
植物体中でのシアル酸の合成
植物体においてシアル酸を合成する方法、及びシアル酸を合成することが可能である植物体が提供される。さらに、植物体においてシアル化されたタンパク質を産生する方法も提供される。シアル酸を合成する方法は、N−アセチルノイラミン酸(Neu5Ac)シンターゼ又はNeu5Acリアーゼをコードするヌクレオチド配列を含む植物体を提供することと、該ヌクレオチド配列を発現させ、それによってシアル酸を合成することを含む。植物体は、エピメラーゼ、CMP−Neu5Acシンターゼ、CMP−Neu5Acトランスポーター及びシアリルトランスフェラーゼのうちの1又は1を超えるものをコードするヌクレオチド配列を共発現してもよい。

【発明の詳細な説明】
【技術分野】
【0001】
(発明の分野)
本発明は、植物体におけるシアル酸の合成に関する。さらに、本発明はシアル酸を産生する方法及び植物体を提供し、これら植物体から産生されるシアル化タンパク質を提供する。
【背景技術】
【0002】
(発明の背景)
植物は、組換え医薬タンパク質を製造するための、コストの低い、且つ混入汚染について安全な製造現場となる可能性がある。アミノ酸配列、立体配置及び生物活性に関する限り、植物で産生された組換えタンパク質のほとんどは哺乳類で製造されたものと区別できない。さらに、哺乳類の糖タンパク質は、それらがトランスジェニック植物で発現された場合、効率的にグリコシル化される。しかしながら、植物は、動物の糖タンパク質で見い出されるものとは異なるN−グリカンを持つ分子を産生する(非特許文献1)。このことは、これらのタンパク質上の植物特異的な糖エピトープがヒトにおいて免疫応答を誘発する(非特許文献2)と共に、シアル化配列のような哺乳類型のエピトープが存在しないことが血流からのそれらの迅速なクリアランスを誘導する可能性があるので、植物で作製された医薬品の使用を限定する可能性がある。その結果、植物で作製された医薬のN−グリコシル化を制御することがヒトの治療法で使用することの前提条件となっている。
【0003】
最近、植物体でのリモデリング戦略が出現し、ヒトと適合可能な糖質特性を持つ植物由来の抗体が得られている。作戦の一部には、小胞体におけるプランティボディの保持が関与し(非特許文献3;非特許文献4;非特許文献5)、他の作戦には、哺乳類のグリコシルトランスフェラーゼによる植物の形質転換が関与していた。たとえば、植物のN−グリコシル化は、ヒトのβ(1,4)ガラクトシルトランスフェラーゼ(非特許文献6;非特許文献7)による植物の形質転換によって部分的にヒト化することができる。形質転換された植物におけるマウスの抗体の発現は結果として、相当するマウスのIgGで認められるものに類似するガラクトシル化特性を抱く植物由来の抗体を産生した(非特許文献7)。
【0004】
哺乳類のIgGは、Fcドメインに位置するNグリコシル化の保存された部位で二分岐N−グリカンを持つ。これらのオリゴ糖は弱くシアル化されており、末端Neu5Acが存在しないので、抗体の機能や安定性を妨害することはない。対照的に、そのほかの循環性糖タンパク質はほとんどシアル化された2、3又は4分岐のN−グリカンを有する。これらグリカンにおける末端シアル酸の存在は、多数の生体機能に必要とされ、その第1は、循環系におけるタンパク質の半減期の制御である。末端シアル酸の非存在下では、糖タンパク質は肝臓のアシアロ糖タンパク質受容体によって検出され、血清からクリアランスされ、これらのタンパク質の半減期を短く、無効なものにする(非特許文献8)。従って、非シアル化植物によって作製された医薬は、ヒトに注射された場合、血流から迅速に除かれる可能性があり、たとえば、タバコに由来するEpoはin vitroでは生物学的に活性を持っていたが、赤血球造血組織に到達する前に循環から除かれるためにin vivoでは、非機能性だった(非特許文献9)。
【0005】
プランティボディに連結するN−グリカンのヒト様N−グリカンへのリモデリングは、共基質として内因性UDP−Galを使用するトランスフェラーゼである、ヒトのβ(1,4)ガラクトシルトランスフェラーゼ(非特許文献6および7)の発現によって、植物ではすでに部分的に達成されている。哺乳類のシアリルトランスフェラーゼも植物に導入されており、機能性であり、正確にゴルジ装置を標的とすることが実証されている(非特許文献10)。しかしながら、内因性オリゴ多糖類のシアル化は認められなかった。植物におけるシアル酸及びシアル化機構の存在は依然として議論のあることころである。しかしながら、ヒトに存在する主なシアル酸であるNeu5Ac、同様にその前駆体、N−アセチルマンノサミン(D−ManNAc)は、検出可能な量で植物にて合成されるとは思われない(非特許文献11)。その結果、植物のN−グリカンのシアル化オリゴ糖へのグリコ操作は、合成を触媒することが可能な内因性酵素の共発現、Neu5Acの活性化及びゴルジ装置への移行を必要とする。
【0006】
哺乳類及び細菌では、Neu5Acの同化作用及び異化作用は、異なった経路を介して生じる(非特許文献12)。主な部類の2つの酵素がNeu−5Acを形成するのに必要とされる。N−アセチルノイラミネートリアーゼ(Neu5Acリアーゼ)は、可逆的反応におけるNeu5AcのN−アセチルマンノサミン(D−ManNAc)とピルビン酸塩への開裂を触媒することによってシアル酸の異化作用に関与する。高濃度のD−ManNAcとピルビン酸塩にて、平衡は、Neu5Acの合成に移行しうる。グルコサミン2−エピメラーゼ活性に連動して、E.coli由来のNeu5AcリアーゼをD−GlcNAcからのNeu5Acの大量生産に利用した(非特許文献13)。或いは、NeuBのようなNeu5Acシンターゼは、ピルビン酸ホルホエノール(PEP)へのManNAcの縮合を触媒し、シアル酸の生合成に直接関与する(非特許文献14で概説された)。
【非特許文献1】Lerouge, P., Cabanes−Macheteau, M., Rayon, C, Fitchette−Laine, A.−C, Gomord, V. and Faye, L. (1998) N−glycoprotein biosynthesis: recent development and future trends.Plant MoI. Biol, 38, 31−48
【非特許文献2】Bardor M, Faveeuw C, Fitchette A−C, Gilbert D, Galas L, Trottein F, Faye L and Lerouge P. (2003) Immureactivity in mammals of two typical plant glyco−epitopes, core−alpha(l,3)−fucose and core−xylose. Glycobiology, 13, 427−434
【非特許文献3】Ko, K., Tekoah, Y., Rudd, P.M., Harvey, D.J., Dwek, R.A., Spitsin, S., Hanlon, C.A., Rupprecht C, Dietzschold, B., Golovkin, M. and Koprowski, H. (2003) Function and glycosylation of plant−derived antiviral monoclonal antibody. Proc. Natl Acad. ScL USA, 101, 8013−8018
【非特許文献4】Sriraman, R., Bardor, M., Sack, M., Vaquero, C, Faye, L., Fischer, R., Finnern R. and Lerouge, P. (2004) Recombinant anti−hCG antibodies retained in the endoplasmic reticulum of transformed plants lack core xylose and core D(l,3)−fucose residues. Plant Biotech. J., 2, 279−287
【非特許文献5】Triguero, A., Cabrera, G., Cremata, J., Yuen, C−T., Wheeler J. and Ramirez N.I. (2005) Plant−derived mouse IgG monoclonal antibody fused to KDEL endoplasmic reticulum−retention signal is TV−glycosylated homogeneously throughout the plant with mostly high−mannose−type N−glycans. Plant Biotech. J. 3, 449−457
【非特許文献6】Palacpac, N. Q., Yoshida, S., Sakai, H., Kimura, Y., Fujiyama, K., Yoshida, T. and Seki, T. (1999) Stable expression of human beta 1,4−galactosyltransferase in plant cells modifies N−linked glycosylation patterns. Proc. Natl. Acad. Sci. USA 96, 4692− 4697
【非特許文献7】Bakker, H., Bardor, M., Molhoff, J., Gomord, V., Elbers, I., Stevens, L., Jordi, W., Lommen, A., Faye, L., Lerouge, P. and Bosch D. (2001) Humanized glycans on antibodies produced by transgenic plants. Proc. Natl. Acad. ScL USA, 98, 2899−2904
【非特許文献8】Kelm, S. And Schauer, R. (1997) Sialic acids in molecular and cellular interactions. Int. Rev. Cytol. 175, 137−240
【非特許文献9】Matsumoto, S., Ikura, K., Ueda, M. and Sasaki, R. (1995) Characterization of a human glycoprotein (erythropoietin) produced in cultured tobacco cells. Plant MoI. Biol.27, 1163−1172
【非特許文献10】Wee, E.Q., Sherrier, D.J., Prime, T.A. and Dupree, P. (1998) Targeting of active sialyltransferase to the plant Golgi apparatus. Plant Cell, 10, 1759−1768
【非特許文献11】Seveno, M., Bardor, M., Paccalet, T., Gomord, V., Lerouge, P. and Faye, L. (2004)Glycoprotein sialylation in plants? Nature Biotech., 22, 5−6
【非特許文献12】Angatta, T. and Varki, A. (2002) Chemical diversity in the sialic acids and related a−keto acids: an evolutionary perspertive. Chem Rev., 102, 439−469
【非特許文献13】Maru, I., Ohnishi, J., Ohta, Y. and Tsukada, Y. (1998) Simple and large−scale production of N−acetylneuraminic acid from N−acetyl−D−glucosamine and pyruvate using N− acyl−D−glucosamine epimerase and N−acetylneuraminate layse. Carbohydr. Res.,306, 575−578
【非特許文献14】Tanner, M. E. (2005) The enzymes of sialic acid biosynthesis. Bioorg. Chem., 33, 216−228
【発明の開示】
【課題を解決するための手段】
【0007】
(発明の要旨)
本発明は、植物体におけるシアル酸の合成に関する。さらに、本発明は、シアル酸を産生する方法及び植物体、並びにこれら植物体から産生されるシアル化タンパク質に関する。
【0008】
本発明の目的は、植物体においてシアル酸を産生する改良された方法を提供することである。
【0009】
本発明によれば、
i)N−アセチルノイラミン酸(Neu5Ac)シンターゼ又はNeu5Acリアーゼをコードするヌクレオチド配列である、植物体で活性のある調節領域と作動可能に連結されたヌクレオチド配列を含む植物体を提供することと、
ii)植物体を生育させ、ヌクレオチド配列を発現させ、それによってシアル酸を合成することを含む、シアル酸、たとえば、Neu5Acの合成方法が提供される。
【0010】
さらに、生育させる工程の後、シアル酸を植物体から回収してもよい。調節領域は、構成的プロモーター、誘導性プロモーター、組織特異的プロモーター及び発育プロモーターからなる群より選択されてもよい。
【0011】
本発明はまた、上記で定義された方法(方法A)に関するものであり、その際、提供する工程では、植物体はさらに、植物体内で活性がある1又は1を超えた第2の調節領域に作動可能に連結された、エピメラーゼ、CMP−Neu5Acシンターゼ、CMP−Neu5Acトランスポーター、ガラクトシルトランスフェラーゼ及びシアリルトランスフェラーゼの1又は1を超えるものをコードする第2のヌクレオチド配列を含む。第2のヌクレオチド配列は、目的のヌクレオチド配列の発現と共に共発現される。さらに第2の調節領域は、構成的プロモーター、誘導性プロモーター、組織特異的プロモーター及び発育プロモーターからなる群より選択されてもよい。
【0012】
本発明はまた、上述の方法(方法A)に関するものであり、その際、Neu5Acシンターゼ若しくはNeu5Acリアーゼをコードするヌクレオチド配列、エピメラーゼ、CMP−Neu5Acシンターゼ若しくはCMP−Neu5Acトランスポーターの1若しくは1を超えるものをコードする第2のヌクレオチド配列、又は目的のヌクレオチド配列及び第2のヌクレオチド配列の両方は、植物体内での発現のためにコドンが最適化されている。
【0013】
本発明は、
i)Neu5Acシンターゼ、Neu5Acリアーゼ、エピメラーゼ、CMP−Neu5Acシンターゼ、CMP−Neu5Acトランスポーター、ガラクトシルトランスフェラーゼ及びシアリルトランスフェラーゼをコードする1又は1を超える第1のヌクレオチド配列、並びに目的のタンパク質をコードする第2のヌクレオチド配列を発現する植物体を提供することと、
ii)植物体を生育させ、第1及び第2のヌクレオチド配列を発現させ、それによって目的のタンパク質を産生し、その際、目的のタンパク質がシアル化されることを含む、
目的のタンパク質を産生する方法(B)を提供する。
【0014】
好ましくは、シアル化される目的のタンパク質は、ジ−、トリ−又はテトラ−分岐N−グリカンを含む。
【0015】
本発明はまた、上述の方法(方法B)に関するものであり、その際、シアル化タンパク質は植物体から抽出される。さらに、シアル化タンパク質は単離され、精製されてもよい。
【0016】
本発明は、植物体で活性のある調節領域に作動可能に連結されたNeu5Acシンターゼ又はNeu5Acリアーゼをコードするヌクレオチド配列を含む植物体、植物細胞又は種子を提供する。植物体、植物細胞又は種子は、植物内で活性のある1又は1を超える第2の調節領域に作動可能に連結された、エピメラーゼ、CMP−Neu5Acシンターゼ、CMP−Neu5Acトランスポーター、ガラクトシルトランスフェラーゼ及びシアリルトランスフェラーゼの1又は1を超えるものをコードする第2のヌクレオチド配列をさらに含んでもよい。さらに、調節領域及び第2の調節領域は、構成的プロモーター、誘導性プロモーター、組織特異的プロモーター及び発育プロモーターからなる群より選択されてもよい。
【0017】
本発明はまた、上述の方法(方法B)に関するものであり、その際、Neu5Acシンターゼ、Neu5Acリアーゼ、エピメラーゼ、CMP−Neu5Acシンターゼ、CMP−Neu5Acトランスポーター、ガラクトシルトランスフェラーゼ及びシアリルトランスフェラーゼをコードする1又は1を超える第1のヌクレオチド配列、目的のタンパク質をコードする第2のヌクレオチド配列、又は第1の配列及び第2の配列の両方は、植物体、植物細胞内又は種子内で発現させるためにコドンが最適化されている。
【0018】
本発明は、植物体で活性のある調節領域と作動可能に連結されたNeu5Acシンターゼ又はNeu5Acリアーゼをコードするヌクレオチド配列をコードするヌクレオチド配列を含む植物体、植物細胞又は種子を包含する。植物体、植物細胞又は種子はさらに、植物体内で活性がある1又は1を超えた第2の調節領域に作動可能に連結された、エピメラーゼ、CMP−Neu5Acシンターゼ、CMP−Neu5Acトランスポーター、ガラクトシルトランスフェラーゼ及びシアリルトランスフェラーゼの1又は1を超えるものをコードする第2のヌクレオチド配列を含んでもよい。さらに、Neu5Acシンターゼ若しくはNeu5Acリアーゼをコードするヌクレオチド配列、エピメラーゼ、CMP−Neu5Acシンターゼ、CMP−Neu5Acトランスポーター、ガラクトシルトランスフェラーゼ及びシアリルトランスフェラーゼの1又は1を超えるものをコードする第2のヌクレオチド配列、又は目的の配列及び第2の配列の両方は、植物体、植物細胞内又は種子内で発現させるためにコドンが最適化されている。
【0019】
本発明はまた、Neu5Acシンターゼ、Neu5Acリアーゼ、エピメラーゼ、CMP−Neu5Acシンターゼ、CMP−Neu5Acトランスポーター、ガラクトシルトランスフェラーゼ及びシアリルトランスフェラーゼをコードする1又は1を超える第1のヌクレオチド配列、目的のタンパク質をコードする第2のヌクレオチド配列を含む植物体、植物細胞又は種子を提供するものであり、第1及び第2のヌクレオチド配列は、植物体で活性がある1又は1を超える調節領域に作動可能に連結される。1又は1を超える調節領域は、構成的プロモーター、誘導性プロモーター、組織特異的プロモーター及び発育プロモーターからなる群より選択されてもよい。さらに、Neu5Acシンターゼ、Neu5Acリアーゼ、エピメラーゼ、CMP−Neu5Acシンターゼ、CMP−Neu5Acトランスポーター、ガラクトシルトランスフェラーゼ及びシアリルトランスフェラーゼをコードする1又は1を超える第1のヌクレオチド配列、目的のタンパク質をコードする第2のヌクレオチド配列、又は第1の配列及び第2の配列の両方は、植物体、植物細胞内又は種子内で発現させるためにコドンが最適化されている。
【0020】
本発明はまた、N−アセチルノイラミン酸(Neu5Ac)シンターゼ又はNeu5Acリアーゼをコードするヌクレオチド配列である、植物体で活性がある調節領域と作動可能に連結されたヌクレオチド配列によって植物体又は植物体の一部を一過性に形質転換することと、ヌクレオチド配列を発現させ、それによってシアル酸を合成することを含むシアル酸の合成方法(方法C)も提供する。さらに、Neu5Ac又はNeu5Acリアーゼは、植物体又は植物体の一部から回収されてもよい。
【0021】
本発明はまた、上述の方法(方法C)に関するものであり、その際、植物体又は植物体の一部を一過性に形質転換する工程はさらに、植物体内で活性がある1又は1を超える第2の調節領域に作動可能に連結された、エピメラーゼ、CMP−Neu5Acシンターゼ、CMP−Neu5Acトランスポーターの1又は1を超えるものをコードする第2のヌクレオチド配列を含み、第2のヌクレオチド配列は、目的のヌクレオチド配列の発現と共に共発現される。
【0022】
本発明は、
i)Neu5Acシンターゼ、Neu5Acリアーゼ、エピメラーゼ、CMP−Neu5Acシンターゼ、CMP−Neu5Acトランスポーター、ガラクトシルトランスフェラーゼ及びシアリルトランスフェラーゼをコードする1又は1を超える第1のヌクレオチド配列、及び目的のタンパク質をコードする第2のヌクレオチド配列を発現する構築物によって植物体又は植物体の一部を一過性に形質転換することと、
ii)目的のタンパク質を産生し、その際目的のタンパク質がシアル化されることを含む
目的のタンパク質を産生する方法(方法D)を提供する。
【0023】
シアル化される目的のタンパク質は、ジ−、トリ−又はテトラ−分岐N−グリカンを含んでもよい。さらに、シアル化タンパク質は植物体又は植物体の一部から抽出されてもよい。シアル化目的のタンパク質はまた単離され、精製されてもよい。
【0024】
本発明はまた、上述の方法(方法D)に関するものであり、その際、産生する工程の後、シアル化目的のタンパク質を含む植物物質は、被験体に経口投与される。たとえば、産生する工程の後、植物体又は植物体の一部を最小限に加工して最小限に加工された植物物質を作製してもよく、シアル化された目的のタンパク質を含む最小限に加工された植物物質を被験体に経口投与してもよい。
【0025】
本明細書で記載されるように、植物体におけるNeu5Ac合成酵素、Neu5Acリアーゼ及びNeuB2の発現は結果的に植物組織内に機能的酵素の蓄積を生じる。Neu5Ac合成酵素は、任意の植物体、たとえば、タバコ及びMedicago sativa(アルファルファ)、分子農法に関する幾つかの農業的利点から恩恵を受ける多年生のマメ科作物で発現されてもよいが、これらに限定されない(Busse, et al., 2001)。
【0026】
本発明の要約は、本発明の特徴を必ずしもすべて記載するわけではない。
【0027】
本発明のこれらの特徴及びそのほかの特徴は、添付の図面を参照する説明からさらに明らかになるであろう。
【発明を実施するための最良の形態】
【0028】
(詳細な説明)
本発明は、植物体におけるシアル酸の合成に関する。さらに、本発明は、シアル酸を発現する方法及び植物体、並びにこれら植物体から産生されるシアル化タンパク質を提供する。
【0029】
以下の記載が、好ましい実施態様である。
【0030】
本発明は、植物体内でN−アセチルノイラミン酸(Neu5Ac)を合成する方法を提供する。Neu5Acリアーゼは、可逆的反応においてNeu5AcをManNAcとピルビン酸塩に切断するのを触媒することによって細菌にてシアル酸を異化する。この反応は可逆的なので、適切な前駆体の存在下でNeu5Acを合成するのにNeu5Acリアーゼを使用してもよい。Neu5Acの産生に関する代替方法には、Neu5Acシンターゼの使用が関与する。Neu5Acシンターゼは、D−ManNAcとPEPとの縮合によってNeu5Acを形成するのを触媒する。
【0031】
従って、本発明は、Neu5Acシンターゼ又はNeu5Acリアーゼをコードするヌクレオチド配列であって、植物体で活性のある調節領域と作動可能に連結されたヌクレオチド配列を含む植物体を提供することと、植物体を生育させ、ヌクレオチド配列を発現させてNeu5Acを合成することを含むNeu5Acを合成する方法を提供する。或いは、方法には、植物体又は植物体の一部の内部におけるNeu5Acの一過性の産生が関与してもよい。
【0032】
そのように産生されるNeu5Acは、植物体から回収され、当該技術で既知の方法を用いたin vitroでのタンパク質のシアル化に使用されてもよい。或いは、Neu5Acは、植物体内で共発現される目的のタンパク質のシアル化のための内因性基質として使用されてもよい。
【0033】
所望であれば、植物体内でNeu5Acを合成するための、N−アセチルマンノサミン(D−ManNAc)を含むが、これに限定されない基質のレベルは、エピメラーゼ、CMP−Neu5Acシンターゼ、CMP−Neu5Acトランスポーターの1又は1を超えるものをコードする1又は1を超える追加のヌクレオチド配列を植物体内で共発現させることによって高められてもよい。たとえば、植物体内で、UDP−GlcNAc2−エピメラーゼ、たとえば、細菌のUDP−GlcNAc2−エピメラーゼ、又は内因性のUDP−GlcNAc2をManNAcに変換するそのほかの供給源からのエピメラーゼを発現させることによってManNAcを合成してもよい。或いは、ManNAc−6−リン酸を製造し、その後、ホスファターゼによって加水分解してもよい。このアプローチによって、GlcNAc−6リン酸2−エピメラーゼ、たとえば、細菌のGlcNAc−6リン酸2−エピメラーゼ、又は哺乳類のUDP−GlcNAc2−エピメラーゼ/ManNAcキナーゼが植物体内で発現される。Neu5Acシンターゼ又はNeu5Acリアーゼをコードするヌクレオチド配列の発現と共にこの第2のヌクレオチド配列を共発現させることによって、高いレベルのNeu5Acが産生される。しかしながら、1以上のこれら酵素の内因性の活性が植物体の内部に存在してもよいので、1以上の上記ヌクレオチド配列の共発現の必要性は、選択される宿主に依存する。
【0034】
細胞質のシアル酸からのN−グリカンのシアル化を確実にするには、細菌又は哺乳類のCMP−Nue5Acシンターゼ、哺乳類のCMP−Nue5Acトランスポーター、哺乳類のガラクトシルトランスフェラーゼ(シアル酸をN−グリカンに変換する前のガラクトースの添加にために)及び哺乳類のシアリルトランスフェラーゼを植物体内で共発現させてもよい。本発明に従って植物体内で産生されたNeu5Acは、CMP−Nue5Acシンターゼを介したCMP−N−アセチルノイラミン酸(CMP−Nue5Ac)の合成のための基質として使用されてもよく、CMP−Nue5Acは次いで、植物体内で共発現される目的のタンパク質のシアル化のための基質として使用される。この場合、植物体は、シアリルトランスフェラーゼをコードするヌクレオチド配列を含んでもよい。哺乳類のシアリルトランスフェラーゼ及び哺乳類のCMP−Nue5Acシンターゼの植物体における発現は実証されている(参照によって本明細書に組み入れられるWee et al., 1998, Misaki R,. et al., 2006)。しかしながら、1以上のこれら酵素の内因性の活性が植物体内で存在してもよいので、1以上の上記ヌクレオチド配列を共発現する必要性は、選択される宿主植物体に依存してもよい。
【0035】
ヌクレオチド配列が植物体内で共発現する場合、標準の形質転換手法である、一過性の形質転換手法を用いて所望のヌクレオチド配列のそれぞれを植物体に導入してもよいし、又は1以上の所望のヌクレオチド配列をそれぞれ発現する2種の植物体を交配してヌクレオチド配列の必要とされる組み合わせを共発現する植物体を得てもよい。
【0036】
従って、本発明はまた、シアル化された目的のタンパク質の産生のための基盤として使用されてもよい植物体を作製する方法も提供する。本方法は、Neu5Acシンターゼ、Neu5Acリアーゼ、エピメラーゼ、CMP−Neu5Acシンターゼ、CMP−Neu5Acトランスポーター、ガラクトシルトランスフェラーゼ及びシアリルトランスフェラーゼをコードする1又は1を超える第1のヌクレオチド配列を発現する植物体を提供することと、1以上のヌクレオチド配列を発現することを含む。目的のタンパク質を産生するために、標準技法、たとえば、形質転換法を用いて、目的のタンパク質をコードする第2のヌクレオチド配列を基盤植物体に導入し、第2のヌクレオチド配列を発現させるか、又は交配された植物体の子孫の中で産生された目的のタンパク質がシアル化されるように目的のタンパク質を発現している植物体と基盤植物体を交配させる。
【0037】
本発明はまた、Neu5Acシンターゼ、Neu5Acリアーゼ、エピメラーゼ、CMP−Neu5Acシンターゼ、CMP−Neu5Acトランスポーター、ガラクトシルトランスフェラーゼ及びシアリルトランスフェラーゼをコードする1又は1を超える第1のヌクレオチド配列並びに目的のタンパク質をコードする第2のヌクレオチド配列を発現する植物体を提供することと、植物体を生育させ、第1及び第2のヌクレオチド配列を発現させ、それによって目的のタンパク質を産生し、その際目的のタンパク質がシアル化されることを含む、目的のタンパク質を産生する方法にも関する。好ましくは、シアル化される目的のタンパク質は、ジ−、トリ−又はテトラ−分岐N−グリカンを含む。シアル化タンパク質は植物体から抽出してもよく、所望であれば、シアル化タンパク質は、常法を用いて単離及び精製してもよい。再び、所望の構築物によって植物体を安定的に形質転換してもよいし、又は、所望の構築物によって植物体又は植物体の一部を一過性に形質転換してもよい。
【0038】
Neu5Acシンターゼ、Neu5Acリアーゼ、エピメラーゼ、CMP−Neu5Acシンターゼ、CMP−Neu5Acトランスポーター、ガラクトシルトランスフェラーゼ及びシアリルトランスフェラーゼをコードするヌクレオチド配列は、コドンを最適化して植物体内での発現レベルを高めてもよい。コドンの最適化によって、植物体内でのコドン使用法にアプローチするために、構造遺伝子又はその断片のオリゴヌクレオチド基本単位の合成及びそれに続く酵素の集合のための適切なDNAヌクレオチドの選択を意味する。
【0039】
植物体内での外来配列の発現を最適化するために、野生型であっても合成配列であってもよいヌクレオチド配列が使用されてもよく、又は、相当するタンパク質、たとえば、Neu5Acシンターゼ、Neu5Acリアーゼ、エピメラーゼ、CMP−Neu5Acシンターゼ、CMP−Neu5Acトランスポーター、ガラクトシルトランスフェラーゼ及びシアリルトランスフェラーゼ、目的のタンパク質、若しくはそれらの組み合わせが、非修飾ヌクレオチド配列によってコードされる場合に産生されるよりも高いレベルで産生される。たとえば、限定について考慮しないが、配列は、合成配列であってもよく、たとえば、BLAST(GenBankを介して利用可能;初期設定パラメーターを用いて)のような、しかし、これに限定されない配列比較法を用いて決定されるように、野生型配列と少なくとも80%の相同性を含んで、植物体内でのコドン利用について配列が最適化されてもよい。たとえば、抗原特性のような、しかし、これに限定されない有用な生物特性を示す目的のタンパク質又はその誘導体をコードする配列の断片又は一部が植物組織の中で発現されてもよいことも企図される。
【0040】
Neu5Acシンターゼ、Neu5Acリアーゼ、エピメラーゼ、CMP−Neu5Acシンターゼ、CMP−Neu5Acトランスポーター、ガラクトシルトランスフェラーゼ、シアリルトランスフェラーゼ及び目的のタンパク質の発現レベル及び導入遺伝子のタンパク質産生をできるだけ多くするために、核酸配列を調べて、Sardana at al.(Plant Cell Reports, 15:677−681, 1996)が概説したものに類似する手順を用いてコーディング領域を修飾し、植物体における遺伝子の発現のために最適化してもよい。双子葉植物において高度に発現される遺伝子のコドン利用の表は、Murray et al (Nuc. Acids Res., 17:477−498, 1989)を含めて幾つかの供給元から利用可能である。
【0041】
従って、本発明は、N−アセチルノイラミン酸(Neu5Ac)シンターゼ又はNeu5Acリアーゼをコードするヌクレオチド配列であって、植物体で活性のある調節領域と作動可能に連結されたヌクレオチド配列を含む植物体を提供することと、植物体を生育させ、ヌクレオチド配列を発現させ、それによってシアル酸を合成することを含むシアル酸を合成する方法を提供する。Neu5Acシンターゼ又はNeu5Acリアーゼをコードするヌクレオチド配列は、植物体内での発現のためにコドンが最適化されてもよい。さらに、提供する工程では、植物体はさらに、植物体内で活性がある1又は1を超えた第2の調節領域に作動可能に連結された、エピメラーゼ、CMP−Neu5Acシンターゼ又はCMP−Neu5Acトランスポーターの1又は1を超えるものをコードする第2のヌクレオチド配列を含み、第2のヌクレオチド配列は、目的のヌクレオチド配列の発現と共に共発現される。エピメラーゼ、CMP−Neu5Acシンターゼ又はCMP−Neu5Acトランスポーターの1又は1を超えるものをコードする第2のヌクレオチド配列は、植物体内で発現されるためにコドンが最適化されてもよい。
【0042】
さらに、本発明は、Neu5Acシンターゼ、Neu5Acリアーゼ、エピメラーゼ、CMP−Neu5Acシンターゼ、CMP−Neu5Acトランスポーター、ガラクトシルトランスフェラーゼ、シアリルトランスフェラーゼをコードする1又は1を超える第1のヌクレオチド配列、及び目的のタンパク質をコードする第2のヌクレオチド配列を発現する植物体を提供することと、植物体を生育させ、第1及び第2のヌクレオチド配列を発現させ、それによって目的のタンパク質を産生する目的のタンパク質を産生する方法を提供する。Neu5Acシンターゼ、Neu5Acリアーゼ、エピメラーゼ、CMP−Neu5Acシンターゼ、CMP−Neu5Acトランスポーター、ガラクトシルトランスフェラーゼ、シアリルトランスフェラーゼをコードする1又は1を超える第1のヌクレオチド配列、目的のタンパク質をコードする第2のヌクレオチド配列、又は第1ヌクレオチド配列及び第2のヌクレオチド配列の両方は、植物体内で発現されるためにコドンが最適化されてもよい。
【0043】
さらに、本発明は、植物体で活性のある調節領域に作動可能に連結されたNeu5Acシンターゼ又はNeu5Acリアーゼをコードするヌクレオチド配列を含む植物体、植物細胞又は種子に関する。植物体、植物細胞又は種子は、植物内で活性のある1又は1を超える第2の調節領域に作動可能に連結された、エピメラーゼ、CMP−Neu5Acシンターゼ、CMP−Neu5Acトランスポーター、ガラクトシルトランスフェラーゼ及びシアリルトランスフェラーゼの1又は1を超えるものをコードする第2のヌクレオチド配列をさらに含んでもよい。Neu5Acシンターゼ又はNeu5Acリアーゼをコードするヌクレオチド配列、エピメラーゼ、CMP−Neu5Acシンターゼ、CMP−Neu5Acトランスポーターの1又は1を超えるものをコードする第2のヌクレオチド配列、又は該ヌクレオチド配列及び第2のヌクレオチド配列の両方は、植物体、植物細胞又は種子内で発現されるためにコドンが最適化されてもよい。
【0044】
本発明はまた、Neu5Acシンターゼ、Neu5Acリアーゼ、エピメラーゼ、CMP−Neu5Acシンターゼ、CMP−Neu5Acトランスポーター、ガラクトシルトランスフェラーゼ及びシアリルトランスフェラーゼをコードする1又は1を超える第1のヌクレオチド配列、目的のタンパク質をコードする第2のヌクレオチド配列を含む植物体、植物細胞又は種子を含み、第1及び第2のヌクレオチド配列は、植物体で活性がある1又は1を超える調節領域に作動可能に連結される。Neu5Acシンターゼ、Neu5Acリアーゼ、エピメラーゼ、CMP−Neu5Acシンターゼ、CMP−Neu5Acトランスポーター、ガラクトシルトランスフェラーゼ及びシアリルトランスフェラーゼをコードする1又は1を超える第1のヌクレオチド配列、目的のタンパク質をコードする第2のヌクレオチド配列、又は第1のヌクレオチド配列及び第2のヌクレオチド配列の両方は、植物体内で発現されるためにコドンが最適化されてもよい。
【0045】
「作動可能に連結する」によって、特定の配列が直接又は間接的に相互作用し、たとえば、遺伝子発現の介在又は修飾のような意図される機能を実行することを意味する。作動可能に連結された配列の相互作用は、たとえば、作動可能に連結された配列と相互作用するタンパク質が介在してもよい。転写調節領域が介在する又はそれによって修飾されるべき目的の配列の転写を行えるように配列が機能的に接続される場合、転写調節領域と目的の配列は作動可能に連結される。
【0046】
用語「植物物体」によって、植物体に由来する任意の物質を意味する。植物物体は、植物体全体、その組織、細胞又は任意の分画を含んでもよい。さらに、植物物体は、植物体の細胞内植物成分、細胞外植物成分、液体、又は固形抽出物、又はそれらの組み合わせを含んでもよい。さらに、植物物体は、植物体の葉、茎、果実、根、又はそれらの組み合わせからの植物体、植物細胞、組織、液体抽出物又はそれらの組み合わせを含んでもよい。植物物体は、加工工程の対象とされていない植物体又はその一部を含んでもよい。しかしながら、植物物体は、以下で定義するような最小限の加工工程、又はクロマトグラフィ、電気泳動などを含むが、これらに限定されない当該技術で共通して知られる技法を用いた部分的な又は実質的なタンパク質精製を含む綿密な工程の対象とされてもよいことも企図される。
【0047】
用語「最小限の加工」によって、植物抽出物、ホモジネート、ホモジネートの分画などを得るために部分的に精製される目的のタンパク質を含む植物物体、たとえば、植物体又はその一部を意味する。部分精製は、植物体の細胞構造を破壊し、それによって可溶性の植物成分、及びたとえば、それらに限定されないが、遠心分離、濾過又はそれらの組み合わせによって分離されてもよい不溶性の植物成分を含む組成物を創製することを含んでもよいが、これらに限定されない。この点で、葉又はそのほかの組織の細胞外空間に分泌されたタンパク質は、吸引又は遠心抽出を用いて容易に得ればよく、加圧下でローラーを通過させる又は粉砕するなどによって組織を抽出し、細胞外空間から離れたタンパク質を圧搾する又は遊離させればよい。最小限の加工には、可溶性タンパク質の粗抽出物の調製が関与すればよく、これらの調製物は、二次的な植物生成物からの混入が無視できるほどだからである。さらに、最小限の加工には、可溶性タンパク質の葉からの水性抽出、その後の好適な塩による沈殿が関与してもよい。そのほかの方法には、抽出物の直接的な使用を可能にするための大規模浸漬及び分泌液抽出が挙げられる。
【0048】
植物物質又は組織の形態での植物物体は、経口にて被験体に送達されてもよい。植物物体は、栄養補助食品の一部として他の食物とともに投与されてもよく、カプセルに内包されてもよい。植物物体又は組織は、濃縮して美味しさを改善する又は高めてもよく、必要に応じてそのほかの物質、成分又は医薬賦形剤とともに提供されてもよい。
【0049】
不均質な目的のタンパク質を含む植物体を、必要性と状況に応じて様々な方法で、被験体、たとえば、動物又はヒトに投与してもよいことが企図される。たとえば、タンパク質が経口で投与されるのであれば、植物体を収穫し、直接被験体に与えてもよいし、与える前に収穫した組織を乾燥してもよいし、収穫を行わずに動物が植物体を食べられるようにしてもよい。収穫された植物体組織に関する本発明の範囲内で、動物の餌の中に食品補助として提供されることも考慮される。植物体組織がほとんど又は全くさらに加工することなく動物に与えられるのであれば、投与される植物体組織は食用であることが好ましい。さらに、植物体から得られる目的のタンパク質は、食品補助として使用する前に、加工しない形態、部分精製した形態、又は生成した形態に抽出されてもよい。この後者の場合、タンパク質は食用又は非食用の植物体のいずれかから産生されればよい。
【0050】
さらに詳細には実施例で記載されるように、Neu5Acリアーゼ及びNeu5Acリアーゼ−FLAG(形質転換体で組換えタンパク質を免疫的に検出するためにC末端にてFLAGエピトープをタグとして付けたNeu5Acリアーゼ)を植物体に導入した。抗FLAG抗体を用いたウエスタンブロット分析によって、形質転換された細胞にMWr32kDaのタンパク質が存在することが明らかにされた(図1a)。Neu5Acリアーゼ又はNeu5Acリアーゼ−FLAGのいずれかを発現している植物体から得られた抽出物にて、in vitro及びin vivoの両方でリアーゼ活性が検出可能だった。非形質転換植物体にて内因性のリアーゼ活性は検出されなかった。しかしながら、D−ManNAcとピルビン酸塩の存在下にて組換え産生されたNeu5Acリアーゼを用いたNeu5Acの合成は認められた(図2bを参照のこと)。従って、組換え的に発現されたNeu5Acリアーゼは、植物体にて生物学的に活性がある。
【0051】
Neu5Acシンターゼ(たとえば、NeuB2、しかし、これに限定されない)及びNeu5Acシンターゼ−FLAG(形質転換体で組換えタンパク質を免疫的に検出するためにC末端にてFLAGエピトープをタグとして付けたNeu5Acシンターゼ)を植物に導入した。抗FLAG抗体を用いたウエスタンブロット分析によって、形質転換された細胞にMWr37kDaのタンパク質が存在することが明らかにされた。Neu5Acシンターゼ又はNeu5Acシンターゼ−FLAGのいずれかを発現している植物体から得られた抽出物にて、in vitro及びin vivoの両方でシンターゼ活性が検出可能だった。非形質転換植物体にて内因性のシンターゼ活性は検出されなかった。しかしながら、D−ManNAcとPEPの存在下にて組換え産生されたNeu5Acシンターゼを用いたNeu5Acの合成は認められた(図3b、cを参照のこと)。従って、組換え的に発現されたNeu5Acシンターゼは、植物体にて生物学的に活性がある。
【0052】
「類縁体」又は「誘導体」は、ヌクレオチド配列が、標的遺伝子又は配列の発現を停止する、標的配列の発現を低下させる、又は標的配列にコードされるタンパク質の合成又は活性を低下させる特性を保持するという条件で、サイレンシングヌクレオチド配列への置換、欠失又は付加を含む。たとえば、核酸配列の誘導体及び類縁体は通常、サイレンシング核酸配列に80%を超える類似性を示す。配列の類似性は、初期設定プログラム(プログラム:blastn、データベース:nr、Expect10、フィルター:低複雑性、アライメント:ペアで、ワールドサイズ:II)を用いたBLASTアルゴリズム(GenBank、ncbi.nlm.nih.gov/cgi−bin/BLAST)の使用によって決定されてもよい。その類縁体又は誘導体はまた、厳密なハイブリッド形成の条件下で(Maniatis et al., in Molecular Cloning (A Laboratory Manual), Cold Spring Harbor Laboratory, 1982, p. 387−389, 又はAusubel at al., (eds), 1989, Current Protocol in Molecular Biology, Vol. 1, Green Publishing Associates, Inc., and John Wiley & Sons, Inc., New Yolk p.2.10.3を参照のこと)配列が標的遺伝子の発現を停止する特性を示すという条件で本明細書に記載される配列の1つにハイブリッド形成するヌクレオチド配列も含む。そのような厳密なハイブリッド形成条件の例の1つは、7%SDS、1mMのEDTA、0.5MのNa2HPO4、pH7.2中で65ECにて16〜20時間、たとえば、[γ−32P]dATP標識されたプローブのような、しかし、これに限定されない好適なプローブとのハイブリッド形成であってもよい。続いて、5%SDS、1mMのEDTA、40mMのNa2HPO4、pH7.2で30分間洗浄し、続いて1%SDS、1mMのEDTA、40mMのNa2HPO4、pH7.2で30分間洗浄すること。この緩衝液での洗浄を繰り返してバックグランドを低減させてもよい。
【0053】
「調節領域」、「調節要素」または「プロモーター」によって、常とは限らないが、通常、遺伝子のタンパク質コーディング領域の上流にある核酸の部分を意味し、DNA又はRNAのいずれか、又はDNA及びRNAの両方で構成されてもよい。調節領域が目的の遺伝子によって活性を持つ場合、目的の遺伝子と作動関係にある場合、目的の遺伝子と作動可能に連結される場合、これが、結果として目的の遺伝子の発現を生じてもよい。調節要素は、臓器特異性又は発生上の若しくは一時的な遺伝子の活性化の制御に介在することが可能である。「調節領域」には、プロモーター要素、基本のプロモーター活性を示すコアプロモーター要素、外部刺激に反応して誘導可能である要素、たとえば、負の調節要素又は転写エンハンサーのようなプロモーター活性に介在する要素が含まれる。本明細書で使用されるとき、「調節領域」には、転写に続いて活性を持つ要素、たとえば、翻訳及び転写のエンハンサー、翻訳及び転写のリプレッサー、上流活性化配列並びにmRNA不安定性決定基のような遺伝子発現を調節する調節要素が挙げられる。これら後者の要素の幾つかは、コーディング領域の近傍に位置してもよい。
【0054】
この開示の背景で、用語「調節要素」又は「調節領域」は通常、常とは限らないが、普通、構造遺伝子の上流(5’)のDNAの配列を言い、RNAポリメラーゼ及び/又は特定の部位で開始する転写に必要とされるそのほかの因子に対して認識を提供することによってコーディング領域の発現を制御する。しかしながら、配列のイントロン又は3’の中に位置するそのほかのヌクレオチド配列も目的のコーディング領域の発現の調節に寄与してもよいことが理解されるべきである。RNAポリメラーゼ又は特定の部位からの開始を確実にするそのほかの転写因子に対して認識を提供する調節因子の例は、プロモーター要素である。すべてとは限らないが、ほとんどの真核生物のプロモーター要素は、普通、転写開始部位のおよそ25塩基対上流に位置するアデノシンとチミジンのヌクレオチド塩基対から構成される保存された核酸配列であるTATAボックスを含有する。プロモーター要素は、転写の開始に反応性である基本のプロモーター要素、同様に、遺伝子発現を加減するほかの調節要素(上記で列記したような)を含む。
【0055】
発育上調節されるもの、誘導可能なもの又は構成的なものを含めて幾つかの種類の調節領域が存在する。発育上調節される、又はその制御下で遺伝子の差次的発現を制御する調節領域は、特定の臓器又は組織の発育中、特定の時間でその臓器又は組織の中で活性化される。しかしながら、一部の発育上調節される調節領域は、特定の発育段階で特定の臓器又は組織で優先的に活性化されてもよく、それらは、発育調節的に活性を持ってもよいし、又は植物体内で同様に他の臓器又は組織における基本的なレベルで活性を持ってもよい。組織特異的な調節領域の例は、たとえば、具体的に見た調節領域には、ナピンプロモーター及びクルシフェリンプロモーターが挙げられる(Rask et al., 1998, J. Plant Physiol., 152:595−599; Bilodeau et al., 1994, Plant Cell 14:125−130)。
【0056】
誘導可能な調節領域は、誘導因子に反応して1以上のDNA配列又は遺伝子の転写を直接又は間接的に活性化することが可能であるものである。誘導因子の非存在下では、DNA配列又は遺伝子は転写されない。通常、誘導可能な調節領域に特異的に結合して転写を活性化するタンパク質因子は、不活性の形態で存在してもよく、次いで直接又は間接的に誘導因子によって活性形態に変換される。しかしながら、タンパク質因子は不在であってもよい。誘導因子は、たとえば、タンパク質、代謝産物、増殖調節剤、除草剤若しくはフェノール化合物のような化学物質、熱、寒冷、塩によって直接強いられる生理的ストレス、又は毒性要素又はウイルスのような病原体又は病原剤の作用を間接的に介したものであることができる。たとえば、噴霧する、水で薄める、加熱することによって又は類似の方法によって細胞又は植物体に誘導因子を外から適用することによって、誘導可能な調節領域を含有する植物細胞を誘導因子に暴露してもよい。誘導可能な調節領域は、植物遺伝子又は非植物遺伝子のいずれかに由来してもよい(たとえば、Garz C and Lenk I.R.P., Trends Plant Sci., 3:352−358, 1998)。強力な誘導性プロモーターの例には、テトラサイクリン誘導性プロモーター(参照によって組み入れられるCratz C., 1997, Ann. Rev. Plant Physiol. Plant Mol. Biol., 48:89−108)、ステロイド誘導性プロモーター(参照によって組み入れられるAoyama T. and Chua, N.H., 1997, Plant J., 2:397−404)、及びエタノール誘導性プロモーター(参照によって組み入れられるSalter M.G. et al., 1998, Plant Journal 16:127−132; Caddick M. X. et al., 1998, Nature Biotech., 16:177−180)、サイトキニン誘導性IB6及びCKII遺伝子(参照によって組み入れられるBrandstater I. and Kieber, J.J., 1998, Plant Cell 10: 1009−1019; Kakimoto T., 1996, Science, 274:982−985)及びオーキシン誘導性要素DR5(参照によって組み入れられるUlmasov, T. et al., 1997, Plant Cell, 9:1963−1971)が挙げられるが、これらに限定されない。
【0057】
構成的な調節領域は、植物体の種々の部分の全体にわたって、且つ植物体の生育全体にわたって継続して遺伝子の発現を指向する。既知の構成的な調節要素の例には、CaMV35S転写物(Odell et al., 1985, Nature, 313:810−812)、コメのアクチン1(Zhang at al., 1991, Plant Cell, 3:1155−1165)、アクチン2(An et al., 1996, Plant J., 10:107−121)、又はtms(参照によって本明細書に組み入れられる米国特許第5,428,147号)、及びトリオセホスフェートイソメラーゼ1(Xu et al., 1994, Plant Physiol., 106:459−467)遺伝子、トウモロコシのユビキチン1遺伝子(Cornejo et al., 1993, Plant Mol Biol., 29:637−646)、Arabidopsisのユビキチン1及び6の遺伝子(Holtorf et al., 1995, Plant Mol. Biol., 29:637−646)、及びタバコの翻訳開始因子4A遺伝子(Mandel et al., 1995, Plant Mol. Biol., 29:995−1004)に関連するプロモーターが挙げられる。用語「構成的な」は、本明細書で使用されるとき、構成的な調節領域の制御下にある遺伝子があらゆる種類の細胞にて同一レベルで発現されるが、該遺伝子は、多量の変異が認められることが多いにもかかわらず、広い範囲の細胞種で発現されることを必ずしも指し示さない。
【0058】
本発明の1又は1を超えるヌクレオチド配列は、本発明のヌクレオチド配列、又は構築物又はベクターで形質転換される好適な植物体宿主で発現されてもよい。好適な宿主の例には、アルファルファ、カノーラ、Brassica spp、トウモロコシ、タバコ、アルファルファ、ジャガイモ、朝鮮ニンジン、エンドウ豆、カラスムギ、コメ、大豆、小麦、大麦、ヒマワリ、及び綿を含む農作物が挙げられるが、これらに限定されない。
【0059】
従って、本発明は、植物体で活性のある調節領域と作動可能に連結された、Neu5Acシンターゼ又はNeu5Acリアーゼをコードするヌクレオチド配列を含む植物体、植物細胞又は種子も提供する。さらに植物体、植物細胞又は種子は、植物体内で活性がある1又は1を超えた第2の調節領域に作動可能に連結された、エピメラーゼ、CMP−Neu5Acシンターゼ、CMP−Neu5Acトランスポーター、ガラクトシルトランスフェラーゼ及びシアリルトランスフェラーゼの1又は1を超えるものをコードする第2のヌクレオチド配列を含んでもよい。
【0060】
本発明はまた、Neu5Acシンターゼ、Neu5Acリアーゼ、エピメラーゼ、CMP−Neu5Acシンターゼ、CMP−Neu5Acトランスポーター、ガラクトシルトランスフェラーゼ及びシアリルトランスフェラーゼをコードする1又は1を超える第1のヌクレオチド配列、目的のタンパク質をコードする第2のヌクレオチド配列を含む植物体、植物細胞又は種子を提供するものであり、第1及び第2のヌクレオチド配列は、植物体で活性がある1又は1を超える調節領域に作動可能に連結される。
【0061】
本発明の1以上のキメラ遺伝構築物は、さらに3’非翻訳領域を含むことができる。3’非翻訳領域は、ポリアデニル化シグナル及びmRNAのプロセッシング又は遺伝子発現を達成することが可能であるそのほかの調節シグナルを含有するDNA断片を含む遺伝子の部分を言う。ポリアデニル化シグナルは普通、mRNA前駆体の3’末端へのポリアデニル酸のトラックの付加を達成することを特徴とする。ポリアデニル化シグナルは、変異が珍しくはないが、標準形態5’−AATAAA−3’に対する相同性の存在によって一般に認識される。本発明の1以上のキメラ遺伝構築物は、必要に応じてさらに、エンハンサー、翻訳エンハンサー又は転写エンハンサーのいずれかを含むことができる。これらのエンハンサーは当業者に周知であり、ATG開始コドンと隣接する配列を含むことができる。開始コドンは、配列全体の翻訳を確実にするようにコーディング配列の読み取りフレームに同調しなければならない。
【0062】
好適な3’領域の非限定例は、たとえば、ノパリンシンターゼ(Nos遺伝子)のような(Ti)プラスミド遺伝子並びに大豆保存タンパク質遺伝子及びリブロース−1,5−ビスホスフェートカルボキシラーゼの小型サブユニット(ssRUBISCO)遺伝子のような植物遺伝子を誘導するアグロバクテリウム腫瘍のポリアデニル化シグナルを含有する3’転写非翻訳領域である。
【0063】
形質転換した植物細胞での同定に役立つように、本発明の構築物は、植物体で選抜可能なマーカーを含むようにさらに操作してもよい。有用な選抜可能なマーカーには、たとえば、ゲンタマイシン、ハイグロマイシン、カナマイシンのような抗生剤、又はホスホノトリシン、グリホサート、クロロスルフロンなどのような除草剤のような化学物質に耐性を提供する酵素が挙げられる。同様に、たとえば、GUS(β−グルクロニダーゼ)のような色変化によって同定可能な化合物の産生、又はたとえば、ルシフェラーゼ若しくはGFPのような発光を提供する酵素を使用してもよい。
【0064】
また、本発明の考慮される部分は、本発明のキメラ遺伝構築物を含有するトランスジェニック植物体、植物細胞又は種子である。植物細胞から植物体全体を再生させる方法も当該技術では既知である。一般に、形質転換された植物細胞は適切な培地で培養され、培地は、抗生剤のような選抜剤を含有してもよく、その際、選抜可能なマーカーを使用して形質転換した植物細胞の同定を円滑にする。いったん、カルスが形成されると、既知の方法に準じて適切な植物ホルモンを用いることによって芽形成を促し、芽を植物再生用の発根培地に移す。次いで、種子から、又は栄養繁殖技法を用いて、該植物体を使用して繰り返し世代を確立してもよい。組織培養を用いることなくトランスジェニック植物体を生成することもできる。
【0065】
形質転換又は一時的な発現の影響を受け易い宿主生物の範囲内での発現のために、本発明の調節要素を目的のコーディング領域と組み合わせてもよい。そのような生物には、単子葉植物及び双子葉植物両方の植物が挙げられるが、これらに限定されず、たとえば、トウモロコシ、シリアル植物、小麦、大麦、タバコ、Brassica、大豆、豆、エンドウ豆、アルファルファ、ジャガイモ、トマト、朝鮮ニンジン及びArabidopsisが挙げられるが、これらに限定されない。
【0066】
これら生物の一時的な発現、形質転換及び再生の方法は、当該技術で確立され、当業者に既知である。形質転換された植物体及び再生された植物体を入手する方法は本発明に決定的ではない。
【0067】
「形質転換」によって、遺伝子型として、表現型として、又はその両方で明らかにされる遺伝情報の種間の移転を意味する。キメラ遺伝構築物から宿主への遺伝情報の種間移転は、遺伝性であってもよく、遺伝情報の移転は安定であるとみなされてもよく、又は移転は、一時的であってもよく、遺伝情報の移転は遺伝性ではない。本発明はさらに、安定な発現系又は一時的な発現系のいずれかと共に使用するのに好適なキメラ遺伝子構築物を含む好適なベクターをさらに含む。
【0068】
Tiプラスミド、Riプラスミド、植物ウイルスベクターを用いて、直接的なDNA形質転換、微量注入、エレクトロポレーションによって本発明の構築物を植物細胞に導入することができる。そのような技法の概説については、たとえば、Weissbach and Weissbach, Methods for Plant Molecular Biology, Academic Press, New Yolk, VIII, pp 421−463, 1988; Geierson and Corey, Plant Molecular Biology, 2d, Ed(1988),;及びMIki and lyer, Fundamentals of Gene Transfer in Plants, in Plant Metabolism. 2dEd, DT, Dennis, DH Turpin, DD Lefebrve, DB, Layzell (eds), Addison Wesly, (Langmans社, London pp561−579(1997)を参照のこと。そのほかの方法には、直接的なDNAの取り込み、リポソームの使用、エレクトロポレーション、たとえば、プロトプラストの使用、微量注入、微量発射若しくはウイスカー、及び真空浸透が挙げられる。たとえば、Bilang et al., (Gene, 100:247−250, 1991), Scheid et al., (Mol. Gen. Genet.,228:104−112, 1991), guerche et al., (Plant Science 52:111−116, 1987), Neuhause et al., (Theor. Appl. genet., 75:30−36, 1987), Klein et al., Nature 327:70−73, 1987; Howell et al., (Science, 208:1265, 1980), Horsch et al., (Sceinec 227:1229−1231, 1985); DeBlock et al., Plant Physiology 91:694−701, 1989; Methods for Plant Molecular Biology (Weissbach and Weissback, eds, Academic Press Inc., 1988; Methods in Plant Molecular Biology (Scheler and Zielinski eds., Academic Press Inc., 1989); Liu and Lomonossoff (J. Virol. Meth., 105:343−348, 2002); 米国特許第4,945,050号、同第5,036,006号及び同第5,100,792号、1995年5月10日出願の米国特許出願出願番号第08/438,666号、及び1992年9月25日出願の同第07/951,715号(すべて参照によって本明細書に組み入れられる)を参照のこと。
【0069】
以下に記載されるように、一過性の発現方法を用いて本発明の構築物を発現させてもよい(参照によって本明細書に組み入れられるLiu Lomonossodd, 2002, Journal of Virological Methods 105:343−348を参照のこと)。これらの方法には、たとえば、アグロ−植菌又はアグロ−浸潤が挙げられるが、これらに限定されない。しかしながら、上で述べたように他の一過性の方法も使用してもよい。アグロ−植菌又はアグロ−浸潤のいずれかによって、所望の核酸を含むアグロバクテリウムの混合物は、組織、たとえば、葉の細胞間間隙、植物体(茎、葉及び花を含む)の空間、植物体(茎、根、花)のそのほかの部分、又は植物体全体に入る。上皮を越境させた後、アグロバクテリウムが感染し、細胞内にDNAのコピーを転移させる。t−DNAは、エピソームで転写され、mRNAが翻訳され、感染した細胞における目的のタンパク質の産生をもたらす。しかしながら、t−DNAの核への継代は一過性である。
【0070】
「目的の遺伝子」、「目的のヌクレオチド配列」又は「目的のコーディング領域」によって、宿主生物、たとえば、植物体の中で発現されるべき遺伝子、ヌクレオチド配列、又はコーディング領域を意味する。これらの用語は相互交換可能に使用される。そのような目的のヌクレオチド配列には、その産物が工業用の酵素、タンパク質、サプリメント、餌、食品又は餌と食品両方の用途での中立的な付加価値のある産物、又はその断片である遺伝子又はコーディング領域が挙げられるが、これらに限定されない。目的のヌクレオチド配列又は目的のコーディング領域はまた、薬学上活性のあるタンパク質、たとえば、増殖因子、増殖調節剤、免疫若しくはワクチン植菌に有用な抗体、抗原及びその断片、又はその誘導体をコードする遺伝子を含んでもよい。そのようなタンパク質には、IL−1〜IL−24、IL−26及びIL−27、サイトカイン、エリスロポイエチン(EPO)、インスリン、G−CSF、GM−CSF、hPG−CSF、M−CSF、又はこれらの組み合わせ、インターフェロン、たとえば、インターフェロン−α、インターフェロン−β、インターフェロン−γ、血液凝固因子、たとえば、因子VIII、因子IX、又はtPAhGH、受容体、受容体作動薬、抗体、神経ポリペプチド、インスリン、ワクチン、たとえば、上皮増殖因子、角化細胞増殖因子、形質転換増殖因子、増殖調節剤のような、しかし、これらに限定されない増殖因子、抗原、自己抗原、その断片、又はこれらの組み合わせが挙げられるが、これらに限定されない。
【0071】
目的の遺伝子が、植物体にとって直接又は間接的に毒性である産物をコードするのであれば、本発明の方法を使用することによって、目的の遺伝子を指向された組織内又は植物体の生育の所望の段階で選択的に発現させることによって、植物体全体にわたってそのような毒性を低減してもよい。
【0072】
目的のコーディング領域又は目的のヌクレオチド配列は、形質転換される、又は本発明のヌクレオチド配列、核酸分子、又は遺伝構築物を含む好適な植物体宿主で発現されてもよい。好適な宿主の例には、Arabidopsis、並びにカノーラ、Brassica spp、トウモロコシ、タバコ、アルファルファ、ジャガイモ、朝鮮ニンジン、エンドウ豆、カラスムギ、コメ、大豆、小麦、大麦、ヒマワリ、及び綿を含む農作物が挙げられるが、これらに限定されない。
【0073】
植物体におけるシアル酸の合成、たとえば、Neu5Acの合成は、組み換えNeu5Acリアーゼ又はNeu5Acシンターゼを発現させることによって実証された。E.coliに由来するNeu5Acリアーゼ及びC.jejuniに由来するNeuB2はそれぞれ、アグロバクテリウムが介在した形質転換によって、又は植物細胞で一過性に発現させた場合、タバコBY2細胞、アルファルファ植物体の細胞質で発現された。組換えタンパク質の分解が認められなかったということは、この区分で酵素が安定であることを示している。BY2細胞で発現されたNeu5Acリアーゼは、可逆的反応にてNeu5AcのManNAcとピルビン酸塩への開裂を触媒することができた。ピルビン酸塩とManNAcの存在下でNeu5Acの合成も認められた。植物体の細胞質のpH及び最も重要な作物の温度に一致するpH7及び25〜37℃にてNeu5Acリアーゼは生物学的に活性があった。さらに、外因性Neu5Acの存在下で行われた供給実験によって酵素が植物体で機能的であることが明らかにされた。
【0074】
Neu5Acシンターゼである、C.jejuni由来のNeuB2は、タバコBY2で発現された場合、D−ManNAcとPEPの存在下でNeu5Acを合成することが認められた。アルファルファ植物体でのNeuB2の発現によって結果として機能的酵素の蓄積が生じた。従って、植物体における微生物のNeu5Acリアーゼ及びNeu5Acシンターゼの発現は結果として、Neu5Acを合成できる酵素の細胞質での産生を生じる。
【0075】
適切なアミノ糖基質と共にNeu5Acリアーゼ又はNeu5Acシンターゼを供給するために、内因性GlcNAcをManNAcに変換することができるエピメラーゼを植物体で共発現させてもよい。この点で、タバコBY2細胞における機能的CMP−Neu5Acシンターゼ及びCMP−Neu5Acトランスポーターの発現が報告されている(Misaki et al., 2006)。NeuB2と共にCMP−Neu5Acシンターゼ、Neu5Acトランスポーター、又はCMP−Neu5Acシンターゼ及びNeu5Acトランスポーターの両方を共発現させることによって、Neu5Acの産生を高めてもよい。N−アセチルマンノサミン(ManNAc)の植物体内での合成は幾つかの方法によって達成されてもよい。たとえば、不可逆的反応にてUDP−GlcNAcをManNAcに変換するUDP−GlcNAc2エピメラーゼ、たとえば、細菌のUDP−GlcNAc2エピメラーゼを植物体内で発現させることによってManNAcを合成してもよい。UDP−GlcNAcは、N−グリカン合成経路を供給するので細胞質に存在する。そのほかの供給元からのGlcNAc2エピメラーゼを発現させることによってManNAcの合成を達成してもよい。或いは、ManNAc−6−ホスフェートを形成し、その後、ホスファターゼで加水分解してもよい(トランスジェニック植物体中で)。このアプローチによって、GlcNAc−6−ホスフェート2エピメラーゼ、たとえば、微生物のGlcNAc−6−ホスフェート2エピメラーゼ又は哺乳類のUDP−GlcNAc−2エピメラーゼ/ManNAcキナーゼが植物体内で発現される。
【0076】
細胞質のシアル酸からのN−グリカンのシアル化を確実にするには、細菌又は哺乳類のCMP−Neu5Acシンターゼ、哺乳類のCMP−Neu5Acトランスポーター、哺乳類のガラクトシルトランスフェラーゼ及び哺乳類のシアリルトランスフェラーゼを植物体内で共発現してもよい(Misaki, R. et al., 2006)。
【0077】
以下の実施例にて本発明をさらに説明する。
【実施例】
【0078】
方法
合成FLAG配列ポリペプチド(Arg−Tyr−Lys−Asp−Asp−Asp−Asp−Lys:配列番号1)に対するポリクローナル抗体をEurogentec(Seraing, Belgium)のウサギで調製した。C18Bond−ElutカートリッジはVarian(Sugarland, TX)から得た。タバコ細胞及びM.sativaのクローニング実験及び形質転換には、それぞれ、Escherichia coliDH−5α及びAgrobacterium tumefaciensLBA4404を用いた。Gomordら(1998)で記載されたように、Nicotiana tabacum Bright Yellow2(BY2)細胞を増殖させた。
【0079】
(Neu5Acリアーゼ及びNeuB2の遺伝子のクローニング及び植物体の発現ベクターの構築)
Neu5Acリアーゼ及びNeuB2の遺伝子をPCRで増幅した。Neu5Acリアーゼの遺伝子は、以下のプライマーを用いたPCRによってE.coliのK1ゲノムDNAから増幅させた:
リアーゼP1:
5’−AATAGGCCATTACGGCCATGGCAACGAATTTACGTGG−3’(配列番号6)、および
リアーゼP2:
5’−AATAGGCCGAGGCGGCCTCACCCGCGCTCTTGCAT−3’(配列番号7)。
neuB2遺伝子については、以下のプライマーを用いてDNA断片を得た:
neuB2−P1:
5’−AATAGGCCATTACGGCCATGAAAAAAACTTTAATC−3’(配列番号8)、および
neuB2−P2:
5’−AATAGGCCGAGGCGGCCTTACTCACGGATAAGCTC−3’(配列番号9)。
【0080】
これら増幅したDNAを、Neu5Acリアーゼ遺伝子用のプラスミドベクターpDNR−LIB及びneuB2遺伝子用のバイナリープラスミドベクターpCAMBIA2300のカリフラワーモザイクウイルス(CaMV)35Sプロモーターの下に置いた。CaMV35Sプロモーター、Neu5Acリアーゼ及びノパリンシンターゼ(Nos)ターミネーターの発現カセットをカナマイシン耐性遺伝子と共に植物発現ベクターpBLTI121(Pagny et al., 2000)に導入した。
【0081】
pBLTINeu5Acリアーゼ−FLAG及びpBLTIneuB2−FLAGプラスミドを生成するために、タンパク質のC末端にコードされたFLAGペプチドで遺伝子を増幅するように設計された以下の4つのプライマーを用いた:
リアーゼ−FLAG−P1:
5’−CGGGGTACCAGAATGGCAACGAATTTACGTGGC−3’(配列番号2)、および
リアーゼ−FLAG−P2:
5’−GCCGAGCTCTCACTTGTCATCGTCATCCTTGTAATCCATCCCGCGCTCTTGCATCAACTG−3’(配列番号3)、
neuB2−FLAG−P1:
5’−CGGGGTACCAGAGAGATGAAAAAAACTTTAATCATCGC−3’(配列番号4)、および
neuB2−FLAG−P2:
5’−GCCGAGCTCTCACTTGTCATCGTCATCCTTGTAATCCATCTCACGGATAAGCTCATCTTC−3’(配列番号5)。
【0082】
以下のプログラム:94℃にて1分間の変性、58℃にて1分間のアニーリング及び72℃にて3分間の重合を30サイクルによるPCRによってこれらの増幅された配列を生成した。PCR産物をpCR−BLUNTII−TOPO(登録商標)(Invitrogen)にてクローニングした。植物細胞で組換えタンパク質を発現する前に、我々は、配列決定によって修飾したcDNA構築物すべてを確認した。その後、挿入物をKpnIとSacIで消化し、KpnI−SacIで消化したpBLTI121にクローニングした。熱ショック形質転換を介して、各ベクター(pBLTI121又はpCAMBIA2300)をAgrobacterium tumefaciensのLBA4404株に導入した。
【0083】
(BY2細胞における発現)
タバコBY2細胞は、MurashigeとSkoog(1962)の培地で維持し、形質転換に用いた。pBLTI121に由来する構築物をアグロバクテリウム(LBA4404)(Hofgen and Willmitzer, 1988)に移入した。100μg/mLのカナマイシンを含有するYEB培地にてトランスジェニックアグロバクテリウム細胞を選抜し、Gromordら(1998)で記載されているように浮遊培養しているタバコの細胞を形質転換するのに用いた。抗生剤(カナマイシン100μg/mL及びセフォタキシム250μg/mL)を含有するMS培地にて形質転換体を選抜し、維持した。各形質転換体からゲノムDNA及びmRNAを調製し、PCR及びRT−PCRによって目的の遺伝子がタバコの浮遊培養細胞に挿入され、そこで発現されていることを確認した。免疫スクリーニングの後、組換えタンパク質を産生している微小カルスを用いてトランスジェニック細胞の浮遊培養を開始した(Gomord et al., 1998)。
【0084】
(アルファルファ植物体における発現)
以下の改変と共に、Tianら(Tian et al., 2002)に記載されるように本質的に、アルファルファの形質転換を行った。Agrobacterium tumefaciensAGL1を用いて、アルファルファ遺伝子型R2336を形質転換した。共培養の工程は、0.8〜1のODで未稀釈の培養物と共に行い、1.5%スクロースの代わりに3%のスクロースをSh2K培地で使用した。
【0085】
(Neu5Acリアーゼ及びNeuB2の活性をアッセイするための細胞抽出物の調製)
4日間培養したBY2浮遊培養細胞1g又はM.sativaの新鮮な葉600ngを回収して、プロテイナーゼ阻害剤(ペプスタチン1μg/mL、E64:1μg/mL及びPMSF:1mM、シグマ)を含有する溶液A(100mMのTris−HCl緩衝液(pH=7.4)にて粉砕した。細胞抽出物を4℃にて10000gで10分間遠心し、硫酸アンモニウム(最終濃度80%)でタンパク質を沈殿させ、スペクトラ/Por(10000Daで切り捨て)と共に、溶液B(Neu5Acリアーゼアッセイ用には100mMのTris−HCl緩衝液(pH=7.4)又はNeuB2用にはpH=8.5、及び10mMのMgCl2)に対して透析した。次いで、タンパク質を酵素アッセイ又は免疫検出に利用した。
【0086】
(Neu5Acリアーゼ−FLAG及びNeuB2−FLAGの免疫検出)
変性緩衝液(20mMのTris−HCl、pH6.8、3%のβ−メルカプトエタノール、5%(v/v)のグリセロール及び1%(w/v)のSDS)にてタンパク質を可溶化し、5分間煮沸して、15%ポリアクリルアミドゲル中でのSDS−PAGEによって分離した。次いで、タンパク質をニトロセルロース膜上に移した。免疫検出のために、FLAGエピトープに対して生成したウサギ抗血清によって膜を探査した。西洋ワサビのペルオキシダーゼを結合したヤギ抗ウサギ抗体と共にインキュベートし、続いて4−クロロナフトールを用いた顕色又は化学発光反応によってタンパク質を検出した。
【0087】
(Neu5Acリアーゼ及びシンターゼのアッセイ)
可溶性の酵素活性は、4mMのPEP、4mMのNADH、20mMのNaHCO3及び10mMのDTEと共に細胞抽出物をインキュベートすることによって測定される。10分後、340nmでの吸収の減少によってNADHの酸化を測定した。Neu5Acリアーゼのリアーゼ活性は、形質転換体細胞抽出物をNeu5Acと共にインキュベートした後、ManNAcの形成を測定することによってアッセイした。プロテイナーゼ阻害剤(ペプスタチン1μg/mL、E64:1μg/mL及びPMSF:1mM、シグマ)及び40mMのNeu5Acを含有する溶液B(100mMのTris−HCl緩衝液(pH=7.4)及び10mMのMgCl2)にて37℃で細胞抽出物を2時間インキュベートした。Neu5Acリアーゼのシンターゼ活性は、形質転換体細胞抽出物をManNAcとピルビン酸塩と共にインキュベートした後、Neu5Acの形成を測定することによってアッセイした。プロテイナーゼ阻害剤(ペプスタチン1μg/mL、E64:1μg/mL及びPMSF:1mM、シグマ)及び20mMのManNAcと40mMのピルビン酸塩を含有する溶液B(100mMのTris−HCl緩衝液(pH=7.4)及び10mMのMgCl2)にて37℃で細胞抽出物を2時間インキュベートした。NeuB2のシンターゼ活性は、形質転換体細胞抽出物をManNAcとPEPと共にインキュベートした後、Neu5Acの形成を測定することによってアッセイした。プロテイナーゼ阻害剤(ペプスタチン1μg/mL、E64:1μg/mL及びPMSF:1mM、シグマ)及び10mMのManNAcと10mMのPEPを含有する溶液B(100mMのTris−HCl緩衝液(pH=7.4)及び10mMのMgCl2)にて37℃で細胞抽出物を2時間インキュベートした。80℃にて5分間加熱することによって反応を停止し、C18Bond−Eluntカートリッジにて水による連続溶出によって精製し、GC−EI−MS分析用に凍結乾燥して得た。
【0088】
(供給実験)
4日間培養したタバコBY2細胞を10mMのNeu5Ac又は30mMのManNAcと共にBY2培地中で37℃にて2日間インキュベートし、それぞれ、Neu5Acリアーゼ又はNeu5Acシンターゼのin vivoでの活性をアッセイした。2日後、Neu5Ac又はManNAcを含まないBY2培地でBY2細胞を洗浄し、回収した。70%エタノール中にて70℃で15分間加熱し、酵素を不活化し、次いで陶器のホモゲナイザーで粉砕した。70℃にて70%エタノールで2回、ホモジネートを洗浄した。残ったペレット及び上清をそれぞれ、細胞壁及び細胞質の遊離の単糖類の代表とみなした。次いで、ガスクロマトグラフィによって上清分画の単糖類を分析した。
【0089】
(GC分析)
酵素のアッセイについては、反応混合物を先ずC18Seppakカートリッジにおける精製工程に提示した。単糖類は100%の水中に溶出された。凍結乾燥の後、500μLの2Mの無水メタノール−HClによる80℃での16時間の加メタノール分解に試料を提示した。メタノールを蒸発した後、20μLの無水酢酸及び20μLのピリジンを添加することによって試料を再びアセチル化した。得られたN−アセチルメチルグリコシド(メチルエステル)を乾燥し、次いでそのTMS−誘導体に変換し、ガスクロマトグラフィで分離した。ガスクロマトグラフは、水素炎イオン化検出器、固定相としてのCP−Sil5CPを伴ったWCOT溶融石英のキャピラリーカラム(長さ25m、i.d.0.25mm)及びガスベクターとしてのヘリウムを備えていた。オーブンの温度プログラムは、120℃にて2分間、1分間当たり10℃で160℃まで、1分間当たり1.5℃で220℃まで、次いで1分間当たり20℃にて280℃までであった。ピークの積分及び標準の単糖類によって確立された反応因子を用いた相当する分子の値の決定によって糖の定量を行った。
【0090】
(植物における一過性発現)
(アグロバクテリウムの増殖)
上述の所望のDNA構築物を担うバイナリーベクターを含有するアグロバクテリウムのクローンは、それぞれ25μg/mL及び50μg/mLのカルベニシリン及びカナマイシンを含有する2mLのYEB培地又はLB培地にて28℃で24時間増殖させた。これら培養物10μLを出発植菌として用いて、25mLのYEB誘導培地(YEB培地、pHを5.6に調整した10mMの2(N−モルフォリノ)エタンスルホン酸(MES)、25mg/Lのカルベニシリン、20μMのカナマイシン)の培養を生成した。回転振盪器(220rpm)インキュベーターにて28℃で18時間、又は600nmでの光学密度が0.8〜1に達するまで後者を増殖させた。
【0091】
(非トランスジェニックタバコの生育)
温室にてNicotiana benthamiana及びNicotiana tabacumの植物体を泥炭系基材中で種子から生育させた。苗木は当初苗床で育て、その後ポットに移した。植物体には一日2回水を与え、各散布にて180ppmの窒素を与えた。温室の条件は、植物体のレベルで20ワット/m2の人工照明にて長日条件(16時間明/8時間暗のサイクル)のもと、昼間25℃及び夜間21℃に保持された。様々な生育段階で植物体を利用することができるが、5〜8週の生育にて優先的に選択された。
【0092】
(タバコにおける構築物の一過性の発現)
本発明では2つの一過性の発現の方法:アグロ−植菌又はアグロ浸潤が使用された。両方の方法では、目的の転移DNA(t−DNA)を担う2つ又は3つのアグロバクテリウムの培養物の混合物が、葉の細胞間間隙に入ることが強いられる。上皮の物理的障壁をいったん越境すると、アグロバクテリウムは近隣の細胞に感染し、植物細胞にt−DNAのコピーを転移させる。この方法によって、核内のt−DNAの継代は一過性であり、t−DNAに存在する遺伝子はエピソームに転写され、mRNAが翻訳され、感染細胞における目的のタンパク質の産生がもたらされる。アグロ浸潤が制御された減圧を用いるのに対して、アグロ植菌法は、シリンジによる加圧を用いてアグロバクテリウム混合物を植物体の内部に挿入する。
【0093】
前に記載されたように調製されたアグロバクテリウムの培養物を、10000gにて8分間遠心し、同一容量の植菌培地(pH5.6に調整され、100μMのアセトシリンゴンで補完された10mMのMgCl2、10mMのMES)に再浮遊し、植菌に先立って室温(RT、23℃)にて1時間保持した。或いは、植菌に先立って4℃にて24時間、浮遊物を保持した。N.benthamiana及びN.tabacumの一過性の形質転換は、以下の改変と共に、Liu及びLomonossoff(2002, Journal of Virological Methods, 105:343−348)に記載されたように本質的に行った。上述の構築物の発現のために、2種のアグロバクテリウム株の混合物を植菌した。第1の株は上述のクローンの1つを含有し、第2の株は、35Sプロモーターの制御のもとでジャガイモウイルスYからのサイレンシングのHcProサプレッサーを含有した。植菌後、植物体を温室でインキュベートした。温度は、昼間、最低23℃及び夜間21℃に保持された。植物体には一日2回水を与え、各散布にて180ppmの窒素を与えた。バイオマスの回収は、4〜8日後に行った。
【0094】
(形質転換されたバイオマスからの可溶性タンパク質抽出物の調製)
バイオマスを回収した後、又は−80℃にて凍結した後、葉を直接分析した。約0.1〜1gのアグロ−植菌された又はアグロ−浸潤させた葉のバイオマスを秤量し、総タンパク液抽出物の生成に使用した。
【0095】
幾つかの抽出方法を用いて、乳鉢と乳棒によって植物組織を粉砕することによって、ポリトロンを用いることによって、又はRetsch製のMixerMill300(MM300)にて粉砕することによって総タンパク抽出物を生成した。0.1〜1gの植物バイオマスを清浄で予備冷却した乳鉢に移した。冷却した抽出緩衝液(2mMのCaCl2及び4%のブタノールを含有する50mMのTris、150mMのNaCl、pH7.4)を1:3(w/v)の定量にて加え、同様に最終濃度、それぞれ1mM及び10μMのPMSF及びキモスタチンを加えた。均質な調製物が得られるまで、乳棒で葉を粉砕した。次いで植物抽出物を1.5mLの微量チューブに移し、4℃にて20,000gで20分間遠心した。或いは、0.1gの植物組織を0.3mLの抽出緩衝液と共に非滅菌の1.5mLの微量チューブに導入した。各チューブにタングステンビーズを加えた。30Hzでの3分間の撹拌サイクルにボックスを提示した。サイクルを2回繰り返した。次いで、植物抽出物を上述のように遠心した。或いは、ポリトロンを用いて、1gのバイオマスを3mLの抽出緩衝液と共に粉砕した。
【0096】
遠心に続いて、上清を清浄な微量チューブに移し、氷上に保持した。最終的に、BSAを参照タンパク質として用いたBradford法によって個々のタンパク抽出物の総タンパク質含量を測定した。
【0097】
(実施例1)
(タバコBY2細胞におけるE.coliのNeu5Acリアーゼの発現)
Escherichia coliK1(受入番号:D00067)由来のNeu5Acリアーゼをコードする遺伝子をタバコBY2細胞に導入した。E.coliK1のNeu5Acリアーゼを含有するプラスミドpBLTI121によるアグロバクテリウムを介在させた形質転換の後、トランスジェニックBY2のカルスを生成した。別の構築物はそのC末端にFLAGエピトープでタグを付け、形質転換体における組換えタンパク質を免疫的に検出できるようにした。カナマイシン耐性によって選抜された形質転換体をmRNAについてRT−PCRによって分析した。Neu5Acリアーゼ転写物を発現している48形質転換体から36及びNeu5Acリアーゼ−FLAG転写物を発現している50形質転換体から30が得られた。さらに高いmRNAの発現レベルを抱くカルスを、Neu5Acリアーゼ活性の性状分析用の浮遊培養に移した。Neu5Acリアーゼ−FLAGの存在は、ウエスタンブロット分析によって、形質転換されたBY2細胞の細胞質タンパク質抽出物にて割り出された。抗FLAG抗体を用いて形質転換した細胞にて32kDaの見かけのMWを持つ単一のタンパク質バンドが特異的に免疫検出された(図1a)。
【0098】
Neu5Acリアーゼ及びNeu5Acリアーゼ−FLAGを発現している浮遊培養されたBY2細胞の可溶性タンパク抽出物にて酵素的分析を行った。抽出物は両方共にリアーゼ活性を示した。タグの付いていないリアーゼを発現している細胞から単離されたタンパク抽出物にてさらなる分析を実施した。先ず、これらの抽出物をNeu5Acの存在下でインキュベートしてリアーゼ活性を調べた。図1b及び1cは、pH7及び37℃にてNeu5Acの非存在下(図1b)又は存在下(図1c)にてNeu5Acリアーゼタンパク質抽出物をインキュベートすることによって形成された最終産物のGC特性を示す。抽出物をNeu5Acと共にインキュベートした場合、3つのシグナル(ピーク1は内因性シグナルの肩部である)が明瞭に検出された。これらのシグナルは、標準のManNAcのピラノース型(ピーク1)及びフラノース型(ピーク2及び3)に類似する保持時間で溶出した。試料の電子衝撃質量分析(GC−EI MS)と併せたガスクロマトグラフィによって、これらシグナルがManNAcp(図1e)及びManNAcf(図1f)の1−O−メチルペルシリル誘導体に対するシグナルに割り振られることが確認された。m/z=173及び186での特徴的なイオンは、普通、アミノ糖について認められるような窒素原子を含有する断片に割り振られた。
【0099】
同様の条件で野生型のタバコBY2細胞に由来する細胞質のタンパク抽出物をNeu5Acとインキュベートしても、ManNAcを形成しなかった(データを示さず)ということは、植物体において内因性のリアーゼが存在しないことを実証している。このデータは、E.coliのNeu5Acリアーゼ遺伝子によって形質転換されたBY2細胞が、Neu5AcをD−ManNAcに開裂できる機能的な酵素を発現したことを示している。
【0100】
組換え酵素の最適pHは、4〜10のpHの範囲で実施されたアッセイにおいて生成されたD−ManNAcのGCの定量化に基づいて約7であることが明らかにされた(データは示さず)。さらに、組換え酵素は、25〜37℃の範囲で見られる高いリアーゼ活性を持つ温度依存性の活性を示した。pH7及び37℃にて、形質転換細胞に由来する可溶性タンパク質抽出物は1時間で、10μMのNeu5Acから0.5μMのManNAcを形成した。15℃未満では、残留する活性のみが検出された。
【0101】
pH7及び37℃にてD−ManNAc及びピルビン酸塩と共に、形質転換されたタバコBY2細胞のタンパク抽出物をインキュベートすることによってNeu5Acを合成する組換えNeu5Acリアーゼの能力が割り出された。図2a及び図2bはそれぞれ、基質の非存在下又は存在下でインキュベートした後の最終産物のGC特性を示す。対照の特性(図2a)と比べた場合、D−ManNAcとピルビン酸塩の存在下で行った反応のGC特性は、Neu5Acについて予測される保持時間でシグナルを示した(図2bでの四角)。このシグナルの電子衝撃質量スペクトル(EI−MS)(図2c)は、窒素含有断片に割り当てられたm/z=186でのイオンと同様にNeu5Ac断片に特異的なm/z=298及び420での特徴的なイオンを示した。このデータは、組換えリアーゼがD−ManNAcとピルビン酸塩の存在下でNeu5Acを合成することができることを示している。
【0102】
10mMのNeu5AcをタバコBY2細胞に供給することによってNeu5Acリアーゼの植物体での活性を測定した。Neu5AcのタバコBY2細胞に対する毒性を調べたところ、ヨウ化プロピジウム及びフルオレセインジアセテートを用いて細胞の生存率を調べることによって48時間にわたって毒性効果は認められなかった。23℃〜37℃の範囲の温度で48時間後のGCにより細胞質の単糖類を分析することによってD−ManNAcの形成を測定した。D−ManNAcはあらゆる処理で検出された(図1d)。GCによるD−ManNAcの定量は、23℃に比べて37℃にてこのアミノ糖の含量で25倍の増加を示した。これらin vivoの実験は、Neu5Acリアーゼが植物体で生物学的に活性を持ち、外因的に供給された基質に作用することができることを明らかにしている。
【0103】
(タバコBY2及びアルファルファ植物体におけるCampylobactor jejuniのNeuB2の発現)
Neu5Acシンターゼである、Campylobactor jejuniに由来するNeuB2(受入番号:NC002163)は、ManNAcとPEPの縮合によるNeu5Acの形成を触媒する。neuB2のDNAを含有するプラスミドpBLTI121によるアグロバクテリウムが介在する形質転換の後、トランスジェニックBY2カルスを生成した。タンパク質の免疫的検出のために、第2の構築物にはそのC末端にFLAGエピトープによるタグを付けた。カナマイシン耐性について選抜した形質転換体をRT−PCRによってmRNAレベルで分析した。最も高いmRNA発現レベルを抱くカルスを分析用の浮遊培養に移した。次いで、NeuB2−FLAGの配列によって形質転換されたBY2細胞から単離した可溶性タンパク抽出物のウエスタンブロット分析によって形質転換されたBY2細胞におけるNeuB2の蓄積を測定した。図3aで説明されるように、抗FLAG抗体は、シンターゼの予測分子量と一致するMW=37kDaにて単一タンパク質バンドを特異的に認識した。neuB2はまた、アグロバクテリウムが介在する形質転換及びin vitroの植物体の再生によってアルファルファ植物体にも導入された(Tian at al., 2002)。34の形質転換された植物体から、29がneuB2の転写物を発現していることが明らかにされた。
【0104】
細菌のNeu5Acシンターゼを発現している、形質転換された細胞及び植物体を分析する前に、内因性のNeu5Acシンターゼ活性の存在を調べた。野生型のタバコBY2細胞及びアルファルファ植物体から得た可溶性タンパク抽出物をD−ManNAc及びPEPと共にインキュベートした。アッセイで形成された単糖類をGCによって分離し、GC−EI−MSによって性状分析した。Neu5Acに割り当てられたピーク及びEI−MSでの特徴的なイオンが検出されなかったということは、植物体がPEPとManNAcを縮合してNeu5Acを形成することができる内因性の酵素を発現していないことを示している。
【0105】
neuB2遺伝子によって形質転換されたタバコBY2細胞及びアルファルファ植物体から単離した可溶性タンパク抽出物と共にD−ManNAcとPEPをインキュベートすることによって植物体で発現された組換えNeuB2のシンターゼ活性を測定した。図3b及び3cは、pH8及び37℃にてD−ManNAcとPEPの非存在下(図3b)又は存在下(図3c)で形質転換されたアルファルファ抽出物をインキュベートすることによって得たGC特性を示す。シンターゼの基質と共にインキュベートした後、Neu5Acについて予測される保持時間にて溶出されたピークが特異的に検出された。このピークのEI−MSは、m/z=298及び420で特徴的なイオンと共に、標準のNeu5Acの1つに類似する断片化パターンを示した。D−ManNacの非存在下でインキュベートした後のGC特性の相当する領域のEI−MSスペクトルにはそれらのイオンは検出されなかった(図3b)。
【0106】
Neu2B又はNeu2B−FLAG配列を発現しているタバコBY2細胞の分析によって同じ結果が得られた。
【0107】
従って、タバコBY2細胞及びアルファルファ植物体両方におけるneuB2の発現は、結果として機能的なNeu5Acシンターゼの産生を生じる。
【0108】
【表1−1】
【0109】
【表1−2】
【0110】
【表1−3】
【0111】
【表1−4】
引用はすべて参照によって本明細書に援用される。
【0112】
1以上の実施態様に関して本発明を説明してきた。しかしながら、クレームで定義されるような本発明の範囲から逸脱することなく多数の変化及び改変が可能であることが、当業者に明らかになるであろう。
【図面の簡単な説明】
【0113】
【図1】図1aは、抗FLAG抗体を用いた、野生型(ライン1)又はNeu5Acリアーゼ−FLAGを発現しているトランスジェニックタバコBY2細胞(ライン2)から抽出した可溶性タンパク質のウエスタンブロット分析を示す。図1b及び図1cは、Neu5Acを発現しているタバコBY2から単離された細胞質タンパク質を単独で(図1b)又はNeu5Acと共に(図1c)pH7にて37℃でインキュベートした後、得られた最終産物のガスクロマトグラフィの特性を示す。図1dは、10mMの外因性Neu5Acと共に37℃にて48時間養ったNue5Acを発現しているタバコBY2細胞の細胞質単糖類のGC特性を示す。図1e及び図1fは、特性(図1c)で検出されたピーク1(図1e)並びにピーク2及び3(図1f)の電子衝撃質量スペクトルを示す。D−ManNAcの1−O−メチルペルシリル誘導体の主要な断片イオンが示される。
【図2】図2a及び図2bは、Neu5Acを発現しているタバコBY2から単離された細胞質タンパク質を単独で(図2a)又はD−ManNAc及びピルビン酸塩と共に(図2b)pH7にて37℃でインキュベートした後、得られた最終産物のガスクロマトグラフィの特性を示す。図2cは、特性(図2b)で現れているピークの電子衝撃質量スペクトルを示す。N−アセチルノイラミン酸の1−O−メチルメチルエステルペルシリル誘導体の主要断片イオンが示される。
【図3】図3aは、抗FLAG抗体を用いた、野生型(ライン1)又はNeuB2−FLAGを発現しているトランスジェニックタバコBY2細胞(ライン2)から抽出した可溶性タンパク質のウエスタンブロット分析を示す。図3b及び図3cは、NeuB2を発現しているアルファルファ植物体の葉から単離された可溶性タンパク質を単独で(図3b)又はD−ManNAc及びPEPと共に(図3c)pH8にて37℃でインキュベートした後、得られた最終産物のガスクロマトグラフィの特性を示す。図3dは、特性(図3c)で現れているピークの電子衝撃質量スペクトルを示す。N−アセチルノイラミン酸の1−O−メチルメチルエステルペルシリル誘導体の主要断片イオンが示される。
【技術分野】
【0001】
(発明の分野)
本発明は、植物体におけるシアル酸の合成に関する。さらに、本発明はシアル酸を産生する方法及び植物体を提供し、これら植物体から産生されるシアル化タンパク質を提供する。
【背景技術】
【0002】
(発明の背景)
植物は、組換え医薬タンパク質を製造するための、コストの低い、且つ混入汚染について安全な製造現場となる可能性がある。アミノ酸配列、立体配置及び生物活性に関する限り、植物で産生された組換えタンパク質のほとんどは哺乳類で製造されたものと区別できない。さらに、哺乳類の糖タンパク質は、それらがトランスジェニック植物で発現された場合、効率的にグリコシル化される。しかしながら、植物は、動物の糖タンパク質で見い出されるものとは異なるN−グリカンを持つ分子を産生する(非特許文献1)。このことは、これらのタンパク質上の植物特異的な糖エピトープがヒトにおいて免疫応答を誘発する(非特許文献2)と共に、シアル化配列のような哺乳類型のエピトープが存在しないことが血流からのそれらの迅速なクリアランスを誘導する可能性があるので、植物で作製された医薬品の使用を限定する可能性がある。その結果、植物で作製された医薬のN−グリコシル化を制御することがヒトの治療法で使用することの前提条件となっている。
【0003】
最近、植物体でのリモデリング戦略が出現し、ヒトと適合可能な糖質特性を持つ植物由来の抗体が得られている。作戦の一部には、小胞体におけるプランティボディの保持が関与し(非特許文献3;非特許文献4;非特許文献5)、他の作戦には、哺乳類のグリコシルトランスフェラーゼによる植物の形質転換が関与していた。たとえば、植物のN−グリコシル化は、ヒトのβ(1,4)ガラクトシルトランスフェラーゼ(非特許文献6;非特許文献7)による植物の形質転換によって部分的にヒト化することができる。形質転換された植物におけるマウスの抗体の発現は結果として、相当するマウスのIgGで認められるものに類似するガラクトシル化特性を抱く植物由来の抗体を産生した(非特許文献7)。
【0004】
哺乳類のIgGは、Fcドメインに位置するNグリコシル化の保存された部位で二分岐N−グリカンを持つ。これらのオリゴ糖は弱くシアル化されており、末端Neu5Acが存在しないので、抗体の機能や安定性を妨害することはない。対照的に、そのほかの循環性糖タンパク質はほとんどシアル化された2、3又は4分岐のN−グリカンを有する。これらグリカンにおける末端シアル酸の存在は、多数の生体機能に必要とされ、その第1は、循環系におけるタンパク質の半減期の制御である。末端シアル酸の非存在下では、糖タンパク質は肝臓のアシアロ糖タンパク質受容体によって検出され、血清からクリアランスされ、これらのタンパク質の半減期を短く、無効なものにする(非特許文献8)。従って、非シアル化植物によって作製された医薬は、ヒトに注射された場合、血流から迅速に除かれる可能性があり、たとえば、タバコに由来するEpoはin vitroでは生物学的に活性を持っていたが、赤血球造血組織に到達する前に循環から除かれるためにin vivoでは、非機能性だった(非特許文献9)。
【0005】
プランティボディに連結するN−グリカンのヒト様N−グリカンへのリモデリングは、共基質として内因性UDP−Galを使用するトランスフェラーゼである、ヒトのβ(1,4)ガラクトシルトランスフェラーゼ(非特許文献6および7)の発現によって、植物ではすでに部分的に達成されている。哺乳類のシアリルトランスフェラーゼも植物に導入されており、機能性であり、正確にゴルジ装置を標的とすることが実証されている(非特許文献10)。しかしながら、内因性オリゴ多糖類のシアル化は認められなかった。植物におけるシアル酸及びシアル化機構の存在は依然として議論のあることころである。しかしながら、ヒトに存在する主なシアル酸であるNeu5Ac、同様にその前駆体、N−アセチルマンノサミン(D−ManNAc)は、検出可能な量で植物にて合成されるとは思われない(非特許文献11)。その結果、植物のN−グリカンのシアル化オリゴ糖へのグリコ操作は、合成を触媒することが可能な内因性酵素の共発現、Neu5Acの活性化及びゴルジ装置への移行を必要とする。
【0006】
哺乳類及び細菌では、Neu5Acの同化作用及び異化作用は、異なった経路を介して生じる(非特許文献12)。主な部類の2つの酵素がNeu−5Acを形成するのに必要とされる。N−アセチルノイラミネートリアーゼ(Neu5Acリアーゼ)は、可逆的反応におけるNeu5AcのN−アセチルマンノサミン(D−ManNAc)とピルビン酸塩への開裂を触媒することによってシアル酸の異化作用に関与する。高濃度のD−ManNAcとピルビン酸塩にて、平衡は、Neu5Acの合成に移行しうる。グルコサミン2−エピメラーゼ活性に連動して、E.coli由来のNeu5AcリアーゼをD−GlcNAcからのNeu5Acの大量生産に利用した(非特許文献13)。或いは、NeuBのようなNeu5Acシンターゼは、ピルビン酸ホルホエノール(PEP)へのManNAcの縮合を触媒し、シアル酸の生合成に直接関与する(非特許文献14で概説された)。
【非特許文献1】Lerouge, P., Cabanes−Macheteau, M., Rayon, C, Fitchette−Laine, A.−C, Gomord, V. and Faye, L. (1998) N−glycoprotein biosynthesis: recent development and future trends.Plant MoI. Biol, 38, 31−48
【非特許文献2】Bardor M, Faveeuw C, Fitchette A−C, Gilbert D, Galas L, Trottein F, Faye L and Lerouge P. (2003) Immureactivity in mammals of two typical plant glyco−epitopes, core−alpha(l,3)−fucose and core−xylose. Glycobiology, 13, 427−434
【非特許文献3】Ko, K., Tekoah, Y., Rudd, P.M., Harvey, D.J., Dwek, R.A., Spitsin, S., Hanlon, C.A., Rupprecht C, Dietzschold, B., Golovkin, M. and Koprowski, H. (2003) Function and glycosylation of plant−derived antiviral monoclonal antibody. Proc. Natl Acad. ScL USA, 101, 8013−8018
【非特許文献4】Sriraman, R., Bardor, M., Sack, M., Vaquero, C, Faye, L., Fischer, R., Finnern R. and Lerouge, P. (2004) Recombinant anti−hCG antibodies retained in the endoplasmic reticulum of transformed plants lack core xylose and core D(l,3)−fucose residues. Plant Biotech. J., 2, 279−287
【非特許文献5】Triguero, A., Cabrera, G., Cremata, J., Yuen, C−T., Wheeler J. and Ramirez N.I. (2005) Plant−derived mouse IgG monoclonal antibody fused to KDEL endoplasmic reticulum−retention signal is TV−glycosylated homogeneously throughout the plant with mostly high−mannose−type N−glycans. Plant Biotech. J. 3, 449−457
【非特許文献6】Palacpac, N. Q., Yoshida, S., Sakai, H., Kimura, Y., Fujiyama, K., Yoshida, T. and Seki, T. (1999) Stable expression of human beta 1,4−galactosyltransferase in plant cells modifies N−linked glycosylation patterns. Proc. Natl. Acad. Sci. USA 96, 4692− 4697
【非特許文献7】Bakker, H., Bardor, M., Molhoff, J., Gomord, V., Elbers, I., Stevens, L., Jordi, W., Lommen, A., Faye, L., Lerouge, P. and Bosch D. (2001) Humanized glycans on antibodies produced by transgenic plants. Proc. Natl. Acad. ScL USA, 98, 2899−2904
【非特許文献8】Kelm, S. And Schauer, R. (1997) Sialic acids in molecular and cellular interactions. Int. Rev. Cytol. 175, 137−240
【非特許文献9】Matsumoto, S., Ikura, K., Ueda, M. and Sasaki, R. (1995) Characterization of a human glycoprotein (erythropoietin) produced in cultured tobacco cells. Plant MoI. Biol.27, 1163−1172
【非特許文献10】Wee, E.Q., Sherrier, D.J., Prime, T.A. and Dupree, P. (1998) Targeting of active sialyltransferase to the plant Golgi apparatus. Plant Cell, 10, 1759−1768
【非特許文献11】Seveno, M., Bardor, M., Paccalet, T., Gomord, V., Lerouge, P. and Faye, L. (2004)Glycoprotein sialylation in plants? Nature Biotech., 22, 5−6
【非特許文献12】Angatta, T. and Varki, A. (2002) Chemical diversity in the sialic acids and related a−keto acids: an evolutionary perspertive. Chem Rev., 102, 439−469
【非特許文献13】Maru, I., Ohnishi, J., Ohta, Y. and Tsukada, Y. (1998) Simple and large−scale production of N−acetylneuraminic acid from N−acetyl−D−glucosamine and pyruvate using N− acyl−D−glucosamine epimerase and N−acetylneuraminate layse. Carbohydr. Res.,306, 575−578
【非特許文献14】Tanner, M. E. (2005) The enzymes of sialic acid biosynthesis. Bioorg. Chem., 33, 216−228
【発明の開示】
【課題を解決するための手段】
【0007】
(発明の要旨)
本発明は、植物体におけるシアル酸の合成に関する。さらに、本発明は、シアル酸を産生する方法及び植物体、並びにこれら植物体から産生されるシアル化タンパク質に関する。
【0008】
本発明の目的は、植物体においてシアル酸を産生する改良された方法を提供することである。
【0009】
本発明によれば、
i)N−アセチルノイラミン酸(Neu5Ac)シンターゼ又はNeu5Acリアーゼをコードするヌクレオチド配列である、植物体で活性のある調節領域と作動可能に連結されたヌクレオチド配列を含む植物体を提供することと、
ii)植物体を生育させ、ヌクレオチド配列を発現させ、それによってシアル酸を合成することを含む、シアル酸、たとえば、Neu5Acの合成方法が提供される。
【0010】
さらに、生育させる工程の後、シアル酸を植物体から回収してもよい。調節領域は、構成的プロモーター、誘導性プロモーター、組織特異的プロモーター及び発育プロモーターからなる群より選択されてもよい。
【0011】
本発明はまた、上記で定義された方法(方法A)に関するものであり、その際、提供する工程では、植物体はさらに、植物体内で活性がある1又は1を超えた第2の調節領域に作動可能に連結された、エピメラーゼ、CMP−Neu5Acシンターゼ、CMP−Neu5Acトランスポーター、ガラクトシルトランスフェラーゼ及びシアリルトランスフェラーゼの1又は1を超えるものをコードする第2のヌクレオチド配列を含む。第2のヌクレオチド配列は、目的のヌクレオチド配列の発現と共に共発現される。さらに第2の調節領域は、構成的プロモーター、誘導性プロモーター、組織特異的プロモーター及び発育プロモーターからなる群より選択されてもよい。
【0012】
本発明はまた、上述の方法(方法A)に関するものであり、その際、Neu5Acシンターゼ若しくはNeu5Acリアーゼをコードするヌクレオチド配列、エピメラーゼ、CMP−Neu5Acシンターゼ若しくはCMP−Neu5Acトランスポーターの1若しくは1を超えるものをコードする第2のヌクレオチド配列、又は目的のヌクレオチド配列及び第2のヌクレオチド配列の両方は、植物体内での発現のためにコドンが最適化されている。
【0013】
本発明は、
i)Neu5Acシンターゼ、Neu5Acリアーゼ、エピメラーゼ、CMP−Neu5Acシンターゼ、CMP−Neu5Acトランスポーター、ガラクトシルトランスフェラーゼ及びシアリルトランスフェラーゼをコードする1又は1を超える第1のヌクレオチド配列、並びに目的のタンパク質をコードする第2のヌクレオチド配列を発現する植物体を提供することと、
ii)植物体を生育させ、第1及び第2のヌクレオチド配列を発現させ、それによって目的のタンパク質を産生し、その際、目的のタンパク質がシアル化されることを含む、
目的のタンパク質を産生する方法(B)を提供する。
【0014】
好ましくは、シアル化される目的のタンパク質は、ジ−、トリ−又はテトラ−分岐N−グリカンを含む。
【0015】
本発明はまた、上述の方法(方法B)に関するものであり、その際、シアル化タンパク質は植物体から抽出される。さらに、シアル化タンパク質は単離され、精製されてもよい。
【0016】
本発明は、植物体で活性のある調節領域に作動可能に連結されたNeu5Acシンターゼ又はNeu5Acリアーゼをコードするヌクレオチド配列を含む植物体、植物細胞又は種子を提供する。植物体、植物細胞又は種子は、植物内で活性のある1又は1を超える第2の調節領域に作動可能に連結された、エピメラーゼ、CMP−Neu5Acシンターゼ、CMP−Neu5Acトランスポーター、ガラクトシルトランスフェラーゼ及びシアリルトランスフェラーゼの1又は1を超えるものをコードする第2のヌクレオチド配列をさらに含んでもよい。さらに、調節領域及び第2の調節領域は、構成的プロモーター、誘導性プロモーター、組織特異的プロモーター及び発育プロモーターからなる群より選択されてもよい。
【0017】
本発明はまた、上述の方法(方法B)に関するものであり、その際、Neu5Acシンターゼ、Neu5Acリアーゼ、エピメラーゼ、CMP−Neu5Acシンターゼ、CMP−Neu5Acトランスポーター、ガラクトシルトランスフェラーゼ及びシアリルトランスフェラーゼをコードする1又は1を超える第1のヌクレオチド配列、目的のタンパク質をコードする第2のヌクレオチド配列、又は第1の配列及び第2の配列の両方は、植物体、植物細胞内又は種子内で発現させるためにコドンが最適化されている。
【0018】
本発明は、植物体で活性のある調節領域と作動可能に連結されたNeu5Acシンターゼ又はNeu5Acリアーゼをコードするヌクレオチド配列をコードするヌクレオチド配列を含む植物体、植物細胞又は種子を包含する。植物体、植物細胞又は種子はさらに、植物体内で活性がある1又は1を超えた第2の調節領域に作動可能に連結された、エピメラーゼ、CMP−Neu5Acシンターゼ、CMP−Neu5Acトランスポーター、ガラクトシルトランスフェラーゼ及びシアリルトランスフェラーゼの1又は1を超えるものをコードする第2のヌクレオチド配列を含んでもよい。さらに、Neu5Acシンターゼ若しくはNeu5Acリアーゼをコードするヌクレオチド配列、エピメラーゼ、CMP−Neu5Acシンターゼ、CMP−Neu5Acトランスポーター、ガラクトシルトランスフェラーゼ及びシアリルトランスフェラーゼの1又は1を超えるものをコードする第2のヌクレオチド配列、又は目的の配列及び第2の配列の両方は、植物体、植物細胞内又は種子内で発現させるためにコドンが最適化されている。
【0019】
本発明はまた、Neu5Acシンターゼ、Neu5Acリアーゼ、エピメラーゼ、CMP−Neu5Acシンターゼ、CMP−Neu5Acトランスポーター、ガラクトシルトランスフェラーゼ及びシアリルトランスフェラーゼをコードする1又は1を超える第1のヌクレオチド配列、目的のタンパク質をコードする第2のヌクレオチド配列を含む植物体、植物細胞又は種子を提供するものであり、第1及び第2のヌクレオチド配列は、植物体で活性がある1又は1を超える調節領域に作動可能に連結される。1又は1を超える調節領域は、構成的プロモーター、誘導性プロモーター、組織特異的プロモーター及び発育プロモーターからなる群より選択されてもよい。さらに、Neu5Acシンターゼ、Neu5Acリアーゼ、エピメラーゼ、CMP−Neu5Acシンターゼ、CMP−Neu5Acトランスポーター、ガラクトシルトランスフェラーゼ及びシアリルトランスフェラーゼをコードする1又は1を超える第1のヌクレオチド配列、目的のタンパク質をコードする第2のヌクレオチド配列、又は第1の配列及び第2の配列の両方は、植物体、植物細胞内又は種子内で発現させるためにコドンが最適化されている。
【0020】
本発明はまた、N−アセチルノイラミン酸(Neu5Ac)シンターゼ又はNeu5Acリアーゼをコードするヌクレオチド配列である、植物体で活性がある調節領域と作動可能に連結されたヌクレオチド配列によって植物体又は植物体の一部を一過性に形質転換することと、ヌクレオチド配列を発現させ、それによってシアル酸を合成することを含むシアル酸の合成方法(方法C)も提供する。さらに、Neu5Ac又はNeu5Acリアーゼは、植物体又は植物体の一部から回収されてもよい。
【0021】
本発明はまた、上述の方法(方法C)に関するものであり、その際、植物体又は植物体の一部を一過性に形質転換する工程はさらに、植物体内で活性がある1又は1を超える第2の調節領域に作動可能に連結された、エピメラーゼ、CMP−Neu5Acシンターゼ、CMP−Neu5Acトランスポーターの1又は1を超えるものをコードする第2のヌクレオチド配列を含み、第2のヌクレオチド配列は、目的のヌクレオチド配列の発現と共に共発現される。
【0022】
本発明は、
i)Neu5Acシンターゼ、Neu5Acリアーゼ、エピメラーゼ、CMP−Neu5Acシンターゼ、CMP−Neu5Acトランスポーター、ガラクトシルトランスフェラーゼ及びシアリルトランスフェラーゼをコードする1又は1を超える第1のヌクレオチド配列、及び目的のタンパク質をコードする第2のヌクレオチド配列を発現する構築物によって植物体又は植物体の一部を一過性に形質転換することと、
ii)目的のタンパク質を産生し、その際目的のタンパク質がシアル化されることを含む
目的のタンパク質を産生する方法(方法D)を提供する。
【0023】
シアル化される目的のタンパク質は、ジ−、トリ−又はテトラ−分岐N−グリカンを含んでもよい。さらに、シアル化タンパク質は植物体又は植物体の一部から抽出されてもよい。シアル化目的のタンパク質はまた単離され、精製されてもよい。
【0024】
本発明はまた、上述の方法(方法D)に関するものであり、その際、産生する工程の後、シアル化目的のタンパク質を含む植物物質は、被験体に経口投与される。たとえば、産生する工程の後、植物体又は植物体の一部を最小限に加工して最小限に加工された植物物質を作製してもよく、シアル化された目的のタンパク質を含む最小限に加工された植物物質を被験体に経口投与してもよい。
【0025】
本明細書で記載されるように、植物体におけるNeu5Ac合成酵素、Neu5Acリアーゼ及びNeuB2の発現は結果的に植物組織内に機能的酵素の蓄積を生じる。Neu5Ac合成酵素は、任意の植物体、たとえば、タバコ及びMedicago sativa(アルファルファ)、分子農法に関する幾つかの農業的利点から恩恵を受ける多年生のマメ科作物で発現されてもよいが、これらに限定されない(Busse, et al., 2001)。
【0026】
本発明の要約は、本発明の特徴を必ずしもすべて記載するわけではない。
【0027】
本発明のこれらの特徴及びそのほかの特徴は、添付の図面を参照する説明からさらに明らかになるであろう。
【発明を実施するための最良の形態】
【0028】
(詳細な説明)
本発明は、植物体におけるシアル酸の合成に関する。さらに、本発明は、シアル酸を発現する方法及び植物体、並びにこれら植物体から産生されるシアル化タンパク質を提供する。
【0029】
以下の記載が、好ましい実施態様である。
【0030】
本発明は、植物体内でN−アセチルノイラミン酸(Neu5Ac)を合成する方法を提供する。Neu5Acリアーゼは、可逆的反応においてNeu5AcをManNAcとピルビン酸塩に切断するのを触媒することによって細菌にてシアル酸を異化する。この反応は可逆的なので、適切な前駆体の存在下でNeu5Acを合成するのにNeu5Acリアーゼを使用してもよい。Neu5Acの産生に関する代替方法には、Neu5Acシンターゼの使用が関与する。Neu5Acシンターゼは、D−ManNAcとPEPとの縮合によってNeu5Acを形成するのを触媒する。
【0031】
従って、本発明は、Neu5Acシンターゼ又はNeu5Acリアーゼをコードするヌクレオチド配列であって、植物体で活性のある調節領域と作動可能に連結されたヌクレオチド配列を含む植物体を提供することと、植物体を生育させ、ヌクレオチド配列を発現させてNeu5Acを合成することを含むNeu5Acを合成する方法を提供する。或いは、方法には、植物体又は植物体の一部の内部におけるNeu5Acの一過性の産生が関与してもよい。
【0032】
そのように産生されるNeu5Acは、植物体から回収され、当該技術で既知の方法を用いたin vitroでのタンパク質のシアル化に使用されてもよい。或いは、Neu5Acは、植物体内で共発現される目的のタンパク質のシアル化のための内因性基質として使用されてもよい。
【0033】
所望であれば、植物体内でNeu5Acを合成するための、N−アセチルマンノサミン(D−ManNAc)を含むが、これに限定されない基質のレベルは、エピメラーゼ、CMP−Neu5Acシンターゼ、CMP−Neu5Acトランスポーターの1又は1を超えるものをコードする1又は1を超える追加のヌクレオチド配列を植物体内で共発現させることによって高められてもよい。たとえば、植物体内で、UDP−GlcNAc2−エピメラーゼ、たとえば、細菌のUDP−GlcNAc2−エピメラーゼ、又は内因性のUDP−GlcNAc2をManNAcに変換するそのほかの供給源からのエピメラーゼを発現させることによってManNAcを合成してもよい。或いは、ManNAc−6−リン酸を製造し、その後、ホスファターゼによって加水分解してもよい。このアプローチによって、GlcNAc−6リン酸2−エピメラーゼ、たとえば、細菌のGlcNAc−6リン酸2−エピメラーゼ、又は哺乳類のUDP−GlcNAc2−エピメラーゼ/ManNAcキナーゼが植物体内で発現される。Neu5Acシンターゼ又はNeu5Acリアーゼをコードするヌクレオチド配列の発現と共にこの第2のヌクレオチド配列を共発現させることによって、高いレベルのNeu5Acが産生される。しかしながら、1以上のこれら酵素の内因性の活性が植物体の内部に存在してもよいので、1以上の上記ヌクレオチド配列の共発現の必要性は、選択される宿主に依存する。
【0034】
細胞質のシアル酸からのN−グリカンのシアル化を確実にするには、細菌又は哺乳類のCMP−Nue5Acシンターゼ、哺乳類のCMP−Nue5Acトランスポーター、哺乳類のガラクトシルトランスフェラーゼ(シアル酸をN−グリカンに変換する前のガラクトースの添加にために)及び哺乳類のシアリルトランスフェラーゼを植物体内で共発現させてもよい。本発明に従って植物体内で産生されたNeu5Acは、CMP−Nue5Acシンターゼを介したCMP−N−アセチルノイラミン酸(CMP−Nue5Ac)の合成のための基質として使用されてもよく、CMP−Nue5Acは次いで、植物体内で共発現される目的のタンパク質のシアル化のための基質として使用される。この場合、植物体は、シアリルトランスフェラーゼをコードするヌクレオチド配列を含んでもよい。哺乳類のシアリルトランスフェラーゼ及び哺乳類のCMP−Nue5Acシンターゼの植物体における発現は実証されている(参照によって本明細書に組み入れられるWee et al., 1998, Misaki R,. et al., 2006)。しかしながら、1以上のこれら酵素の内因性の活性が植物体内で存在してもよいので、1以上の上記ヌクレオチド配列を共発現する必要性は、選択される宿主植物体に依存してもよい。
【0035】
ヌクレオチド配列が植物体内で共発現する場合、標準の形質転換手法である、一過性の形質転換手法を用いて所望のヌクレオチド配列のそれぞれを植物体に導入してもよいし、又は1以上の所望のヌクレオチド配列をそれぞれ発現する2種の植物体を交配してヌクレオチド配列の必要とされる組み合わせを共発現する植物体を得てもよい。
【0036】
従って、本発明はまた、シアル化された目的のタンパク質の産生のための基盤として使用されてもよい植物体を作製する方法も提供する。本方法は、Neu5Acシンターゼ、Neu5Acリアーゼ、エピメラーゼ、CMP−Neu5Acシンターゼ、CMP−Neu5Acトランスポーター、ガラクトシルトランスフェラーゼ及びシアリルトランスフェラーゼをコードする1又は1を超える第1のヌクレオチド配列を発現する植物体を提供することと、1以上のヌクレオチド配列を発現することを含む。目的のタンパク質を産生するために、標準技法、たとえば、形質転換法を用いて、目的のタンパク質をコードする第2のヌクレオチド配列を基盤植物体に導入し、第2のヌクレオチド配列を発現させるか、又は交配された植物体の子孫の中で産生された目的のタンパク質がシアル化されるように目的のタンパク質を発現している植物体と基盤植物体を交配させる。
【0037】
本発明はまた、Neu5Acシンターゼ、Neu5Acリアーゼ、エピメラーゼ、CMP−Neu5Acシンターゼ、CMP−Neu5Acトランスポーター、ガラクトシルトランスフェラーゼ及びシアリルトランスフェラーゼをコードする1又は1を超える第1のヌクレオチド配列並びに目的のタンパク質をコードする第2のヌクレオチド配列を発現する植物体を提供することと、植物体を生育させ、第1及び第2のヌクレオチド配列を発現させ、それによって目的のタンパク質を産生し、その際目的のタンパク質がシアル化されることを含む、目的のタンパク質を産生する方法にも関する。好ましくは、シアル化される目的のタンパク質は、ジ−、トリ−又はテトラ−分岐N−グリカンを含む。シアル化タンパク質は植物体から抽出してもよく、所望であれば、シアル化タンパク質は、常法を用いて単離及び精製してもよい。再び、所望の構築物によって植物体を安定的に形質転換してもよいし、又は、所望の構築物によって植物体又は植物体の一部を一過性に形質転換してもよい。
【0038】
Neu5Acシンターゼ、Neu5Acリアーゼ、エピメラーゼ、CMP−Neu5Acシンターゼ、CMP−Neu5Acトランスポーター、ガラクトシルトランスフェラーゼ及びシアリルトランスフェラーゼをコードするヌクレオチド配列は、コドンを最適化して植物体内での発現レベルを高めてもよい。コドンの最適化によって、植物体内でのコドン使用法にアプローチするために、構造遺伝子又はその断片のオリゴヌクレオチド基本単位の合成及びそれに続く酵素の集合のための適切なDNAヌクレオチドの選択を意味する。
【0039】
植物体内での外来配列の発現を最適化するために、野生型であっても合成配列であってもよいヌクレオチド配列が使用されてもよく、又は、相当するタンパク質、たとえば、Neu5Acシンターゼ、Neu5Acリアーゼ、エピメラーゼ、CMP−Neu5Acシンターゼ、CMP−Neu5Acトランスポーター、ガラクトシルトランスフェラーゼ及びシアリルトランスフェラーゼ、目的のタンパク質、若しくはそれらの組み合わせが、非修飾ヌクレオチド配列によってコードされる場合に産生されるよりも高いレベルで産生される。たとえば、限定について考慮しないが、配列は、合成配列であってもよく、たとえば、BLAST(GenBankを介して利用可能;初期設定パラメーターを用いて)のような、しかし、これに限定されない配列比較法を用いて決定されるように、野生型配列と少なくとも80%の相同性を含んで、植物体内でのコドン利用について配列が最適化されてもよい。たとえば、抗原特性のような、しかし、これに限定されない有用な生物特性を示す目的のタンパク質又はその誘導体をコードする配列の断片又は一部が植物組織の中で発現されてもよいことも企図される。
【0040】
Neu5Acシンターゼ、Neu5Acリアーゼ、エピメラーゼ、CMP−Neu5Acシンターゼ、CMP−Neu5Acトランスポーター、ガラクトシルトランスフェラーゼ、シアリルトランスフェラーゼ及び目的のタンパク質の発現レベル及び導入遺伝子のタンパク質産生をできるだけ多くするために、核酸配列を調べて、Sardana at al.(Plant Cell Reports, 15:677−681, 1996)が概説したものに類似する手順を用いてコーディング領域を修飾し、植物体における遺伝子の発現のために最適化してもよい。双子葉植物において高度に発現される遺伝子のコドン利用の表は、Murray et al (Nuc. Acids Res., 17:477−498, 1989)を含めて幾つかの供給元から利用可能である。
【0041】
従って、本発明は、N−アセチルノイラミン酸(Neu5Ac)シンターゼ又はNeu5Acリアーゼをコードするヌクレオチド配列であって、植物体で活性のある調節領域と作動可能に連結されたヌクレオチド配列を含む植物体を提供することと、植物体を生育させ、ヌクレオチド配列を発現させ、それによってシアル酸を合成することを含むシアル酸を合成する方法を提供する。Neu5Acシンターゼ又はNeu5Acリアーゼをコードするヌクレオチド配列は、植物体内での発現のためにコドンが最適化されてもよい。さらに、提供する工程では、植物体はさらに、植物体内で活性がある1又は1を超えた第2の調節領域に作動可能に連結された、エピメラーゼ、CMP−Neu5Acシンターゼ又はCMP−Neu5Acトランスポーターの1又は1を超えるものをコードする第2のヌクレオチド配列を含み、第2のヌクレオチド配列は、目的のヌクレオチド配列の発現と共に共発現される。エピメラーゼ、CMP−Neu5Acシンターゼ又はCMP−Neu5Acトランスポーターの1又は1を超えるものをコードする第2のヌクレオチド配列は、植物体内で発現されるためにコドンが最適化されてもよい。
【0042】
さらに、本発明は、Neu5Acシンターゼ、Neu5Acリアーゼ、エピメラーゼ、CMP−Neu5Acシンターゼ、CMP−Neu5Acトランスポーター、ガラクトシルトランスフェラーゼ、シアリルトランスフェラーゼをコードする1又は1を超える第1のヌクレオチド配列、及び目的のタンパク質をコードする第2のヌクレオチド配列を発現する植物体を提供することと、植物体を生育させ、第1及び第2のヌクレオチド配列を発現させ、それによって目的のタンパク質を産生する目的のタンパク質を産生する方法を提供する。Neu5Acシンターゼ、Neu5Acリアーゼ、エピメラーゼ、CMP−Neu5Acシンターゼ、CMP−Neu5Acトランスポーター、ガラクトシルトランスフェラーゼ、シアリルトランスフェラーゼをコードする1又は1を超える第1のヌクレオチド配列、目的のタンパク質をコードする第2のヌクレオチド配列、又は第1ヌクレオチド配列及び第2のヌクレオチド配列の両方は、植物体内で発現されるためにコドンが最適化されてもよい。
【0043】
さらに、本発明は、植物体で活性のある調節領域に作動可能に連結されたNeu5Acシンターゼ又はNeu5Acリアーゼをコードするヌクレオチド配列を含む植物体、植物細胞又は種子に関する。植物体、植物細胞又は種子は、植物内で活性のある1又は1を超える第2の調節領域に作動可能に連結された、エピメラーゼ、CMP−Neu5Acシンターゼ、CMP−Neu5Acトランスポーター、ガラクトシルトランスフェラーゼ及びシアリルトランスフェラーゼの1又は1を超えるものをコードする第2のヌクレオチド配列をさらに含んでもよい。Neu5Acシンターゼ又はNeu5Acリアーゼをコードするヌクレオチド配列、エピメラーゼ、CMP−Neu5Acシンターゼ、CMP−Neu5Acトランスポーターの1又は1を超えるものをコードする第2のヌクレオチド配列、又は該ヌクレオチド配列及び第2のヌクレオチド配列の両方は、植物体、植物細胞又は種子内で発現されるためにコドンが最適化されてもよい。
【0044】
本発明はまた、Neu5Acシンターゼ、Neu5Acリアーゼ、エピメラーゼ、CMP−Neu5Acシンターゼ、CMP−Neu5Acトランスポーター、ガラクトシルトランスフェラーゼ及びシアリルトランスフェラーゼをコードする1又は1を超える第1のヌクレオチド配列、目的のタンパク質をコードする第2のヌクレオチド配列を含む植物体、植物細胞又は種子を含み、第1及び第2のヌクレオチド配列は、植物体で活性がある1又は1を超える調節領域に作動可能に連結される。Neu5Acシンターゼ、Neu5Acリアーゼ、エピメラーゼ、CMP−Neu5Acシンターゼ、CMP−Neu5Acトランスポーター、ガラクトシルトランスフェラーゼ及びシアリルトランスフェラーゼをコードする1又は1を超える第1のヌクレオチド配列、目的のタンパク質をコードする第2のヌクレオチド配列、又は第1のヌクレオチド配列及び第2のヌクレオチド配列の両方は、植物体内で発現されるためにコドンが最適化されてもよい。
【0045】
「作動可能に連結する」によって、特定の配列が直接又は間接的に相互作用し、たとえば、遺伝子発現の介在又は修飾のような意図される機能を実行することを意味する。作動可能に連結された配列の相互作用は、たとえば、作動可能に連結された配列と相互作用するタンパク質が介在してもよい。転写調節領域が介在する又はそれによって修飾されるべき目的の配列の転写を行えるように配列が機能的に接続される場合、転写調節領域と目的の配列は作動可能に連結される。
【0046】
用語「植物物体」によって、植物体に由来する任意の物質を意味する。植物物体は、植物体全体、その組織、細胞又は任意の分画を含んでもよい。さらに、植物物体は、植物体の細胞内植物成分、細胞外植物成分、液体、又は固形抽出物、又はそれらの組み合わせを含んでもよい。さらに、植物物体は、植物体の葉、茎、果実、根、又はそれらの組み合わせからの植物体、植物細胞、組織、液体抽出物又はそれらの組み合わせを含んでもよい。植物物体は、加工工程の対象とされていない植物体又はその一部を含んでもよい。しかしながら、植物物体は、以下で定義するような最小限の加工工程、又はクロマトグラフィ、電気泳動などを含むが、これらに限定されない当該技術で共通して知られる技法を用いた部分的な又は実質的なタンパク質精製を含む綿密な工程の対象とされてもよいことも企図される。
【0047】
用語「最小限の加工」によって、植物抽出物、ホモジネート、ホモジネートの分画などを得るために部分的に精製される目的のタンパク質を含む植物物体、たとえば、植物体又はその一部を意味する。部分精製は、植物体の細胞構造を破壊し、それによって可溶性の植物成分、及びたとえば、それらに限定されないが、遠心分離、濾過又はそれらの組み合わせによって分離されてもよい不溶性の植物成分を含む組成物を創製することを含んでもよいが、これらに限定されない。この点で、葉又はそのほかの組織の細胞外空間に分泌されたタンパク質は、吸引又は遠心抽出を用いて容易に得ればよく、加圧下でローラーを通過させる又は粉砕するなどによって組織を抽出し、細胞外空間から離れたタンパク質を圧搾する又は遊離させればよい。最小限の加工には、可溶性タンパク質の粗抽出物の調製が関与すればよく、これらの調製物は、二次的な植物生成物からの混入が無視できるほどだからである。さらに、最小限の加工には、可溶性タンパク質の葉からの水性抽出、その後の好適な塩による沈殿が関与してもよい。そのほかの方法には、抽出物の直接的な使用を可能にするための大規模浸漬及び分泌液抽出が挙げられる。
【0048】
植物物質又は組織の形態での植物物体は、経口にて被験体に送達されてもよい。植物物体は、栄養補助食品の一部として他の食物とともに投与されてもよく、カプセルに内包されてもよい。植物物体又は組織は、濃縮して美味しさを改善する又は高めてもよく、必要に応じてそのほかの物質、成分又は医薬賦形剤とともに提供されてもよい。
【0049】
不均質な目的のタンパク質を含む植物体を、必要性と状況に応じて様々な方法で、被験体、たとえば、動物又はヒトに投与してもよいことが企図される。たとえば、タンパク質が経口で投与されるのであれば、植物体を収穫し、直接被験体に与えてもよいし、与える前に収穫した組織を乾燥してもよいし、収穫を行わずに動物が植物体を食べられるようにしてもよい。収穫された植物体組織に関する本発明の範囲内で、動物の餌の中に食品補助として提供されることも考慮される。植物体組織がほとんど又は全くさらに加工することなく動物に与えられるのであれば、投与される植物体組織は食用であることが好ましい。さらに、植物体から得られる目的のタンパク質は、食品補助として使用する前に、加工しない形態、部分精製した形態、又は生成した形態に抽出されてもよい。この後者の場合、タンパク質は食用又は非食用の植物体のいずれかから産生されればよい。
【0050】
さらに詳細には実施例で記載されるように、Neu5Acリアーゼ及びNeu5Acリアーゼ−FLAG(形質転換体で組換えタンパク質を免疫的に検出するためにC末端にてFLAGエピトープをタグとして付けたNeu5Acリアーゼ)を植物体に導入した。抗FLAG抗体を用いたウエスタンブロット分析によって、形質転換された細胞にMWr32kDaのタンパク質が存在することが明らかにされた(図1a)。Neu5Acリアーゼ又はNeu5Acリアーゼ−FLAGのいずれかを発現している植物体から得られた抽出物にて、in vitro及びin vivoの両方でリアーゼ活性が検出可能だった。非形質転換植物体にて内因性のリアーゼ活性は検出されなかった。しかしながら、D−ManNAcとピルビン酸塩の存在下にて組換え産生されたNeu5Acリアーゼを用いたNeu5Acの合成は認められた(図2bを参照のこと)。従って、組換え的に発現されたNeu5Acリアーゼは、植物体にて生物学的に活性がある。
【0051】
Neu5Acシンターゼ(たとえば、NeuB2、しかし、これに限定されない)及びNeu5Acシンターゼ−FLAG(形質転換体で組換えタンパク質を免疫的に検出するためにC末端にてFLAGエピトープをタグとして付けたNeu5Acシンターゼ)を植物に導入した。抗FLAG抗体を用いたウエスタンブロット分析によって、形質転換された細胞にMWr37kDaのタンパク質が存在することが明らかにされた。Neu5Acシンターゼ又はNeu5Acシンターゼ−FLAGのいずれかを発現している植物体から得られた抽出物にて、in vitro及びin vivoの両方でシンターゼ活性が検出可能だった。非形質転換植物体にて内因性のシンターゼ活性は検出されなかった。しかしながら、D−ManNAcとPEPの存在下にて組換え産生されたNeu5Acシンターゼを用いたNeu5Acの合成は認められた(図3b、cを参照のこと)。従って、組換え的に発現されたNeu5Acシンターゼは、植物体にて生物学的に活性がある。
【0052】
「類縁体」又は「誘導体」は、ヌクレオチド配列が、標的遺伝子又は配列の発現を停止する、標的配列の発現を低下させる、又は標的配列にコードされるタンパク質の合成又は活性を低下させる特性を保持するという条件で、サイレンシングヌクレオチド配列への置換、欠失又は付加を含む。たとえば、核酸配列の誘導体及び類縁体は通常、サイレンシング核酸配列に80%を超える類似性を示す。配列の類似性は、初期設定プログラム(プログラム:blastn、データベース:nr、Expect10、フィルター:低複雑性、アライメント:ペアで、ワールドサイズ:II)を用いたBLASTアルゴリズム(GenBank、ncbi.nlm.nih.gov/cgi−bin/BLAST)の使用によって決定されてもよい。その類縁体又は誘導体はまた、厳密なハイブリッド形成の条件下で(Maniatis et al., in Molecular Cloning (A Laboratory Manual), Cold Spring Harbor Laboratory, 1982, p. 387−389, 又はAusubel at al., (eds), 1989, Current Protocol in Molecular Biology, Vol. 1, Green Publishing Associates, Inc., and John Wiley & Sons, Inc., New Yolk p.2.10.3を参照のこと)配列が標的遺伝子の発現を停止する特性を示すという条件で本明細書に記載される配列の1つにハイブリッド形成するヌクレオチド配列も含む。そのような厳密なハイブリッド形成条件の例の1つは、7%SDS、1mMのEDTA、0.5MのNa2HPO4、pH7.2中で65ECにて16〜20時間、たとえば、[γ−32P]dATP標識されたプローブのような、しかし、これに限定されない好適なプローブとのハイブリッド形成であってもよい。続いて、5%SDS、1mMのEDTA、40mMのNa2HPO4、pH7.2で30分間洗浄し、続いて1%SDS、1mMのEDTA、40mMのNa2HPO4、pH7.2で30分間洗浄すること。この緩衝液での洗浄を繰り返してバックグランドを低減させてもよい。
【0053】
「調節領域」、「調節要素」または「プロモーター」によって、常とは限らないが、通常、遺伝子のタンパク質コーディング領域の上流にある核酸の部分を意味し、DNA又はRNAのいずれか、又はDNA及びRNAの両方で構成されてもよい。調節領域が目的の遺伝子によって活性を持つ場合、目的の遺伝子と作動関係にある場合、目的の遺伝子と作動可能に連結される場合、これが、結果として目的の遺伝子の発現を生じてもよい。調節要素は、臓器特異性又は発生上の若しくは一時的な遺伝子の活性化の制御に介在することが可能である。「調節領域」には、プロモーター要素、基本のプロモーター活性を示すコアプロモーター要素、外部刺激に反応して誘導可能である要素、たとえば、負の調節要素又は転写エンハンサーのようなプロモーター活性に介在する要素が含まれる。本明細書で使用されるとき、「調節領域」には、転写に続いて活性を持つ要素、たとえば、翻訳及び転写のエンハンサー、翻訳及び転写のリプレッサー、上流活性化配列並びにmRNA不安定性決定基のような遺伝子発現を調節する調節要素が挙げられる。これら後者の要素の幾つかは、コーディング領域の近傍に位置してもよい。
【0054】
この開示の背景で、用語「調節要素」又は「調節領域」は通常、常とは限らないが、普通、構造遺伝子の上流(5’)のDNAの配列を言い、RNAポリメラーゼ及び/又は特定の部位で開始する転写に必要とされるそのほかの因子に対して認識を提供することによってコーディング領域の発現を制御する。しかしながら、配列のイントロン又は3’の中に位置するそのほかのヌクレオチド配列も目的のコーディング領域の発現の調節に寄与してもよいことが理解されるべきである。RNAポリメラーゼ又は特定の部位からの開始を確実にするそのほかの転写因子に対して認識を提供する調節因子の例は、プロモーター要素である。すべてとは限らないが、ほとんどの真核生物のプロモーター要素は、普通、転写開始部位のおよそ25塩基対上流に位置するアデノシンとチミジンのヌクレオチド塩基対から構成される保存された核酸配列であるTATAボックスを含有する。プロモーター要素は、転写の開始に反応性である基本のプロモーター要素、同様に、遺伝子発現を加減するほかの調節要素(上記で列記したような)を含む。
【0055】
発育上調節されるもの、誘導可能なもの又は構成的なものを含めて幾つかの種類の調節領域が存在する。発育上調節される、又はその制御下で遺伝子の差次的発現を制御する調節領域は、特定の臓器又は組織の発育中、特定の時間でその臓器又は組織の中で活性化される。しかしながら、一部の発育上調節される調節領域は、特定の発育段階で特定の臓器又は組織で優先的に活性化されてもよく、それらは、発育調節的に活性を持ってもよいし、又は植物体内で同様に他の臓器又は組織における基本的なレベルで活性を持ってもよい。組織特異的な調節領域の例は、たとえば、具体的に見た調節領域には、ナピンプロモーター及びクルシフェリンプロモーターが挙げられる(Rask et al., 1998, J. Plant Physiol., 152:595−599; Bilodeau et al., 1994, Plant Cell 14:125−130)。
【0056】
誘導可能な調節領域は、誘導因子に反応して1以上のDNA配列又は遺伝子の転写を直接又は間接的に活性化することが可能であるものである。誘導因子の非存在下では、DNA配列又は遺伝子は転写されない。通常、誘導可能な調節領域に特異的に結合して転写を活性化するタンパク質因子は、不活性の形態で存在してもよく、次いで直接又は間接的に誘導因子によって活性形態に変換される。しかしながら、タンパク質因子は不在であってもよい。誘導因子は、たとえば、タンパク質、代謝産物、増殖調節剤、除草剤若しくはフェノール化合物のような化学物質、熱、寒冷、塩によって直接強いられる生理的ストレス、又は毒性要素又はウイルスのような病原体又は病原剤の作用を間接的に介したものであることができる。たとえば、噴霧する、水で薄める、加熱することによって又は類似の方法によって細胞又は植物体に誘導因子を外から適用することによって、誘導可能な調節領域を含有する植物細胞を誘導因子に暴露してもよい。誘導可能な調節領域は、植物遺伝子又は非植物遺伝子のいずれかに由来してもよい(たとえば、Garz C and Lenk I.R.P., Trends Plant Sci., 3:352−358, 1998)。強力な誘導性プロモーターの例には、テトラサイクリン誘導性プロモーター(参照によって組み入れられるCratz C., 1997, Ann. Rev. Plant Physiol. Plant Mol. Biol., 48:89−108)、ステロイド誘導性プロモーター(参照によって組み入れられるAoyama T. and Chua, N.H., 1997, Plant J., 2:397−404)、及びエタノール誘導性プロモーター(参照によって組み入れられるSalter M.G. et al., 1998, Plant Journal 16:127−132; Caddick M. X. et al., 1998, Nature Biotech., 16:177−180)、サイトキニン誘導性IB6及びCKII遺伝子(参照によって組み入れられるBrandstater I. and Kieber, J.J., 1998, Plant Cell 10: 1009−1019; Kakimoto T., 1996, Science, 274:982−985)及びオーキシン誘導性要素DR5(参照によって組み入れられるUlmasov, T. et al., 1997, Plant Cell, 9:1963−1971)が挙げられるが、これらに限定されない。
【0057】
構成的な調節領域は、植物体の種々の部分の全体にわたって、且つ植物体の生育全体にわたって継続して遺伝子の発現を指向する。既知の構成的な調節要素の例には、CaMV35S転写物(Odell et al., 1985, Nature, 313:810−812)、コメのアクチン1(Zhang at al., 1991, Plant Cell, 3:1155−1165)、アクチン2(An et al., 1996, Plant J., 10:107−121)、又はtms(参照によって本明細書に組み入れられる米国特許第5,428,147号)、及びトリオセホスフェートイソメラーゼ1(Xu et al., 1994, Plant Physiol., 106:459−467)遺伝子、トウモロコシのユビキチン1遺伝子(Cornejo et al., 1993, Plant Mol Biol., 29:637−646)、Arabidopsisのユビキチン1及び6の遺伝子(Holtorf et al., 1995, Plant Mol. Biol., 29:637−646)、及びタバコの翻訳開始因子4A遺伝子(Mandel et al., 1995, Plant Mol. Biol., 29:995−1004)に関連するプロモーターが挙げられる。用語「構成的な」は、本明細書で使用されるとき、構成的な調節領域の制御下にある遺伝子があらゆる種類の細胞にて同一レベルで発現されるが、該遺伝子は、多量の変異が認められることが多いにもかかわらず、広い範囲の細胞種で発現されることを必ずしも指し示さない。
【0058】
本発明の1又は1を超えるヌクレオチド配列は、本発明のヌクレオチド配列、又は構築物又はベクターで形質転換される好適な植物体宿主で発現されてもよい。好適な宿主の例には、アルファルファ、カノーラ、Brassica spp、トウモロコシ、タバコ、アルファルファ、ジャガイモ、朝鮮ニンジン、エンドウ豆、カラスムギ、コメ、大豆、小麦、大麦、ヒマワリ、及び綿を含む農作物が挙げられるが、これらに限定されない。
【0059】
従って、本発明は、植物体で活性のある調節領域と作動可能に連結された、Neu5Acシンターゼ又はNeu5Acリアーゼをコードするヌクレオチド配列を含む植物体、植物細胞又は種子も提供する。さらに植物体、植物細胞又は種子は、植物体内で活性がある1又は1を超えた第2の調節領域に作動可能に連結された、エピメラーゼ、CMP−Neu5Acシンターゼ、CMP−Neu5Acトランスポーター、ガラクトシルトランスフェラーゼ及びシアリルトランスフェラーゼの1又は1を超えるものをコードする第2のヌクレオチド配列を含んでもよい。
【0060】
本発明はまた、Neu5Acシンターゼ、Neu5Acリアーゼ、エピメラーゼ、CMP−Neu5Acシンターゼ、CMP−Neu5Acトランスポーター、ガラクトシルトランスフェラーゼ及びシアリルトランスフェラーゼをコードする1又は1を超える第1のヌクレオチド配列、目的のタンパク質をコードする第2のヌクレオチド配列を含む植物体、植物細胞又は種子を提供するものであり、第1及び第2のヌクレオチド配列は、植物体で活性がある1又は1を超える調節領域に作動可能に連結される。
【0061】
本発明の1以上のキメラ遺伝構築物は、さらに3’非翻訳領域を含むことができる。3’非翻訳領域は、ポリアデニル化シグナル及びmRNAのプロセッシング又は遺伝子発現を達成することが可能であるそのほかの調節シグナルを含有するDNA断片を含む遺伝子の部分を言う。ポリアデニル化シグナルは普通、mRNA前駆体の3’末端へのポリアデニル酸のトラックの付加を達成することを特徴とする。ポリアデニル化シグナルは、変異が珍しくはないが、標準形態5’−AATAAA−3’に対する相同性の存在によって一般に認識される。本発明の1以上のキメラ遺伝構築物は、必要に応じてさらに、エンハンサー、翻訳エンハンサー又は転写エンハンサーのいずれかを含むことができる。これらのエンハンサーは当業者に周知であり、ATG開始コドンと隣接する配列を含むことができる。開始コドンは、配列全体の翻訳を確実にするようにコーディング配列の読み取りフレームに同調しなければならない。
【0062】
好適な3’領域の非限定例は、たとえば、ノパリンシンターゼ(Nos遺伝子)のような(Ti)プラスミド遺伝子並びに大豆保存タンパク質遺伝子及びリブロース−1,5−ビスホスフェートカルボキシラーゼの小型サブユニット(ssRUBISCO)遺伝子のような植物遺伝子を誘導するアグロバクテリウム腫瘍のポリアデニル化シグナルを含有する3’転写非翻訳領域である。
【0063】
形質転換した植物細胞での同定に役立つように、本発明の構築物は、植物体で選抜可能なマーカーを含むようにさらに操作してもよい。有用な選抜可能なマーカーには、たとえば、ゲンタマイシン、ハイグロマイシン、カナマイシンのような抗生剤、又はホスホノトリシン、グリホサート、クロロスルフロンなどのような除草剤のような化学物質に耐性を提供する酵素が挙げられる。同様に、たとえば、GUS(β−グルクロニダーゼ)のような色変化によって同定可能な化合物の産生、又はたとえば、ルシフェラーゼ若しくはGFPのような発光を提供する酵素を使用してもよい。
【0064】
また、本発明の考慮される部分は、本発明のキメラ遺伝構築物を含有するトランスジェニック植物体、植物細胞又は種子である。植物細胞から植物体全体を再生させる方法も当該技術では既知である。一般に、形質転換された植物細胞は適切な培地で培養され、培地は、抗生剤のような選抜剤を含有してもよく、その際、選抜可能なマーカーを使用して形質転換した植物細胞の同定を円滑にする。いったん、カルスが形成されると、既知の方法に準じて適切な植物ホルモンを用いることによって芽形成を促し、芽を植物再生用の発根培地に移す。次いで、種子から、又は栄養繁殖技法を用いて、該植物体を使用して繰り返し世代を確立してもよい。組織培養を用いることなくトランスジェニック植物体を生成することもできる。
【0065】
形質転換又は一時的な発現の影響を受け易い宿主生物の範囲内での発現のために、本発明の調節要素を目的のコーディング領域と組み合わせてもよい。そのような生物には、単子葉植物及び双子葉植物両方の植物が挙げられるが、これらに限定されず、たとえば、トウモロコシ、シリアル植物、小麦、大麦、タバコ、Brassica、大豆、豆、エンドウ豆、アルファルファ、ジャガイモ、トマト、朝鮮ニンジン及びArabidopsisが挙げられるが、これらに限定されない。
【0066】
これら生物の一時的な発現、形質転換及び再生の方法は、当該技術で確立され、当業者に既知である。形質転換された植物体及び再生された植物体を入手する方法は本発明に決定的ではない。
【0067】
「形質転換」によって、遺伝子型として、表現型として、又はその両方で明らかにされる遺伝情報の種間の移転を意味する。キメラ遺伝構築物から宿主への遺伝情報の種間移転は、遺伝性であってもよく、遺伝情報の移転は安定であるとみなされてもよく、又は移転は、一時的であってもよく、遺伝情報の移転は遺伝性ではない。本発明はさらに、安定な発現系又は一時的な発現系のいずれかと共に使用するのに好適なキメラ遺伝子構築物を含む好適なベクターをさらに含む。
【0068】
Tiプラスミド、Riプラスミド、植物ウイルスベクターを用いて、直接的なDNA形質転換、微量注入、エレクトロポレーションによって本発明の構築物を植物細胞に導入することができる。そのような技法の概説については、たとえば、Weissbach and Weissbach, Methods for Plant Molecular Biology, Academic Press, New Yolk, VIII, pp 421−463, 1988; Geierson and Corey, Plant Molecular Biology, 2d, Ed(1988),;及びMIki and lyer, Fundamentals of Gene Transfer in Plants, in Plant Metabolism. 2dEd, DT, Dennis, DH Turpin, DD Lefebrve, DB, Layzell (eds), Addison Wesly, (Langmans社, London pp561−579(1997)を参照のこと。そのほかの方法には、直接的なDNAの取り込み、リポソームの使用、エレクトロポレーション、たとえば、プロトプラストの使用、微量注入、微量発射若しくはウイスカー、及び真空浸透が挙げられる。たとえば、Bilang et al., (Gene, 100:247−250, 1991), Scheid et al., (Mol. Gen. Genet.,228:104−112, 1991), guerche et al., (Plant Science 52:111−116, 1987), Neuhause et al., (Theor. Appl. genet., 75:30−36, 1987), Klein et al., Nature 327:70−73, 1987; Howell et al., (Science, 208:1265, 1980), Horsch et al., (Sceinec 227:1229−1231, 1985); DeBlock et al., Plant Physiology 91:694−701, 1989; Methods for Plant Molecular Biology (Weissbach and Weissback, eds, Academic Press Inc., 1988; Methods in Plant Molecular Biology (Scheler and Zielinski eds., Academic Press Inc., 1989); Liu and Lomonossoff (J. Virol. Meth., 105:343−348, 2002); 米国特許第4,945,050号、同第5,036,006号及び同第5,100,792号、1995年5月10日出願の米国特許出願出願番号第08/438,666号、及び1992年9月25日出願の同第07/951,715号(すべて参照によって本明細書に組み入れられる)を参照のこと。
【0069】
以下に記載されるように、一過性の発現方法を用いて本発明の構築物を発現させてもよい(参照によって本明細書に組み入れられるLiu Lomonossodd, 2002, Journal of Virological Methods 105:343−348を参照のこと)。これらの方法には、たとえば、アグロ−植菌又はアグロ−浸潤が挙げられるが、これらに限定されない。しかしながら、上で述べたように他の一過性の方法も使用してもよい。アグロ−植菌又はアグロ−浸潤のいずれかによって、所望の核酸を含むアグロバクテリウムの混合物は、組織、たとえば、葉の細胞間間隙、植物体(茎、葉及び花を含む)の空間、植物体(茎、根、花)のそのほかの部分、又は植物体全体に入る。上皮を越境させた後、アグロバクテリウムが感染し、細胞内にDNAのコピーを転移させる。t−DNAは、エピソームで転写され、mRNAが翻訳され、感染した細胞における目的のタンパク質の産生をもたらす。しかしながら、t−DNAの核への継代は一過性である。
【0070】
「目的の遺伝子」、「目的のヌクレオチド配列」又は「目的のコーディング領域」によって、宿主生物、たとえば、植物体の中で発現されるべき遺伝子、ヌクレオチド配列、又はコーディング領域を意味する。これらの用語は相互交換可能に使用される。そのような目的のヌクレオチド配列には、その産物が工業用の酵素、タンパク質、サプリメント、餌、食品又は餌と食品両方の用途での中立的な付加価値のある産物、又はその断片である遺伝子又はコーディング領域が挙げられるが、これらに限定されない。目的のヌクレオチド配列又は目的のコーディング領域はまた、薬学上活性のあるタンパク質、たとえば、増殖因子、増殖調節剤、免疫若しくはワクチン植菌に有用な抗体、抗原及びその断片、又はその誘導体をコードする遺伝子を含んでもよい。そのようなタンパク質には、IL−1〜IL−24、IL−26及びIL−27、サイトカイン、エリスロポイエチン(EPO)、インスリン、G−CSF、GM−CSF、hPG−CSF、M−CSF、又はこれらの組み合わせ、インターフェロン、たとえば、インターフェロン−α、インターフェロン−β、インターフェロン−γ、血液凝固因子、たとえば、因子VIII、因子IX、又はtPAhGH、受容体、受容体作動薬、抗体、神経ポリペプチド、インスリン、ワクチン、たとえば、上皮増殖因子、角化細胞増殖因子、形質転換増殖因子、増殖調節剤のような、しかし、これらに限定されない増殖因子、抗原、自己抗原、その断片、又はこれらの組み合わせが挙げられるが、これらに限定されない。
【0071】
目的の遺伝子が、植物体にとって直接又は間接的に毒性である産物をコードするのであれば、本発明の方法を使用することによって、目的の遺伝子を指向された組織内又は植物体の生育の所望の段階で選択的に発現させることによって、植物体全体にわたってそのような毒性を低減してもよい。
【0072】
目的のコーディング領域又は目的のヌクレオチド配列は、形質転換される、又は本発明のヌクレオチド配列、核酸分子、又は遺伝構築物を含む好適な植物体宿主で発現されてもよい。好適な宿主の例には、Arabidopsis、並びにカノーラ、Brassica spp、トウモロコシ、タバコ、アルファルファ、ジャガイモ、朝鮮ニンジン、エンドウ豆、カラスムギ、コメ、大豆、小麦、大麦、ヒマワリ、及び綿を含む農作物が挙げられるが、これらに限定されない。
【0073】
植物体におけるシアル酸の合成、たとえば、Neu5Acの合成は、組み換えNeu5Acリアーゼ又はNeu5Acシンターゼを発現させることによって実証された。E.coliに由来するNeu5Acリアーゼ及びC.jejuniに由来するNeuB2はそれぞれ、アグロバクテリウムが介在した形質転換によって、又は植物細胞で一過性に発現させた場合、タバコBY2細胞、アルファルファ植物体の細胞質で発現された。組換えタンパク質の分解が認められなかったということは、この区分で酵素が安定であることを示している。BY2細胞で発現されたNeu5Acリアーゼは、可逆的反応にてNeu5AcのManNAcとピルビン酸塩への開裂を触媒することができた。ピルビン酸塩とManNAcの存在下でNeu5Acの合成も認められた。植物体の細胞質のpH及び最も重要な作物の温度に一致するpH7及び25〜37℃にてNeu5Acリアーゼは生物学的に活性があった。さらに、外因性Neu5Acの存在下で行われた供給実験によって酵素が植物体で機能的であることが明らかにされた。
【0074】
Neu5Acシンターゼである、C.jejuni由来のNeuB2は、タバコBY2で発現された場合、D−ManNAcとPEPの存在下でNeu5Acを合成することが認められた。アルファルファ植物体でのNeuB2の発現によって結果として機能的酵素の蓄積が生じた。従って、植物体における微生物のNeu5Acリアーゼ及びNeu5Acシンターゼの発現は結果として、Neu5Acを合成できる酵素の細胞質での産生を生じる。
【0075】
適切なアミノ糖基質と共にNeu5Acリアーゼ又はNeu5Acシンターゼを供給するために、内因性GlcNAcをManNAcに変換することができるエピメラーゼを植物体で共発現させてもよい。この点で、タバコBY2細胞における機能的CMP−Neu5Acシンターゼ及びCMP−Neu5Acトランスポーターの発現が報告されている(Misaki et al., 2006)。NeuB2と共にCMP−Neu5Acシンターゼ、Neu5Acトランスポーター、又はCMP−Neu5Acシンターゼ及びNeu5Acトランスポーターの両方を共発現させることによって、Neu5Acの産生を高めてもよい。N−アセチルマンノサミン(ManNAc)の植物体内での合成は幾つかの方法によって達成されてもよい。たとえば、不可逆的反応にてUDP−GlcNAcをManNAcに変換するUDP−GlcNAc2エピメラーゼ、たとえば、細菌のUDP−GlcNAc2エピメラーゼを植物体内で発現させることによってManNAcを合成してもよい。UDP−GlcNAcは、N−グリカン合成経路を供給するので細胞質に存在する。そのほかの供給元からのGlcNAc2エピメラーゼを発現させることによってManNAcの合成を達成してもよい。或いは、ManNAc−6−ホスフェートを形成し、その後、ホスファターゼで加水分解してもよい(トランスジェニック植物体中で)。このアプローチによって、GlcNAc−6−ホスフェート2エピメラーゼ、たとえば、微生物のGlcNAc−6−ホスフェート2エピメラーゼ又は哺乳類のUDP−GlcNAc−2エピメラーゼ/ManNAcキナーゼが植物体内で発現される。
【0076】
細胞質のシアル酸からのN−グリカンのシアル化を確実にするには、細菌又は哺乳類のCMP−Neu5Acシンターゼ、哺乳類のCMP−Neu5Acトランスポーター、哺乳類のガラクトシルトランスフェラーゼ及び哺乳類のシアリルトランスフェラーゼを植物体内で共発現してもよい(Misaki, R. et al., 2006)。
【0077】
以下の実施例にて本発明をさらに説明する。
【実施例】
【0078】
方法
合成FLAG配列ポリペプチド(Arg−Tyr−Lys−Asp−Asp−Asp−Asp−Lys:配列番号1)に対するポリクローナル抗体をEurogentec(Seraing, Belgium)のウサギで調製した。C18Bond−ElutカートリッジはVarian(Sugarland, TX)から得た。タバコ細胞及びM.sativaのクローニング実験及び形質転換には、それぞれ、Escherichia coliDH−5α及びAgrobacterium tumefaciensLBA4404を用いた。Gomordら(1998)で記載されたように、Nicotiana tabacum Bright Yellow2(BY2)細胞を増殖させた。
【0079】
(Neu5Acリアーゼ及びNeuB2の遺伝子のクローニング及び植物体の発現ベクターの構築)
Neu5Acリアーゼ及びNeuB2の遺伝子をPCRで増幅した。Neu5Acリアーゼの遺伝子は、以下のプライマーを用いたPCRによってE.coliのK1ゲノムDNAから増幅させた:
リアーゼP1:
5’−AATAGGCCATTACGGCCATGGCAACGAATTTACGTGG−3’(配列番号6)、および
リアーゼP2:
5’−AATAGGCCGAGGCGGCCTCACCCGCGCTCTTGCAT−3’(配列番号7)。
neuB2遺伝子については、以下のプライマーを用いてDNA断片を得た:
neuB2−P1:
5’−AATAGGCCATTACGGCCATGAAAAAAACTTTAATC−3’(配列番号8)、および
neuB2−P2:
5’−AATAGGCCGAGGCGGCCTTACTCACGGATAAGCTC−3’(配列番号9)。
【0080】
これら増幅したDNAを、Neu5Acリアーゼ遺伝子用のプラスミドベクターpDNR−LIB及びneuB2遺伝子用のバイナリープラスミドベクターpCAMBIA2300のカリフラワーモザイクウイルス(CaMV)35Sプロモーターの下に置いた。CaMV35Sプロモーター、Neu5Acリアーゼ及びノパリンシンターゼ(Nos)ターミネーターの発現カセットをカナマイシン耐性遺伝子と共に植物発現ベクターpBLTI121(Pagny et al., 2000)に導入した。
【0081】
pBLTINeu5Acリアーゼ−FLAG及びpBLTIneuB2−FLAGプラスミドを生成するために、タンパク質のC末端にコードされたFLAGペプチドで遺伝子を増幅するように設計された以下の4つのプライマーを用いた:
リアーゼ−FLAG−P1:
5’−CGGGGTACCAGAATGGCAACGAATTTACGTGGC−3’(配列番号2)、および
リアーゼ−FLAG−P2:
5’−GCCGAGCTCTCACTTGTCATCGTCATCCTTGTAATCCATCCCGCGCTCTTGCATCAACTG−3’(配列番号3)、
neuB2−FLAG−P1:
5’−CGGGGTACCAGAGAGATGAAAAAAACTTTAATCATCGC−3’(配列番号4)、および
neuB2−FLAG−P2:
5’−GCCGAGCTCTCACTTGTCATCGTCATCCTTGTAATCCATCTCACGGATAAGCTCATCTTC−3’(配列番号5)。
【0082】
以下のプログラム:94℃にて1分間の変性、58℃にて1分間のアニーリング及び72℃にて3分間の重合を30サイクルによるPCRによってこれらの増幅された配列を生成した。PCR産物をpCR−BLUNTII−TOPO(登録商標)(Invitrogen)にてクローニングした。植物細胞で組換えタンパク質を発現する前に、我々は、配列決定によって修飾したcDNA構築物すべてを確認した。その後、挿入物をKpnIとSacIで消化し、KpnI−SacIで消化したpBLTI121にクローニングした。熱ショック形質転換を介して、各ベクター(pBLTI121又はpCAMBIA2300)をAgrobacterium tumefaciensのLBA4404株に導入した。
【0083】
(BY2細胞における発現)
タバコBY2細胞は、MurashigeとSkoog(1962)の培地で維持し、形質転換に用いた。pBLTI121に由来する構築物をアグロバクテリウム(LBA4404)(Hofgen and Willmitzer, 1988)に移入した。100μg/mLのカナマイシンを含有するYEB培地にてトランスジェニックアグロバクテリウム細胞を選抜し、Gromordら(1998)で記載されているように浮遊培養しているタバコの細胞を形質転換するのに用いた。抗生剤(カナマイシン100μg/mL及びセフォタキシム250μg/mL)を含有するMS培地にて形質転換体を選抜し、維持した。各形質転換体からゲノムDNA及びmRNAを調製し、PCR及びRT−PCRによって目的の遺伝子がタバコの浮遊培養細胞に挿入され、そこで発現されていることを確認した。免疫スクリーニングの後、組換えタンパク質を産生している微小カルスを用いてトランスジェニック細胞の浮遊培養を開始した(Gomord et al., 1998)。
【0084】
(アルファルファ植物体における発現)
以下の改変と共に、Tianら(Tian et al., 2002)に記載されるように本質的に、アルファルファの形質転換を行った。Agrobacterium tumefaciensAGL1を用いて、アルファルファ遺伝子型R2336を形質転換した。共培養の工程は、0.8〜1のODで未稀釈の培養物と共に行い、1.5%スクロースの代わりに3%のスクロースをSh2K培地で使用した。
【0085】
(Neu5Acリアーゼ及びNeuB2の活性をアッセイするための細胞抽出物の調製)
4日間培養したBY2浮遊培養細胞1g又はM.sativaの新鮮な葉600ngを回収して、プロテイナーゼ阻害剤(ペプスタチン1μg/mL、E64:1μg/mL及びPMSF:1mM、シグマ)を含有する溶液A(100mMのTris−HCl緩衝液(pH=7.4)にて粉砕した。細胞抽出物を4℃にて10000gで10分間遠心し、硫酸アンモニウム(最終濃度80%)でタンパク質を沈殿させ、スペクトラ/Por(10000Daで切り捨て)と共に、溶液B(Neu5Acリアーゼアッセイ用には100mMのTris−HCl緩衝液(pH=7.4)又はNeuB2用にはpH=8.5、及び10mMのMgCl2)に対して透析した。次いで、タンパク質を酵素アッセイ又は免疫検出に利用した。
【0086】
(Neu5Acリアーゼ−FLAG及びNeuB2−FLAGの免疫検出)
変性緩衝液(20mMのTris−HCl、pH6.8、3%のβ−メルカプトエタノール、5%(v/v)のグリセロール及び1%(w/v)のSDS)にてタンパク質を可溶化し、5分間煮沸して、15%ポリアクリルアミドゲル中でのSDS−PAGEによって分離した。次いで、タンパク質をニトロセルロース膜上に移した。免疫検出のために、FLAGエピトープに対して生成したウサギ抗血清によって膜を探査した。西洋ワサビのペルオキシダーゼを結合したヤギ抗ウサギ抗体と共にインキュベートし、続いて4−クロロナフトールを用いた顕色又は化学発光反応によってタンパク質を検出した。
【0087】
(Neu5Acリアーゼ及びシンターゼのアッセイ)
可溶性の酵素活性は、4mMのPEP、4mMのNADH、20mMのNaHCO3及び10mMのDTEと共に細胞抽出物をインキュベートすることによって測定される。10分後、340nmでの吸収の減少によってNADHの酸化を測定した。Neu5Acリアーゼのリアーゼ活性は、形質転換体細胞抽出物をNeu5Acと共にインキュベートした後、ManNAcの形成を測定することによってアッセイした。プロテイナーゼ阻害剤(ペプスタチン1μg/mL、E64:1μg/mL及びPMSF:1mM、シグマ)及び40mMのNeu5Acを含有する溶液B(100mMのTris−HCl緩衝液(pH=7.4)及び10mMのMgCl2)にて37℃で細胞抽出物を2時間インキュベートした。Neu5Acリアーゼのシンターゼ活性は、形質転換体細胞抽出物をManNAcとピルビン酸塩と共にインキュベートした後、Neu5Acの形成を測定することによってアッセイした。プロテイナーゼ阻害剤(ペプスタチン1μg/mL、E64:1μg/mL及びPMSF:1mM、シグマ)及び20mMのManNAcと40mMのピルビン酸塩を含有する溶液B(100mMのTris−HCl緩衝液(pH=7.4)及び10mMのMgCl2)にて37℃で細胞抽出物を2時間インキュベートした。NeuB2のシンターゼ活性は、形質転換体細胞抽出物をManNAcとPEPと共にインキュベートした後、Neu5Acの形成を測定することによってアッセイした。プロテイナーゼ阻害剤(ペプスタチン1μg/mL、E64:1μg/mL及びPMSF:1mM、シグマ)及び10mMのManNAcと10mMのPEPを含有する溶液B(100mMのTris−HCl緩衝液(pH=7.4)及び10mMのMgCl2)にて37℃で細胞抽出物を2時間インキュベートした。80℃にて5分間加熱することによって反応を停止し、C18Bond−Eluntカートリッジにて水による連続溶出によって精製し、GC−EI−MS分析用に凍結乾燥して得た。
【0088】
(供給実験)
4日間培養したタバコBY2細胞を10mMのNeu5Ac又は30mMのManNAcと共にBY2培地中で37℃にて2日間インキュベートし、それぞれ、Neu5Acリアーゼ又はNeu5Acシンターゼのin vivoでの活性をアッセイした。2日後、Neu5Ac又はManNAcを含まないBY2培地でBY2細胞を洗浄し、回収した。70%エタノール中にて70℃で15分間加熱し、酵素を不活化し、次いで陶器のホモゲナイザーで粉砕した。70℃にて70%エタノールで2回、ホモジネートを洗浄した。残ったペレット及び上清をそれぞれ、細胞壁及び細胞質の遊離の単糖類の代表とみなした。次いで、ガスクロマトグラフィによって上清分画の単糖類を分析した。
【0089】
(GC分析)
酵素のアッセイについては、反応混合物を先ずC18Seppakカートリッジにおける精製工程に提示した。単糖類は100%の水中に溶出された。凍結乾燥の後、500μLの2Mの無水メタノール−HClによる80℃での16時間の加メタノール分解に試料を提示した。メタノールを蒸発した後、20μLの無水酢酸及び20μLのピリジンを添加することによって試料を再びアセチル化した。得られたN−アセチルメチルグリコシド(メチルエステル)を乾燥し、次いでそのTMS−誘導体に変換し、ガスクロマトグラフィで分離した。ガスクロマトグラフは、水素炎イオン化検出器、固定相としてのCP−Sil5CPを伴ったWCOT溶融石英のキャピラリーカラム(長さ25m、i.d.0.25mm)及びガスベクターとしてのヘリウムを備えていた。オーブンの温度プログラムは、120℃にて2分間、1分間当たり10℃で160℃まで、1分間当たり1.5℃で220℃まで、次いで1分間当たり20℃にて280℃までであった。ピークの積分及び標準の単糖類によって確立された反応因子を用いた相当する分子の値の決定によって糖の定量を行った。
【0090】
(植物における一過性発現)
(アグロバクテリウムの増殖)
上述の所望のDNA構築物を担うバイナリーベクターを含有するアグロバクテリウムのクローンは、それぞれ25μg/mL及び50μg/mLのカルベニシリン及びカナマイシンを含有する2mLのYEB培地又はLB培地にて28℃で24時間増殖させた。これら培養物10μLを出発植菌として用いて、25mLのYEB誘導培地(YEB培地、pHを5.6に調整した10mMの2(N−モルフォリノ)エタンスルホン酸(MES)、25mg/Lのカルベニシリン、20μMのカナマイシン)の培養を生成した。回転振盪器(220rpm)インキュベーターにて28℃で18時間、又は600nmでの光学密度が0.8〜1に達するまで後者を増殖させた。
【0091】
(非トランスジェニックタバコの生育)
温室にてNicotiana benthamiana及びNicotiana tabacumの植物体を泥炭系基材中で種子から生育させた。苗木は当初苗床で育て、その後ポットに移した。植物体には一日2回水を与え、各散布にて180ppmの窒素を与えた。温室の条件は、植物体のレベルで20ワット/m2の人工照明にて長日条件(16時間明/8時間暗のサイクル)のもと、昼間25℃及び夜間21℃に保持された。様々な生育段階で植物体を利用することができるが、5〜8週の生育にて優先的に選択された。
【0092】
(タバコにおける構築物の一過性の発現)
本発明では2つの一過性の発現の方法:アグロ−植菌又はアグロ浸潤が使用された。両方の方法では、目的の転移DNA(t−DNA)を担う2つ又は3つのアグロバクテリウムの培養物の混合物が、葉の細胞間間隙に入ることが強いられる。上皮の物理的障壁をいったん越境すると、アグロバクテリウムは近隣の細胞に感染し、植物細胞にt−DNAのコピーを転移させる。この方法によって、核内のt−DNAの継代は一過性であり、t−DNAに存在する遺伝子はエピソームに転写され、mRNAが翻訳され、感染細胞における目的のタンパク質の産生がもたらされる。アグロ浸潤が制御された減圧を用いるのに対して、アグロ植菌法は、シリンジによる加圧を用いてアグロバクテリウム混合物を植物体の内部に挿入する。
【0093】
前に記載されたように調製されたアグロバクテリウムの培養物を、10000gにて8分間遠心し、同一容量の植菌培地(pH5.6に調整され、100μMのアセトシリンゴンで補完された10mMのMgCl2、10mMのMES)に再浮遊し、植菌に先立って室温(RT、23℃)にて1時間保持した。或いは、植菌に先立って4℃にて24時間、浮遊物を保持した。N.benthamiana及びN.tabacumの一過性の形質転換は、以下の改変と共に、Liu及びLomonossoff(2002, Journal of Virological Methods, 105:343−348)に記載されたように本質的に行った。上述の構築物の発現のために、2種のアグロバクテリウム株の混合物を植菌した。第1の株は上述のクローンの1つを含有し、第2の株は、35Sプロモーターの制御のもとでジャガイモウイルスYからのサイレンシングのHcProサプレッサーを含有した。植菌後、植物体を温室でインキュベートした。温度は、昼間、最低23℃及び夜間21℃に保持された。植物体には一日2回水を与え、各散布にて180ppmの窒素を与えた。バイオマスの回収は、4〜8日後に行った。
【0094】
(形質転換されたバイオマスからの可溶性タンパク質抽出物の調製)
バイオマスを回収した後、又は−80℃にて凍結した後、葉を直接分析した。約0.1〜1gのアグロ−植菌された又はアグロ−浸潤させた葉のバイオマスを秤量し、総タンパク液抽出物の生成に使用した。
【0095】
幾つかの抽出方法を用いて、乳鉢と乳棒によって植物組織を粉砕することによって、ポリトロンを用いることによって、又はRetsch製のMixerMill300(MM300)にて粉砕することによって総タンパク抽出物を生成した。0.1〜1gの植物バイオマスを清浄で予備冷却した乳鉢に移した。冷却した抽出緩衝液(2mMのCaCl2及び4%のブタノールを含有する50mMのTris、150mMのNaCl、pH7.4)を1:3(w/v)の定量にて加え、同様に最終濃度、それぞれ1mM及び10μMのPMSF及びキモスタチンを加えた。均質な調製物が得られるまで、乳棒で葉を粉砕した。次いで植物抽出物を1.5mLの微量チューブに移し、4℃にて20,000gで20分間遠心した。或いは、0.1gの植物組織を0.3mLの抽出緩衝液と共に非滅菌の1.5mLの微量チューブに導入した。各チューブにタングステンビーズを加えた。30Hzでの3分間の撹拌サイクルにボックスを提示した。サイクルを2回繰り返した。次いで、植物抽出物を上述のように遠心した。或いは、ポリトロンを用いて、1gのバイオマスを3mLの抽出緩衝液と共に粉砕した。
【0096】
遠心に続いて、上清を清浄な微量チューブに移し、氷上に保持した。最終的に、BSAを参照タンパク質として用いたBradford法によって個々のタンパク抽出物の総タンパク質含量を測定した。
【0097】
(実施例1)
(タバコBY2細胞におけるE.coliのNeu5Acリアーゼの発現)
Escherichia coliK1(受入番号:D00067)由来のNeu5Acリアーゼをコードする遺伝子をタバコBY2細胞に導入した。E.coliK1のNeu5Acリアーゼを含有するプラスミドpBLTI121によるアグロバクテリウムを介在させた形質転換の後、トランスジェニックBY2のカルスを生成した。別の構築物はそのC末端にFLAGエピトープでタグを付け、形質転換体における組換えタンパク質を免疫的に検出できるようにした。カナマイシン耐性によって選抜された形質転換体をmRNAについてRT−PCRによって分析した。Neu5Acリアーゼ転写物を発現している48形質転換体から36及びNeu5Acリアーゼ−FLAG転写物を発現している50形質転換体から30が得られた。さらに高いmRNAの発現レベルを抱くカルスを、Neu5Acリアーゼ活性の性状分析用の浮遊培養に移した。Neu5Acリアーゼ−FLAGの存在は、ウエスタンブロット分析によって、形質転換されたBY2細胞の細胞質タンパク質抽出物にて割り出された。抗FLAG抗体を用いて形質転換した細胞にて32kDaの見かけのMWを持つ単一のタンパク質バンドが特異的に免疫検出された(図1a)。
【0098】
Neu5Acリアーゼ及びNeu5Acリアーゼ−FLAGを発現している浮遊培養されたBY2細胞の可溶性タンパク抽出物にて酵素的分析を行った。抽出物は両方共にリアーゼ活性を示した。タグの付いていないリアーゼを発現している細胞から単離されたタンパク抽出物にてさらなる分析を実施した。先ず、これらの抽出物をNeu5Acの存在下でインキュベートしてリアーゼ活性を調べた。図1b及び1cは、pH7及び37℃にてNeu5Acの非存在下(図1b)又は存在下(図1c)にてNeu5Acリアーゼタンパク質抽出物をインキュベートすることによって形成された最終産物のGC特性を示す。抽出物をNeu5Acと共にインキュベートした場合、3つのシグナル(ピーク1は内因性シグナルの肩部である)が明瞭に検出された。これらのシグナルは、標準のManNAcのピラノース型(ピーク1)及びフラノース型(ピーク2及び3)に類似する保持時間で溶出した。試料の電子衝撃質量分析(GC−EI MS)と併せたガスクロマトグラフィによって、これらシグナルがManNAcp(図1e)及びManNAcf(図1f)の1−O−メチルペルシリル誘導体に対するシグナルに割り振られることが確認された。m/z=173及び186での特徴的なイオンは、普通、アミノ糖について認められるような窒素原子を含有する断片に割り振られた。
【0099】
同様の条件で野生型のタバコBY2細胞に由来する細胞質のタンパク抽出物をNeu5Acとインキュベートしても、ManNAcを形成しなかった(データを示さず)ということは、植物体において内因性のリアーゼが存在しないことを実証している。このデータは、E.coliのNeu5Acリアーゼ遺伝子によって形質転換されたBY2細胞が、Neu5AcをD−ManNAcに開裂できる機能的な酵素を発現したことを示している。
【0100】
組換え酵素の最適pHは、4〜10のpHの範囲で実施されたアッセイにおいて生成されたD−ManNAcのGCの定量化に基づいて約7であることが明らかにされた(データは示さず)。さらに、組換え酵素は、25〜37℃の範囲で見られる高いリアーゼ活性を持つ温度依存性の活性を示した。pH7及び37℃にて、形質転換細胞に由来する可溶性タンパク質抽出物は1時間で、10μMのNeu5Acから0.5μMのManNAcを形成した。15℃未満では、残留する活性のみが検出された。
【0101】
pH7及び37℃にてD−ManNAc及びピルビン酸塩と共に、形質転換されたタバコBY2細胞のタンパク抽出物をインキュベートすることによってNeu5Acを合成する組換えNeu5Acリアーゼの能力が割り出された。図2a及び図2bはそれぞれ、基質の非存在下又は存在下でインキュベートした後の最終産物のGC特性を示す。対照の特性(図2a)と比べた場合、D−ManNAcとピルビン酸塩の存在下で行った反応のGC特性は、Neu5Acについて予測される保持時間でシグナルを示した(図2bでの四角)。このシグナルの電子衝撃質量スペクトル(EI−MS)(図2c)は、窒素含有断片に割り当てられたm/z=186でのイオンと同様にNeu5Ac断片に特異的なm/z=298及び420での特徴的なイオンを示した。このデータは、組換えリアーゼがD−ManNAcとピルビン酸塩の存在下でNeu5Acを合成することができることを示している。
【0102】
10mMのNeu5AcをタバコBY2細胞に供給することによってNeu5Acリアーゼの植物体での活性を測定した。Neu5AcのタバコBY2細胞に対する毒性を調べたところ、ヨウ化プロピジウム及びフルオレセインジアセテートを用いて細胞の生存率を調べることによって48時間にわたって毒性効果は認められなかった。23℃〜37℃の範囲の温度で48時間後のGCにより細胞質の単糖類を分析することによってD−ManNAcの形成を測定した。D−ManNAcはあらゆる処理で検出された(図1d)。GCによるD−ManNAcの定量は、23℃に比べて37℃にてこのアミノ糖の含量で25倍の増加を示した。これらin vivoの実験は、Neu5Acリアーゼが植物体で生物学的に活性を持ち、外因的に供給された基質に作用することができることを明らかにしている。
【0103】
(タバコBY2及びアルファルファ植物体におけるCampylobactor jejuniのNeuB2の発現)
Neu5Acシンターゼである、Campylobactor jejuniに由来するNeuB2(受入番号:NC002163)は、ManNAcとPEPの縮合によるNeu5Acの形成を触媒する。neuB2のDNAを含有するプラスミドpBLTI121によるアグロバクテリウムが介在する形質転換の後、トランスジェニックBY2カルスを生成した。タンパク質の免疫的検出のために、第2の構築物にはそのC末端にFLAGエピトープによるタグを付けた。カナマイシン耐性について選抜した形質転換体をRT−PCRによってmRNAレベルで分析した。最も高いmRNA発現レベルを抱くカルスを分析用の浮遊培養に移した。次いで、NeuB2−FLAGの配列によって形質転換されたBY2細胞から単離した可溶性タンパク抽出物のウエスタンブロット分析によって形質転換されたBY2細胞におけるNeuB2の蓄積を測定した。図3aで説明されるように、抗FLAG抗体は、シンターゼの予測分子量と一致するMW=37kDaにて単一タンパク質バンドを特異的に認識した。neuB2はまた、アグロバクテリウムが介在する形質転換及びin vitroの植物体の再生によってアルファルファ植物体にも導入された(Tian at al., 2002)。34の形質転換された植物体から、29がneuB2の転写物を発現していることが明らかにされた。
【0104】
細菌のNeu5Acシンターゼを発現している、形質転換された細胞及び植物体を分析する前に、内因性のNeu5Acシンターゼ活性の存在を調べた。野生型のタバコBY2細胞及びアルファルファ植物体から得た可溶性タンパク抽出物をD−ManNAc及びPEPと共にインキュベートした。アッセイで形成された単糖類をGCによって分離し、GC−EI−MSによって性状分析した。Neu5Acに割り当てられたピーク及びEI−MSでの特徴的なイオンが検出されなかったということは、植物体がPEPとManNAcを縮合してNeu5Acを形成することができる内因性の酵素を発現していないことを示している。
【0105】
neuB2遺伝子によって形質転換されたタバコBY2細胞及びアルファルファ植物体から単離した可溶性タンパク抽出物と共にD−ManNAcとPEPをインキュベートすることによって植物体で発現された組換えNeuB2のシンターゼ活性を測定した。図3b及び3cは、pH8及び37℃にてD−ManNAcとPEPの非存在下(図3b)又は存在下(図3c)で形質転換されたアルファルファ抽出物をインキュベートすることによって得たGC特性を示す。シンターゼの基質と共にインキュベートした後、Neu5Acについて予測される保持時間にて溶出されたピークが特異的に検出された。このピークのEI−MSは、m/z=298及び420で特徴的なイオンと共に、標準のNeu5Acの1つに類似する断片化パターンを示した。D−ManNacの非存在下でインキュベートした後のGC特性の相当する領域のEI−MSスペクトルにはそれらのイオンは検出されなかった(図3b)。
【0106】
Neu2B又はNeu2B−FLAG配列を発現しているタバコBY2細胞の分析によって同じ結果が得られた。
【0107】
従って、タバコBY2細胞及びアルファルファ植物体両方におけるneuB2の発現は、結果として機能的なNeu5Acシンターゼの産生を生じる。
【0108】
【表1−1】
【0109】
【表1−2】
【0110】
【表1−3】
【0111】
【表1−4】
引用はすべて参照によって本明細書に援用される。
【0112】
1以上の実施態様に関して本発明を説明してきた。しかしながら、クレームで定義されるような本発明の範囲から逸脱することなく多数の変化及び改変が可能であることが、当業者に明らかになるであろう。
【図面の簡単な説明】
【0113】
【図1】図1aは、抗FLAG抗体を用いた、野生型(ライン1)又はNeu5Acリアーゼ−FLAGを発現しているトランスジェニックタバコBY2細胞(ライン2)から抽出した可溶性タンパク質のウエスタンブロット分析を示す。図1b及び図1cは、Neu5Acを発現しているタバコBY2から単離された細胞質タンパク質を単独で(図1b)又はNeu5Acと共に(図1c)pH7にて37℃でインキュベートした後、得られた最終産物のガスクロマトグラフィの特性を示す。図1dは、10mMの外因性Neu5Acと共に37℃にて48時間養ったNue5Acを発現しているタバコBY2細胞の細胞質単糖類のGC特性を示す。図1e及び図1fは、特性(図1c)で検出されたピーク1(図1e)並びにピーク2及び3(図1f)の電子衝撃質量スペクトルを示す。D−ManNAcの1−O−メチルペルシリル誘導体の主要な断片イオンが示される。
【図2】図2a及び図2bは、Neu5Acを発現しているタバコBY2から単離された細胞質タンパク質を単独で(図2a)又はD−ManNAc及びピルビン酸塩と共に(図2b)pH7にて37℃でインキュベートした後、得られた最終産物のガスクロマトグラフィの特性を示す。図2cは、特性(図2b)で現れているピークの電子衝撃質量スペクトルを示す。N−アセチルノイラミン酸の1−O−メチルメチルエステルペルシリル誘導体の主要断片イオンが示される。
【図3】図3aは、抗FLAG抗体を用いた、野生型(ライン1)又はNeuB2−FLAGを発現しているトランスジェニックタバコBY2細胞(ライン2)から抽出した可溶性タンパク質のウエスタンブロット分析を示す。図3b及び図3cは、NeuB2を発現しているアルファルファ植物体の葉から単離された可溶性タンパク質を単独で(図3b)又はD−ManNAc及びPEPと共に(図3c)pH8にて37℃でインキュベートした後、得られた最終産物のガスクロマトグラフィの特性を示す。図3dは、特性(図3c)で現れているピークの電子衝撃質量スペクトルを示す。N−アセチルノイラミン酸の1−O−メチルメチルエステルペルシリル誘導体の主要断片イオンが示される。
【特許請求の範囲】
【請求項1】
シアル酸を合成する方法であって、
i)N−アセチルノイラミン酸(Neu5Ac)シンターゼ又はNeu5Acリアーゼをコードするヌクレオチド配列を含む植物体を提供する工程であって、該ヌクレオチド配列は、該植物体中で活性のある調節領域と作動可能に連結されている、工程、
ii)該植物体を生育させ、該ヌクレオチド配列を発現させ、それによってシアル酸を合成する工程
を包含する、方法。
【請求項2】
前記シアル酸がNeu5Acであり、前記生育させる工程の後、前記Neu5Acが前記植物体から回収される、請求項1に記載の方法。
【請求項3】
前記調節領域が、構成的プロモーター、誘導性プロモーター、組織特異的プロモーター及び発育プロモーターからなる群より選択される、請求項1に記載の方法。
【請求項4】
前記提供する工程では、前記植物体が、該植物体内で活性のある1又は1を超える第2の調節領域に作動可能に連結された、エピメラーゼ、CMP−Neu5Acシンターゼ、又はCMP−Neu5Acトランスポーターのうちの1又は1を超えるものをコードする第2のヌクレオチド配列をさらに含み、該第2のヌクレオチド配列は、該ヌクレオチド配列の発現と共に共発現される、請求項1に記載の方法。
【請求項5】
前記第2の調節領域が、構成的プロモーター、誘導性プロモーター、組織特異的プロモーター及び発育プロモーターからなる群より選択される、請求項4に記載の方法。
【請求項6】
前記Neu5Acシンターゼ若しくはNeu5Acリアーゼをコードするヌクレオチド配列、前記エピメラーゼ、CMP−Neu5Acシンターゼ、若しくはCMP−Neu5Acトランスポーターのうちの1若しくは1を超えるものをコードする第2のヌクレオチド配列、又は該ヌクレオチド配列及び該第2のヌクレオチド配列の両方は、前記植物体内での発現のためにコドンが最適化されている、請求項4に記載の方法。
【請求項7】
目的のタンパク質を産生する方法であって、
i)Neu5Acシンターゼ、Neu5Acリアーゼ、エピメラーゼ、CMP−Neu5Acシンターゼ、CMP−Neu5Acトランスポーター、ガラクトシルトランスフェラーゼ、シアリルトランスフェラーゼをコードする1又は1を超える第1のヌクレオチド配列、及び該目的のタンパク質をコードする第2のヌクレオチド配列を発現する植物体を提供する工程と、
ii)該植物体を生育させ、第1及び第2のヌクレオチド配列を発現させ、それによって該目的のタンパク質を産生する工程であって、ここで該目的のタンパク質がシアル化されている、工程
を包含する、方法。
【請求項8】
前記シアル化された目的のタンパク質は、ジ−、トリ−又はテトラ−分岐N−グリカンを含む、請求項7に記載の方法。
【請求項9】
前記生育させる工程の後、前記植物体から前記シアル化されたタンパク質が抽出される、請求項7に記載の方法。
【請求項10】
前記シアル化された目的のタンパク質が単離され、精製される、請求項9に記載の方法。
【請求項11】
前記生育させる工程の後、目的のシアル化されたタンパク質を含む植物物質が被験体に経口投与される、請求項7に記載の方法。
【請求項12】
前記生育させる工程の後、前記植物体が最小限に加工され、前記シアル化された目的のタンパク質を含む最小限に加工された植物物質が被験体に経口投与される、請求項7に記載の方法。
【請求項13】
前記シアル化された目的のタンパク質が被験体に投与される、請求項10に記載の方法。
【請求項14】
Neu5Acシンターゼ、Neu5Acリアーゼ、エピメラーゼ、CMP−Neu5Acシンターゼ、CMP−Neu5Acトランスポーター、ガラクトシルトランスフェラーゼ、シアリルトランスフェラーゼをコードする1若しくは1を超える第1のヌクレオチド配列、前記目的のタンパク質をコードする第2のヌクレオチド配列、又は該第1のヌクレオチド配列及び該第2のヌクレオチド配列の両方は、前記植物体内での発現のためにコドンが最適化されている、請求項7に記載の方法。
【請求項15】
前記植物体内で活性のある調節領域と作動可能に連結されたNeu5Acシンターゼ又はNeu5Acリアーゼをコードするヌクレオチド配列を含む、植物体、植物細胞又は種子。
【請求項16】
前記植物体内、植物細胞内又は前記種子内で活性のある1又は1を超える第2の調節領域に作動可能に連結された、エピメラーゼ、CMP−Neu5Acシンターゼ、CMP−Neu5Acトランスポーター、ガラクトシルトランスフェラーゼ及びシアリルトランスフェラーゼのうちの1又は1を超えるものをコードする第2のヌクレオチド配列をさらに含む、請求項15に記載の植物体、植物細胞又は種子。
【請求項17】
前記第1及び第2の調節領域が、構成的プロモーター、誘導性プロモーター、組織特異的プロモーター及び発育プロモーターからなる群より選択される、請求項16に記載の植物体、植物細胞又は種子。
【請求項18】
Neu5Acシンターゼ若しくはNeu5Acリアーゼをコードするヌクレオチド配列、前記エピメラーゼ、CMP−Neu5Acシンターゼ、若しくはCMP−Neu5Acトランスポーターのうちの1若しくは1を超えるものをコードする第2のヌクレオチド配列、又は該ヌクレオチド配列及び第2のヌクレオチド配列の両方は、前記植物体内、前記植物細胞内又は前記種子内での発現のためにコドンが最適化されている、請求項16に記載の植物体、植物細胞又は種子。
【請求項19】
Neu5Acシンターゼ、Neu5Acリアーゼ、エピメラーゼ、CMP−Neu5Acシンターゼ、CMP−Neu5Acトランスポーター、ガラクトシルトランスフェラーゼ、およびシアリルトランスフェラーゼをコードする1又は1を超える第1のヌクレオチド配列、ならびに目的のタンパク質をコードする第2のヌクレオチド配列を含む植物体、植物細胞又は種子であって、該第1のヌクレオチド配列及び該第2のヌクレオチド配列は、該植物体内、該植物細胞内又は該種子内で活性のある1又は1を超える調節領域に作動可能に連結される、植物体、植物細胞又は種子。
【請求項20】
前記1又は1を超える調節領域が、構成的プロモーター、誘導性プロモーター、組織特異的プロモーター及び発育プロモーターからなる群より選択される、請求項19に記載の植物体、植物細胞又は種子。
【請求項21】
前記Neu5Acシンターゼ、Neu5Acリアーゼ、エピメラーゼ、CMP−Neu5Acシンターゼ、CMP−Neu5Acトランスポーター、ガラクトシルトランスフェラーゼ、シアリルトランスフェラーゼをコードする1若しくは1を超える第1のヌクレオチド配列、目的のタンパク質をコードする第2のヌクレオチド配列、又は該第1のヌクレオチド配列及び該第2のヌクレオチド配列の両方は、前記植物体内、植物細胞内又は前記種子内での発現のためにコドンが最適化されている、請求項19に記載の植物体、植物細胞又は種子。
【請求項22】
シアル酸を合成する方法であって、
N−アセチルノイラミン酸(Neu5Ac)シンターゼ又はNeu5Acリアーゼをコードするヌクレオチド配列によって植物体又は該植物体の一部を一過性に形質転換する工程であって、該ヌクレオチド配列は、該植物体で活性のある調節領域と作動可能に連結されている、工程と、
該ヌクレオチド配列を発現させ、それによってシアル酸を合成する工程と
を包含する、方法。
【請求項23】
前記シアル酸がNeu5Acであり、前記発現する工程の後、前記Neu5Acが前記植物体又は前記植物体の一部から回収される、請求項22に記載の方法。
【請求項24】
前記植物体又は該植物体の一部を一過性に形質転換する工程において、該植物体内で活性のある1又は1を超える第2の調節領域に作動可能に連結された、エピメラーゼ、CMP−Neu5Acシンターゼ、又はCMP−Neu5Acトランスポーターのうちの1又は1を超えるものをコードする第2のヌクレオチド配列をさらに含み、該第2のヌクレオチド配列が、該ヌクレオチド配列の発現と共に共発現される、請求項22に記載の方法。
【請求項25】
目的のタンパク質を産生する方法であって、
i)Neu5Acシンターゼ、Neu5Acリアーゼ、エピメラーゼ、CMP−Neu5Acシンターゼ、CMP−Neu5Acトランスポーター、ガラクトシルトランスフェラーゼ、シアリルトランスフェラーゼをコードする1又は1を超える第1のヌクレオチド配列、および該目的のタンパク質をコードする第2のヌクレオチド配列を発現する構築物によって植物体又は該植物体の一部を一過性に形質転換することと、
ii)該目的のタンパク質を産生する工程であって、ここで該目的のタンパク質がシアル化されている、工程
を包含する、方法。
【請求項26】
前記シアル化される目的のタンパク質が、ジ−、トリ−又はテトラ−分岐N−グリカンを含む、請求項25に記載の方法。
【請求項27】
前記産生する工程の後、前記シアル化されたタンパク質が前記植物体又は該植物体の一部から抽出される、請求項25に記載の方法。
【請求項28】
前記シアル化された目的のタンパク質が単離され、精製される、請求項27に記載の方法。
【請求項29】
前記産生する工程の後、前記シアル化された目的のタンパク質を含む植物物質が被験体に経口投与される、請求項25に記載の方法。
【請求項30】
前記産生する工程の後、前記植物体又は該植物体の一部が最小限に加工されて最小限に加工された植物物質が作製され、該シアル化された目的のタンパク質を含む最小限に加工された植物物質が被験体に経口投与される、請求項25に記載の方法。
【請求項1】
シアル酸を合成する方法であって、
i)N−アセチルノイラミン酸(Neu5Ac)シンターゼ又はNeu5Acリアーゼをコードするヌクレオチド配列を含む植物体を提供する工程であって、該ヌクレオチド配列は、該植物体中で活性のある調節領域と作動可能に連結されている、工程、
ii)該植物体を生育させ、該ヌクレオチド配列を発現させ、それによってシアル酸を合成する工程
を包含する、方法。
【請求項2】
前記シアル酸がNeu5Acであり、前記生育させる工程の後、前記Neu5Acが前記植物体から回収される、請求項1に記載の方法。
【請求項3】
前記調節領域が、構成的プロモーター、誘導性プロモーター、組織特異的プロモーター及び発育プロモーターからなる群より選択される、請求項1に記載の方法。
【請求項4】
前記提供する工程では、前記植物体が、該植物体内で活性のある1又は1を超える第2の調節領域に作動可能に連結された、エピメラーゼ、CMP−Neu5Acシンターゼ、又はCMP−Neu5Acトランスポーターのうちの1又は1を超えるものをコードする第2のヌクレオチド配列をさらに含み、該第2のヌクレオチド配列は、該ヌクレオチド配列の発現と共に共発現される、請求項1に記載の方法。
【請求項5】
前記第2の調節領域が、構成的プロモーター、誘導性プロモーター、組織特異的プロモーター及び発育プロモーターからなる群より選択される、請求項4に記載の方法。
【請求項6】
前記Neu5Acシンターゼ若しくはNeu5Acリアーゼをコードするヌクレオチド配列、前記エピメラーゼ、CMP−Neu5Acシンターゼ、若しくはCMP−Neu5Acトランスポーターのうちの1若しくは1を超えるものをコードする第2のヌクレオチド配列、又は該ヌクレオチド配列及び該第2のヌクレオチド配列の両方は、前記植物体内での発現のためにコドンが最適化されている、請求項4に記載の方法。
【請求項7】
目的のタンパク質を産生する方法であって、
i)Neu5Acシンターゼ、Neu5Acリアーゼ、エピメラーゼ、CMP−Neu5Acシンターゼ、CMP−Neu5Acトランスポーター、ガラクトシルトランスフェラーゼ、シアリルトランスフェラーゼをコードする1又は1を超える第1のヌクレオチド配列、及び該目的のタンパク質をコードする第2のヌクレオチド配列を発現する植物体を提供する工程と、
ii)該植物体を生育させ、第1及び第2のヌクレオチド配列を発現させ、それによって該目的のタンパク質を産生する工程であって、ここで該目的のタンパク質がシアル化されている、工程
を包含する、方法。
【請求項8】
前記シアル化された目的のタンパク質は、ジ−、トリ−又はテトラ−分岐N−グリカンを含む、請求項7に記載の方法。
【請求項9】
前記生育させる工程の後、前記植物体から前記シアル化されたタンパク質が抽出される、請求項7に記載の方法。
【請求項10】
前記シアル化された目的のタンパク質が単離され、精製される、請求項9に記載の方法。
【請求項11】
前記生育させる工程の後、目的のシアル化されたタンパク質を含む植物物質が被験体に経口投与される、請求項7に記載の方法。
【請求項12】
前記生育させる工程の後、前記植物体が最小限に加工され、前記シアル化された目的のタンパク質を含む最小限に加工された植物物質が被験体に経口投与される、請求項7に記載の方法。
【請求項13】
前記シアル化された目的のタンパク質が被験体に投与される、請求項10に記載の方法。
【請求項14】
Neu5Acシンターゼ、Neu5Acリアーゼ、エピメラーゼ、CMP−Neu5Acシンターゼ、CMP−Neu5Acトランスポーター、ガラクトシルトランスフェラーゼ、シアリルトランスフェラーゼをコードする1若しくは1を超える第1のヌクレオチド配列、前記目的のタンパク質をコードする第2のヌクレオチド配列、又は該第1のヌクレオチド配列及び該第2のヌクレオチド配列の両方は、前記植物体内での発現のためにコドンが最適化されている、請求項7に記載の方法。
【請求項15】
前記植物体内で活性のある調節領域と作動可能に連結されたNeu5Acシンターゼ又はNeu5Acリアーゼをコードするヌクレオチド配列を含む、植物体、植物細胞又は種子。
【請求項16】
前記植物体内、植物細胞内又は前記種子内で活性のある1又は1を超える第2の調節領域に作動可能に連結された、エピメラーゼ、CMP−Neu5Acシンターゼ、CMP−Neu5Acトランスポーター、ガラクトシルトランスフェラーゼ及びシアリルトランスフェラーゼのうちの1又は1を超えるものをコードする第2のヌクレオチド配列をさらに含む、請求項15に記載の植物体、植物細胞又は種子。
【請求項17】
前記第1及び第2の調節領域が、構成的プロモーター、誘導性プロモーター、組織特異的プロモーター及び発育プロモーターからなる群より選択される、請求項16に記載の植物体、植物細胞又は種子。
【請求項18】
Neu5Acシンターゼ若しくはNeu5Acリアーゼをコードするヌクレオチド配列、前記エピメラーゼ、CMP−Neu5Acシンターゼ、若しくはCMP−Neu5Acトランスポーターのうちの1若しくは1を超えるものをコードする第2のヌクレオチド配列、又は該ヌクレオチド配列及び第2のヌクレオチド配列の両方は、前記植物体内、前記植物細胞内又は前記種子内での発現のためにコドンが最適化されている、請求項16に記載の植物体、植物細胞又は種子。
【請求項19】
Neu5Acシンターゼ、Neu5Acリアーゼ、エピメラーゼ、CMP−Neu5Acシンターゼ、CMP−Neu5Acトランスポーター、ガラクトシルトランスフェラーゼ、およびシアリルトランスフェラーゼをコードする1又は1を超える第1のヌクレオチド配列、ならびに目的のタンパク質をコードする第2のヌクレオチド配列を含む植物体、植物細胞又は種子であって、該第1のヌクレオチド配列及び該第2のヌクレオチド配列は、該植物体内、該植物細胞内又は該種子内で活性のある1又は1を超える調節領域に作動可能に連結される、植物体、植物細胞又は種子。
【請求項20】
前記1又は1を超える調節領域が、構成的プロモーター、誘導性プロモーター、組織特異的プロモーター及び発育プロモーターからなる群より選択される、請求項19に記載の植物体、植物細胞又は種子。
【請求項21】
前記Neu5Acシンターゼ、Neu5Acリアーゼ、エピメラーゼ、CMP−Neu5Acシンターゼ、CMP−Neu5Acトランスポーター、ガラクトシルトランスフェラーゼ、シアリルトランスフェラーゼをコードする1若しくは1を超える第1のヌクレオチド配列、目的のタンパク質をコードする第2のヌクレオチド配列、又は該第1のヌクレオチド配列及び該第2のヌクレオチド配列の両方は、前記植物体内、植物細胞内又は前記種子内での発現のためにコドンが最適化されている、請求項19に記載の植物体、植物細胞又は種子。
【請求項22】
シアル酸を合成する方法であって、
N−アセチルノイラミン酸(Neu5Ac)シンターゼ又はNeu5Acリアーゼをコードするヌクレオチド配列によって植物体又は該植物体の一部を一過性に形質転換する工程であって、該ヌクレオチド配列は、該植物体で活性のある調節領域と作動可能に連結されている、工程と、
該ヌクレオチド配列を発現させ、それによってシアル酸を合成する工程と
を包含する、方法。
【請求項23】
前記シアル酸がNeu5Acであり、前記発現する工程の後、前記Neu5Acが前記植物体又は前記植物体の一部から回収される、請求項22に記載の方法。
【請求項24】
前記植物体又は該植物体の一部を一過性に形質転換する工程において、該植物体内で活性のある1又は1を超える第2の調節領域に作動可能に連結された、エピメラーゼ、CMP−Neu5Acシンターゼ、又はCMP−Neu5Acトランスポーターのうちの1又は1を超えるものをコードする第2のヌクレオチド配列をさらに含み、該第2のヌクレオチド配列が、該ヌクレオチド配列の発現と共に共発現される、請求項22に記載の方法。
【請求項25】
目的のタンパク質を産生する方法であって、
i)Neu5Acシンターゼ、Neu5Acリアーゼ、エピメラーゼ、CMP−Neu5Acシンターゼ、CMP−Neu5Acトランスポーター、ガラクトシルトランスフェラーゼ、シアリルトランスフェラーゼをコードする1又は1を超える第1のヌクレオチド配列、および該目的のタンパク質をコードする第2のヌクレオチド配列を発現する構築物によって植物体又は該植物体の一部を一過性に形質転換することと、
ii)該目的のタンパク質を産生する工程であって、ここで該目的のタンパク質がシアル化されている、工程
を包含する、方法。
【請求項26】
前記シアル化される目的のタンパク質が、ジ−、トリ−又はテトラ−分岐N−グリカンを含む、請求項25に記載の方法。
【請求項27】
前記産生する工程の後、前記シアル化されたタンパク質が前記植物体又は該植物体の一部から抽出される、請求項25に記載の方法。
【請求項28】
前記シアル化された目的のタンパク質が単離され、精製される、請求項27に記載の方法。
【請求項29】
前記産生する工程の後、前記シアル化された目的のタンパク質を含む植物物質が被験体に経口投与される、請求項25に記載の方法。
【請求項30】
前記産生する工程の後、前記植物体又は該植物体の一部が最小限に加工されて最小限に加工された植物物質が作製され、該シアル化された目的のタンパク質を含む最小限に加工された植物物質が被験体に経口投与される、請求項25に記載の方法。
【図1】

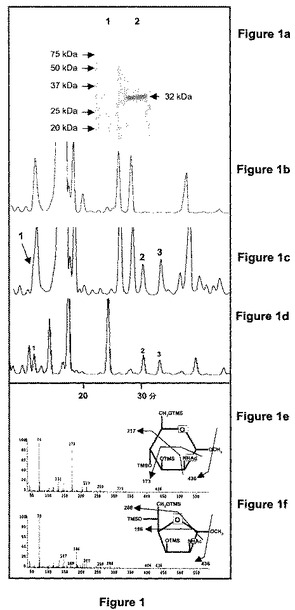
【図2】
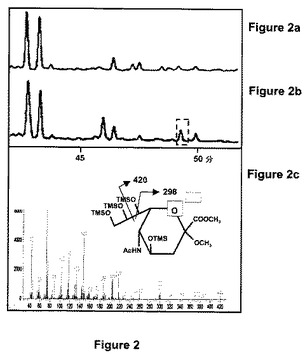
【図3】

【図2】
【図3】
【公表番号】特表2009−525729(P2009−525729A)
【公表日】平成21年7月16日(2009.7.16)
【国際特許分類】
【出願番号】特願2008−553589(P2008−553589)
【出願日】平成19年2月9日(2007.2.9)
【国際出願番号】PCT/CA2007/000197
【国際公開番号】WO2007/090290
【国際公開日】平成19年8月16日(2007.8.16)
【出願人】(502121395)メディカゴ インコーポレイテッド (13)
【出願人】(508242056)センター ナショナル デ ラ ルシェルシュ サイエンティフィック (6)
【Fターム(参考)】
【公表日】平成21年7月16日(2009.7.16)
【国際特許分類】
【出願日】平成19年2月9日(2007.2.9)
【国際出願番号】PCT/CA2007/000197
【国際公開番号】WO2007/090290
【国際公開日】平成19年8月16日(2007.8.16)
【出願人】(502121395)メディカゴ インコーポレイテッド (13)
【出願人】(508242056)センター ナショナル デ ラ ルシェルシュ サイエンティフィック (6)
【Fターム(参考)】
[ Back to top ]