソタイプ特異的抗体で被覆されたElispotプレートに直接分別した。分別された細胞を標準的なElispot法に従って処理した。右のパネルは、選別された領域におけるIg分泌性細胞の数を示す。大部分のBlimpgfp細胞は、Igを分泌するが、全てのBlimpgfp陰性細胞が、検査したいずれのIgアイソタイプも分泌しない。図5Cは、様々な刺激に応答しての種々のBlimpgfp発現を示すグラフ図である。高度に精製された小さな休止B細胞を、i)組換えCD40LおよびIL-4;ii)CD40L、IL-4およびIL-5;iii)LPS;iv)LPSおよびIL-4;v)LPSおよび抗IgDモノクローナル抗体で刺激した。4日培養した後、細胞を回収し、Synd-1特異的抗体で染色し、フローサイトメトリーで分析した。フローサイトメトリー分析の2パラメータープロットをここに示す。
【図6】活性化B細胞を移植したマウスの分析結果を示すグラフ図である。Blimpgfp/+マウスの精製された休止脾臓B細胞を20ug/mL LPSの存在下で3日間活性化した。3×106の細胞(約2×106のgfp陽性細胞を含有、すなわち、抗体分泌性細胞、A)をLPSで3回洗浄し、WT受容個体に静脈内注射により移植した。7日後、それらの受容マウスをドナーASCの存在について分析した(B)。
【図7】図7Aは、Blimpgfp/+×blimpgfp/+交雑からのマウス骨の遺伝子型解析結果を表にまとめたものである。図7Bは、離乳したマウス(左)またはE9.5日目の胚(右)の遺伝子型解析の代表的PCR結果の写真図である。
【図8】20ug/mLのLPSの存在下で培養し、3日目にGFP陽性、すなわち抗体分泌性細胞、の存在について分析した、Blimpgfp/gftおよびBlimpgfp/+再構成マウスの脾細胞の写真図およびグラフ図である(A)。その後、両方の培養物のGFP陽性細胞を選別し(B、ゲートR1)、ELIspotアッセイにおいて分析した。Blimpgfp/+細胞は、60〜70%の抗体分泌性細胞を生じ(B、下のパネル左)、Blimpgfp/gftは、5〜7%しか抗体分泌性細胞をもたらさず、ヘテロ接合細胞によって生じたスポットと比較して小さなELIspotしか生じなかった(B、下のパネル、右)。ELISPOTプレートの単一代表ウエルにおけるIgMおよびκ鎖の検出(200個のgfp陽性細胞をインプット)。
【図9】骨髄由来マクロファージ(BMM)および血液単球のFACS分析の結果のグラフ図である。骨髄細胞を10ng/mL rMCSFの存在下で7日間培養し、培地を交換し、非付着細胞を培養3日目および5日目に除去した。付着細胞(BMM)をBlimpgfp発現について分析した(左パネル)。さらに、MacI/Gr1二重陽性血球細胞をFACSで分析した(右パネル)(黒線-野生型、赤線-Blimpgfp/+)。
【図10】インビトロで産生させた樹状細胞(DC)のFACS分析を示すグラフ図である。骨髄細胞を100ng/mL Flt3リガンドの存在下で8日間培養した。その後、細胞を、さらに24時間培養(左カラム)、またはCpG(1.5uM)、GMCSF(50ng/mL)、gIFN(20ng/mL)およびIL-4(20ng/mL)で刺激(中央カラム)もしくは1ug/mLのLPSで刺激(右カラム)した。形質細胞性DCおよび従来型DCについてのBlimpgfp発現をヒストグラムで示す(実線-野生型、点線-Blimpgfp/+)。
【図11】インビボおよびインビトロでのT細胞のFACS分析を示すグラフ図である。Blimpgfp/+マウスの胸腺T細胞(左)およびリンパ節T細胞(中央)およびインビトロ活性化CD4+/CD8+精製リンパ節細胞(右)をFACSにおいて分析した。ゲートされたT細胞集団のBlimpgfp発現レベルをヒストグラムで示す(下のパネル;黒線-野生型;赤線-Blimpgfp/+)。
【図12】Blimpgfp/+レポーターマウスにおいて検出することができ、成熟刺激により誘導することができるNK系列におけるBlimp-1発現を示すグラフ図である。A)インビボ脾臓NK細胞は、GFP+である。B)Blimpgfp/+脾臓から選別されたNK細胞を4日間、IL-15中で、その後、2日間、示されているサイトカイン中で培養した。mfiは、Blimpgfpの蛍光係数の平均である。C)IL-15+/-IL-21においてエクスビボで7日間培養した、+/+NK細胞におけるBlimp-1の発現。マウスBlimp-1に対するモノクローナル抗体を使用してBlimp-1発現を検出し、α-アクチンに対するヤギポリクローナル抗体を負荷対照として使用した。
【図13−1】マウスBlimp-1/PRDM-1のcDNAおよび予測アミノ酸配列を示す図である。コード配列を大文字で示す。
【図13−2】図13−1の続きを示す図である。
【図13−3】図13−2の続きを示す図である。
【図13−4】図13−3の続きを示す図である。
【図14】図13におけるヌクレオチド配列から誘導されたマウスBlimp-1/PRDM-1のアミノ酸配列(大文字)を示す図である。
【図15−1】ヒトBlimp-1/PRDM-1のcDNAおよび予測アミノ酸配列を示す図である。コード配列を大文字で示す。
【図15−2】図15−1の続きを示す図である。
【図16】図15におけるヌクレオチド配列から誘導されたヒトBlimp-1/PRDM-1のアミノ酸配列(大文字)を示す図である。
【図17−1】マウスBlimp-1のゲノムヌクレオチド配列を示す図である。このゲノム遺伝子座は、太字大文字の8エキソンを含む。ATGおよび停止コドンに下線を引いてある。
【図17−2】図17−1の続きを示す図である。
【図17−3】図17−2の続きを示す図である。
【図17−4】図17−3の続きを示す図である。
【図17−5】図17−4の続きを示す図である。
【図17−6】図17−5の続きを示す図である。
【図18−1】ヒトBlimp-1のゲノムヌクレオチド配列を示す図である。このゲノム遺伝子座は、大文字、太字の8エキソンを含む。ATGおよび停止コドンに下線を引いてある。
【図18−2】図18−1の続きを示す図である。
【図18−3】図18−2の続きを示す図である。
【図18−4】図18−3の続きを示す図である。
【図18−5】図18−4の続きを示す図である。
【図18−6】図18−5の続きを示す図である。
【図18−7】図18−6の続きを示す図である。
【図19A】Blimp-1が、活性化/記憶T細胞において発現されることを示すグラフ図である。Blimp-1欠失T細胞は、活性化/記憶表現型を有する。A)示されている表現型の、ゲートされたCD4+またはCD8+脾臓T細胞を、GFP蛍光および活性化状態について検査した。Blimpgfp/+マウス由来のCD62LlowCD4+ T細胞およびCD44highCD8+ T細胞の大部分は、GFPが低く、少数のCD4+ T細胞(低または高CD62L)のみが、高GFPである。Blimp GFPは、Blimpgfp.gfp T細胞の同じ集団内で強く発現される(ドットブロット);Blimp-1欠失CD4+およびCD8+脾臓T細胞の表現型(ヒストグラム、実線blimp-1+/+、点線blimp-1gfp/gfp)。
【図19B】Blimp-1が、活性化/記憶T細胞において発現されることを示すグラフ図である。Blimp-1欠失T細胞は、活性化/記憶表現型を有する。B)インビトロ培養によりBlimp-1発現が誘導される。ナイーブ(naive)CD4+ T細胞をTh1/Th2分極条件下で2ラウンド成長させた。
【図19C】Blimp-1が、活性化/記憶T細胞において発現されることを示すグラフ図である。Blimp-1欠失T細胞は、活性化/記憶表現型を有する。C)Th1またはTh2条件下で上と同じように成長させたCD4+細胞における野生型Blimp-1タンパク質についてのウエスタンブロット。抗Zap-70を負荷対照として使用した。形質細胞分化を誘導するようにLPSで4日間刺激したB細胞を陽性対照として使用した。
【図19D】Blimp-1が、活性化/記憶T細胞において発現されることを示すグラフ図である。Blimp-1欠失T細胞は、活性化/記憶表現型を有する。D)示されている遺伝子型で再構成されたマウスを、HSV感染後、gB特異的四量体を使用して優性gB498-505エピトープに特異的なCD8+ T細胞の出現について分析した。HSV感染データは、各々の遺伝子型の少なくとも3匹のマウスの代表である。
【図19E】Blimp-1が、活性化/記憶T細胞において発現されることを示すグラフ図である。Blimp-1欠失T細胞は、活性化/記憶表現型を有する。E)インビトロ培養gB特異的CTLは、正常な細胞毒性機能を示す。
【図20A】Blimp-1陽性CD4+ T細胞集団の分子分析の図である。A)Blimp-1は、CD25+サプレッサーT細胞において発現される。CD25+およびCD25-CD4+ T細胞を選別し(左のパネル)、ナイーブCD4+をTh1およびTh2分極条件下で分化させ(右のパネル)、全ての細胞集団をRT-PCR分析に供した。
【図20B】Blimp-1陽性CD4+ T細胞集団の分子分析の図である。B)Blimp-1欠失CD4+細胞は、大量のIFNgを分泌し、IL-10分泌欠損を示した。CD4+細胞は、脾臓からエクスビボで選別し、抗CD3およびCD28結合プレートで24時間再刺激したか、またはTh1もしくはTh2細胞にインビトロで分化させ、再刺激に供した。上清中のIL-10およびIFNgは、ELISAで検出した。
【図21】Blimp-1欠失マウスが、致死性リンパ球増殖亢進症候群を発現することを示す図である。A)対照細胞またはBlimpgfp/gfp胎児肝臓由来幹細胞で再構成したRag1-/-マウスの組織学的検査。Blimpgfp/gfp再構築マウスは、瀕死の場合、犠牲にした。Blimpgfp/+マウスからの正常な組織学的外観の器官を、Blimpgfp/gfpマウスで観察されたリンパ球浸潤と対比させる。B)Blimpgfp/+再構成マウスではなく、Blimpgfp/gfp再構成マウスにおける病的状態の急速な発症。検査した各遺伝子型の動物の数を示す。
【図22A】Blimp-1が、T細胞の恒常性増殖を調節することを示すデータのグラフ図である。A)示されている遺伝子型由来の3×106のナイーブCD4+またはCD8+脾臓T細胞を非放射線照射Rag2-/-受容個体に養子的に移入した。受容個体を体重減少および窮迫の徴候についてモニターした。グラフは、3〜4週間にわたる体重の変化率を示す。4〜6匹のマウスを各遺伝子型の細胞で再構成した。
【図22B】Blimp-1が、T細胞の恒常性増殖を調節することを示すデータのグラフ図である。B)体重が10%より多く減少したマウスを犠牲にし、脾臓T細胞数を判定した。
【図22C】Blimp-1が、T細胞の恒常性増殖を調節することを示すデータのグラフ図である。C)Blimpgfp/gfp CD8+ T細胞の移入後の代表Rag2-/-マウスの脾腫。
【図22D】Blimp-1が、T細胞の恒常性増殖を調節することを示すデータのグラフ図である。D)移入の3週間後のドナーT細胞におけるBlimpgfp発現のフローサイトメトリー分析。
【図23】Blimp-1が、二次刺激後に誘導され、サイトカイン応答性を調節することを示すデータの図である。A)示されている遺伝子型のナイーブCD8+ T細胞を抗CD3/CD28およびIL-2の存在下で5日間培養し、その後、5日間、IL-21を含有する二次培養物に再接種した。B)上と同じ細胞培養物を、示されているサイトカインの組み合わせと共にまたはそれらを伴わずに抗CD3/CD28で刺激した。一次または二次培養物において、示されている時点で、全生細胞数を判定した。CD44high活性化/記憶CD8+細胞は、一次培養のみに供した。
【発明を実施するための形態】
【0028】
好ましい態様の詳細な説明
本発明は、造血細胞系の細胞もしくは胚性細胞を同定および単離するための、および/または造血細胞もしくは胚性細胞の分化をモニターするための方法の開発に、一部基づいており、この方法は、(レポーターによる)ポリペプチドの存在の検出または定量を含み、その存在は、細胞の最終分化に関連している。
【0029】
特に好ましい態様において、ポリペプチドは、Blimpまたはその一部、フラグメントもしくは機能的な形態であり、これらは、レポーター分子と共発現される。
【0030】
従って、本発明の一つの局面は、発現されるとき、レポーター分子と共発現されるポリペプチドを産生する、ポリペプチドをコードする遺伝物質を含み、ポリペプチドが、細胞における最終分化への決定付けを含む細胞表現型に関連している、遺伝子改変細胞またはそのような細胞を含む非ヒト生物を提供する。
【0031】
さらなる局面において、本発明は、発現されるとき、レポーター分子と共発現されるBlimpまたはその一部、フラグメントもしくは機能的な形態を産生する、Blimpポリペプチドをコードする改変Blimp遺伝子を含み、Blimpの存在が、細胞における最終分化への決定付けを含む細胞表現型に関連している、遺伝子改変細胞またはそのような細胞を含む非ヒト生物を提供する。
【0032】
さらに好ましい局面において、本発明は、Blimpコード配列またはその一部、フラグメントもしくは機能的な形態およびレポーター分子コード配列を含む、Blimp mRNA転写産物をコードする改変Blimp遺伝子を含み、Blimpの存在が、細胞における最終分化への決定付けを含む細胞表現型に関連している、遺伝子改変細胞またはそのような細胞を含む非ヒト生物を提供する。
【0033】
好ましくは、レポーター分子コード配列は、Blimp対立遺伝子のイントロンに挿入される。このようにして、改変Blimp対立遺伝子は、内因性Blimp調節要素の制御下で2シストロンのRNAからレポーターを共産生する。
【0034】
用語「共発現」および「共産生」は、同時または実質的に同時の2つ以上の核酸領域の転写(1つまたは複数のRNAとして発現される)および同時または実質的に同時な、それらのその後の翻訳(1つまたは複数のポリペプチドとして産生される)を指すように本明細書においては広義に用いている。好ましくは、Blimpまたはその一部、フラグメントもしくは機能的な形態とレポーター分子の両方をコードする1つの転写産物が発現される。各場合、レポーターの発現は、レポートされる分子の発現に作動可能に連結している。
【0035】
本明細書において「細胞表現型」への言及は、細胞の分子特性または機能特性を包含する。例えば、ASC細胞は、Blimp-1(分子マーカー)を発現し、ならびになかでも、高いIg分泌率、MHCクラスII分子の不在および低い表面Igレベルを示すことにより、他のB細胞とは機能的に区別される。本明細書において用いる場合、この用語は、調節されたレベルのBlimp-1発現の分子特性に加えて、全範囲の分子もしくは機能特性、または任意の特定の分子もしくは機能特性への言及である。
【0036】
本遺伝子改変細胞またはそのような細胞を含む非ヒト生物は、ヒト、非ヒト霊長類、家畜、愛玩生物、実験検査生物、爬虫類または両生類などであるが、これらに限定されない、任意の生物由来の細胞または遺伝物質を含むことができる。好ましくは、本遺伝子改変生物は、マウスまたは他の実験検査動物、例えばラット、モルモット、ブタ、ウサギもしくはヒツジである。
【0037】
本明細書において用いる場合、単数形「ある(a、an)」および「その(the)」は、その文脈が明確に別様に指示していない限り、複数の局面を含む。従って、例えば、「細胞」への言及は、単細胞のみでなく2個以上の細胞を含み;「遺伝子」への言及は、1つの遺伝子のみでなく2つ以上の遺伝子を含むなどである。
【0038】
本発明の改変遺伝子は、B細胞系列細胞などの造血系の細胞における最終分化についてのマーカーである。
【0039】
「遺伝子改変細胞」への言及は、その改変前の形態を基準にして変更されているゲノムおよびその後代のコード領域または非コード領域からのヌクレオチドの配列を含むように操作された任意の細胞への言及である。特に、細胞は、最終分化の遺伝子マーカーおよびレポーター分子コード配列を共発現するように遺伝子改変される。好ましくは、細胞は、Blimpまたはその一部、フラグメントもしくは機能的な部分およびレポーター分子を共発現するように遺伝子改変される。レポーター分子は、同定可能なシグナルを直接または間接的に生じることができる任意の分子であり得る。蛍光または他の発光レポーター分子が、特に好ましい。
【0040】
便利には、最初にターゲティング構築物を用いて、細胞または生物体内で改変遺伝子配列を発生させる。一般に、しかし排他的でなく、ターゲティング構築物は、相同組換えにより標的配列を改変する。または、改変された遺伝子配列を人工染色体を使用して導入することができる。ターゲティングまたは他の構築物は、例えば、Sambrook, Molecular Cloning: A Laboratoy Manual, 3rd Edition, CSHLP, CSH, NY, 2001 ; Ausubel (Ed) Current Protocols in Molecular Biology, 5th Edition, John Wiley & Sons, Inc, NY, 2002などの、分子生物学実験室マニュアルに記載されている当技術分野において周知の方法を用いて、産生させ、標的細胞に導入することができる。ターゲティング構築物は、エレクトロポレーション、ウイルス媒介移入またはマイクロインジェクションなどの任意の方法により細胞に導入することができる。一般には選択マーカーを利用して、ターゲティング構築物が首尾よく組みこまれた細胞を最初に同定する。当業者には理解されるように、本改変生物は、一定の細胞においてのみBlimp対立遺伝子およびレポーター分子を発現するように遺伝子改変することができる。
【0041】
一つの特定の態様において、本発明は、ターゲティング構築物としての使用に適する核酸構築物を提供し、構築物は、Blimp-1の対立遺伝子の全てまたは一部分およびレポーター構築物を含む。構築物は、機能的に活性なBlimp-1ポリペプチドまたは機能的に不活性なBlimp-1ポリペプチドをコードする遺伝物質を含む。特定の態様において、構築物は、DNA結合モチーフを含むジンクフィンガードメインを欠く部分的Blimp-1ポリペプチドをコードする。特に好ましい態様において、構築物は、核酸構築物のリコンビナーゼ媒介欠失および標的遺伝子配列への核酸構築物の相同組換えを助長する部位に隣接している。または、構築物は、それがエピソーム的に複製する宿主細胞に導入することができる。
【0042】
遺伝子改変生物は、Hogan et al., Manipulating the Mouse Embryo : A Laboratory Manual, Cold Spring Harbour Laboratory Press, CSH NY, 1986 ; Mansour et al., Nature 336 :348-352,1988 ; Pickert, Transgenic Animal Technology: A Laboratory Handbook, Academic Press, San Diago, CA, 1994に記載されているような当技術分野において周知の技法を用いて産生させる。胚性幹細胞(ES細胞)を含む幹細胞を発生の胚盤胞期に受容生物の胚に導入する。そこではそれらを内細胞塊に組み込むことができ、この内細胞塊で、それらは、受容生物の生殖細胞系に発達し寄与する。ES細胞は、インビトロで維持されている移植前の胚から便利に得られる(Robertson et al., Nature 322 : 445-448, 1986)。正確なターゲティングが確認されたら、改変細胞を胚盤胞期または桑実期または他の適する発達段階に注入して、キメラ生物を発生させる。または改変細胞を解離胚細胞と凝集させて、凝集キメラを形成することができる。その後、そのキメラ生物を、適する雌里親生物に移植し、胚を出産日まで発達させることができる。キメラ後代を繁殖させて、ターゲティング構築物によりもたらされたヌクレオチド配列を各細胞のゲノムが含有する子孫を得る。遺伝子改変生物は、ヘテロ接合改変を含むことがあり、または両方の対立遺伝子が影響を受けることがある。
【0043】
本発明によると、驚くべきことに、Blimp-1は、ASC経路に沿って分化する決定付けではなく、ASCによる抗体の産生に不可欠であると判定される。従って、B細胞系列細胞におけるBlimpの(例えば、レポーター分子との共発現による)同定は、細胞が、ASCに分化するように決定付けられているか、ASCに分化したことを示す。
【0044】
さらに、本明細書において開示するように、Blimpは、T細胞恒常性を含むリンパ球恒常性および最終分化するようになるT細胞の能力に不可欠である。生体哺乳動物におけるBlimpの不在は、進行性多臓器リンパ球増殖性疾患を導く。
【0045】
従って、本発明のもう一つの局面は、発現されるとき、レポーター分子と共発現されるポリペプチドを産生するポリペプチドをコードする遺伝物質を含む、遺伝子改変細胞またはそのような細胞を含む非ヒト生物を提供し、レポーター分子の検出は、細胞表現型および/または最終分化への細胞の決定付けの指標となる。
【0046】
さらなる態様において、本発明は、発現されるとき、レポーター分子と共発現されるBlimpまたはその一部、フラグメントもしくは機能的な形態を産生する、Blimpポリペプチドをコードする改変Blimp遺伝子を含む、遺伝子改変細胞またはそのような細胞を含む非ヒト生物を提供し、レポーター分子の検出は、細胞表現型および/または最終分化への細胞の決定付けの指標となる。
【0047】
さらに好ましい局面において、本発明は、Blimpコード配列またはその一部、フラグメントもしくは機能的な形態およびレポーター分子コード配列を含む、Blimp mRNA転写産物をコードする改変Blimp遺伝子を含む、遺伝子改変細胞またはそのような細胞を含む非ヒト生物を提供し、この場合、レポーター分子の検出は、細胞表現型および/または最終分化への細胞の決定付けの指標となる。
【0048】
好ましくは、レポーター分子コード配列は、Blimp対立遺伝子のイントロンに挿入される。このようにして、改変Blimp対立遺伝子は、内因性Blimp調節要素の制御下で2シストロンのRNAからレポーターを共産生する。
【0049】
従って、本発明のもう一つの局面は、発現されるとき、レポーター分子と共発現されるポリペプチドを産生する、ポリペプチドをコードする遺伝物質を含む、遺伝子改変細胞またはそのような細胞を含む非ヒト生物を提供し、この場合、造血系の細胞におけるレポーター分子の検出は、細胞表現型および/または最終分化への細胞の決定付けの指標となる。
【0050】
さらなる局面において、本発明は、発現されるとき、レポーター分子と共発現されるBlimpまたはその一部、フラグメントもしくは機能的な形態を産生する、Blimpポリペプチドをコードする改変Blimp遺伝子を含む、遺伝子改変細胞またはそのような細胞を含む非ヒト生物を提供し、B細胞におけるレポーター分子の検出は、レポーター分子活性を有する細胞がASCへの分化に決定付けられていることの指標となる。
【0051】
さらに好ましい局面において、本発明は、Blimpコード配列またはその一部、フラグメントもしくは機能的な形態およびレポーター分子コード配列を含む、Blimp mRNA転写産物をコードする改変Blimp遺伝子を含む、遺伝子改変細胞またはそのような細胞を含む非ヒト生物を提供し、この場合、T細胞におけるレポーター分子の検出は、レポーター分子活性を有する細胞が活性化T細胞であることの指標となる。
【0052】
好ましくは、レポーター分子コード配列は、Blimp対立遺伝子のイントロンに挿入される。このようにして、改変Blimp対立遺伝子は、内因性Blimp調節要素の制御下で2シストロンのRNAからレポーターを共産生する。
【0053】
本明細書においてBlimp-1遺伝子またはその核酸発現産物(RNA)への言及は、その相同体、部分、フラグメント、機能的な形態(低ストリンジェンシー条件下でそれにハイブリダイズするか、最適なアラインメント後に全部分または機能的な部分に対する有意な配列類似性、例えば少なくとも約60%の配列類似性を含む機能的変異体または誘導体を含む)を含む。Blimp-1ポリペプチドまたはタンパク質への言及は、その全ての相同体、部分、フラグメントまたは機能的な形態(最適なアラインメント後に少なくとも60%のアミノ酸配列類似性を有する機能的変異体またな誘導体を含む)を含むように広義で用いられている。
【0054】
本分子の機能的な部分としては、その特定の機能、例えば基質結合、三次コンフォメーションまたは転写活性に重要である完全長分子の部分が挙げられる。転写開始部位は、容易にマッピングされ、プロモーター活性を付与する部位は容易に同定される(例えば、Tunyaplin et al., Nucleic Acid Research 28 (24):4846-4855, 2000参照)。機能的な部分は、分子の発現および活性の調節に重要である。機能的変異体または誘導体は、参照分子の発現および活性の調節に重要な機能活性の少なくとも1つを保持する。Blimp-1に関しては、その発現が、最終分化、ASCによるIg分泌の誘導、およびT細胞の活性化に関連している。
【0055】
改変Blimp遺伝子は、機能的に活性なBlimpポリペプチド、機能的に不活性なBlimpポリペプチド、および/または部分的Blimpポリペプチド、例えばDNA結合モチーフを含むジンクフィンガードメインを欠く、ポリペプチドもしくはペプチドなどをコードすることができる。用語「ポリペプチド」および「タンパク質」は、本発明では交換可能に用いている。
【0056】
ペプチド形の「部分」は、5個未満のアミノ酸を含むエピトープほどの小ささであってもよいし、または数百キロダルトンほどの大きさであってもよい。相同体と比較されるポリペプチド配列の長さは、一般に、少なくとも約16のアミノ酸、通常は少なくとも約20残基、さらに通常は少なくとも約24残基、典型的には少なくとも約28残基および好ましくは約35より多い残基であろう。核酸分子の「一部」は、少なくとも約10ヌクレオチド、または好ましくは約13ヌクレオチド、またはさらに好ましくは少なくとも約20ヌクレオチドの最小サイズを有すると定義し、少なくとも約35ヌクレオチドの最小サイズを有することもある。この定義は、10〜35の範囲内の全てのサイズのヌクレオチド、ならびに50、100、300、500、600ヌクレオチドを含む35より多くのヌクレオチド、またはこれらの値の範囲内の任意の数のヌクレオチドを有する核酸分子を含む。
【0057】
本発明は、変異体Blimpポリペプチドをコードする改変Blimp対立遺伝子も企図する。「変異体」ポリペプチドとしては、天然タンパク質のN末端および/もしくはC末端での1つまたは複数のアミノ酸の欠失(いわゆるトランケーション)もしくは付加;天然タンパク質における1つまたは複数の部位での1つまたは複数のアミノ酸の欠失または付加;または天然タンパク質における1つまたは複数の部位での1つまたは複数のアミノ酸の置換により、天然タンパク質から誘導されるタンパク質が挙げられる。本発明に包含される変異体タンパク質は、生物活性であり、すなわち、天然タンパク質の望ましい生物活性を保持し続けている(すなわち、それらは、例えばc-mycおよび/またはCIITAの転写リプレッサーである)。または、変異体Blimpポリペプチドは、非機能的である。このような変異体は、例えば、遺伝的多形の結果または人間が操作した結果として生じ得る。天然Blimpポリペプチドの生物活性変異体は、デフォルトパラメータを使用する本明細書の他の箇所で説明する配列アラインメントプログラムによって判定して、天然タンパク質についてのアミノ酸配列と少なくとも40%、50%、60%、70%、一般には少なくとも75%、80%、85%、好ましくは約90%から95%またはそれより多く、さらに好ましくは約98%以上の配列類似性を有するであろう。Blimpポリペプチドの生物活性変異体は、ポリペプチドとは、一般に100、50もしくは20個という多さのアミノ酸残基、または好適には1〜15個という少なさのアミノ酸残基、1〜10個という少なさ、例えば6〜10個、5個という少なさ、4、3、2個という少なさ、またはさらには1個のアミノ酸残基が異なり得る。
【0058】
Blimpポリペプチドは、アミノ酸置換、欠失、トランケーションおよび挿入を含む様々な方法で変更することができる。このような操作のための方法は、当技術分野において一般に公知である。例えば、Blimpポリペプチドのアミノ酸配列変異体は、コードされている核酸配列の突然変異により作製することができる。突然変異誘発およびヌクレオチド配列変更のための方法は、当技術分野において周知である。例えば、Kunkel (Proc. Natl. Acad. Sci. USA 82 : 488-492,1985), Kunkel et al.,(Methods in Enzymol. 154 : 367-382,1987),米国特許第4,873,192号, Watson et al. ("Molecular Biology of the Gene", Fourth Edition, Benjamin/Cummings, Menlo Park, Calif., 1987)およびこれらに引用されている参考文献を参照されたい。対象となるタンパク質の生物活性に影響を及ぼす、または及ぼさない適切なアミノ酸置換基についてのガイダンスは、Dayhoffらのモデル(Natl. Biomed. Res. Found 5 : 345-358, 1978)において見出すことができる。例えば、DNA結合モチーフを含有するジンクフィンガードメインの全てまたは一部の欠失は、機能的に不活性であるBlimp変異体を生じさせるであろう。一部の態様において、動物モデルは、一部または全ての組織において遺伝子改変非機能的Blimp対立遺伝子がヘテロ接合性であるが、他の対立遺伝子は、機能的Blimpポリペプチドを発現することができる機能的Blimp対立遺伝子を含む。他の態様において、特定の細胞または組織ではBlimpを発現しないヘテロ接合性動物を産生させる。または1つまたは2つの改変Blimp対立遺伝子により細胞、組織または非ヒト生物において機能的Blimpを産生させることができる。点突然変異またはトランケーションにより作製されたコンビナトリアルライブラリの遺伝子産物をスクリーングするための方法、および選択された特性を有する遺伝子産物についてcDNAライブラリをスクリーニングするための方法は、当技術分野において公知である。このような方法は、Blimpポリペプチドのコンビナトリアル突然変異誘発により作製された遺伝子ライブラリの迅速なスクリーニングに適応できる。ライブラリの中の機能的突然変異体の頻度を増す技法である、再帰的集合型突然変異誘発(Recursive ensemble mutagenesis)(REM)をスクリーニングアッセイと併用して、Blimpポリペプチド変異体を同定することができる(Arkin et al., Proc. Natl. Acad. Sci, USA 89 : 7811-7815, 1992; Delgrave et al. Protein Engineering 6:327-331, 1993)。より詳細に下で論じるような、保存的置換、例えば、1つのアミノ酸と類似した特性を有する別のアミノ酸との交換が望ましいこともある。
【0059】
親Blimpアミノ酸配列と比較すると、変異体Blimpポリペプチドは、配列に沿った様々な位置での保存的アミノ酸配列置換を含有し得る。「保存的アミノ酸置換」は、アミノ酸残基が類似した側鎖を有するアミノ酸残基で置換されることである。類似した側鎖を有するアミノ酸残基のファミリーは、当技術分野において定義されており、それらは、一般に、次のように下位分類することができる:
【0060】
酸性:この残基は、生理pHでHイオンを喪失するため負の電荷を有し、およびこの残基は、水溶液に誘引されて、ペプチドの配座の表面位置を求め、ペプチドが生理pHの水性媒体中にあるときにはそこに収容される。酸性側鎖を有するアミノ酸としては、グルタミン酸およびアスパラギン酸が挙げられる。
【0061】
塩基性:この残基は、生理pHでまたはその1または2pH単位(例えば、ヒスチジン)内でHイオンと会合するため正の電荷を有し、およびこの残基は、水溶液に誘引されて、ペプチドの配座の表面位置を求め、ペプチドが生理pHの水性媒体中にあるときにはそこに収容される。塩基性側鎖を有するアミノ酸としては、アルギニン、リジンおよびヒスチジンが挙げられる。
【0062】
荷電:これらの残基は、生理pHで帯電しており、従って、酸性側鎖または塩基性側鎖を有するアミノ酸(すなわち、グルタミン酸、アスパラギン酸、アルギニン、リジンおよびヒスチジン)を含む。
【0063】
疎水性:これらの残基は、生理pHで帯電しておらず、およびその残基は、水溶液に反発して、ペプチドの配座の内部位置を求め、ペプチドが水性媒体中にあるときにはそこに収容される。疎水性側鎖を有するアミノ酸としては、チロシン、バリン、イソロイシン、ロイシン、メチオニン、フェニルアラニンおよびトリプトファンが挙げられる。
【0064】
中性/極性:これらの残基は、生理pHで帯電していないが、その残基は、ペプチドの配座の内部位置を求めてペプチドが水性媒体中にあるときにそこに収容されることになるほどには水溶液と反発しない。中性/極性側鎖を有するアミノ酸としては、アスパラギン酸、グルタミン酸、システイン、ヒスチジン、セリンおよびトレオニンが挙げられる。
【0065】
本明細書においては、一定のアミノ酸を、それらの側鎖がたとえ極性基を欠く場合であっても疎水性をもたらすほどには大きくないことから、「小さい」とも特性付けしている。プロリンを除き、「小さい」アミノ酸は、少なくとも1つの極性基が1つの側鎖上にあるときには4つ以下の炭素原子を有するアミノ酸、およびそうでないときには3つ以下の炭素原子を有するアミノ酸である。小さい側鎖を有するアミノ酸としては、グリシン、セリン、アラニンおよびトレオニンが挙げられる。遺伝子にコードされている第二アミノ酸プロリンは、ペプチド鎖の二次コンフォメーションに対するその公知の作用のため、特別な場合である。プロリンの構造は、その側鎖がαアミノ基の窒素ならびにα炭素に結合している点で、他の全ての天然由来アミノ酸とは異なる。しかし、幾つかのアミノ酸類似性マトリックス(例えば、Dayhoff et al., 1978 (前記);および Gonnet et al., Science 256 (5062): 1443-1445,1992に例えば開示されているようなPAM120マトリックスおよびPAM250マトリックス)は、グリシン、セリン、アラニンおよびトレオニンと同じグループにプロリンを含む。従って、本発明の目的のために、プロリンは、「小さい」アミノ酸と分類する。
【0066】
極性または非極性と分類するために必要とされる誘引度または反発度は、自由裁量であり、従って、本発明が特に企図するアミノ酸は、様々に分類されている。特に名を挙げていない大部分のアミノ酸は、公知の性質を基に分類することができる。
【0067】
アミノ酸残基は、環状または非環状;芳香族性または非芳香族性;残基の側鎖置換基を基準にした自明の分類;および小さいまたは大きいとさらに下位に分類することができる。残基は、追加の極性置換基が存在するという条件ではカルボキシル炭素を含め合計4個以下の炭素原子を含有する場合;そうでなければ3個以下の場合、小さいと見なす。勿論、小さい残基は、必ず非芳香族である。それらの構造特性に依存して、アミノ酸残基は、2つ以上のクラスに分類することができる。天然由来タンパク質アミノ酸について、この理論体系による下位分類法を表Aに提示する。
【0068】
(表A)アミノ酸下位分類法
【0069】
保存的アミノ酸置換も、側鎖に基づくグループ分けを含む。例えば、脂肪族側鎖を有するアミノ酸のグループは、グリシン、アラニン、バリン、ロイシンおよびイソロイシンであり;脂肪族-ヒドロキシル側鎖を有するアミノ酸のグループは、セリンおよびトレオニンであり;アミド含有側鎖を有するアミノ酸のグループは、アスパラギンおよびグルタミンであり;芳香族側鎖を有するアミノ酸のグループは、フェニルアラニン、チロシンおよびトリプトファンであり;塩基性側鎖を有するアミノ酸のグループは、リジン、アルギニンおよびヒスチジンであり;ならびに硫黄含有側鎖を有するアミノ酸のグループは、システインおよびメチオニンである。例えば、イソロイシンもしくはバリンでのロイシンの置換、グルタミン酸のアスパラギン酸の置換、セリンでのトレオニンの置換、または構造的に関連したアミノ酸でのアミノ酸の同様の置換は、結果として生じる変異体ポリペプチドの特性に重要な影響を及ぼさないであろうと予測するのが妥当である。アミノ酸変更が、結果として機能的Blimpポリペプチドを生じさせるかどうかは、その活性をアッセイすることにより容易に判定することができる。保存的置換を、下記表B中、例示的な置換の見出しの下に示す。さらに好ましい置換を、好ましい置換の見出しの下に示す。本発明の範囲に入るアミノ酸置換は、一般に、(a)その置換領域内のペプチド骨格の構造、(b)標的部位における分子の電荷もしくは疎水性、または(c)側鎖の嵩、を維持することに対するそれらの影響が有意に異ならない置換を選択することにより、達成することができる。置換導入後、変異体を生物活性についてスクリーニングする。
【0070】
(表B)例示的なおよび好ましいアミノ酸置換
【0071】
または、保存的置換を行うために類似したアミノ酸をその側鎖の同一性に基づき3つのカテゴリーにグループ分けすることができる。Zubay, G., Biochemistry, third edition, Wm. C. Brown Publishers (1993)に記載されているように、第一のグループは、グルタミン酸、アスパラギン酸、アルギニン、リジン、ヒスチジンを含み、これらは全て、荷電側鎖を有し;第二のグループは、グリシン、セリン、トレオニン、システイン、チロシン、グルタミン、アスパラギンを含み、および第三のグループは、ロイシン、イソロイシン、バリン、アラニン、プロリン、フェニルアラニン、トリプトファン、メチオニンを含む。
【0072】
従って、Blimpポリペプチド中の予想される非必須なアミノ酸は、典型的に、同じ側鎖ファミリーからの別のアミノ酸残基で置換される。または飽和突然変異誘発などによりBlimpポリヌクレオチドコード配列の全てまたは一部に沿ってランダムに突然変異を導入することができ、結果として生じた突然変異体を親ポリペプチドの活性についてスクリーニングして、その活性を保持する突然変異体を同定することができる。コード配列の突然変異誘発後、コードされたペプチドを組換え発現させることができ、そのポリペプチドの活性を判定することができる。
【0073】
従って、本発明は、天然由来Blimpポリペプチド配列の変異体またはそれらの生物活性フラグメントも企図し、この場合の変異体は、1つまたは複数のアミノ酸残基の付加、欠失または置換により天然由来配列とは区別される。一般に、変異体は、例えば配列番号:2および4のいずれか一方に記載の親Blimpポリペプチド配列との少なくとも約30、40、50、55、60、65、70、75、80、85、90、91、92、93、94、95、96、97、98、99%の類似性を示すであろう。望ましくは、変異体は、例えば配列番号:2および4のいずれか一方に記載の参照Blimpポリペプチド配列との少なくとも30、40、50、55、60、65、70、75、80、85、90、91、92、93、94、95、96、97、98、99%の配列同一性を有するであろう。さらに、1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、30、40、50、60、70、80、90、100以上のアミノ酸の付加、欠失または置換により天然または親配列とは異なるが、親Blimpポリペプチドの特性を保持する配列が考えられる。Blimpポリペプチドは、本明細書において定義するようなストリンジェンシー条件、特に高ストリンジェンシー条件下で、Blimpポリヌクレオチド配列にハイブリダイズするポリヌクレオチドまたはその非コード鎖によりコードされるポリペプチドも含む。
【0074】
一部の態様において、変異体ポリペプチドは、少なくとも1個、しかし50、40、30、20、15、10、8、6、5、4、3または2個未満のアミノ酸残基がBlimp配列と異なる。別の態様において、変異体ポリペプチドは、少なくとも1%、しかし20%、15%、10%または5%未満の残基が配列番号:2および4のいずれか一方における対応する配列と異なる。(この比較にアラインメントが必要な場合、最大類似性のために配列をアラインしなければならない。欠失もしくは挿入またはミスマッチからの「ループド」アウト("Looped" out)配列の差異を考慮に入れる)。好適には、差異は、非必須残基における差異もしくは変更または保存的置換である。
【0075】
「非必須」アミノ酸残基は、態様ポリペプチドの野生型配列から変更することができるが、その活性の1つまたは複数を撤廃しない、または実質的に変更しない残基である。好適には、変更は、これらの活性の1つを実質的に変更せず、例えば、活性は、野生型の少なくとも20%、40%、60%、70%または80%である。「必須」アミノ酸残基は、本発明のBlimpポリペプチドの野生型配列から変更されたとき、結果的に親分子の活性が撤廃される、例えば、野生型活性の20%未満が存在する、残基である。
【0076】
他の態様において、変異体ポリペプチドは、例えば配列番号:2および4のいずれか一方に記載のBlimpポリペプチドの対応する配列との、少なくとも約50%、55%、60%、65%、70%、75%、80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%またはそれより高い類似性を有するアミノ酸配列を含む。
【0077】
本発明は、任意の哺乳動物または動物(鳥類を含む)被験者からの、例えばヒト、非ヒト霊長類、家畜、実験動物、愛玩動物または野生動物からのBlimpを包含する。「Blimp」への言及は、上記のいずれかの種由来のBlimpまたはBlimpならびにその構造的もしくは進化的等価物または相同体を含む。例えば、本発明は、配列番号:2もしくは4との少なくとも約60%の類似性、または配列番号:1、3、5もしくは6との少なくとも約60%の同一性を実質的に有するアミノ酸配列を有するBlimpまたはBlimpを包含する。少なくとも約60%への言及は、60、61、62、63、64%およびそれに続く100%までの級数のうちの連続的な数字を含む。
【0078】
核酸形での分子の機能誘導体は、低ストリンジェンシー条件下で分子またはその相補形にハイブリダイズできるヌクレオチド配列を含む核酸分子を含む。
【0079】
本明細書において用いる用語「類似性」または「同一性」は、ヌクレオチドまたはアミノ酸レベルでの比較される配列間の正確な同一性を含む。ヌクレオチドレベルで同一性がない場合、「類似性」は、それにもかかわらず構造的、機能的、生化学的および/または配座レベルで互いに関連している異なるアミノ酸を結果的に生じる配列間の差異を含む。アミノ酸レベルで同一性がない場合、「類似性」は、それにもかかわらず構造的、機能的、生化学的および/または配座レベルで互いに関連しているアミノ酸を含む。特に好ましい態様において、ヌクレオチドおよびアミノ酸配列比較は、類似性ではなく同一性のレベルで行われる。
【0080】
2つ以上のポリヌクレオチドまたはポリペプチド間の配列の関係を記述するために使用する用語としては、「参照配列」、「比較ウインドウ」、「配列類似性」「配列同一性」、「配列類似率」、「配列同一率」、「実質的に類似した」および「実質的同一性」が挙げられる。「参照配列」は、長さで、少なくとも12、しかししばしば15〜18、多くの場合、少なくとも25以上、例えば30個のモノマー単位(ヌクレオチドおよびアミノ酸残基を含む)である。2つのポリヌクレオチドは、各々、(1)その2つのヌクレオチドの間で類似している配列(すなわち、完全ポリヌクレオチド配列の一部だけ)、および(2)その2つのポリヌクレオチドの間で互いに異なる配列を含み得るので、典型的に、2つの(またはそれより多い)ポリヌクレオチドの間の配列比較は、局所的な配列類似性領域を同定および比較するように「比較ウインドウ」全体にわたってその2つポリヌクレオチドの配列を比較することによって行われる。「比較ウインドウ」は、参照配列と比較される典型的には12個の連続する残基の概念セグメントを指す。2つの配列の最適なアラインメントのために、比較ウインドウは、(付加または欠失を含まない)参照配列と比較して約20%以下の付加または欠失(すなわち、ギャップ)を含み得る。比較ウインドウをアラインするための配列の最適なアラインメントは、コンピュータによるアルゴリズムの実現(米国、ウィスコンシン州、Madison、575 Science Drive、Genetics Computer Group、Wisconsin Genetics Software Package Release 7.0におけるGAP、BESTFIT、FASTA、およびTFASTA)によって、または検査、および選択された任意の様々な方法により生成される最良アラインメント(すなわち、比較ウインドウ全体にわたって最高の相同率を結果として生じるもの)によって行うことができる。例えば、Altschul et al., Nucl. Acids Res. 25. 3389, 1997により開示されているようなプログラムのBLASTファミリーを参照することもできる。配列分析の詳細な論考は、Ausubel et al., Current Protocols in Molecular Biology John Wiley & Sons Inc, 1994-1998, Chapter 15)のUnit 19.3において見出すことができる。
【0081】
本明細書において用いる用語「配列類似性」および「配列同一性」は、配列が、比較ウインドウ全体にわたってヌクレオチドごとのベースまたはアミノ酸ごとのベースで同一であるか、機能的にもしくは構造的に類似していることの程度を指す。従って、例えば「配列同一率」は、比較ウインドウ全体にわたって最適にアラインされた2つの配列を比較し、両方の配列において同一の核酸塩基(例えば、A、T、C、G、I)または同一のアミノ酸残基(例えば、Ala、Pro、Ser、Thr、Gly、Val、Leu、Ile、Phe、Tyr、Trp、Lys、Arg、His、Asp、Glu、Asn、Gln、CysおよびMet)が発生する位置の数を決定してマッチした位置の数を得、そのマッチした位置の数を比較ウインドウ内の全位置数(すなわち、そのウインドウのサイズ)で割り、その結果に100を掛けて配列同一率を得ることによって計算する。本発明の目的のために、「配列同一性」は、DNASISコンピュータプログラム(米国、カリフォルニア州、South San FranciscoのHitachi Software engineering Co.,Ltd.から入手できる、Windows用Version 2.5)により、そのソフトウェアに添付されている参照マニュアルにおいて使用されているような標準デフォルトを使用して計算される「マッチ率」を意味すると理解されよう。同様のコメントが配列類似性に関連して適用される。
【0082】
さらに、Blimp相同体または誘導体は、低ストリンジェンシー条件下で、配列番号:1、3、5もしくは6、またはそれらからの相補形にハイブリダイズすることができると定義することができる。
【0083】
本明細書において低ストリンジェンシーへの言及は、ハイブリダイゼーションのために少なくとも約0から少なくとも約15% v/vのホルムアミドおよび少なくとも約1Mから少なくとも約2Mの塩、ならびに洗浄条件のために少なくとも約1Mから少なくとも約2Mの塩を含み、包含する。一般に、低ストリンジェンシーは、約25〜30℃から約42℃である。この温度は、変更することができ、使用される温度が高いほど、ホルムアルデヒドが置換され、および/または代替ストリンジェンシー条件がもたらされる。必要な場合には、ハイブリダイゼーションのために少なくとも約16% v/vから少なくとも約30% v/vのホルムアミドおよび少なくとも約0.5Mから少なくとも約0.9Mの塩、ならびに洗浄条件のために少なくとも約0.5Mから少なくとも約0.9Mの塩を含み、包含する中ストリンジェンシー、またはハイブリダイゼーションのために少なくとも約31% v/vから少なくとも約50% v/vのホルムアミドおよび少なくとも約0.01Mから少なくとも約0.15Mの塩、ならびに洗浄条件のために少なくとも約0.01Mから少なくとも約0.15Mの塩を含み、包含する、高ストリンジェンシーなどの代替ストリンジェンシー条件を適用することができる。一般に、洗浄は、Tm=69.3+0.41(G+C)%(Marmur et al., J. Mol. Biol. 5: 109, 1962)で行われる。しかし、2本鎖DNAのTmは、ミスマッチ塩基対の数が1%増加するごとに1℃減少する(Bonner et al., Eur. J. Biochem. 46: 83, 1974)。ホルムアミドは、これらのハイブリダイゼーション条件では任意である。従って、特に好ましいストリンジェンシーレベルは、次のように定義される:低ストリンジェンシーは、25〜42℃で、6×SSC緩衝液、0.1% w/v SDSであり;中ストリンジェンシーは、20℃から65℃の範囲の温度で、2×SSC緩衝液、0.1% w/v SDSであり;高ストリンジェンシーは、少なくとも65℃の温度で、0.1×SSC緩衝液、0.1% w/v SDSである。
【0084】
好ましくは、改変Blimp遺伝子は、Blimpの対立遺伝子の全てまたは一部分を含む核酸構築物を使用し、それにレポーター分子をコードする核酸配列を挿入することにより改変される。
【0085】
レポーター分子は、便利には、レポーター発現カセットまたはレポーター構築物によりコードされる。レポーター構築物は、Blimp-1調節要素の制御下で導入され得、および細胞、組織または生物におけるBlimp-1発現パターンを忠実にレポートすることができる。
【0086】
「レポーター」とは、レポーター遺伝子により一般にコードされ、レポーターアッセイで測定される任意の分子、タンパク質またはポリペプチドを意味する。レポーターは、遺伝子配列の活性の理解を可能にする検出可能なシグナルを提供する。それらは、活性を直接レポートすることができ、または下流の標的の活性をモニターすることにより間接的にモニターすることができる。レポータータンパク質は、他のタンパク質と区別可能でなければならず、理想的には容易に定量されなけらばならない。それによって決定されたエピトープと抗体の間の反応性は、任意で二次またはさらに進んだ抗体と共に、容易に利用することができる。一般的なレポータータンパク質としては、ルシフェラーゼ、クロラムフェニコールトランスフェラーゼ(CAT)、β-ガラクトシダーゼ(B-gal)、または蛍光タンパク質、例えば緑色蛍光タンパク質(GFP)が挙げられる。本明細書においてGFPへの言及は、任意の蛍光または発光タンパク質(クラゲまたは他の生物に由来するものを含む)、およびDSRed、HcRed(Clontech)またはhrGFP(Stratagene)などの色変異体を含む全ての相同体、誘導体、類似体を包含することを意味する。好ましくは、レポーター発現カセットは、蛍光または他の発光GFPをコードする。GFPレポーターは、生細胞において容易に検出することができ、細胞選別用途に特に有用であり、好ましい。
【0087】
蛍光または発光マーカーの例は、次の表2に含まれているものであるが、これらに限定されない中から選択することができる。
【0088】
(表2)
1 Ex:ピーク励起波長(nm)
2 Em:ピーク発光波長(nm)
【0089】
蛍光発光を分析する任意の好適な方法は、本発明に包含される。これに関して、本発明は、例えばLakowicz et al., Biophys. J. 72: 567, 1997により開示されているような、二光子および三光子時間分解蛍光顕微鏡;例えば参照として本明細書に組み入れられるEriksson et al., Biophys. J. 2: 64, 1993に開示されているような、蛍光寿命イメージング;および例えばYouvan et al., Biotechnology et Elia 3: 1-18, 1997により開示されているような、蛍光共鳴エネルギー移動を含むが、それらに限定されない技法を企図する。
【0090】
本発明に従って使用することができる例示的な蛍光体としては、Dowerら(国際特許公開公報第93/06121号)により開示されたものが挙げられる。好ましくは、蛍光染料を利用する。蛍光色素を本レポーター分子に組み込むために、任意の適する蛍光染料を使用することができる。例えば、非常に多数の蛍光色素を記載している米国特許第5,573,909号(Singerら)および同第5,326,692号(Brinkleyら)を参照することができる。米国特許第5,227,487号、同第5,274,113号、同第5,405,975号、同第5,433,986号、同第5,442,045号、同第5,451,663号、同第5,453,517号、同第5,459,276号、同第5,515,864号、同第5,648,270号および同第5,723,218号に記載されている蛍光色素も参照することができる。
【0091】
最新のフローサイトメータは、これらのタスクを1秒あたり100,000細胞/粒子まで行うことができる。フィルターおよびダイクロイックミラーの光学アレイの使用により、異なる蛍光波長を同時に分離、検出することができる。加えて、異なる励起波長を有する多数のレーザーを使用することができる。それ故、様々な蛍光体を使用して、ターゲティングし、例えば個々の細胞の細胞内および細胞外特性を検査することができる。散乱光測定により、個々の細胞のサイズ、形状、粒度および/または複雑性ならびに、それ故、対象となる特定の集団への所属を分類することもできる(Shapiro, Practical flow cytometry, 3rd Ed., Brisbane, Wiley-Liss, 1995)。
【0092】
本発明の方法において使用することができる好適なフローサイトメトリーとしては、単一励起レーザー、一般には488nmのスペクトル線を用いて15mWで動作するアルゴンイオン空気冷却レーザー、を使用して、5〜9の光学パラメータを測定するもの(表3参照)が挙げられる。より最新式のフローサイトメータは、アルゴンイオンレーザー(488または514nm)に加えてHeNeレーザー(633nm)またはHeCdレーザー(325nm)などの多重励起レーザーを使用することができる。担体についての、異なる光学検出可能/定量可能な属性に対応する光学パラメータを、フローサイトメータによって測定して、その担体のコード(または多次元空間でのアドレス指定能力)をもたらす質的および/または量的情報のマトリックスを生じることができる。
【0093】
例えば、Biggsら(Cytometry 36: 36-45, 1999)は、細胞を免疫表現型解析する(すなわち、分類する)の目的のために、3つの励起レーザーを使用する11パラメータのフローサイトメータを構築し、前方および側方散乱測定に加えて9つの区別可能なフルオロフォア(fluorophore)の使用を実証した。現在市販されている最大パラメータ数は、17:前方散乱、側方散乱、および各々が5つの蛍光検出器を有する3つの励起レーザー、である。全てのパラメータを適切に使用することができるかどうかは、吸光係数、量子収率、および全ての蛍光体の間のスペクトルのオーバーラップ量に依存する(Malemed et al., "Flow cytometry and sorting" ,2nd Ed., New York, Wiley-Liss, 1990)。しかし、本発明は、いずれの特定のフローサイトメータにも、いずれの特定のパラメータセットにも限定されないことは理解されるだろう。これに関して、本発明は、従来のフローサイトメータの代わりに、例えばFu et al., Nature Biotechnology 17: 1109-1111, 1999に開示されているような微細加工フローサイトメータの使用も企図する。
【0094】
(表3)フローサイトメータにより測定することができる例示的な光学パラメータ
* 488nm励起レーザーを使用
† バンドパス(bandpass)フィルタの幅
# ロングパス(longpass)フィルタ
【0095】
この選別能力を有するフローサイトメータは、「蛍光活性化細胞選別装置」(FACS)として公知である。従って、検出可能にユニークな担体の集団を得る本発明の方法における選別段階は、蛍光活性化細胞選別(FACS)などによるフローサイトメトリー技法によって行うことができる。とはいえ、本発明に関して、FACSは、より正確には「蛍光活性化担体または固体支持体選別」である(例えば、"Methods in Cell Biology" Vol. 33, Darzynkiewica, Z. and Crissman, H. A. , eds. , Academic Press参照)。
【0096】
さらなる実施形態において、本発明は、造血細胞系の細胞を表現型解析および/またはモニターするための方法を提供し、この方法は、改変Blimp遺伝子を含む遺伝子改変細胞またはそのような細胞を含む非ヒト生物のスクリーニングを含み、この場合、遺伝子の発現は、細胞表現型および/または最終分化への細胞の決定付けの指標となる。造血細胞としては、B細胞、T細胞、樹状細胞、マクロファージおよびナチュラルキラー細胞、顆粒球、好酸球、赤血球、巨核球、骨髄、間質、脾細胞前駆細胞ならびにこれらの誘導体が挙げられるが、それらに限定されるわけではない。
【0097】
好ましくは、改変Blimp遺伝子は、発現されるとき、レポーター分子と共発現される、Blimpまたはその一部、フラグメントもしくは機能的な形態を産生する、Blimpコード配列またはその一部、フラグメントもしくは機能形態およびレポーター分子コード配列を含むBlimp mRNA転写産物をコードし、この場合、レポーター分子の検出は、細胞表現型および/または最終分化への細胞の決定付けの指標となる。
【0098】
さらなる態様において、レポーター活性またはレポーター活性の変更を示す細胞を、レポーター活性を示さない細胞の中から単離または選択する。レポーター活性細胞の単離は、フローサイトメトリー、レーザー走査サイトメトリー、クロマトグラフィーおよび/または他の同等の手順によるものであり得る。加えて、さらなる選択マーカーを使用して、本発明の改変細胞を単離または選択することができる。フローサイトメトリー単離が特に好ましい。
【0099】
好ましくは、細胞は、B細胞系列の細胞集団において同定または単離されたASCである。
【0100】
従って、本発明は、実質的にB細胞の集団から実質的に精製されたASC集団を単離する方法を提供し、該方法は、改変Blimp遺伝子を含む遺伝子改変細胞またはそのような細胞を含む非ヒト生物を、ASCへの分化を誘導することができる薬剤または組成物と接触させることを含み、該遺伝子の発現または活性は、レポーター構築物によりレポートされ、ならびに該レポーター活性の検出は、レポーター分子活性を有する細胞がASCであることの指標となり、必要な場合には、該生物からB細胞を単離し、レポーター分子の活性を基にASCを単離する。
【0101】
好ましくは、改変細胞は、発現されるとき、レポーター分子と共発現されるBlimpまたはその一部、フラグメントもしくは機能的な形態を産生する、Blimpコード配列またはその一部、フラグメントもしくは機能的な形態およびレポーター分子コード配列を含むBlimp mRNA転写産物をコードする改変Blimp遺伝子を含み、レポーター活性は、レポーター分子活性を有する細胞がASCであることの指標となる。
【0102】
好ましくは、細胞のスクリーニングは、蛍光レポーター分子のフローサイトメトリー分析により達成される。
【0103】
B細胞は、例えば密度勾配遠心分離、フローサイトメトリー、または磁性ビーズの使用により、生物またはサンプルから便利に単離される。B細胞を選択的に、クローン的にもしくは多クローン的にまたは別様に有効に活性化し、それらのASCへの分化を誘導する任意の薬剤または組成物を包含する。多クローン性活性化因子の一例は、LPSである。
【0104】
一つの態様において、レポーターはGFPであり、該ASCは、フローサイトメトリーにより単離される。
【0105】
「実質的に精製された」とは、ASCが、細胞の少なくとも約60から95%、好ましくは少なくとも約97%、さらに好ましくは少なくとも約99%を含み、例えば、少なくとも約60、61、62、63、64%およびそれに続く100%までの級数における連続的な数字である。または、未選別細胞より約100,000倍の濃縮が企図される。
【0106】
本発明は、ワクチンの抗原性、またはASCによる抗体産生を強化または抑制する薬剤の能力を検査するための方法も提供し、低減されたレポーター活性は、抗体応答およびレポーター活性を下方調節もしくは阻害する薬剤の指標となり、または対照を基準にして強化されたレポーター活性は、抗体応答の陽性調節因子である薬剤の指標となる。この局面によると、この方法は以下の段階を含む:
(i)遺伝子改変細胞またはそのような細胞を含む非ヒト動物に薬剤またはワクチンを投与する段階であって、細胞または生物は、発現されるとき、レポーター分子と共発現される、Blimpまたはその一部、フラグメントもしくは機能的な形態を産生する、Blimpポリペプチドをコードする改変Blimp-1遺伝子を含む、段階;
(ii)その存在が、細胞表現型およびASCによる抗体産生を調節する該薬剤またはワクチンの能力の指標となる、レポーター分子の存在について細胞または生物を検査する段階。
【0107】
もう一つの態様において、本発明は、遺伝子組成物またはタンパク質性組成物を含むワクチンの抗原性または免疫原性を検査するための方法を提供し、この方法は以下の段階を含む:
(i)遺伝子改変細胞またはそのような細胞を含む非ヒト動物にワクチンを投与する段階であって、細胞または生物は、発現されるとき、レポーター分子と共発現される、Blimpまたはその一部、フラグメントもしくは機能的な形態を産生する、Blimpポリペプチドをコードする改変Blimp-1遺伝子を含む、段階;ならびに
(ii)その存在が、T細胞および/またはB細胞の活性化を調節するワクチンの能力の指標となる、レポーター分子の存在について細胞または生物を検査する段階。
【0108】
一部の態様において、Blimp遺伝子は、Blimpコード配列またはその一部、フラグメントもしくは機能的な形態およびレポーター分子コード配列を含む、Blimp mRNA転写産物をコードする。他の態様において、レポーター分子コード配列は、Blimp対立遺伝子のイントロン内に挿入される。さらなる態様において、改変Blimp対立遺伝子は、ホモ接合またはヘテロ接合の形態で存在する。アッセイの目的に依存して、一部の態様において、改変Blimp対立遺伝子は、機能的Blimp転写因子またはその機能的な部分、形態、相同体もしくは変異体をコードする。他の態様において、改変Blimp対立遺伝子は、非機能的Blimp転写因子またはその非機能的な部分、形態、相同体もしくは変異体をコードする。実例となる態様において、細胞または遺伝物質は、ヒト、非ヒト霊長類、家畜、愛玩生物、実験検査生物、爬虫類または両生類に由来する。実験検査動物の例としては、齧歯動物(マウスを含む)、モルモット、ブタ、カモ、ウサギまたはヒツジが挙げられる。
【0109】
本明細書において開示する方法の幾つかの実例となる態様において、細胞は、造血細胞または胚細胞である。本明細書において開示する場合のBlimpは、B細胞とT細胞両方の最終分化に不可欠であり、従って、好ましい細胞系列は、リンパ球細胞である。特定の態様において、リンパ球細胞タイプは、B細胞およびT細胞から選択される。B細胞の場合、その最終分化形態は、ASCであり、これらの細胞は、本明細書において開示する方法を使用して、さらに、実質的に精製することができる。細胞がB細胞である場合、最終分化T細胞としては、CD4+ T細胞およびCD8+ T細胞が挙げられるが、これらに限定されるわけではない。便利には、レポーター分子の検出は、細胞表現型および/または特定の条件下または試験薬剤の存在下での最終分化への細胞の決定付けの指標または予測となる。さらにいっそいう便利には、レポーター分子は、蛍光または発光レポーター分子である。
【0110】
本発明は、細胞分化を調節する際に使用するための、本明細書において記載の方法により同定される、Blimp-1発現またはBlimp-1活性の拮抗薬および作動薬を含む、ASCなどであるが、これに限定されない、細胞の最終分化の拮抗薬または作動薬にも関する。この調節成分、作動薬または拮抗薬が向けられる分子を、本明細書においては総称して「標的」または「標的分子」と呼ぶ。
【0111】
従って、もう一つの態様では、本発明は、造血細胞における最終分化の作動薬または拮抗薬についてのインビボまたはインビトロスクリーニング方法を提供し、この方法は、以下の段階を含む:遺伝子改変細胞またはそのような細胞を含む非ヒト動物に1つまたは複数の薬剤を暴露する段階であって、細胞または生物は、発現されるとき、レポーター分子と共発現されるBlimpまたはその一部、フラグメントもしくは機能的な形態を産生する、Blimpポリペプチドをコードする改変Blimp-1遺伝子を含む、段階;およびレポーター分子の存在またはレポーター分子のレベルの変化について細胞または生物を検査する段階であって、レポーター分子の存在は、最終分化を作動し、またはそれに拮抗する1つまたは複数の薬剤の能力の指標となる、段階。Blimpの作動薬は、造血細胞の最終分化を直接または間接的に誘導し、ならびに例えば癌および/または自己免疫疾患の治療または予防の際に、ならびに病的感染への適切な免疫応答を促進する際に有用である。最終分化細胞の発生を阻害する分子は、狼瘡患者などの自己免疫患者において、またはアレルギーの場合などの免疫機能不全を治療する際に有用である。
【0112】
好ましくは、改変細胞は、発現されるとき、レポーター分子と共発現されるBlimpまたはその一部、フラグメント、相同体、変異体、誘導体、機能的な形態もしくは非機能的な形態を産生する、Blimpコード配列またはその一部、フラグメントもしくは機能的な形態およびレポーター分子コード配列を含むBlimp mRNA転写産物をコードする改変Blimp遺伝子を含む造血細胞であり、ここで、レポーター活性は、レポーター分子活性を有する細胞が最終分化するかまたは最終分化に決定付けられていることの指標となる。さらに好ましくは、造血細胞は、リンパ球系列の細胞である。一部の態様において、最終分化細胞はASCであり、他の態様において、最終分化細胞は、CD4 T細胞および/またはCD8 T細胞である。改変Blimp対立遺伝子は、細胞、組織または非ヒト生物中にホモ接合またはヘテロ接合の形態で存在する。さらに、特定の用途によっては、Blimp対立遺伝子は、転写(機能)活性Blimpポリペプチドを発現する。従って、あるアッセイでは、細胞における最終分化を調節または誘導するために機能的Blimpポリペプチドを有することが有用であろう。他の形態では、改変Blimp対立遺伝子は、機能的Blimpを発現せず、Blimp対立遺伝子が発現されたかどうか、またはBlimp発現のレベルが機能Blimpポリペプチドを産生することができる細胞において調節されたかどうかを、レポーター活性の検出によって判定することで十分であろう。
【0113】
細胞(インビトロ)アッセイは、特に便利であり、細胞においてその活性を容易に検出することができるレポーター分子とカップリングさせたときのアッセイは、ハイスループットスクリーニングに理想的に適する。当業者には公知であるように、多数の異なる形式が利用可能である。一つの有用な例は、Ulleras et al., Toxicology 206 (2) : 245-256, 2005に記載されている。
【0114】
分子または分化状態の「調節」は、その機能活性または分化の全てまたは一部を完全にまたは部分的に阻害または低減または下方調節すること、およびその機能活性または分化の全てまたは一部を強化または上方調節することを含む。分子が、遺伝子配列である場合、その機能活性は、例えば、その結合能力または転写もしくは翻訳活性、またはその半減期を調節することにより、調節することができる。分子が、コードされたポリペプチドである場合、その機能活性は、例えば、その結合能力、その半減期、細胞もしくは膜における位置、またはその酵素活性を調節することにより、調節することができる。調節成分は、調節を達成する作動薬または拮抗薬である。強化された分化は、低減された細胞分裂の指標にもなる。
【0115】
拮抗薬または作動薬の一例は、タンパク質、ポリペプチドまたはペプチドである。これらの用語は、交換可能に用いることができる。これらの用語は、アミノ酸のポリマーおよびその同等物を指し、特定長の産物を指さず、従って、ポリペプチド、ペプチド、オリゴペプチドおよびタンパク質は、ポリペプチドの一定義に包含される。これらの用語は、ポリペプチドの改変、例えば、グリコシル化、アセチル化、リン酸化なども除外しない。この定義に包含されるものは、例えば、表4に与えるものなどの、例えば非天然アミノ酸を含むアミノ酸の1つまたは複数の類似体を含有するポリペプチド、または置換されている結合を有するポリペプチドである。このようなポリペプチドは、細胞に進入することができるために必要であり得る。アミノ酸の化学的類似体を有するポリペプチドは、プロテアーゼ媒介消化に対して、より耐性であり得る。拮抗薬または作動薬の一例は、Blimpの化学的類似体である。拮抗薬および作動薬は、例えばc-myc発現がBlimp-1により抑制されるなど、Blimpが相互作用する分子に作用することができる。
【0116】
遺伝分子は、作動薬および拮抗薬調節成分へも発達させられる。用語「遺伝分子」、「核酸」、「ヌクレオチド」および「ポリヌクレオチド」は、当業者には容易に理解されるように、RNA、cDNA、ゲノムDNA、合成形および混合型ポリマー、センスとアンチセンス鎖の両方を含み、化学的にもしくは生化学的に改変されていてもよいし、または非天然もしくは誘導体化ヌクレオチド塩基を含有していてもよい。このような改変としては、例えば、ラベル;メチル化;1つまたは複数の天然由来ヌクレオチドの類似体(例えば、モルホリン環)での置換;ヌクレオチド間調節、例えば非荷電結合(例えば、メチルホスホネート、ホスホトリエステル、ホスホアミデート、カルバメートなど)、荷電結合(例えば、ホスホロチオエート、ホスホロジチオエートなど)、ペンデント部分(pendent moiety)(例えば、ポリペプチド)、インターカレーター(intercalator)(例えば、アクリジン、ソラレンなど)、キレート化剤、アルキル化剤および改変結合(例えば、α-アノマー核酸など)が挙げられる。水素結合および他の化学的相互作用により指定配列に結合する能力に関してポリヌクレオチドを模倣する合成分子も含まれる。このような分子は当技術分野において公知であり、このような分子としては、例えば、分子骨格内でペプチド結合がリン酸エステル結合と置き換わっているものが挙げられる。アンチセンス分子の改変は周知であり、Kurrek, Eur. J. Biochem. 270:1628-1644, 2003に要約されている。
【0117】
例えばアンチセンスポリヌクレオチド配列は、転写産物のサイレンシングに有用である。さらに、Blimp遺伝子座の全てまたは一部を含有するポリヌクレオチドベクターを、プロモータの制御下、アンチセンス配向に配置し、細胞に導入することができる。細胞内でのこのようなアンチセンス構築物の発現は、標的転写および/または翻訳に干渉するであろう。そのような分子は、自己免疫状態における免疫応答の抑制に特に有用であり得る。さらに、共抑制、およびRNAiまたはsiRNAを誘導するメカニズムを利用することもできる。またはアンチセンスもしくはセンス分子を直接投与することができる。後者の形態では、アンチセンスまたはセンス分子を組成物に調合し、その後、任意の数の手段により標的細胞に投与することができる。
【0118】
アンチセンスおよびセンス分子に関する変形は、モルホリンヌクレオチド誘導体およびホスホロジアミデート結合からなるオリゴヌクレオチドであるモルホリノの使用を伴う(例えば、Summerton and Weller, Antisense and Nucleic Acid Drug Development 7: 187-195,1997)。このような化合物を胚に注入し、mRNAでの干渉の影響を観察する。
【0119】
一つの態様において、本発明は、Blimpをコードする核酸分子の機能または作用を調節する際に使用するためにオリゴヌクレオチドおよび類似の化学種などの化合物を利用する。すなわち、オリゴヌクレオチドが、転写または翻訳後遺伝子サイレンシングを誘導する。これは、内因性リガンドをコードする1つまたは複数の核酸分子と特異的にハイブリダイズするオリゴヌクレオチドを提供することにより達成される。オリゴヌクレオチドは、細胞に直接提供してもよいし、または細胞内で発生させてもよい。本明細書において用いる場合、用語「標的核酸」および「阻害剤をコードする核酸分子」は、阻害剤をコードするDNA、そのようなDNAから転写されたRNA(プレmRNAおよびmRNAまたはそれらの一部を含む)、ならびにまたそのようなRNAから誘導されたcDNAを包含するように、便宜上、用いている。本発明の化合物とその標的核酸のハイブリダイゼーションを、一般に、「アンチセンス」と呼ぶ。従って、本発明の一部の好ましい態様の実施に含まれると考えられる好ましいメカニズムを、本明細書においては「アンチセンス阻害」と呼ぶ。典型的に、このようなアンチセンス阻害は、少なくとも1つの鎖またはセグメントが切断、分解または別様に作動不能にされるような、オリゴヌクレオチド鎖またはセグメントの水素結合に基づくハイブリダイゼーションに基づく。これに関して、そのようなアンチセンス阻害は、特定の核酸分子およびそれらの機能をターゲットにするのが、本発明では好ましい。
【0120】
干渉されるDNAの機能としては、複製および転写を挙げることができる。例えば、複製および転写は、内因性細胞テンプレート、ベクター、プラスミド構築物またはその他由来であり得る。干渉されるRNAの機能としては、タンパク質翻訳部位へのRNAの転移、RNA合成部位から遠い細胞内の部位へのRNAの転移、RNAからのタンパク質の翻訳、1つまたは複数のRNA種を生じさせるようなRNAのスプライシング、およびRNA内に束縛されるかRNAにより助長され得るRNAの関与する触媒活性または複合体形成、といった機能を挙げることができる。一例では、標的機能へのこのような干渉の結果、Blimpレベルが低下する。本発明の状況において、「調節」および「発現の調節」は、遺伝子をコードする核酸分子、例えばDNAまたはRNAの量またはレベルの増加(刺激)または低下(阻害)のいずれかを意味する。阻害は、多くの場合、発現の調節の好ましい形態であり、mRNAは、多くの場合、好ましい標的核酸である。
【0121】
アンチセンス化合物は、標的核酸への化合物の結合が標的核酸の正常な機能に干渉して活性を喪失させるとき、ならびに特異的結合が望まれる条件下、すなわちインビボアッセイまたは治療的処置の場合には生理条件下、およびインビトロアッセイの場合にはアッセイを行う条件下で、非標的核酸配列へのアンチセンス化合物の非特異的結合を回避するために十分な程度の相補性があるとき、特異的にハイブリダイズすることができる。
【0122】
本明細書において用いる場合、「相補的な」は、オリゴマー化合物の2つの核酸塩基間の正確な対合能力を指す。例えば、オリゴヌクレオチド(オリゴヌクレオチド化合物)の一定の位置の核酸塩基が、標的核酸の一定の位置の核酸塩基と水素結合することができる場合(標的核酸は、DNA、RNAまたはオリゴヌクレオチド分子である)には、そのオリゴヌクレオチドと標的核酸間の水素結合の位置は、相補的位置と考える。そのオリゴヌクレオチドとさらなるDNA、RNAまたはオリゴヌクレオチド分子は、各分子中の十分な数の相補的位置が、互いに水素結合することができる核酸塩基によって占有されているとき、互いに相補的である。従って、「特異的にハイブリダイズ可能な」および「相補的な」は、安定で特異的な結合が、オリゴヌクレオチドと標的核酸の間に発生するような十分な数の核酸塩基に関する正確な対合または相補性の十分な程度を示すために使用する用語である。
【0123】
本発明によると、化合物としては、アンチセンスオリゴマー化合物、アンチセンスオリゴヌクレオチド、リボザイム、外部ガイド配列(EGS)オリゴヌクレオチド、可変スプライサー(alternate splicer)、プライマー、プローブ、および標的核酸の少なくとも一部分にハイブリダイズする他のオリゴマー化合物が挙げられる。従って、これらの化合物は、1本鎖、2本鎖、環状またはヘアピンオリゴマー化合物の形態で導入することができ、ならびに内部または末端バルジ(bulge)またはループなどの構造要素を含有することができる。系に導入されると、本発明の化合物は、標的核酸の改変を果たすように、1つまたは複数の酵素または構造タンパク質の作用を惹起する。このような酵素の1つの非限定的な例は、RNアーゼH(RNA:DNA二重鎖のRNA鎖を切断する細胞エンドヌクレアーゼ)である。「DNA様」である1本鎖アンチセンス化合物が、RNアーゼHを惹起することは、当技術分野においては公知である。従って、RNアーゼHの活性化は、結果としてRNA標的の切断を生じさせ、それにより遺伝子発現のオリゴヌクレオチド媒介阻害の効率が大きく上昇する。同様の役割が、酵素のRNアーゼIIIおよびリボヌクレアーゼLファミリーのものなどの他のリボヌクレアーゼにおいて仮定されている。
【0124】
アンチセンス化合物の好ましい形態は、1本鎖アンチセンスオリゴヌクレオチドであるが、多くの種において、2本鎖構造、例えば2本鎖RNA(dsRNA)分子の導入は、遺伝子またはその関連遺伝子産物の機能の、強力で特異的なアンチセンス媒介低下を誘導することが証明されている。この現象は、植物においても動物においても発生する。
【0125】
本発明の状況において、用語「オリゴマー化合物」は、複数のモノマー単位を含むポリマーまたはオリゴマーを指す。本発明の状況において、用語「オリゴヌクレオチド」は、リボ核酸(RNA)もしくはデオキシリボ核酸(DNA)またはその模倣体、キメラ、類似体および相同体のオリゴマーまたはポリマーを指す。この用語は、天然由来核酸塩基、糖および共有結合性ヌクレオシド間(骨格)結合からなるオリゴヌクレオチド、ならびに同様に機能する天然由来タンパク質を有するオリゴヌクレオチドを包含する。このような改変または置換オリゴヌクレオチドは、例えば細胞取り込み向上、標的核酸に対する親和性向上およびヌクレアーゼの存在下での安定性上昇などの望ましい特性のため、多くの場合、天然形態より好ましい。
【0126】
オリゴヌクレオチドは、本発明の化合物の好ましい形態であるが、本発明は、本明細書において記載するものなどのオリゴヌクレオチド、類似体および模倣体を含むが、これらに限定されるわけではない、化合物の他のファミリーも企図する。
【0127】
翻訳開始コドンと翻訳終結コドンの間の領域を指すことが当技術分野において公知である、オープンリーディングフレーム(ORF)または「コード領域」は、有効にターゲティングすることができる領域である。本発明の状況の中で、1つの領域は、遺伝子のオープンリーディングフレーム(ORF)の翻訳開始または終結コドンを包含する遺伝子内領域である。
【0128】
他の標的領域としては、翻訳開始コドンから5’方向にあるmRNAの一部を指すことが当技術分野においては公知であり、従って、mRNA(またはその遺伝子の対応するヌクレオチド)の5’キャップ部位と翻訳開始コドンの間のヌクレオチドを含む、5’非翻訳領域(5’UTR);ならびに翻訳終結コドンから3’方向にあるmRNAの一部を指すことが当技術分野においては公知であり、従って、mRNA(またはその遺伝子の対応するヌクレオチド)の翻訳終結コドンと3’末端の間のヌクレオチドを含む、3’非翻訳領域(3’UTR)が挙げられる。mRNAの5’キャップ部位は、5’-5’三リン酸結合によりmRNAの5’の大部分の残基に連結しているN7-メチル化グアノシン残基を含む。mRNAの5’キャップ領域は、5’キャップ構造それ自体、ならびにそのキャップ部位に隣接する最初の50のヌクレオチドを含むと考えられる。5’キャップ領域のターゲティングも好ましい。
【0129】
当技術分野において公知であるように、ヌクレオシドは、塩基と糖の組み合わせである。ヌクレオシドの塩基部分は、一般には複素環式塩基である。そのような複素環式塩基の2つの最も一般的なクラスは、プリンおよびピリミジンである。ヌクレオチドは、そのヌクレオシドの糖部分に共有結合で連結されたリン酸基をさらに含むヌクレオシドである。ペントフラノシル糖を含むヌクレオシドについては、リン酸基をその糖の2’、3’または5’ヒドロキシル部分のいずれかに連結させることができる。オリゴヌクレオチドを形成する場合、リン酸基は、隣接するヌクレオシドと互いに共有結合で連結して、線状ポリマー化合物を形成する。そして次に、この線状ポリマー化合物のそれぞれの末端をさらに連結させて、環状化合物を形成することができるが、線状化合物が一般に好ましい。加えて、線状化合物は、内部核酸塩基相補性を有することがあり、従って、完全または部分的2本鎖化合物を生じさせるような様式でフォールディングすることがある。オリゴヌクレオチド内のリン酸基は、そのオリゴヌクレオチドのヌクレオシド間骨格を形成すると一般に言及されている。RNAおよびDNAの正常な結合または骨格は、3’-5’ホスホジエステル結合である。
【0130】
本発明において有用な好ましいアンチセンス化合物の具体的な例としては、改変された骨格または非天然ヌクレオシド間結合を含有するオリゴヌクレオチドが挙げられる。本明細書において定義するように、改変された骨格を有するオリゴヌクレオチドは、その骨格内にリン原子を保持するもの、およびその骨格内にリン原子を有さないものを含む。本明細書の目的のために、および当技術分野において時として参照されるように、ヌクレオシド間骨格内にリン原子を有さない改変オリゴヌクレオチドもオリゴヌクレオシドとみなすことができる。
【0131】
中にリン原子を含有する好ましい改変オリゴヌクレオチド骨格としては、例えば、正常な3’-5’結合を有する、ホスホロチオエート、キラルホスホロチオエート、ホスホロジチオエート、ホスホトリエステル、アミノアルキルホスホトリエステル、メチルおよび他のアルキルホスホネート(3’-アルキレンホスホネート、5’-アルキレンホスホネートを含む)およびキラルホスホネート、ホスフィネート、ホスホルアミデート(3’-アミノホスホルアミデートを含む)およびアミノアルキルホスホルアミデート、チオノホスホルアミデート、チオノアルキルホスホネート、チオノアルキルホスホトリエステル、セレノホスフェートおよびボラノホスフェート、および、これらの2’-5’連結型類似体、ならびに1つまたは複数のヌクレオチド間結合が3’-3’、5’-5’、または2’-2’結合である、逆極性を有するものが挙げられる。逆極性を有する好ましいオリゴヌクレオチドは、3’の大部分のヌクレオチド間結合に単一3’-3’結合、すなわち、無塩基であり得る(核酸塩基が欠失しているか、その位置にヒドロキシル基を有する)単一の逆ヌクレオシド残基、を含む。様々な塩、混合塩および遊離酸形態も含まれる。
【0132】
本発明の単離または組換え作動薬および拮抗薬は、直接使用されるか、薬学的試薬または他の試薬としてのそれらの有効性を向上させるために、当技術分野において周知の方法によりさらに改変することができる。活性化合物についての重要な考慮事項としては、調合物および送達方法が挙げられる。
【0133】
作動薬または拮抗薬は、遺伝子形またはタンパク質形の標的の全てまたは一部により決定される分子、例えば、抗体、模倣体またはアンチセンス分子を含む。
【0134】
この点に関しては、抗イディオタイプ抗体、キメラ抗体およびヒト化抗体を含む抗体が有用であり、現在、それらの作製は、当業者には日常的である。ペプチドまたは非ペプチド模倣体は、機能のために重要である標的分子の残基を同定することにより、標的の作動薬として開発することができる。標的分子と相互作用し、改善された薬理学的特性を有する分子を設計するために、モデリングを使用することができる。全てのそのような分子は、細胞に進入できるように改変する必要があるだろう。
【0135】
合理的な創薬により、対象となる生物活性ポリペプチドのまたはそれらが相互作用する小分子(例えば、作動薬、拮抗薬、阻害剤またはエンハンサー)の構造類似体を産生させて、例えばポリペプチドのさらに活性もしくは安定な形態である薬物、または例えばポリペプチドの機能を強化するか、ポリペプチドの機能に干渉する薬物を形成することができる。例えば、Hodgson(Bio/Technology 9: 19-21, 1991)を参照されたい。一つのアプローチでは、X線結晶学により、コンピュータモデリングにより、または最も典型的にはアプローチの併用により、対象となるタンパク質の三次元構造を、先ず、決定する。ポリペプチドの構造に関する有用な情報は、相同タンパク質の構造に基づくモデリングによっても得ることができる。合理的な創薬の一例は、HIVプロテアーゼ阻害剤の開発である(Erickson et al., Science 249 : 527-533,1990)。加えて、アラニンスキャンによって標的分子を分析することができる(Wells, Methods Enzymol. 202 : 2699-2705, 1991)。この技法では、アミノ酸残基をAlaにより置換し、ペプチド活性に対するその影響を判定する。そのペプチドの各アミノ酸残基をこの要領で分析して、そのペプチドの重要な領域を決定する。
【0136】
機能的アッセイにより選択した標的特異的抗体を単離すること、およびその後、その結晶構造を解明することも可能である。原則として、このアプローチは、その後の創薬の基礎となり得るファーマコア(Pharmacore)を生じさせる。機能的薬理活性抗体に対する抗イディオタイプ抗体(抗id)を作製することにより、タンパク質結晶学を全て回避することができる。鏡像の鏡像として、抗idの結合部位は、元の受容体の類似体であると予想されよう。その後、その抗idを使用して、化学的または生物学的に産生させたペプチドバンクのバンクからペプチドを同定および単離することができる。その後、選択されたペプチドはファーマコアとしての機能を果たすであろう。
【0137】
本明細書において考えられる類似体は、側鎖の改変;ペプチド、ポリペプチドまたはタンパク質合成中の非天然アミノ酸および/またはそれらの誘導体の組み込み;架橋剤の使用;ならびにタンパク質性分子またはそれらの類似体に配座的拘束を負わせる他の方法を含むが、これらに限定されるわけではない。
【0138】
本発明により考えられる側鎖改変の例としては、アルデヒドとの反応による還元アルキル化、その後のNaBH4での還元;メチルアセトイミデートでのアミジン化;無水酢酸でのアシル化;シアネートでのアミノ基のカルバモイル化;2,4,6-トリニトロベンゼンスルホン酸(TNBS)でのアミノ基のトリニトロベンジル化;無水コハク酸および無水テトラヒドロフタル酸でのアミノ基のアシル化;ならびにピリドキサル-5-リン酸でのリジンのピリドキシル化、その後のNaBH4での還元などによる、アミノ基の改変が挙げられる。
【0139】
アルギニン残基のグアニジン基は、2,3-ブタンジオン、フェニルグリオキサールおよびグリオキサールなどの試薬での複素環式縮合生成物の形成によって改変することができる。
【0140】
カルボキシル基は、O-アシルイソウレア形成によるカルボジイミド活性化、その後の、例えば対応するアミドへの、二次的誘導体化によって改変することができる。
【0141】
スルフヒドリル基は、ヨード酢酸またはヨードアセトアミドでのカルボキシメチル化;システイン酸への過ギ酸酸化;他のチオール化合物との混合ジスルフィドの形成;マレイミド、無水マレイン酸または他の置換マレイミドとの反応;4-クロロメルクリベンゾエート、4-クロロメルクリフェニルスルホン酸、フェニルメルクリクロライド、2-クロロメルクリ-4-ニトロフェノールおよび他の水銀剤を使用する水銀性誘導体の形成;アルカリ性pHでのシアネートでのカルバモイル化などの方法によって改変することができる。
【0142】
トリプトファン残基は、例えば、N-ブロモスクシンイミドでの酸化または2-ヒドロキシ-5-ニトロベンジルブロマイドもしくはスルフェニルハライドでのインドール環のアルキル化によって改変することができる。一方、チロシン残基は、テトラニトロメタンでニトロ化して3-ニトロチロシン誘導体を形成することによって変性することができる。
【0143】
ヒスチジン残基のイミダゾール環の改変は、ヨード酢酸誘導体でのアルキル化またはジエチルピロカルボネートでのN-カルボエトキシル化によって達成することができる。
【0144】
ペプチド合成中に非天然アミノ酸および誘導体を組み込む例としては、ノルロイシン、4-アミノ酪酸、4-アミノ-3-ヒドロキシ-5-フェニルペンタン酸、6-アミノへキサン酸、t-ブチルグリシン、ノルバリン、フェニルグリシン、オルニチン、サルコシン、4-アミノ-3-ヒドロキシ-6-メチルヘプタン酸、2-チエニルアラニンおよび/またはアミノ酸のD異性体の使用が挙げられるが、これらに限定されるわけではない。本明細書で考えられる非天然アミノ酸のリストを表4に示す。
【0145】
(表4)非典型アミノ酸についてのコード
【0146】
例えば3D配座を安定させるために、架橋剤を使用することができ、例えば、n=1からn=6の(CH2)nスペーサー基を有する二官能性イミドエステル、グルタルアルデヒド、N-ヒドロキシスクシンイミドエステルなどの二官能性架橋剤、ならびにアミノ反応性部分(例えば、N-ヒドロキシスクシンイミド)および別の基に特異的な反応性部分(例えば、マレイミド、またはジチオ部分(SH)またはカルボジイミド(COOH))を通常含有するヘテロ二官能性試薬を使用する。加えて、例えば、CαおよびNα-メチルアミノ酸の組み込み、およびアミノ酸のCα原子とCβ原子の間への二重結合の導入により、ペプチドを配座的に拘束することができる。
【0147】
天然産物、コンビナトリアルまたはファージ提示法は、全て、調節成分をスクリーニングするために利用することができる。ハイスループットスクリーニング法の大量選択を利用することができ、これに本発明の細胞を利用できるようにすることができる。
【0148】
Blimp-1で作動する遺伝子ネットワークの他のメンバーを同定する際、ツーハイブリッドスクリーニングも有用である。標的相互作用および調節成分についてのスクリーニングは、2つの物理的に分けることができる機能的ドメインからなる転写因子を利用する酵母ツーハイブリッド系を使用して行うことができる。最も一般的に使用されるのは、DNA結合ドメインおよび転写活性化ドメインからなる酵母GAL4転写活性化因子である。2つの異なるクローニングベクターを使用して、潜在的結合性タンパク質をコードする遺伝子へのGAL4ドメインの別個の融合体を作製する。これらの融合タンパク質は共発現され、核を標的にし、ならびに相互作用が発生する場合にはレポーター遺伝子(例えば、lacZ)の活性化によって検出可能な表現型が生じる。この場合、例えば、S.セレビジエ(S.cerevisiae)を、cDNA GAL4活性化ドメイン融合体を発現するライブラリまたはベクター、およびGAL4に融合したMyb経路構成要素を発現するベクターで共形質転換する。lacZをレポーター遺伝子として使用する場合、融合タンパク質の共発現により青色が生じるであろう。標的と相互作用する小分子または他の候補化合物は、結果として細胞の色の喪失をもたらすであろう。Munderら(Appl. Microbiol. Biotechnol. 52(3): 311-320, 1999)およびYoungら(Nat. Biotechnol. 16(10): 946-950, 1998)が開示したような酵母ツーハイブリッド系を参照することができる。この系によってこのように同定された分子を、その後、本発明の遺伝子改変生物または遺伝子改変細胞において再検査する。
【0149】
組換え、合成または単離された形態の本作動薬および拮抗薬ならびに1つまたは複数の薬学的に許容される担体、希釈剤または賦形剤を含む治療用薬学的組成物が、さらに企図される。癌の治療または免疫応答の調節が特に企図される。
【0150】
治療法という用語は、ある状態または疾病の治療または予防への言及と解釈するものとする。用語「治療する」および「改善する」は、交換可能に用いている。
【0151】
用語「組成物」または「薬剤」または「薬物」は、所望の薬理および/または生理作用を誘導する化学的化合物を指す。この用語は、塩、エステル、アミド、プロドラッグ、活性代謝産物、類似体などを含むが、これらに限定されるわけではない、本明細書において特に述べる化合物の薬学的に許容されるおよび薬理学的に活性な成分も含む。上の用語を用いるときには、それが、活性薬剤それ自体ならびに薬学的に許容される、薬理学的に活性な塩、エステル、アミド、プロドラッグ、代謝産物、類似体などを含むと理解しなければならない。用語「化合物」は、狭く解釈するべきではなく、ペプチド、ポリペプチドおよびタンパク質ならびに遺伝分子、例えばRNA、DNA、およびその模倣体および化学的類似体まで拡大する。
【0152】
フレーズ「疾病もしくは状態を改善する」または「治療」または「治療的」は、最も広い状況で用いており、疾病もしくは状態、または疾病もしくは状態の1つまたは複数の症状、または疾病もしくは状態の症状の頻度におけるあらゆる測定可能なまたは実質的に有意な改善、ならびにその疾病からの完全回復または状態、その症状もしくはその基礎原因の除去を含む。本発明は、疾病または状態の大きな範囲に応用でき、担当技術者は、症例ごとベースで表現型の評価の正確なパラメータを決定しなければならない。状態は、1つまたは複数の疾病に随伴することもあり、またはそのように関連していないこともある。状態の改善は、あらゆる所望の生理学的または心理学的変化を包含する。
【0153】
本組成物の有効量は、当業者により最良に確立される。本明細書において用いる場合の化合物の「有効量」という用語は、所望の治療または生理効果を生じさせるために十分な薬剤の量を意味する。時には望ましくない効果、例えば副作用が、所望の治療効果とともに現れる。それ故、担当者は、何が適切な「有効量」であるかを判定する際、潜在的利益と潜在的リスクとのバランスをとる。必要とされる正確な量は、被験者の種、年齢および全身の健康状態、投与方式などに依存して、被験者ごとに変わるであろう。従って、正確な「有効量」を特定することができないこともある。しかし、いずれの個々の症例においても、適切な「有効量」は、当業者が日常的な経験のみを利用して決定することができる。
【0154】
組換え、合成または単離形態の本発明の作動薬および拮抗薬、ならびに1つまたは複数の薬学的に許容される担体、希釈剤または賦形剤を含む治療用薬学的組成物が、さらに企図される。癌の治療または免疫応答の調節が、特に企図される。
【0155】
治療法という用語は、ある状態または疾病の治療または予防への言及と解釈するものとする。用語「治療する」および「改善する」は、交換可能に用いている。
【0156】
用語「組成物」または「薬剤」または「薬物」は、所望の薬理および/または生理作用を誘導する化学的化合物を指す。この用語は、塩、エステル、アミド、プロドラッグ、活性代謝産物、類似体などを含むが、これらに限定されるわけではない、本明細書において特に述べる化合物の薬学的に許容されるおよび薬理学的に活性な成分も包含する。上記の用語を用いるときには、それが、活性薬剤それ自体ならびに薬学的に許容される、薬理学的に活性な塩、エステル、アミド、プロドラッグ、代謝産物、類似体などを含むと理解しなければならない。用語「化合物」は、狭く解釈するべきではなく、ペプチド、ポリペプチドおよびタンパク質ならびに遺伝分子、例えばRNA、DNA、およびその模倣体および化学的類似体まで拡大する。
【0157】
フレーズ「疾病もしくは状態を改善する」または「治療」または「治療的」は、最も広い状況で用いており、疾病もしくは状態または疾病もしくは状態の1つまたは複数の症状または疾病もしくは状態の症状の頻度におけるあらゆる測定可能なまたは実質的に有意な改善、ならびにその疾病からの完全回復または状態、その症状もしくはその基礎原因の除去を含む。本発明は、疾病または状態の大きな範囲に応用でき、担当技術者は、症例ごとベースで表現型の評価の正確なパラメータを決定しなければならない。状態は、1つまたは複数の疾病に随伴することもあり、またはそのように関連していいこともある。状態の改善は、あらゆる所望の生理学的または心理学的変化を包含する。
【0158】
本組成物の有効量は、当業者により最良に確立される。本明細書において用いる場合の化合物の「有効量」という用語は、所望の治療または生理効果を生じさせるために十分な薬剤の量を意味する。時には望ましくない効果、例えば副作用が、所望の治療効果とともに現れる。それ故、担当者は、何が適切な「有効量」であるかを判定する際、潜在的利益と潜在的リスクのバランスをとる。必要とされる正確な量は、被験者の種、年齢および全身の健康状態、投与方式などに依存して、被験者ごとに変わるであろう。従って、正確な「有効量」を特定することができないこともある。しかし、いずれの個々の症例においても、適切な「有効量」は、当業者が日常的な経験のみを利用して決定することができる。
【0159】
本発明のポリペプチド、核酸、抗体、ペプチド、化学的類似体、作動薬、拮抗薬または模倣体は、薬学的組成物に調合することができ、薬学的組成物は、従来の薬学的調剤法に従って調製される。例えば、Remington's Pharmaceutical Sciences, 18th Ed. (1990, Mack Publishing, Company, Easton, PA, U.S.A.を)参照されたい。組成物は、活性薬剤または活性薬剤の薬学的に許容される塩を含有することができる。これらの組成物は、活性物質の1つに加えて、薬学的に許容される賦形剤、担体、緩衝液、安定剤または当技術分野において周知の他の材料を含むことができる。そのような材料は、非毒性でなければならず、活性成分の有効度に干渉してはならない。担体は、投与、例えば、静脈内投与、経口投与、髄腔内投与または非経口投与、に望ましい製剤の形態に依存して多種多様な形態を取ることができる。
【0160】
経口投与用の化合物は、固体または液体製剤、例えばカプセル、ピル、錠剤、ロゼンジ、粉末、懸濁液または乳剤に調合することができる。経口剤形の組成物を調製する場合、通常の製薬用媒体、例えば、経口液体製剤(例えば、懸濁液、エリキシルおよび溶液など)の場合には、例えば、水、グリコール、油、アルコール、着香剤、保存料、着色剤、懸濁化剤など;または経口固体製剤(例えば、粉末、カプセルおよび錠剤など)の場合には、デンプン、糖、希釈剤、顆粒化剤、滑沢剤、結合剤、崩壊剤など、のいずれを利用してもよい。投与の容易さのため、錠剤およびカプセルが、最も有利な経口単位剤形の代表であり、この場合、固体の製薬用担体が、明らかに利用される。所望される場合、標準的な技法により錠剤に糖衣または腸溶コーチングを施すことができる。活性薬剤をカプセル化して、それを安定化して胃腸管を通過させ、一方、それと同時に、血液脳関門を通過できるようにすることができる。例えば、国際特許公開公報第96/11698号を参照されたい。
【0161】
非経口投与用の化合物は、製薬用担体に溶解し、溶液または懸濁液のいずれかとして投与することができる。実例となる適する担体は、水、食塩水、デキストロース溶液、フルクトース溶液、エタノール、または動物、植物もしくは合成起源の油である。担体は、他の成分、例えば、保存料、懸濁化剤、可溶化剤、緩衝液なども含有することができる。化合物を髄腔内投与する場合、それらを髄液に溶解することもできる。
【0162】
活性薬剤は、好ましくは、治療有効量で投与される。実際の投与量ならびに投与の速度および時間経過は、治療する状態の性質および重症度に依存するであろう。治療の処方、例えば、投薬量の決定、タイミングなどは、一般担当者または専門家の責任の範囲内であり、典型的には治療すべき疾患、個々の患者の状態、送達部位、投与方法および担当者には公知の他の因子を考慮に入れる。技法およびプロトコルの例は、Remington's Pharmaceutical Sciences,(前記)において見出すことができる。
【0163】
または、抗体もしくは細胞特異的リガンドなどのターゲティング系の使用によるターゲティング療法を用いて、一定の細胞タイプにさらに特異的に活性薬剤を送達することができる。ターゲティングは、様々な理由のため、例えば、その薬剤が許容できないほど毒性である場合、またはそれがあまりにも高い投薬量を別様に必要とする場合、またはそれが別様に標的細胞に侵入することができない場合、望ましいであろう。
【0164】
これらの薬剤を直接投与する代わりに、標的細胞において、例えば、上記のものなどのウイルスベクターにおいて、または米国特許第5,550,050号ならびに国際特許公開公報第92/19195号、同第94/25503号、同第95/01203号、同第95/05452号、同第96/02286号、同第96/02646号、同第96/40871号、同第96/40959号および同第97/12635号に記載されているものなどの細胞ベースの送達系において産生させることができる。ベクターを標的細胞にターゲティングし、発現産物の発現を特定の細胞、発達段階または細胞周期段階に限定することができるだろう。細胞ベースの送達系は、患者体内の所望の標的部位に移植するために設計され、ならびに標的薬剤のコード配列を含有する。または、治療すべき細胞において産生されたまたは治療すべき細胞にターゲティングされた活性化薬剤により活性形態に転化させるために、薬剤を前駆体形で投与することもできる。例えば、欧州特許出願第0 425 731A号および国際特許公開公報第90/07936号を参照されたい。
【0165】
以下の非限定的実施例により、本発明をさらに説明する。
【実施例】
【0166】
実施例1
Blimp-1突然変異対立遺伝子(Blimpgfp)の作製
イントロン3’からエキソン6に、スプライス受容体、3つ全てのリーディングフレーム内の停止コドン、内因性リボソーム認識部位(IRES)、eGFPをコードするcDNA、および転写を停止させるSV40ポリアデニル化シグナルからなるeGFP発現カセットが挿入されている、Blimp-1ターゲティング構築物を産生させた。組み込まれたターゲティングベクターでの胚性幹(ES)細胞の選択を可能にするために、PGK-Neoγ遺伝子もそのイントロンに挿入されている。それらのeGFPカセットおよびNeoγカセットをFrt部位に隣接させて、挿入されたDNAのflpリコンビナーゼ媒介欠失を可能にする。Blimp-1ターゲティング構築物、G418により選択される耐性クローンを用いてC57BL/6 ES細胞をエレクトロポレーションし、5’および3’ゲノムDNAプローブへのサザンハイブリダイゼーションによりスクリーニングした(図1C)。Blimpgfp対立遺伝子を有する4つの正確にターゲティングされたクローン(図1C)を、300個のスクリーニングされたクローンから同定した。これらをBALB/c胚盤胞に注入して、キメラ創始者を得た。これらのキメラを増殖させ、生殖系伝達を1つのクローン(4F3)で達成した。
【0167】
実施例2
Blimp-1の発現パターンの記述を可能にするGFPレポーター
当初、Blimp-1は、ASC分化を受けるように誘導されたBリンパ球においてのみ発現されると報告された(Turner et al., (前記))。しかし、その後の研究により、胚形成中(Chang et al., Mech Dev 117:305, 2002)、および骨髄細胞における(Chang et al., (前記), 2000)Blimp-1のより幅広い発現パターンが明らかになった。Blimpgfp対立遺伝子により、造血系列の中でも、より広くは生物においても、Blimp-1の発現パターンをより完全に定義することができる。上で概説したターゲティング戦略は、結果としてBlimpgfp対立遺伝子を生じさせ、それが、内因性Blimp-1調節要素の制御下で2シストロンのmRNAからGFPを発現し、従って、完全Blimp-1発現パターンを反復すると予測される。加えて、この戦略は、Blimp-1 mRNA転写産物を分断して、DNA結合モチーフを含有するジンクフィンガードメインを欠くトランケートされたバージョンのBlimp-1タンパク質(エキソン1〜6)を産生させる。これに従って、インビトロでLPSを用いて分化するように誘導されたBlimpgfp/+ B細胞のウエスタンブロッティングは、野生型Blimp-1タンパク質バンドおよびトランケート型Blimp-1タンパク質バンドの両方を示した(図1D)。生細胞におけるGFP発現および固定された組織におけるBlimp-1タンパク質のモニタリングにより、インビボおよびインビトロでのBリンパ球の遺伝子活性および分化の運命を単細胞レベルでモニターすることができる。
【0168】
実施例3
ASCにおけるBlimp-1のインビボ発現
Blimpgfp/+マウスにおけるリンパ組織の検査は、高Blimp-1の小集団が、骨髄(0.1〜0.2%)、脾臓(0.4〜0.6%)およびリンパ節(0.1%)において細胞を発現していることを示した(図2)。さらに、GFP+細胞の表現型解析は、それらが、以前に定義された高Synd-1/低B220のASC集団、ならびに以前に十分に特性付けされていなかった低Synd-1〜Synd-1陰性の表現型を表すことを示した(図2、(Underhill et al., (前記))。これらの細胞がASCであることを確認するために、Blimpgfp/+骨髄および脾臓からGFP+細胞を選別し、Ig産生についてのELISpot分析に供した。図3からわかるように、細胞の75〜100%がIg分泌性細胞であった(3回の独立した実験の代表)。さらに、GFP陰性画分の選別は、0.001%のASCの頻度(細胞100,000個あたり1個未満)、これに対して、未選別骨髄におけるこれらの細胞の頻度は、0.05〜0.09%であった(100,000個あたり50〜90個)。従って、Blimpgfp発現性ASCの単離は、未分別細胞の100,000倍の濃縮をもたらし、これらの希少細胞を単離するための実質的に決定的な方法をもたらす。加えて、GFP+ ASC集団における全てのIgアイソタイプを表した(図3)。
【0169】
GFPレポーターマウスを使用してBlimpgfp/+マウスにおけるASCの産生をさらに検査するために、2μgのリポ多糖類(LPS)を静脈内注射し、注射後1〜7日のGFP発現について分析した(LPS注射は、結果として成熟B細胞の多クローン性活性化を生じさせる)。LPS注射は、結果として、注射後3日目にピークに達する脾臓GFP+細胞の数の劇的な増加を、その後、7日周辺に定常状態レベルへの低下を生じさせる(図4A)。ゲートされたGFP+細胞の分析は、ASC分化が、Synd-1+/B220+細胞の出現、続いてSind-1+/B220-の出現に伴い同時波で発生したこと、および最終的に、ASCの画分が、Synd-1-/B220-になることを示した。この分化は、骨髄においても反映され、この場合、GFP+細胞は、Synd-1+/-/B220-定常状態集団を迅速に生じさせるSynd-1+/B220+細胞として、注射後4日目に出現する(図4B)。要約すると、LPS治療は、Blimpgfp/+マウス株を使用して初めて表現型的に定義することができるASC分化の波を誘導する。
【0170】
実施例4
インビトロで誘導されるASCにおけるBlimp-1の発現
ASC系列への決定付けおよびASC系列までの進行に影響を及ぼすパラメータをインビトロで定量分析する方法論を開発した。このシステムは、パーコール(Percoll)勾配遠心分離および磁性ビーズ濃縮により精製し、B細胞増殖およびASCへの分化を誘導する様々な刺激の存在下で培養する、小さな休止B細胞の単離を含む。これらの条件としては、IL-4および抗CD40を使用するT依存性応答の模倣またはLPSを使用するT非依存性反応の模倣が挙げられる。加えて、IL-5を濃度調節しながらこれらの培養物に滴下して分化速度を加速し、および抗IgD(1.19)架橋を行って、抗原特異的応答を活性化することができる。1〜5日目にフローサイトメトリーにより培養物を分析して、BlimpgfpおよびSynd-1+発現性細胞の頻度を測定した。培養物中のASCの数は、ELIspotにより判定した。
【0171】
CD40L/IL-4/IL-5またはLPSを使用するBlimpgfp誘導の時間経過の分析(図5A)は、最初のGFP+細胞が、2日後にLPS培養物において観察されることを示した。その後、4日目における約50%GFP+細胞のピークまで、陽性細胞の数は増加する。対照的に、CD40L/IL-4/IL-5治療は、結果として、より少数のGFP発現性細胞の遅発性誘導を生じさせる。興味深いことに、CD40L/IL-4/IL-5誘導GFP発現性細胞の大部分が、Synd-1+でもあるのに対し、LPSは、Synd-1+とSynd-1-両方のGFP発現性細胞を誘導する(図5B)。全てのGFP+細胞が、実際にASCであるかを判定するために、LPS治療BlimpgfpB細胞からの4つの画分を選別した(図5C)。IgM、IgG3およびIgG2bについてのELIspotアッセイは、培養物中の全てのASCが、GFP+画分(A、B、C)によって表され、Synd-1発現レベルと相関しないことを明確に示す。さらに、ASCの頻度は、GFP+集団間で変わらなかった(図5C)。従って、これらのデータは、Blimpgfpが、ASCの運命のマーカーである一方で、Synd-1+が、インビボと同様にインビトロでもASC活性の亜集団の単なる指標であることを明確に示している(図2)。BlimpgfpおよびSynd-1発現の調節を、上で概説した様々な刺激を使用して検査した。興味深いことに、図5Cに概略を示すように、GFP+/Synd-1+およびGFP+/Synd-1- ASC形成の頻度は、異なる刺激により調節され得る。従って、Blimpgfp発現に続いて、インビトロおよびインビボでのASC誘導の調節を同定するための簡単で決定的な方法論をもたらす。
【0172】
最後に、インビトロ由来Blimpgfp陽性ASCを、静脈内注射の7日後に放射線未照射宿主の骨髄および脾臓において検出することができる(図6)。従って、Blimpgfp対立遺伝子は、インビボでの生存、移動および機能特性に対するASCのインビトロ治療の効果を検査する方法をもたらす。
【0173】
実施例5
Blimp-1は胚形成に必要とされる
ホモ接合のBlimpgfp/gfp動物を産生するために、Blimpgfp/+個体を交雑させた。Blimp-1野生型およびBlimpgfp特異的PCRプライマーを使用して、これらの交雑からの子孫を生後21日目に遺伝子型解析した。Blimpgfp/+マウスが、生存しており健康であったのに対し、Blimpgfp/gfp個体は、同定されなかった。これは、Blimp-1欠失が、結果として胚性致死または早期産後致死を生じさせることを示している(図7)。Blimpgfp/gfp動物が死亡する段階により近い段階を検査するために、Blimpgfp/+マウスの時限交雑から生じた胚を検査した。これらのデータは、Blimpgfp/gfp胚が、胚性段階E15.5ほどもの後期に生存していることを示す。しかし、生存能力がある、より高い齢の個体が、文献に記載されたことはない。Blimp-1が胚形成中に広範に発現されることは公知であり、この発見は、Blimpgfpマウスを使用する分析により支持されている。
【0174】
実施例6
Blimp-1は、抗体産生に不可欠である
Blimpgfp/gfp動物の胚性致死を回避するために、および抗体産生におけるBlimp-1の重要性を直接検査するために、致死量の放射線を照射した同系マウスの胎仔肝臓幹細胞再構成を用いて、造血形全体にわたって機能的なBlimp-1タンパク質を欠く成体マウスを産生させた。これらのBlimpgfp/gfpキメラ動物は、健康であり、検査した全ての造血細胞系列を比較的正常な数、含有している。LPSまたはCD40L/IL-4およびIL-5、いずれかでの刺激後のこれらのマウスにおけるASC集団のインビトロ分析は、主としてsynd-1+であるGFP+ Blimp欠失細胞の存在を示した(図8A)。重要なことに、ELIspotアッセイにより評価したところ、これらの細胞は、抗体を分泌できなかった(図8B)。従って、Blimpgfp、ここに記載するマウスモデルは、ASCを単離するための決定的ツールをもたらすばかりでなく、ホモ接合突然変異Blimpgfp/gfp脾細胞からのBlimp-1発現性細胞の集団の同定を可能にし、それにより、Blimp-1欠失の表現型の根底にあるメカニズムの分析を大いに助長する。
【0175】
実施例7
他の造血系列におけるBlimp-1の発現
Blimpgfpレポーター系により、初めて、造血におけるBlimp-1の発現パターンも定義することができた。上で述べたように、Blimpgfpマウスのリンパ器官の分析は、GFP高産生性集団が、ほぼ排他的にASCであることを示した。しかし、低レベルのGFP産生性細胞も出現した。
【0176】
Blimp-1は、ヒトおよびマウスマクロファージならびに顆粒球により発現されることが報告されている。MCSF-1の存在下での血液単球細胞および骨髄由来マクロファージ培養物のフローサイトメトリー分析は、これらの細胞タイプにおける明確なBlimpgfp発現を示した(図9)。しかし、GFP蛍光は、顆粒球では観察されなかった。対照的に、インビボで単離された樹状細胞は、Blimp-1 mRNA発現を欠く。同様に、flt3Lを有する骨髄細胞の培養から誘導された形質細胞性および従来の樹状細胞は、Blimpgfp蛍光を欠く。しかし、選別された樹状細胞のエクスビボ活性化、またはCpG DNAによるflt3L培養物のインビトロ活性化は、結果として、Blimp-1発現、主として従来の樹状細胞を生じさせた(図10)
【0177】
Blimpgfp/+マウスからの胸腺および休止脾臓の分析は、Blimp-1が、T細胞発達中に発現されないことを実証した。しかし、Blimpgfp発現性T細胞の小集団が、リンパ節に存在した。これらの細胞は、存在する活性化T細胞の小集団を提示する可能性があったため、本発明者らは、コンカナバリンA(T細胞を強力に活性化することが公知である条件)の存在下または不在下、インビトロで、リンパ節T細胞を抗CD3モノクローナル抗体で刺激した。インビトロ活性化T細胞は、Blimpgfpを発現し、インビボ分析を支持した(図12C)。
【0178】
BlimpgfpマウスにおけるNK系列の検査は、検査した他の造血系列とは異なり、NK細胞が、Blimp-1を構成的に発現することを示した。NK細胞は、血液、脾臓および骨髄からNK1.1+/CD122+細胞として同定され、均一にGFP+であることを示した(図12A)。この発現は、IL-15の存在下で培養された成熟NK細胞がGFP+であるためインビトロで維持され、かつ、NK細胞の最終分化を誘導するサイトカイン、例えばIL-21またはIL-12/IL-18によってさらに誘導することができる(図12B)。NK細胞におけるBlimp-1の発現は、Blimp-1特異的モノクローナル抗体でのウエスタンブロッティングによっても確認した。
【0179】
要約すると、Blimpgfpレポーターマウスは、様々な造血系列の後期においてBlimp-1を誘導することが明らかになり、その結果、これらの細胞タイプの成熟の調節因子を同定する方法が得られた。重要なことに、非Bリンパ系細胞タイプにおける比較的低い産生レベルのGFPは、均一ASC集団の単離に干渉しない。
【0180】
実施例8
Blimpgfpマウスを使用する、Blimp-1の癌における役割の検査
ASC分化の検査におけるその有用性に加えて、Blimpgfpレポーターマウスは、この細胞タイプの悪性トランスフォーメーションを検査するために使用することができる。マウスでは形質細胞種およびヒトでは多発性骨髄腫と呼ばれるASCの腫瘍は、IgHイントロン性エンハンサーの制御下、B細胞系列においてv-abl癌遺伝子を発現するEμ-v-ablトランスジェニックマウス(Rosenbaum et al., (前記))において、特異的におよび頻繁に惹起される。これらのマウスをBlimpgfp突然変異マウスと交雑させて、腫瘍の潜在性および発病率に対するBlimp-1遺伝子の一方または両方のコピーの欠失の影響を判定した。2つの結果が考えられる:Blimp-1は、形質細胞分化プログラムを誘導することにより、v-ablトランスフォーメーションの機会に関するウインドウを開くために必要となり得る。このトランスジーンは、初期B細胞における発現にもかかわらず、形質細胞種しか誘導しない(Rosenbaum et al., (前記))。従って、機能的Blimp-1対立遺伝子の欠失により、腫瘍発病率の低下または潜在性の増加が予測されよう。または、大きな比率のv-abl誘導形質細胞種が、再配列されたおよび活性化したc-myc遺伝子も有するので、Mycは、トランスフォーメーションにおける本質的な共同作用性活性であり得る(Rosenbaum et al., (前記))。Blimp-1は、最終ASC分化の間のc-myc発現を抑制すると一般に考えられている(Lin et al., Science 276:596, 1997)。このシナリオでは、機能的Blimp-1の欠失が、持続性c-myc発現を可能にすることとなり、それにより、形質細胞種の発現が加速し得る。
【0181】
Blimp-1が、実際、ASC腫瘍形成において一定の役割を果たすならば、それ故、Blimpgfpレポーター株は、腫瘍進行に対するBlimp-1阻害/誘導の効果を判定するための有用な動物モデルをもたらす。
【0182】
実施例9
T細胞における最終分化の調節におけるBlimpの役割を評価する方法
マウス
Blimpgfpマウス(Kallies et al., J Exp Med 200:967-977, 2004)、Rag1-/-マウスおよびRag2-/-マウスを、C57BL/6バックグラウンドで保持した。Blimpgfp遺伝子型解析および胎仔肝臓キメラを、記載されている(Kallies et al., 2004 (前記))とおり作製した。
【0183】
フローサイトメトリーおよびELISA
CD4(GK1.5)、CD8(53.6.7)、TCRβ(H57-597)、Ly5.2(ALI-4A2)に対するモノクローナル抗体(mAb)を、プロテインG-セファロースカラム(Amersham Pharmacia Biotech)でハイブリドーマ上清から精製し、ビオチン(Pierce Chemical Company)、アロフィコシアニン(APC)およびフィコエリトリン(PE)(ProZyme)に、推奨どおりにコンジュゲートさせた。ビオチン化抗CD25(7D4)およびCD122(Tm-β1)ならびにPEコンジュゲート型抗CD44(IM7)、CD62L(MEL-14)、IFNγ.□□□〜1□.、IL-4(11B11)は、PharMingenから入手した。ビオチン化mAbは、ストレプトアビジン-PEまたはCy5と共に発現された(Southern Biotechnologies Inc.)。LSRサイトメーター(BD Bioscience)で細胞を分析し、高速フローサイトメーター(Moflo cytomation and BD Biosciences)で細胞選別を行った。サイトカインについての細胞内染色は、当技術分野において公知の標準的な手順に従って行った。IFNγおよびIL-10産生についてのELISAは、記載されている(Brady et al., J Immunol 172: 2048-2058, 2004)とおり行った。IL-4 ELISAは、1つのモノクローナル抗体を捕捉試薬として使用し、第二のモノクローナル抗体を検出に使用した。ELISAは、三回重複で行い、組換えタンパク質標準物質を使用して定量した。
【0184】
ウエスタンブロッティング
例えば、Rosenbauerら(Embo J 21:211-20, 2002)が記載したように、全タンパク質抽出物を相当数の細胞から産生させ、ウエスタンブロッティングを行った。抗Blimp-1 mAbは、以前に記載されている(Kallies et al., 2004 (前記))。抗Vav1を使用して、等量のタンパク質負荷を確認した。
【0185】
HSV感染
20μLのPBSで希釈した4×105単純疱疹ウイルス(HSV-1 KOS株)にマウスを感染させた。後ろ足の足蹠と踵の間への皮下注射によって投与した。感染したマウスの脾臓および膝窩リンパ節を、その後、分析のために回収した。
【0186】
インビトロ細胞毒性
gB特異的細胞毒性T細胞リンパ球(CTL)を常用の手順[Belz, 2001]により作製した。感染したマウスから脾臓を除去し、108個の1000Gy照射gB498-505被覆C57BL/6脾細胞を5日間単細胞培養した。細胞毒性は、従来の51Cr放出アッセイで評価した。EL4(H-2b)標的細胞を、Na51Crで1時間標識し、gBペプチドを60分間適用し、2回洗浄し、5,000標的/ウエルでプレーティングした。その後、それらをエフェクター集団と共に5時間インキュベートし、その後、γ計数用に上清を回収した。二倍リンパ球希釈物を三回重複でアッセイし、一方、未処理およびTriton X-100破壊対照は、四回重複で測定した。特異的溶解率は、100×(エフェクターを伴う標的からの51Cr放出-標的のみからの51Cr放出)/(Triton X-100を伴う標的からの51Cr放出)として計算した。T細胞不在下でインキュベートした標的からの51Cr放出レベルは、全Triton X-100媒介51Cr放出の10%未満であった。
【0187】
gB特異的CD8+ T細胞の四量体染色
H-2Kb糖タンパク質と単純疱疹ウイルスの糖タンパクB(gB498-505)由来のペプチド(SSIEFARL)とのMHCクラスI四量体型複合体[Altman, 1996 #102; Allan, 2003 #100]を使用して、ウイルス特異的CD8+ T細胞を同定した。カルボキシ末端膜貫通ドメインがbirAビオチン化モチーフで置換されている組換えH-2Kb分子を、ヒトβ2-ミクログロブリンおよびウイルスペプチドでリフォールディングし、birAでビオチン化し、4:1のモル比でニュートラビジン(neutravidin)-PE(オレゴン州、Eugene、Molecular Probes)と複合させた。室温で、60分間、リンパ球をPBS/BSA/アジド中の四量体型複合体について染色し、その後、抗CD8αAPCで染色し、2回洗浄し、フローサイトメトリーによって分析した。
【0188】
組織学
器官を10%緩衝ホルマリン中で固定し、パラフィンに包埋し、薄片に切り、ヘマトキシリン/エオシンで染色した。
【0189】
実施例10
Blimp発現は、IL-21により誘導することができ、ナイーブCD4+およびCD8+ T細胞の活性化エフェクター細胞への成熟に関する重要な成分であり、正常なリンパ球恒常性に不可欠である
【0190】
Blimpgfp/+マウスからのリンパ器官の検査は、形質細胞において高レベルのGFPを示した。さらに、T細胞集団は、低レベルのGFPを発現した(図19A)。機能的Blimp対立遺伝子を発現しない、Blimpgfp/gfp細胞で再構成されたRag1-/-マウスのさらなる分析は、この同じ集団の顕著な増幅およびGFP蛍光の有意な増加を示した(図20)。これらのマウスのT細胞区画のより広範なフローサイトメトリー分析は、胸腺細胞(データは示していない)およびナイーブT細胞にはGFP蛍光がないことを示した(図20)。
【0191】
対照的に、および本明細書において示すように、Blimp-1は、活性化/記憶タイプCD4+(TCRβ+CD62Llow)およびCD8+(TCRβ+CD44high)T細胞において有意に発現される(図20)。より詳細なFACS分析では、代替マーカーを使用して、Blimp-1欠失マウスにおいて、増幅されたT細胞プールの性質を判定した。野生型胎仔肝臓で再構成されたマウスとは対照的に、Blimpgfp/gfp細胞で再構成されたRag1-/-マウスは、低CD62L、GFP陽性のT細胞集団の強い増幅を示した。さらに、これらのマウスは、低CD27〜CD27陰性のT細胞の数の強い上昇、CD25陽性CD4細胞の増加ならびに明瞭な高CD122集団の欠如を有した。代替マーカー、例えばCD69を使用する分析により、GFP+細胞の活性化状態を確認した(データは示していない)。Blimpgfp/gfpで再構成されたRag1-/-マウスにおけるエフェクターCD4 T細胞数の増加を、単離脾臓CD4 T細胞のエクスビボ刺激により確認し、結果として、8〜10倍高いIFN分泌が生じたが、IL-10の分泌は減少した。
【0192】
抗CD3/CD28およびIL-2(CD8+細胞について)またはTh1/Th2分極条件(CD4+細胞について)の存在下でのナイーブT細胞のインビトロ培養は、一次刺激培養物において低レベルのGFPを示した。しかし、再び刺激すると、Blimpgfp/+およびBlimpgfp/gfpマウスからのCD4およびCD8 T細胞は、迅速にGFP+になった(図19B)。重要なことに、野生型CD4+エフェクターT細胞におけるBlimp-1発現が、ウエスタンブロッティングにより確認された(図19C)。これらのデータは、Blimp-1発現の誘導が、ナイーブCD4+およびCD8+のエフェクター細胞への成熟の一成分であることを証明している。
【0193】
特異的抗原刺激後のBlimp-1発現をさらに評価するために、感染性疾患モデルを使用した。マウスを単純疱疹ウイルス(HSV)に感染させ、感染後、様々な時点で、優性エピトープに特異的な四量体および標準的な細胞毒性機能アッセイを使用してウイルス特異的CD8+ T細胞の一次および記憶応答についてモニターした。
【0194】
これらの実験により抗原特異的T細胞におけるBlimp-1の発現が確認された。また、これらの実験は、Blimpgfp/gfp細胞が、抗原特異的標的細胞を溶解する能力に関して十分機能的であることを示している。
【0195】
Blimpgfp/gfpで再構成されたマウスにおいて観察された活性化T細胞の増幅プールは、Blimp-1不在下での異常応答性または脱調節恒常性を示唆していた。この結論に合わせて、Blimpgfp/gfpで再構成されたマウスは、顕著な体重減少、外皮の皺および下痢を示し、再構成後6週間目から犠牲にしなければならなかった。これらのマウスの組織分析は、広範なリンパ球浸潤、ならびに肺、肝臓および胃腸管を含む様々な器官の炎症を示した(図21A)。組織損傷は、肺および腸において最も顕著であり、このプロセスが、これらのマウスにおいて観察された体重減少、下痢および死亡に関連していた。再構成されたマウスのリンパ器官および肝臓のフローサイトメトリー分析は、全ての器官における完全に活性化したCD4+およびCD8+ T細胞の大きな増幅を示した(図2B)。CD4+細胞は、主としてTh1に偏っていた。さらにBlimpgfp/gfp Rag2-/-胎仔肝臓を有するRag1-/-マウスは、結果として一切死なず、観察された病状にはT細胞が強く関与していた(図21)。
【0196】
無制御T細胞増幅が、T細胞に固有のものであるのかを判定するために、選別されたT細胞のRag1-/-への養子移入実験を行った。これらのアッセイにおいて、野生型T細胞は、恒常性増幅を制限されることによるリンパ球減少性環境に応答する(図22A)。T細胞、すなわち3×106個のCD4+またはCD8+ C57BL/6またはBlimpgfp/gfp、をRag1-/-受容個体に注射し、移入の3週間後に脾臓T細胞の数を評価した。図22Aに示すように、両方の系列のBlimpgfp/gfp T細胞が、劇的に増幅された恒常性増幅能力を有した。さらに、結果として生じたT細胞全てが、GFPを発現し、野生型対照物は、活性化された表現型を提示した(図22)。Blimpgfp/gfp T細胞を受けた受容個体は、図21に記載されているような、体重減少、巨脾腫(図22)および類似した範囲のリンパ球浸潤表現型を急速に発現した。これらのデータは、Blimp-1欠失T細胞が、多臓器浸潤および死を結果として生じさせる異常調節増幅を提示することを実証している。
【0197】
自己免疫性は、機能的CD4+CD25+サプレッサーT細胞の喪失に関連付けられている。Blimp+/gfpおよびBlimpgfp/gfp CD4+ T細胞におけるこの集団の分析は、CD25+ CD4 T細胞におけるGFPの中等度〜強度の発現を示した。これは、調節T細胞集団におけるBlimp-1についての機能を示唆している。インビトロおよびインビボでの機能分析により、Tregについての重要な因子FoxP3、およびTregに関連した他の遺伝子の不変の発現が観察された。Blimp欠失細胞CD4細胞は、高レベルのIFNを分泌し、IL-10分泌不良を示す。
【0198】
インビボでBlimp-1欠失T細胞において見られる異常調節恒常性が、それ自体を強化増幅としてインビトロでも表すかを判定するために、培養ナイーブCD8+ T細胞を抗CD3/CD28ならびに細胞増殖および恒常性を調節することが公知であるサイトカイン(IL-2、IL-15およびIL-21を含む)の存在下でインキュベートした。上の条件下で7日間成長させたナイーブCD8+細胞は、GFP発現、および遺伝子型の間で類似した増殖プロフィールをほとんど示さなかった(図23A)。ナイーブCD4+細胞、およびPMA/イオノマイシンの組み合わせにより刺激したものについて、同様の結果が観察された(データは示していない)。対照的に、IL-2、IL-15またはIL-21の存在下でのCD8+細胞の二次刺激は、結果として、Blimp-1発現の強い誘導を生じさせた(図23A)。この発現プロフィールと一致して、Blimp-1が十分な細胞と比較して、蓄積細胞数の有意な増加が、Blimpgfp/gfp細胞の増殖応答において観察された。予測されたとおり、活性化/記憶CD44high細胞を一次応答のために使用したとき、増殖能に同様の強い差異が観察された。この増加は、全てのサイトカイン条件下で観察されたが、IL-21の存在下で最も顕著であった(図23B)。IL-21は、NODマウスにおける糖尿病感受性遺伝子候補であることが最近明らかになった(King, et al. Cell 117:265-277, 2004)Tヘルパーサイトカインである。このモデルでは、IL-21が、エフェクターT細胞の代謝回転を増加させ、その結果としてリンパ球減少誘導恒常性増殖および糖尿病が生じた(King, et al., 2004 (前記))。興味深いことに、IL-21は、B細胞最終分化におけるBlimp-1の強力な刺激因子であり、またCD8+ T細胞におけるGFPの最も効率的な誘導因子であった。このことは、リンパ球分化におけるこのサイトカインの共通の役割を示唆している。
【0199】
分子およびインビトロ試験によって、Blimp-1は、ヒストンメチルトランスフェラーゼ(Gyory et al., Nat Immunol 5: 299-308, 2004)、デアセチラーゼ(Yu et al., Mol Cell Biol 20:2592-3603, 2000)および標的をサイレンシングさせるためのGrouchoファミリーのコリプレッサー(Ren et al., Genes Dev 13:125-137, 1999)を補充することができる強力な転写リプレッサーであることが示された。プロモータおよびマイクロアレイの研究により、B細胞系列において多数のBlimp-1標的が同定された(Shaffer et al, Immunity 17:51-62, 2002; Shaffer et al., Immunity 21:81-93, 2004)。これらのうち、c-myc(Lin et al., Science 276:596, 1997)、CIITA(Piskurich et al., Nat Immunol 1:526-532, 2000)を含む多数が、T細胞系列で発現される。
【0200】
実施例11
Blimpは、全てのリンパ球における保存的な最終分化プログラムの主調節因子である
要約すると、本明細書において記載のデータは、Blimp-1が、活性化された従来のT細胞において様々な状況下で発現されることを実証している。Blimp-1欠失T細胞を注射した、または突然変異幹細胞で再構成されたマウスが、進行性多器官リンパ球増殖性疾患の結果として死亡するので、Blimp発現は、正常なリンパ球恒常性には不可欠である。加えて、T細胞分化の恒常性を調節することが公知であるIL-21などのサイトカインは、Blimp-1発現の強力な誘導因子であり、Blimp-1不在下、インビトロで強化増殖を支持した。従って、Blimp-1発現は、初期刺激によってではなく、免疫応答の完了に向けて、エフェクターT細胞の分化を誘導し得る。従って、Blimp-1は、T細胞収縮の遺伝プログラムおよび/または免疫恒常性に不可欠である記憶形成を調節するように機能する、同定された最初の転写因子である。
【0201】
BおよびTリンパ球は、それらの発達中に多数の細胞的および遺伝子的類似性、例えば、共通の前駆細胞、定序逐次VDJ組換えおよび類似した発達チェックポイントを共有するが、形質細胞へのB細胞の最終分化が、明確な機能的終点である(Calame et al., Annu Rev Immunol 8:8, 2003)一方で、T細胞発生の最終段階は、ほとんど定義されていない。BおよびT細胞系列内のBlimp-1の類似した機能および発現プロフィールは、その外見的に異なる外観にもかかわらず、Blimp-1が、全てのリンパ球における保存的最終分化プログラムの主調節因子であるという興味深い可能性を提起する。
【0202】
Blimp欠失は、過形成および無制御増殖の原因となり、その一方で、Blimpの発現が、リンパ球恒常性ならびにASC、T細胞およびB細胞を含む造血細胞の最終分化を可能にする。従って、サイトカインおよび他の免疫調節成分、化学物質、ペプチドまたは小もしくは中分子剤は、本明細書において記載のインビトロおよびインビボ細胞モデル系においてスクリーニングして、治療薬または予防薬としての可能性を判定することができる。
【0203】
従って、本モデルレポーター系は、T細胞およびB細胞などの免疫系の細胞における最終分化を調節する薬剤の能力を評価する際に有用である。
【0204】
本明細書において記載の本発明には、明記している以外の様々な変化および変更の余地があることは、当業者には理解されるだろう。本発明が、全てのそのような変化および変更を包含することは、理解されるはずである。本発明は、個々にまたは集合的に、本明細書に引用している、または本明細書において指摘している段階、特徴、組成物および化合物の全て、ならびに段階または特徴のいずれか2つ以上の任意のおよび全ての組み合わせも包含する。
【0205】
参考文献一覧
【特許請求の範囲】
【請求項1】
内因性Blimp調節要素の制御下で発現され、かつ、レポーター分子と共発現されるBlimp(PRDM-1)またはその機能的もしくは非機能的な部分、形態、相同体もしくは変異体を産生する、Blimpポリペプチドをコードする改変Blimp(PRDM-1)遺伝子を含む遺伝子改変細胞またはそのような細胞を含む非ヒト生物であって、細胞内におけるBlimpの存在が、細胞表現型および/または細胞における最終分化への決定付け(commitment)に関連する、細胞または生物。
【請求項2】
Blimp遺伝子が、Blimpコード配列を含むかまたはその機能的もしくは非機能的な部分、形態、相同体もしくは変異体およびレポーター分子コード配列をコードするBlimp mRNA転写産物をコードする、請求項1記載の細胞または生物。
【請求項3】
レポーター分子コード配列がBlimp対立遺伝子のイントロン内に挿入される、請求項2記載の細胞または生物。
【請求項4】
改変Blimp対立遺伝子がホモ接合またはヘテロ接合の形態で存在する、請求項3記載の細胞または非ヒト生物。
【請求項5】
改変Blimp対立遺伝子がヘテロ接合の形態で存在する、請求項4記載の細胞または非ヒト生物。
【請求項6】
改変Blimp対立遺伝子が、機能的Blimp転写因子またはその機能的な部分、形態、相同体もしくは変異体をコードする、請求項1〜5のいずれか一項記載の細胞または非ヒト生物。
【請求項7】
改変Blimp対立遺伝子が、非機能的Blimp転写因子またはその非機能的な部分、形態、相同体もしくは変異体をコードする、請求項1〜5のいずれか一項記載の細胞または非ヒト生物。
【請求項8】
ヒト、非ヒト霊長類、家畜、愛玩生物、実験検査生物、爬虫類または両生類などの任意の生物に由来する細胞または遺伝物質を含む、請求項1記載の細胞または非ヒト生物。
【請求項9】
齧歯動物(マウスを含む)、モルモット、ブタ、カモ、ウサギまたはヒツジなどの実験検査動物に由来する、請求項8記載の細胞または生物。
【請求項10】
細胞が造血細胞または胚細胞である、請求項1〜5のいずれか一項記載の細胞または生物。
【請求項11】
細胞が造血細胞である、請求項10に記載の細胞または生物。
【請求項12】
細胞がリンパ球細胞である、請求項11に記載の細胞または生物。
【請求項13】
細胞が、B細胞およびT細胞から選択されるリンパ球系列の細胞である、請求項12記載の細胞または生物。
【請求項14】
B細胞がASCである、請求項13記載の細胞または生物。
【請求項15】
ASCの実質的に精製された集団である、請求項14記載の細胞。
【請求項16】
T細胞がCD4+ T細胞およびCD8+ T細胞から選択される、請求項13記載の細胞または生物。
【請求項17】
レポーター分子の検出が細胞表現型および/または最終分化への細胞の決定付けの指標となる、請求項1〜16のいずれか一項記載の細胞または生物。
【請求項18】
胚、生殖体またはES細胞の形態で提供される、請求項1〜14、16および17のいずれか一項記載の非ヒト生物。
【請求項19】
レポーター分子が蛍光または発光レポーター分子である、請求項1〜18のいずれか一項記載の細胞または生物。
【請求項20】
内因性Blimp調節要素の制御下で発現され、かつ、Blimpまたはその機能的もしくは非機能的な部分、形態、相同体もしくは変異体およびレポーター分子を共発現する、Blimpタンパク質をコードする改変Blimp遺伝子を含む遺伝子改変造血細胞またはそのような細胞を含む非ヒト動物をスクリーニングする段階を含み、レポーター活性の検出が、細胞表現型および/または最終分化への細胞の決定付けの指標となる、造血系の細胞を表現型解析および/またはモニタリングする方法。
【請求項21】
造血細胞がB細胞、T細胞、樹状細胞、マクロファージ、ナチュラルキラー細胞、顆粒球、赤血球、好酸球、巨核球、骨髄細胞、脾細胞、皮膚細胞または間質細胞ならびにこれらの前駆体および誘導体から選択される細胞である、請求項20記載の方法。
【請求項22】
細胞の表現型解析および/またはモニタリングが蛍光または発光レポーター分子のサイトメトリー分析により達成される、請求項20または21記載の方法。
【請求項23】
レポーター活性を示さない細胞の中から、レポーター活性またはレポーター活性もしくはレベルの変化を示す細胞を単離または選択する段階をさらに含む、請求項20記載の方法。
【請求項24】
レポーター活性細胞の単離が、フローサイトメトリー、レーザー走査サイトメトリー、クロマトグラフィーおよび/または他の同等の手法によるものである、請求項23記載の方法。
【請求項25】
さらなる選択マーカーを使用してレポーター活性細胞を選択する段階をさらに含む、請求項23記載の方法。
【請求項26】
細胞がB細胞系列の細胞の集団において同定または単離されたASCである、請求項20〜25のいずれか一項記載の方法。
【請求項27】
細胞がT細胞の混合集団において同定または単離された活性化/最終分化T細胞である、請求項20〜25のいずれか一項記載の方法。
【請求項28】
以下の段階を含む、ワクチンの抗原性または免疫原性を検査する方法:
(i)遺伝子改変造血細胞またはそのような細胞を含む非ヒト動物にワクチンを投与する段階であって、細胞または生物は、レポーター分子と共発現される、Blimpまたはその一部、フラグメントもしくは機能的な形態を、発現されたときに産生する、Blimpポリペプチドをコードする改変Blimp-1遺伝子を含む、段階;
(ii)その存在が造血細胞において最終分化を誘導するワクチンの能力の指標となるレポーター分子について、細胞または生物を検査する段階。
【請求項29】
レポーター活性の存在がT細胞および/またはB細胞において最終分化を促進するワクチンの能力の指標となる、請求項27記載の方法。
【請求項30】
以下の段階を含む、造血細胞における最終分化の作動薬(agonist)または拮抗薬(antagonist)についてインビトロまたはインビボでスクリーニングする方法:遺伝子改変細胞またはそのような細胞を含む非ヒト動物に1つまたは複数の薬剤を暴露する段階であって、細胞または生物が、レポーター分子と共発現される、Blimpまたはその一部もしくはフラグメントもしくは機能的な形態を、発現されたときに産生する、Blimpポリペプチドをコードする改変Blimp-1遺伝子を含む、段階;ならびに、その存在が、最終分化を作動し、または最終分化に拮抗する1つまたは複数の薬剤の能力の指標となるレポーター分子の存在またはレポーター分子のレベルの変化について、細胞または生物を検査する段階。
【請求項31】
細胞が、Blimpコード配列を含むかまたはその機能的もしくは非機能的な部分、形態、相同体もしくは変異体およびレポーター分子コード配列をコードする、Blimp mRNA転写産物をコードする改変Blimp遺伝子を含む、請求項20、28または30いずれか一項記載の方法。
【請求項32】
レポーター分子コード配列がBlimp対立遺伝子のイントロン内に挿入される、請求項31記載の方法。
【請求項33】
改変Blimp対立遺伝子がホモ接合またはヘテロ接合の形態で存在する、請求項32記載の方法。
【請求項34】
改変Blimp対立遺伝子がヘテロ接合の形態で存在する、請求項33記載の方法。
【請求項35】
改変Blimp対立遺伝子が、機能的Blimp転写因子またはその機能的な部分、形態、相同体もしくは変異体をコードする、請求項31記載の方法。
【請求項36】
改変Blimp対立遺伝子が、非機能的Blimp転写因子またはその非機能的な部分、形態、相同体もしくは変異体をコードする、請求項31記載の方法。
【請求項37】
細胞または遺伝物質が、ヒト、非ヒト霊長類、家畜、愛玩生物、または実験検査生物、爬虫類または両生類などの任意の生物に由来する、請求項31記載の方法。
【請求項38】
実験検査生物が、齧歯動物(マウスを含む)、モルモット、ブタ、カモ、ウサギおよびヒツジから選択される、請求項37記載の方法。
【請求項39】
細胞が、癌性または非癌性の造血または胚細胞である、請求項31記載の方法。
【請求項40】
細胞がリンパ球細胞である、請求項39記載の方法。
【請求項41】
リンパ球細胞がB細胞およびT細胞から選択される、請求項40記載の方法。
【請求項42】
B細胞がASCである、請求項41記載の方法。
【請求項43】
T細胞がCD4+ T細胞およびCD8+ T細胞から選択される、請求項41記載の方法。
【請求項44】
内因性Blimp調節要素の制御下で標的細胞において発現され、かつ、レポーター分子と共発現される、Blimp(PRDM-1)またはその機能的もしくは非機能的な部分、形態、相同体もしくは変異体を産生する、Blimpポリペプチドおよびレポーター分子をコードする改変Blimp(PRDM-1)遺伝子を含む、ターゲティングベクター。
【請求項45】
Blimp遺伝子が、Blimpコード配列を含むかまたはその機能的もしくは非機能的な部分、形態、相同体もしくは変異体およびレポーター分子コード配列をコードする、Blimp mRNA転写産物をコードする、請求項44記載のターゲティングベクター。
【請求項46】
レポーター分子コード配列がBlimp対立遺伝子のイントロン内に挿入される、請求項45記載のターゲティングベクター。
【請求項47】
レポーター分子がGFPである、請求項44記載のターゲティングベクター。
【請求項1】
内因性Blimp調節要素の制御下で発現され、かつ、レポーター分子と共発現されるBlimp(PRDM-1)またはその機能的もしくは非機能的な部分、形態、相同体もしくは変異体を産生する、Blimpポリペプチドをコードする改変Blimp(PRDM-1)遺伝子を含む遺伝子改変細胞またはそのような細胞を含む非ヒト生物であって、細胞内におけるBlimpの存在が、細胞表現型および/または細胞における最終分化への決定付け(commitment)に関連する、細胞または生物。
【請求項2】
Blimp遺伝子が、Blimpコード配列を含むかまたはその機能的もしくは非機能的な部分、形態、相同体もしくは変異体およびレポーター分子コード配列をコードするBlimp mRNA転写産物をコードする、請求項1記載の細胞または生物。
【請求項3】
レポーター分子コード配列がBlimp対立遺伝子のイントロン内に挿入される、請求項2記載の細胞または生物。
【請求項4】
改変Blimp対立遺伝子がホモ接合またはヘテロ接合の形態で存在する、請求項3記載の細胞または非ヒト生物。
【請求項5】
改変Blimp対立遺伝子がヘテロ接合の形態で存在する、請求項4記載の細胞または非ヒト生物。
【請求項6】
改変Blimp対立遺伝子が、機能的Blimp転写因子またはその機能的な部分、形態、相同体もしくは変異体をコードする、請求項1〜5のいずれか一項記載の細胞または非ヒト生物。
【請求項7】
改変Blimp対立遺伝子が、非機能的Blimp転写因子またはその非機能的な部分、形態、相同体もしくは変異体をコードする、請求項1〜5のいずれか一項記載の細胞または非ヒト生物。
【請求項8】
ヒト、非ヒト霊長類、家畜、愛玩生物、実験検査生物、爬虫類または両生類などの任意の生物に由来する細胞または遺伝物質を含む、請求項1記載の細胞または非ヒト生物。
【請求項9】
齧歯動物(マウスを含む)、モルモット、ブタ、カモ、ウサギまたはヒツジなどの実験検査動物に由来する、請求項8記載の細胞または生物。
【請求項10】
細胞が造血細胞または胚細胞である、請求項1〜5のいずれか一項記載の細胞または生物。
【請求項11】
細胞が造血細胞である、請求項10に記載の細胞または生物。
【請求項12】
細胞がリンパ球細胞である、請求項11に記載の細胞または生物。
【請求項13】
細胞が、B細胞およびT細胞から選択されるリンパ球系列の細胞である、請求項12記載の細胞または生物。
【請求項14】
B細胞がASCである、請求項13記載の細胞または生物。
【請求項15】
ASCの実質的に精製された集団である、請求項14記載の細胞。
【請求項16】
T細胞がCD4+ T細胞およびCD8+ T細胞から選択される、請求項13記載の細胞または生物。
【請求項17】
レポーター分子の検出が細胞表現型および/または最終分化への細胞の決定付けの指標となる、請求項1〜16のいずれか一項記載の細胞または生物。
【請求項18】
胚、生殖体またはES細胞の形態で提供される、請求項1〜14、16および17のいずれか一項記載の非ヒト生物。
【請求項19】
レポーター分子が蛍光または発光レポーター分子である、請求項1〜18のいずれか一項記載の細胞または生物。
【請求項20】
内因性Blimp調節要素の制御下で発現され、かつ、Blimpまたはその機能的もしくは非機能的な部分、形態、相同体もしくは変異体およびレポーター分子を共発現する、Blimpタンパク質をコードする改変Blimp遺伝子を含む遺伝子改変造血細胞またはそのような細胞を含む非ヒト動物をスクリーニングする段階を含み、レポーター活性の検出が、細胞表現型および/または最終分化への細胞の決定付けの指標となる、造血系の細胞を表現型解析および/またはモニタリングする方法。
【請求項21】
造血細胞がB細胞、T細胞、樹状細胞、マクロファージ、ナチュラルキラー細胞、顆粒球、赤血球、好酸球、巨核球、骨髄細胞、脾細胞、皮膚細胞または間質細胞ならびにこれらの前駆体および誘導体から選択される細胞である、請求項20記載の方法。
【請求項22】
細胞の表現型解析および/またはモニタリングが蛍光または発光レポーター分子のサイトメトリー分析により達成される、請求項20または21記載の方法。
【請求項23】
レポーター活性を示さない細胞の中から、レポーター活性またはレポーター活性もしくはレベルの変化を示す細胞を単離または選択する段階をさらに含む、請求項20記載の方法。
【請求項24】
レポーター活性細胞の単離が、フローサイトメトリー、レーザー走査サイトメトリー、クロマトグラフィーおよび/または他の同等の手法によるものである、請求項23記載の方法。
【請求項25】
さらなる選択マーカーを使用してレポーター活性細胞を選択する段階をさらに含む、請求項23記載の方法。
【請求項26】
細胞がB細胞系列の細胞の集団において同定または単離されたASCである、請求項20〜25のいずれか一項記載の方法。
【請求項27】
細胞がT細胞の混合集団において同定または単離された活性化/最終分化T細胞である、請求項20〜25のいずれか一項記載の方法。
【請求項28】
以下の段階を含む、ワクチンの抗原性または免疫原性を検査する方法:
(i)遺伝子改変造血細胞またはそのような細胞を含む非ヒト動物にワクチンを投与する段階であって、細胞または生物は、レポーター分子と共発現される、Blimpまたはその一部、フラグメントもしくは機能的な形態を、発現されたときに産生する、Blimpポリペプチドをコードする改変Blimp-1遺伝子を含む、段階;
(ii)その存在が造血細胞において最終分化を誘導するワクチンの能力の指標となるレポーター分子について、細胞または生物を検査する段階。
【請求項29】
レポーター活性の存在がT細胞および/またはB細胞において最終分化を促進するワクチンの能力の指標となる、請求項27記載の方法。
【請求項30】
以下の段階を含む、造血細胞における最終分化の作動薬(agonist)または拮抗薬(antagonist)についてインビトロまたはインビボでスクリーニングする方法:遺伝子改変細胞またはそのような細胞を含む非ヒト動物に1つまたは複数の薬剤を暴露する段階であって、細胞または生物が、レポーター分子と共発現される、Blimpまたはその一部もしくはフラグメントもしくは機能的な形態を、発現されたときに産生する、Blimpポリペプチドをコードする改変Blimp-1遺伝子を含む、段階;ならびに、その存在が、最終分化を作動し、または最終分化に拮抗する1つまたは複数の薬剤の能力の指標となるレポーター分子の存在またはレポーター分子のレベルの変化について、細胞または生物を検査する段階。
【請求項31】
細胞が、Blimpコード配列を含むかまたはその機能的もしくは非機能的な部分、形態、相同体もしくは変異体およびレポーター分子コード配列をコードする、Blimp mRNA転写産物をコードする改変Blimp遺伝子を含む、請求項20、28または30いずれか一項記載の方法。
【請求項32】
レポーター分子コード配列がBlimp対立遺伝子のイントロン内に挿入される、請求項31記載の方法。
【請求項33】
改変Blimp対立遺伝子がホモ接合またはヘテロ接合の形態で存在する、請求項32記載の方法。
【請求項34】
改変Blimp対立遺伝子がヘテロ接合の形態で存在する、請求項33記載の方法。
【請求項35】
改変Blimp対立遺伝子が、機能的Blimp転写因子またはその機能的な部分、形態、相同体もしくは変異体をコードする、請求項31記載の方法。
【請求項36】
改変Blimp対立遺伝子が、非機能的Blimp転写因子またはその非機能的な部分、形態、相同体もしくは変異体をコードする、請求項31記載の方法。
【請求項37】
細胞または遺伝物質が、ヒト、非ヒト霊長類、家畜、愛玩生物、または実験検査生物、爬虫類または両生類などの任意の生物に由来する、請求項31記載の方法。
【請求項38】
実験検査生物が、齧歯動物(マウスを含む)、モルモット、ブタ、カモ、ウサギおよびヒツジから選択される、請求項37記載の方法。
【請求項39】
細胞が、癌性または非癌性の造血または胚細胞である、請求項31記載の方法。
【請求項40】
細胞がリンパ球細胞である、請求項39記載の方法。
【請求項41】
リンパ球細胞がB細胞およびT細胞から選択される、請求項40記載の方法。
【請求項42】
B細胞がASCである、請求項41記載の方法。
【請求項43】
T細胞がCD4+ T細胞およびCD8+ T細胞から選択される、請求項41記載の方法。
【請求項44】
内因性Blimp調節要素の制御下で標的細胞において発現され、かつ、レポーター分子と共発現される、Blimp(PRDM-1)またはその機能的もしくは非機能的な部分、形態、相同体もしくは変異体を産生する、Blimpポリペプチドおよびレポーター分子をコードする改変Blimp(PRDM-1)遺伝子を含む、ターゲティングベクター。
【請求項45】
Blimp遺伝子が、Blimpコード配列を含むかまたはその機能的もしくは非機能的な部分、形態、相同体もしくは変異体およびレポーター分子コード配列をコードする、Blimp mRNA転写産物をコードする、請求項44記載のターゲティングベクター。
【請求項46】
レポーター分子コード配列がBlimp対立遺伝子のイントロン内に挿入される、請求項45記載のターゲティングベクター。
【請求項47】
レポーター分子がGFPである、請求項44記載のターゲティングベクター。
【図1】


【図2】
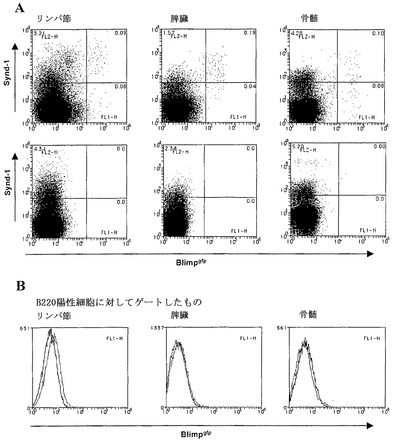
【図3】
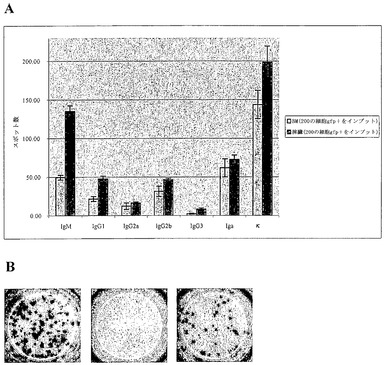
【図4A】
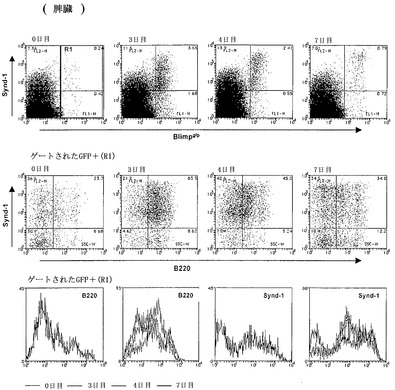
【図4B】
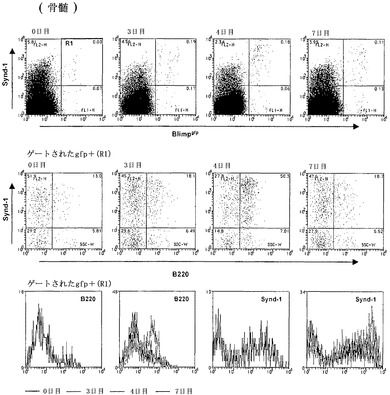
【図5】
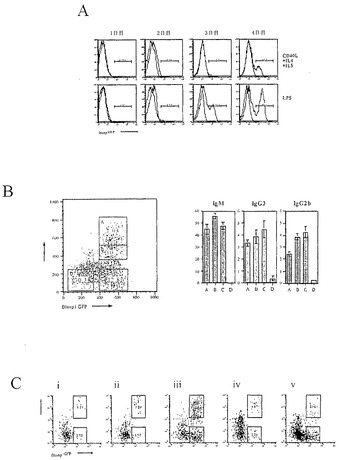
【図6】
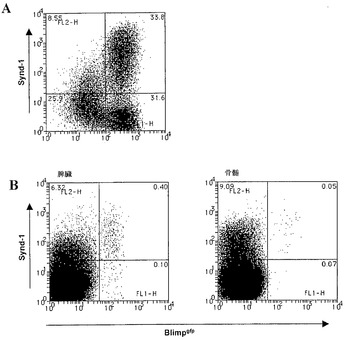
【図7】
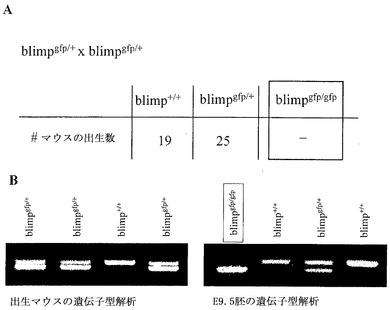
【図8】
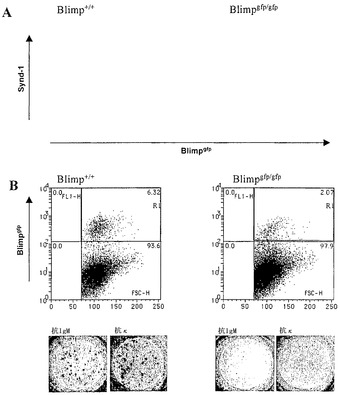
【図9】
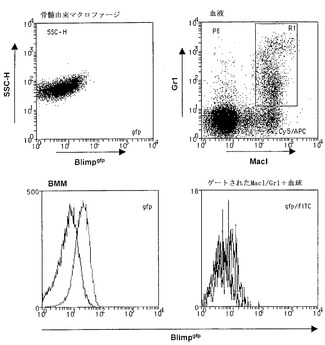
【図10】
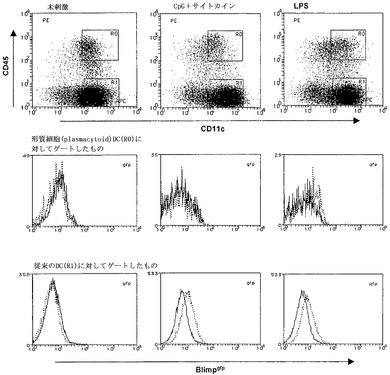
【図11】
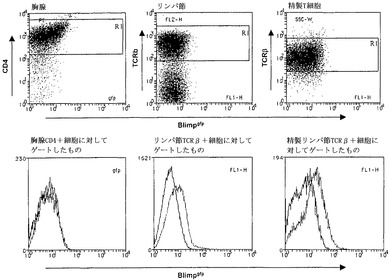
【図12】

【図13−1】

【図13−2】

【図13−3】

【図13−4】

【図14】

【図15−1】

【図15−2】

【図16】

【図17−1】

【図17−2】

【図17−3】

【図17−4】

【図17−5】

【図17−6】

【図18−1】

【図18−2】

【図18−3】

【図18−4】

【図18−5】

【図18−6】

【図18−7】

【図19A】
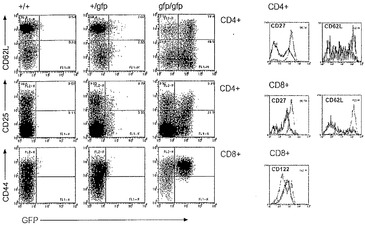
【図19B】
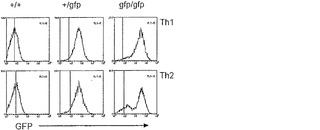
【図19C】

【図19D】

【図19E】

【図20A】
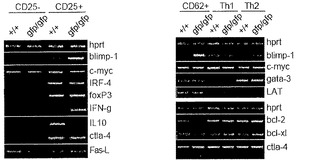
【図20B】

【図21】

【図22A】
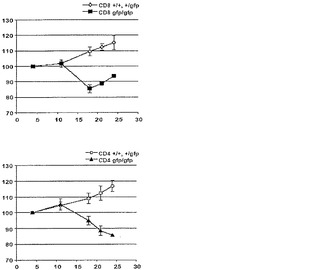
【図22B】
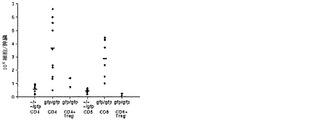
【図22C】

【図22D】
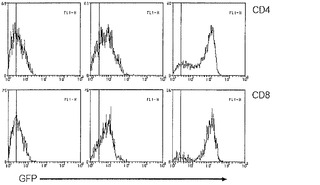
【図23】
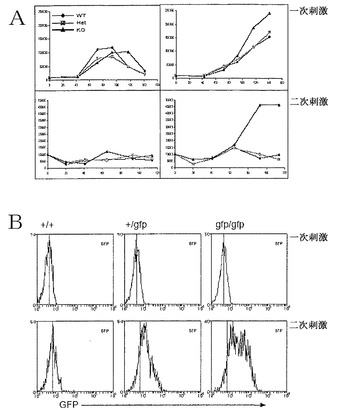
【図2】
【図3】
【図4A】
【図4B】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13−1】
【図13−2】
【図13−3】
【図13−4】
【図14】
【図15−1】
【図15−2】
【図16】
【図17−1】
【図17−2】
【図17−3】
【図17−4】
【図17−5】
【図17−6】
【図18−1】
【図18−2】
【図18−3】
【図18−4】
【図18−5】
【図18−6】
【図18−7】
【図19A】
【図19B】
【図19C】
【図19D】
【図19E】
【図20A】
【図20B】
【図21】
【図22A】
【図22B】
【図22C】
【図22D】
【図23】
【公開番号】特開2012−50431(P2012−50431A)
【公開日】平成24年3月15日(2012.3.15)
【国際特許分類】
【出願番号】特願2011−174435(P2011−174435)
【出願日】平成23年8月10日(2011.8.10)
【分割の表示】特願2006−552420(P2006−552420)の分割
【原出願日】平成17年2月11日(2005.2.11)
【公序良俗違反の表示】
(特許庁注:以下のものは登録商標)
1.WINDOWS
【出願人】(505098650)ウォルター アンド エリザ ホール インスティテュート オブ メディカル リサーチ (11)
【Fターム(参考)】
【公開日】平成24年3月15日(2012.3.15)
【国際特許分類】
【出願日】平成23年8月10日(2011.8.10)
【分割の表示】特願2006−552420(P2006−552420)の分割
【原出願日】平成17年2月11日(2005.2.11)
【公序良俗違反の表示】
(特許庁注:以下のものは登録商標)
1.WINDOWS
【出願人】(505098650)ウォルター アンド エリザ ホール インスティテュート オブ メディカル リサーチ (11)
【Fターム(参考)】
[ Back to top ]