
脊椎動物のDelta遺伝子のヌクレオチドおよびタンパク質配列およびそれに基づく方法

【課題】脊椎動物のDelta遺伝子のヌクレオチド配列、それらによりコードされたタンパク質のアミノ酸配列、並びにその誘導体(例えば、断片)および類似体を提供する。
【解決手段】特定の態様において、脊椎動物デルタタンパク質はヒト・タンパク質である。さらに、デルタタンパク質の細胞内ドメイン、細胞外ドメイン、DSLドメイン、DSLドメインのアミノ末端側のドメイン、膜貫通領域、もしくは1以上のEGF様反復ドメイン、またはこれらの任意の組合せを含むがこれらに限らない、デルタタンパク質の1以上のドメインを含むデルタの断片(並びにその誘導体および類似体)に関する。さらに、デルタ、その誘導体および類似体に対する抗体も示す。デルタタンパク質、その誘導体および類似体の、例えば組換え法による、産生方法も示す。治療および診断法、並びに医薬組成物も示す。
【解決手段】特定の態様において、脊椎動物デルタタンパク質はヒト・タンパク質である。さらに、デルタタンパク質の細胞内ドメイン、細胞外ドメイン、DSLドメイン、DSLドメインのアミノ末端側のドメイン、膜貫通領域、もしくは1以上のEGF様反復ドメイン、またはこれらの任意の組合せを含むがこれらに限らない、デルタタンパク質の1以上のドメインを含むデルタの断片(並びにその誘導体および類似体)に関する。さらに、デルタ、その誘導体および類似体に対する抗体も示す。デルタタンパク質、その誘導体および類似体の、例えば組換え法による、産生方法も示す。治療および診断法、並びに医薬組成物も示す。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、1995年06月28日に出願された米国仮出願(出願番号60/000,589)について優
先権を主張する。この仮出願は、参考文献としてその全体が本明細書中に取込まれるもの
である。
【0002】
1.はじめに
本出願は、脊椎動物のDelta遺伝子およびそれがコードするタンパク質産物、ならびに
その誘導体および類似体に関する。脊椎動物のデルタタンパク質、誘導体、および抗体の
産生もまた提供される。本発明はさらに、治療用組成物と検査方法および治療方法に関す
る。
【背景技術】
【0003】
2.発明の背景
ショウジョウバエ(Drosophila)における遺伝子の分析は、発生経路の複雑度を詳細に分析し、相互作用している遺伝子座を同定する上で非常に有用であった。しかしながら、遺伝的相互作用の根底にあるプロセスの精確な性質を理解するためには、問題となっている遺伝子のタンパク質産物の知識が必要である。
【0004】
脊椎動物の中枢神経系は、異なる細胞の型の完全な混合物であり、そのほぼすべてが同じ細胞源(神経板およびそれに続く神経管を形成する神経上皮)からつくられている。この細胞のシートにおいて、いくつかが神経細胞になるにもかかわらず他の細胞が非神経細胞のままであるように、神経発生を制御する機構は何なのであろう?この答えは、脊椎動物については、実質的には知られていないが、多くの細胞の相互作用と神経発生の間の細胞の予定運命の決定を制御する遺伝子が、ショウジョウバエにおいて十分に特徴付けられている(Campos-Ortega,1993,J.Neurobiol.24:1305-1327)。神経発生の解剖学的内容の全貌は、昆虫と脊椎動物とでは非常に異なるが、細胞レベルにおいては、類似のイベントが保存された分子機構を介して生じているという可能性が残されている。発生学的、遺伝的そして分子的な証拠から、ショウジョウバエにおける初期段階の外胚葉の分化は、細胞の相互作用に依存するということが示唆された(Doeand Goodman, 1985,Dev.Biol.111:206-219; Technau

Neurogenet.2:291-308;de la Concha et al.,1988,Genetics 118:499-508; Xu et al.,1990,Genes Dev.4:464-475;Artavanis-Tsakonas,1988,Trends Genet.4:95-100)。突然変異の分析より、接合的に作用する小さな群の遺伝子、いわゆる神経遺伝子座が上皮と神経経路との間の外胚葉細胞の選択に影響を与えるということが明らかになった(Poulson,1937,Proc.Natl.Acad.Sci.23:133-137;Lehmann et
Wilhelm Roux'sArch.Dev.Biol.193:283-295; Wieschaus et al.,1984,Wilhelm
Roux's Arch.Dev.Biol.193:267-282)。接合神経遺伝子座(Notch(N),Delta(D),mastermind(mam),Enhancerof Split(E(spl)),neuralized(neu),およびbigbrain(bib)のいずれかにおけるヌル突然変異は、腹部および側面の表皮の構造の損失(expense)における神経系の過栄養を生じさせる。この効果は、表皮前駆細胞を神経系に間違ったルートで送ることによるものであり、そして神経遺伝子の機能が神経の予定運命から表皮の予定運命へと神経領域の範囲内で細胞の流れを変えるために必要であることを暗示する。
【0005】
神経前駆体は連続的な波(successive waves)の神経発生の間、神経上皮からショウジョウバエの胚で生じる(Campos-Ortega &Hartenstein,1985,キイロショウジョウバエの胚の発生(Springer-Verlarg,Berlin;New York);Doe,1992,Development116:855-863)。これらの細胞の産生パターンは、前神経遺伝子および神経遺伝子の活性によって、主として決定される。前神経遺伝子は細胞のクラスターを神経の予定運命に前もって処理するが(Skeath& Carroll,1994,Faseb J.8:714-21でレビューされた)、クラスター中のあるサブセットだけが神経前駆体になる。この制限は、神経遺伝子の作用によるものであり、この神経遺伝子は側面阻害(細胞のシグナリングの阻害のある型)を仲介し、それによって神経の予定運命に拘束される細胞が、これに隣接する細胞を拘束されないままにするかまたは非神経系に入るように強制する(Artavanis-Tsakonas& Simpson,1991,Trends Genet.7:403-408; Doe & Goodman,1985,Dev.BIol.111:206-219)。側面阻害をできないようにする突然変異は、神経細胞、すなわち、「神経の」表現型の過剰産生の原因となる(Lehmannet al.,1981,Roux's Arch.Dev.Biol.190:226-229; Lehmann et al.,Roux's Arch.Dev.Biol.192:62-74)。ショウジョウバエにおいては、阻害シグナルはDelta神経遺伝子によってコードされる膜貫通タンパク質によって送達され、このタンパク質は発生期の神経細胞によって表示される(Heitzler& Simpson,1991,Cell 64:1083-1092)。隣接する細胞は、神経遺伝子Notchによってコードされる、膜貫通受容体タンパク質を発現する(Fortini& Artavanis-Tsakonas,1993,Cell 75:1245-1247)。Delta遺伝子はNotch遺伝子座と相互作用可能な遺伝的単位として同定された(Xuet al.,1990,Genes Dev.4:464-475)。
【0006】
突然変異の分析はまた、神経遺伝子の作用が多面発現性であり、唯一胚発生に限定されるものではないことを明らかにした。例えば、個眼の形成、剛毛の形成および翼の形成は細胞相互作用に依存することもまた知られているが、これらは神経突然変異によって影響される(Morgan et al.,1925,Bibliogr.Genet.2:1-226; Welshons,1956,Dros.Inf.Serv.30:157-158;Preiss et al.,1988,EMBO J.7:3917-3927; Shellenbarger and Mohler,1978,Dev.Biol.62:432-446;Technau and Campos-Ortega,1986,Wilhelm Roux's Dev.Biol.195:445-454; Tomlisonand Ready,1987,Dev.Biol.120:366-376; Cagan and Ready,1989,Genes Dev.3:1099-1112)。神経遺伝子はまた、ハエの筋肉、消化管、外分泌系および再生系の正常な発生に必要とされる(Muskavitch,1994,Dev.Biol.166:415-430)。
【0007】
ノッチおよびデルタはいずれも膜を1回だけ貫く膜貫通タンパク質であり(Wharton et al.,1985,Cell 43:567-581; Kidd and Young,1986,Mol.Cell.Biol.
Genes Dev.2:1723-1735)、その細胞外ドメインに複数のタンデムEGF様反復配列を含む(Muskavitch,1994,Dev.Biol.166:415-430)。Notch遺伝子は、大きなN-末端細胞外ドメインを有する〜300kdタンパク質(我々は「Notch」をこのタンパク質を表すために使用する)をコードする。このタンパク質は、36個の表皮増殖因子(EGF)様タンデム反復配列とそれに続く3個の別のシステインを多く含む反復配列とを含み、これをNotch/lin-12反復配列と命名した(Whartonet al.,1985,Cell 43:567-581; Kidd and Young,1986,Mol.Cell.Biol.6:3094-3108;Yochem et al.,1988,Nature 335:547-550)。分子の研究により、ノッチとデルタとは、初期発生の決定に関与する、細胞のコミュニケーション機構の生化学的に相互作用する要素を構成するという示唆が導かれた(Fehonet al.,1990,Cell 61:523-534)。相同体が線虫(Caenorhabditis elegans)で発見され、ここでもNotch関連遺伝子lin-12とDelta関連遺伝子lag-2とは側面阻害に応答可能であった(Sternberg,1993,CurrentBiol.3:763-765; Henderson et al.,1994,Development 120:2913-2924; Greenwald,1994,Curr.Opin.Genet.Dev.4:556-562)。脊椎動物においては幾つかのノッチ相同体も同定されており(Kopan& Weintraub,1993,J.Cell Biol.121:631-641; Lardelli et al.,1994,Mech.Dev.46:123-136;Lardelli & Lendahl,1993,Exp.Cell Res.204:364-372; Weinmaster et al.,1991,Development113:199-205; Weinmaster et al.,1992,Development 116:931-941; Coffman et al.,1990,Science249:1438-1441; Bierkamp & Campos-Ortega,1993,Mech.Dev.43:87-100)、それらは多くの組織と発生の多くの段階で発現される。Notch-1の欠失は、マウスでは中胚葉節の欠損と胎児の死亡を生じさせたが(Swiateket al.,1994,Genes Dev.8:707-719; Conlon et al.,Rossant,J.Development(J.Dev.)121:1533-1545)、構造的なNotch-1の突然変異活性形は、アフリカツメガエル(Xenopus)および培養哺乳類細胞では細胞の分化を阻害するらしい(Coffmanet al.,1993,Cell 73:659-671; Kopan et al.,1994,Development 120:2385-2396; Nyeet al.,1994,Development 120:2421-2430)。
【0008】
EGF様モチーフが、血液凝固カスケードに関連するタンパク質を含めて、種々のタンパク質中で発見された(Furieand Furie,1988,Cell 53:505-518)。特に、このモチーフは血液凝固第IX因子および第X因子などの細胞外タンパク質(Rees etal.,1988,EMBO J.7:2053-2061; Furie and Furie,1988,Cell 53:505-518)、他のショウジョウバエの遺伝子(Knustet al.,1987,EMBO J.761-766; Rothnberg et al.,1988,Cell 55:1047-1059)、およびトロンボモジュリン(Suzukiet al.,1987,EMBOJ.6:1891-1897)およびLDL受容体(Sudhof et al.,1985,Science 228:815-822)などの幾つかの細胞表面受容体タンパク質において見出された。タンパク結合部位を、トロンボモジュリンおよびウロキナーゼ中のEGF反復配列ドメインにマッピングした(Kurosawaet al.,1988,J.Biol.Chem.263:5993-5996; Appella et al.,1987,J.Biol.Chem.262:4437-4440)。
【0009】
上記の参考文献は、これらが本発明に対する先行技術であるということを認めるものと解釈すべきではない。
【発明の概要】
【課題を解決するための手段】
【0010】
3.発明の概要
本発明は、脊椎動物のDelta遺伝子(ニワトリとマウスのDelta遺伝子、および他の種の
関連遺伝子)のヌクレオチド配列、およびそれらがコードするタンパク質のアミノ酸配列
、並びにその誘導体(例えば、断片)と類似体に関する。前述のヌクレオチド配列にハイ
ブリダイズ可能な核酸またはこれと相補的な核酸もまた提供される。特別な実施態様にお
いては、デルタタンパク質は哺乳類のタンパク質であり、好ましくはヒトのタンパク質で
ある。
【0011】
本発明は、脊椎動物のDelta誘導体および類似体に関し、これらは機能的に活性、すな
わち、全長(野生型の)デルタタンパク質に関連する1以上の公知の機能活性を表示する
ことができる。このような機能活性には、抗原性(抗デルタ抗体に結合する(または結合
についてデルタと拮抗する)能力)、免疫原性(デルタに結合する抗体を産生する能力)
、Notchもしくは他のトポリズミック(toporythmic)タンパク質またはそれらの断片への結
合能(または結合についてデルタと拮抗する能力)(「付着性」)、デルタの受容体への
結合能(または結合についてデルタと拮抗する能力)などが含まれるが、これらに限定さ
れるものではない。ここで使用する「トポリズミックタンパク質」は、Notch遺伝子、Del
ta遺伝子、Serrate遺伝子、Enhancerof split遺伝子、およびDeltex、並びにこの相互作
用する遺伝子のセットの他のメンバーのタンパク質産物をいい、他のメンバーは、それら
の遺伝子配列が、例えば、Delta遺伝子、Serrate遺伝子もしくはNotch遺伝子にハイブリ
ダイズする能力、またはそれらとの相同性、または表現型の相互作用を表示する能力もし
くは生化学的に相互作用するそれらのタンパク質産物の能力によって同定されてもよい。
【0012】
本発明はさらに、脊椎動物のDelta遺伝子の断片(およびその誘導体ならびに類似体)
に関し、これらは、細胞内ドメイン、細胞外ドメイン、膜貫通ドメイン、DSLドメイン、D
SLドメインに対するドメインのアミノ末端を含む1以上のデルタタンパク質のドメイン、
または1以上のデルタタンパク質のEGF様(相同)反復配列、または前述のものの組合わ
せを含むが、これらに限定されるものではない。
【0013】
脊椎動物のデルタタンパク質に対する抗体、その誘導体および類似体が、さらに提供さ
れる。
【0014】
脊椎動物のデルタタンパク質、誘導体および類似体の産生方法、例えば、組換え手段に
よる産生方法もまた提供される。
【0015】
本発明はまた、治療方法、検査方法、およびデルタタンパク質と核酸をベースとする組
成物に関する。本発明は、本発明の組成物を投与することによる細胞の予定運命または分
化の障害の治療を提供する。このような治療用組成物(本明細書中では「治療剤(Therape
utics)」という)は、デルタタンパク質とその類似体および誘導体(断片を含む);それ
らに対する抗体;デルタタンパク質、類似体、もしくは誘導体をコードする核酸;並びに
デルタアンチセンス核酸を含む。好適な実施態様においては、本発明の治療剤を投与して
ガン状態を治療し、または前新生物状態または非腫瘍状態から新生物発生または腫瘍状態
への進行を妨げる。他の特別の実施態様においては、本発明の治療剤を投与して神経系の
障害を治療し、または組織の再生および回復を促進させる。
【0016】
ある実施態様においては、ノッチおよび/またはデルタの機能と拮抗するかまたは阻害
する治療剤(以下「拮抗治療剤」という)を治療効果のために投与する。別の実施態様に
おいては、ノッチおよび/またはデルタの機能を促進させる治療剤(以下「アゴニスト治
療剤」という)を治療効果のために投与する。
【0017】
細胞の予定運命の障害、特に、ノッチおよび/またはデルタタンパク質の異常型もしく
は望ましくないレベルの発現もしくは活性または局在化を含む、過剰増殖障害(例えば、
ガン)または低増殖障害は、このようなレベルの検出によって診断することができ、以下
にさらに詳細に説明する。
【0018】
好適な特徴としては、本発明の治療剤は、少なくとも、ノッチタンパク質またはその断
片への結合を仲介するデルタの断片(以下「付着性断片」という)からなるタンパク質で
ある。
3.1.定義
本明細書中で使用するように、下線をつけてあるかまたは斜体にしてある遺伝子名は遺
伝子を示し、対照的に、それがコードするタンパク質産物は遺伝子名に下線を付していな
い。例えば、「Delta」はDelta遺伝子を意味し、「Delta」はDelta遺伝子のタンパク質産
物を表す。
【図面の簡単な説明】
【0019】
【図1A1】図1A1は、ニワトリのデルタ(C-Delta-1)のDNA配列である(配列番号1)。
【図1A2】図1A2は、ニワトリのデルタ(C-Delta-1)のDNA配列である(配列番号1)。
【図1A3】図1A3は、ニワトリのデルタ(C-Delta-1)のDNA配列である(配列番号1)。
【図1B1】図1B1は、別にスプライスされたニワトリのデルタ(C-Delta-1)のDNA配列である(配列番号3)。
【図1B2】図1B2は、別にスプライスされたニワトリのデルタ(C-Delta-1)のDNA配列である(配列番号3)。
【図2】図2は、ニワトリのデルタ(C-Delta-1)の推定アミノ酸配列である(配列番号2)。
【図3A】図3Aは、X-Delta-1(アフリカツメガエルのDelta;配列番号5)およびショウジョウバエのDelta(配列番号6)とを並べて、保存ドメイン構造、EGF反復配列、DSLドメイン、および膜貫通ドメイン(TM)を示した、C-Delta-1の推定アミノ酸配列である(配列番号2)。保存されたアミノ酸配列は四角で囲み、3つともが揃っておりかつN-末端のシステイン残基が揃っていない部分に、それぞれ●をつけた。C-Delta-1の細胞内ドメインとX-Delta-1の細胞内ドメインとは互いに非常によく似ているが、ショウジョウバエのDeltaの対応する部分に有意な相同性を示さなかった。
【図3B】図3Bは、X-Delta-1(アフリカツメガエルのDelta;配列番号5)およびショウジョウバエのDelta(配列番号6)とを並べて、保存ドメイン構造、EGF反復配列、DSLドメイン、および膜貫通ドメイン(TM)を示した、C-Delta-1の推定アミノ酸配列である(配列番号2)。保存されたアミノ酸配列は四角で囲み、3つともが揃っておりかつN-末端のシステイン残基が揃っていない部分に、それぞれ●をつけた。C-Delta-1の細胞内ドメインとX-Delta-1の細胞内ドメインとは互いに非常によく似ているが、ショウジョウバエのDeltaの対応する部分に有意な相同性を示さなかった。
【図4】図4は、C-Delta-1(配列番号2)、ショウジョウバエのDelta(配列番号6)(Vassinet al.,1987,EMBO J.6:3431-3440; Kopczynski et al.,1988,Genes Dev.2:1723-1735)、ショウジョウバエのSerrate(配列番号7)(Fleminget al.,1990,Genes Dev.4:2188-2201;Thomas et al.,1991,Development 111:749-761)、C-Serrate-1(配列番号8)(Myat,Henrique,Ish-Horowiczand Lewis,準備中)、Apx-1(配列番号9)(Melloet al.,1994,Cell 77:95-106)およびLag-2(配列番号10)(Henderson et al.,1994,Development120:2913-2924; Tax et al.,1994 Nature 368:150-154)からのDSLドメインのアライメントを、保存されたシステインのスペースをあけ、ショウジョウバエと脊椎動物のノッチ様タンパク質のために仮定されたリガンドの間で保存されたアミノ酸、およびC.elegans(線虫)のリガンドでさらに保存されたこれらとを示す(四角で囲んだ)。
【図5A】C-Delta-1の発現とC-Notch-1の発現とは、第1日(El)の神経板における神経発生の開始と相関する。左側が前である。全体をマウントしたin situハイブリダイゼーションの検体を図5A−Dに示す。5Eは断面図である。図5Aは、ステージ7において、神経板の大部分と前中胚葉節の中胚葉の根底にある部分でC-Notch-1が発現された。
【図5B】図5Bは、ステージ7におけるC-Delta-1が神経板、将来の後菱脳、第一の中胚葉節のすぐ前方ですでに検出可能である(白い四角形)。この神経ドメインの後部端は、根底にある前中胚葉節の中胚葉(psm)において、非常に強く発現されたドメインの前部の縁とほぼ水平である。神経板中における初期の発現が生じるかもしれず、根底にある中胚葉中での発現は遮蔽されるかもしれない(未発表の結果)。
【図5C】図5Cは、5B中で四角で囲った場所の拡大図であり、C-Delta-1を発現している神経板中で光散乱された細胞(scattered cells)を示す。
【図5D】図5Dでは、ステージ8において、神経板中におけるC-Delta-1の発現が神経板の発生として後に伸びている。この写真において見ることができる標識された神経板細胞のドメイン(カッコをつけた部分)は、前中胚葉節の中胚葉を超えて後ろに続いている。
【図5E】図5Eは、ステージ8における胚の準矢じり形の断面図であり、神経板(組織の背層;カッコをつけた)の光散乱細胞中でC-Delta-1が発現されており、中胚葉節の中胚葉(腹層)中で幅が広くなっている。断面の平面はわずかに斜めになっており、神経板ドメインの後部が失われている(5D参照)。
【図6A】図6A〜6Cは、C-Delta-1発現細胞がBrdUを取込んでいないことを示す。612個のC-Delta-1細胞のうち、581個がBrdU-(76個の断面;6個の胚)であった。図6Aは、細胞周期中のどの時期が神経上皮の細胞の核の頂部−基底部の位置に関連するかを示す線図である。S期の核は基底にある(Fujita,1963,J.Comp.Neurol.120:37-42; Biffo et al.,1992,Histochem.Cytochem.40:535-540)。核は影をつけて示した。
【図6B】図6A〜6Cは、C-Delta-1発現細胞がBrdUを取込んでいないことを示す。図6Bは、BrdUで2時間標識したステージ9の胚の神経管の断面図であり、C-Delta-1発現細胞(青色の地色上で暗く見える)とBrdU標識核(ピンク)を示す。標識核はDNA合成が生じる基底で優性であり、基底のC-Delta-1発現細胞は未だ標識されていない。
【図6C】図6A〜6Cは、C-Delta-1発現細胞がBrdUを取込んでいないことを示す。図6Cは4時間インキュベートしたステージ9の胚の断面図である。多くの標識核がS期を脱しており、管腔の方へ移動しているが、C-Delta-1発現細胞はまだ基底にあり、BrdUで標識されていない。
【図7A】図7Aは、マウスのDelta遺伝子(M-Delta-1)のDNA配列(配列番号11)である。
【図7B】図7Bは、マウスのDelta遺伝子(M-Delta-1)のDNA配列(配列番号11)である。
【図8】図8は、マウスのDelta遺伝子(M-Delta-1)の推定アミノ酸配列(配列番号12)である。
【図9A】図9Aは、マウスのM-Delta-1(配列番号12)とニワトリのC-Delta-1(配列番号2)の推定アミノ酸配列のアライメントを示す。これらは、それらの多くのアミノ酸配列が同一であることを示す。同一のアミノ酸は四角で囲った。2つの遺伝子の間のコンセンサス配列(配列番号13)は下の方に示した。
【図9B】図9Bは、マウスのM-Delta-1(配列番号12)とニワトリのC-Delta-1(配列番号2)の推定アミノ酸配列のアライメントを示す。これらは、それらの多くのアミノ酸配列が同一であることを示す。同一のアミノ酸は四角で囲った。2つの遺伝子の間のコンセンサス配列(配列番号13)は下の方に示した。
【図10A】ヒトDelta(H-Delta-1)のPCR増幅断片のDNA配列(配列番号14)および3つの可能なオープンリーディングフレームを用いる推定アミノ酸配列、第2行目(配列番号15)、第3行目(配列番号16)、第4行目(配列番号17)。
【図10B】ヒトDelta(H-Delta-1)のPCR増幅断片のDNA配列(配列番号14)および3つの可能なオープンリーディングフレームを用いる推定アミノ酸配列、第2行目(配列番号15)、第3行目(配列番号16)、第4行目(配列番号17)。
【図11】ヒトH-Delta-1(上段)とニワトリC-Delta-1(下段)のアライメント。ヒトデルタの推定アミノ酸配列(配列番号18)を上段に示す。ヒトデルタの配列を「肉眼」で判定し、その場合、C-Delta-1との相同性を最大とすることにより適当なリーディングフレームの配列を決定した。図10に示した単一のリーディングフレームはどれも、リーディングフレームシフトを引き起こした図10のDNA配列のエラーのため、正しい配列を与えなかった。
【図12A1】図12A1はクローンHDl 18からのヒトDelta(H-Delta-1)のコンティグ(contig)DNA配列を表す。
【図12A2】図12A2はクローンHDl 18からのヒトDelta(H-Delta-1)のコンティグ(contig)DNA配列を表す。
【図12A3】図12A3はクローンHDl 18からのヒトDelta(H-Delta-1)のコンティグ(contig)DNA配列を表す。
【図12B1】図12B1は図12Aに示したヌクレオチド配列(上段、配列番号33)および3つの可能なオープンリーディングフレームを用いる推定アミノ酸配列、第2行目(配列番号34)、第3行目(配列番号35)、第4行目(配列番号36)を表す。マウスデルタ−1のアミノ酸配列と最大の相同性を有するアミノ酸配列をボックスで囲ってある。このボックスで囲ったアミノ酸配列はヒトデルタの推定アミノ酸配列である。ここで、リーディングフレームシフトは、この配列中の配列決定エラーが存在する場所を示す。図12Aに示した単一のリーディングフレームはどれも、リーディングフレームのシフトを引き起こしたDNA配列中のエラーのため、中断されないアミノ酸配列を与えなかった。Xは未決定のアミノ酸を示し、Nは未決定のヌクレオチドを示す。
【図12B2】図12B2は図12Aに示したヌクレオチド配列(上段、配列番号33)および3つの可能なオープンリーディングフレームを用いる推定アミノ酸配列、第2行目(配列番号34)、第3行目(配列番号35)、第4行目(配列番号36)を表す。マウスデルタ−1のアミノ酸配列と最大の相同性を有するアミノ酸配列をボックスで囲ってある。このボックスで囲ったアミノ酸配列はヒトデルタの推定アミノ酸配列である。ここで、リーディングフレームシフトは、この配列中の配列決定エラーが存在する場所を示す。図12Aに示した単一のリーディングフレームはどれも、リーディングフレームのシフトを引き起こしたDNA配列中のエラーのため、中断されないアミノ酸配列を与えなかった。Xは未決定のアミノ酸を示し、Nは未決定のヌクレオチドを示す。
【図12B3】図12B3は図12Aに示したヌクレオチド配列(上段、配列番号33)および3つの可能なオープンリーディングフレームを用いる推定アミノ酸配列、第2行目(配列番号34)、第3行目(配列番号35)、第4行目(配列番号36)を表す。マウスデルタ−1のアミノ酸配列と最大の相同性を有するアミノ酸配列をボックスで囲ってある。このボックスで囲ったアミノ酸配列はヒトデルタの推定アミノ酸配列である。ここで、リーディングフレームシフトは、この配列中の配列決定エラーが存在する場所を示す。図12Aに示した単一のリーディングフレームはどれも、リーディングフレームのシフトを引き起こしたDNA配列中のエラーのため、中断されないアミノ酸配列を与えなかった。Xは未決定のアミノ酸を示し、Nは未決定のヌクレオチドを示す。
【図12B4】図12B4は図12Aに示したヌクレオチド配列(上段、配列番号33)および3つの可能なオープンリーディングフレームを用いる推定アミノ酸配列、第2行目(配列番号34)、第3行目(配列番号35)、第4行目(配列番号36)を表す。マウスデルタ−1のアミノ酸配列と最大の相同性を有するアミノ酸配列をボックスで囲ってある。このボックスで囲ったアミノ酸配列はヒトデルタの推定アミノ酸配列である。ここで、リーディングフレームシフトは、この配列中の配列決定エラーが存在する場所を示す。図12Aに示した単一のリーディングフレームはどれも、リーディングフレームのシフトを引き起こしたDNA配列中のエラーのため、中断されないアミノ酸配列を与えなかった。Xは未決定のアミノ酸を示し、Nは未決定のヌクレオチドを示す。
【図12B5】図12B5は図12Aに示したヌクレオチド配列(上段、配列番号33)および3つの可能なオープンリーディングフレームを用いる推定アミノ酸配列、第2行目(配列番号34)、第3行目(配列番号35)、第4行目(配列番号36)を表す。マウスデルタ−1のアミノ酸配列と最大の相同性を有するアミノ酸配列をボックスで囲ってある。このボックスで囲ったアミノ酸配列はヒトデルタの推定アミノ酸配列である。ここで、リーディングフレームシフトは、この配列中の配列決定エラーが存在する場所を示す。図12Aに示した単一のリーディングフレームはどれも、リーディングフレームのシフトを引き起こしたDNA配列中のエラーのため、中断されないアミノ酸配列を与えなかった。Xは未決定のアミノ酸を示し、Nは未決定のヌクレオチドを示す。
【図12B6】図12B6は図12Aに示したヌクレオチド配列(上段、配列番号33)および3つの可能なオープンリーディングフレームを用いる推定アミノ酸配列、第2行目(配列番号34)、第3行目(配列番号35)、第4行目(配列番号36)を表す。マウスデルタ−1のアミノ酸配列と最大の相同性を有するアミノ酸配列をボックスで囲ってある。このボックスで囲ったアミノ酸配列はヒトデルタの推定アミノ酸配列である。ここで、リーディングフレームシフトは、この配列中の配列決定エラーが存在する場所を示す。図12Aに示した単一のリーディングフレームはどれも、リーディングフレームのシフトを引き起こしたDNA配列中のエラーのため、中断されないアミノ酸配列を与えなかった。Xは未決定のアミノ酸を示し、Nは未決定のヌクレオチドを示す。
【図13A】マウスM-Delta-1DNA配列(第1行目、配列番号37)とヒトH-Delta-1DNA配列(第2行目、配列番号33)とそれらの共通配列(第3行目、配列番号38)のアライメント。
【図13B】マウスM-Delta-1DNA配列(第1行目、配列番号37)とヒトH-Delta-1DNA配列(第2行目、配列番号33)とそれらの共通配列(第3行目、配列番号38)のアライメント。
【図13C】マウスM-Delta-1DNA配列(第1行目、配列番号37)とヒトH-Delta-1DNA配列(第2行目、配列番号33)とそれらの共通配列(第3行目、配列番号38)のアライメント。
【図13D】マウスM-Delta-1DNA配列(第1行目、配列番号37)とヒトH-Delta-1DNA配列(第2行目、配列番号33)とそれらの共通配列(第3行目、配列番号38)のアライメント。
【図13E】マウスM-Delta-1DNA配列(第1行目、配列番号37)とヒトH-Delta-1DNA配列(第2行目、配列番号33)とそれらの共通配列(第3行目、配列番号38)のアライメント。
【図13F】マウスM-Delta-1DNA配列(第1行目、配列番号37)とヒトH-Delta-1DNA配列(第2行目、配列番号33)とそれらの共通配列(第3行目、配列番号38)のアライメント。
【図13G】マウスM-Delta-1DNA配列(第1行目、配列番号37)とヒトH-Delta-1DNA配列(第2行目、配列番号33)とそれらの共通配列(第3行目、配列番号38)のアライメント。
【図14A】図12Bからのボックスで囲ったアミノ酸配列を表す、コンポサイト・ヒトデルタ(H-Delta-1)アミノ酸配列(それぞれ配列番号39〜65)を示す。「>」は配列が下の行に続くことを示す。「*」は配列の中断を示す。
【図14B】図12Bからのボックスで囲ったアミノ酸配列を表す、コンポサイト・ヒトデルタ(H-Delta-1)アミノ酸配列(それぞれ配列番号39〜65)を示す。「>」は配列が下の行に続くことを示す。「*」は配列の中断を示す。
【発明を実施するための形態】
【0020】
5.発明の詳細な説明
本発明は、脊椎動物Delta遺伝子のヌクレオチド配列およびそれらのコード化タンパク
質のアミノ酸配列に関する。本発明はさらに、脊椎動物デルタタンパク質の断片、並びに
他の誘導体および類似体に関する。このような断片または誘導体をコードする核酸も本発
明の範囲内である。本発明は多くの異なる脊椎動物種のDelta遺伝子とそれらのコード化
タンパク質を提供する。本発明のDelta遺伝子にはニワトリ、マウス、ヒトのDelta、およ
び他の脊椎動物種の関連遺伝子(相同体)が含まれる。特定の実施態様において、Delta
遺伝子およびタンパク質は脊椎動物、特に哺乳動物に由来するものである。本発明の好ま
しい実施態様では、デルタタンパク質がヒトタンパク質である。前記タンパク質および誘
導体の、例えば組換え法による、産生も提供される。
【0021】
本発明は、機能的に活性な本発明のデルタ誘導体および類似体に関し、すなわち、それ
らは全長(野生型)タンパク質と関連した既知の機能的活性を1以上示すことができるも
のである。このような機能的活性として、抗原性〔抗デルタ抗体と結合する(または該結
合に関してデルタと競合する)能力〕、免疫原性〔デルタと結合する抗体を誘導する能力
〕、ノッチ(Notch)または他のトポリズミック(toporythmic)タンパク質もしくはその断片
と結合する(または該結合に関してデルタと競合する)能力(「接着性」)、デルタの受
容体と結合する(または該結合に関してデルタと競合する)能力、細胞の運命または分化
に影響を与える能力、および治療活性が挙げられるが、これらに限らない。本明細書中で
用いる「トポリズミックタンパク質」 とは、Notch、Delta、Serrate、Enhancer of spli
t、およびDeltex、並びにこの相互作用遺伝子ファミリーの他のメンバー(例えば、Delta
、SerrateまたはNotchとハイブリダイズするその遺伝子配列の能力、またはDelta、Serr
ateまたはNotchに対するその相同性、または表現型相互作用を示すその遺伝子の能力によ
って同定しうる)のタンパク質産物を指す。
【0022】
本発明はさらに、デルタタンパク質の細胞内ドメイン、細胞外ドメイン、DSLドメイ
ン、DSLドメインのアミノ末端側の領域、膜貫通領域、膜結合領域、もしくは1以上の
EGF様(相同)反復ドメイン、またはこれらの任意の組合せを含むがこれらに限らない
、デルタタンパク質の1以上のドメインを含むデルタの断片(並びにその誘導体および類
似体)に関する。
【0023】
さらに、脊椎動物デルタ、その誘導体および類似体に対する抗体も提供される。
【0024】
以下で実証するように、デルタは細胞の運命(分化)の決定に関与する発生および他の
生理学的プロセスにおいて、特にノッチのリガンドとして、重要な役割を果たしている。
とりわけ、デルタは中枢神経系の細胞の運命を決定づける上で主要な役割を担っていると
考えられる。本発明の核酸およびアミノ酸配列並びに抗体は、ヒトと他の生物種のデルタ
mRNAおよびタンパク質の検出・定量に、その発現の研究に、デルタとその断片、他の
誘導体および類似体の産生に、分化および他の生理学的プロセスの研究・操作に用いるこ
とができる。本発明はまた、デルタタンパク質および核酸に基づく治療および診断法、並
びに医薬組成物に関する。本発明は、本発明の治療用化合物を投与することによる細胞運
命または分化の障害の治療を提供する。このような治療用化合物(本明細書中では「治療
薬」という)としては、デルタタンパク質、その類似体および誘導体(断片を含む)、そ
れらに対する抗体、デルタタンパク質、類似体または誘導体をコードする核酸、およびデ
ルタ・アンチセンス核酸が挙げられる。好ましい実施態様では、本発明の治療薬は癌状態
を治療するために、あるいは前新生物または非悪性状態から新生物または悪性状態への進
行を防ぐために投与される。他の特定の実施態様では、本発明の治療薬は神経系の障害を
治療するために、または組織の再生および修復を促進するために投与される。
【0025】
一実施態様において、ノッチおよび/またはデルタ機能と拮抗するまたはそれらを阻害
する治療薬(以後「アンタゴニスト治療薬」という)が治療効果のために投与される。他
の実施態様において、ノッチおよび/またはデルタ機能を促進する治療薬(以後「アゴニ
スト治療薬」という)が治療効果のために投与される。
【0026】
細胞運命の疾患、特にノッチおよび/またはデルタタンパク質の発現、活性または局在
化の異常なもしくは望ましくないレベルを伴う過増殖性(例えば、癌)または低増殖性疾
患は、以下で詳述するように、このようなレベルを検出することにより診断することがで
きる。
【0027】
好ましい態様において、本発明の治療薬は、ノッチタンパク質またはその断片への結合
を媒介するデルタの少なくとも断片(本明細書中では「接着性断片」という)からなるタ
ンパク質である。
【0028】
本発明は、特に、ニワトリDelta相同体のクローニング(第6節)、マウスDelta相同体
のクローニング(第7節)、およびヒトDelta相同体のクローニング(第8節)を開示す
る以下の実施例により例示される。
【0029】
制限ではなく、開示を明瞭にするために、本発明の詳細な説明を以下の分節に大別する
。
【0030】
5.1.Delta遺伝子の単離
本発明は、脊椎動物のDelta核酸のヌクレオチド配列に関する。特定の態様では、ヒト
のDelta核酸は、図10(配列番号14)もしくは図12A(配列番号33)に示すcD
NA配列、またはそのコード領域、または脊椎動物のデルタ(Delta)タンパク質をコード
する核酸(例えば、配列番号1、3、11、14または33の配列を有する)を含む。本
発明は、脊椎動物のDelta配列の少なくとも8個のヌクレオチド(すなわち、ハイブリダ
イズ可能な部分)から成る核酸を提供し、他の態様では、核酸が、Delta配列の少なくと
も25個の(連続)ヌクレオチド、50個のヌクレオチド、100個のヌクレオチド、150個のヌ
クレオチド、もしくは200個のヌクレオチド、またはDeltaの全長をコードする配列から成
る。本発明はまた、前記配列もしくはその相補体にハイブリダイズ可能な、または相補的
な核酸にも関する。特定の発明では、脊椎動物のDelta遺伝子の少なくとも10、25、50、1
00もしくは200ヌクレオチド、または全体をコードする領域に相補的な配列を含む核酸が
提供される。特定の態様では、ストリンジェンシーの低い条件下で、脊椎動物(例えば、
哺乳類)のDelta核酸(例えば、配列番号14もしくは配列番号33またはそれらの少な
くとも10、25、50、100もしくは200個のヌクレオチド部分を有する)、またはデルタ誘導
体をコードする核酸にハイブリダイズ可能な核酸が提供される。例としては、それに限定
されないが、そのようなストリンジェンシーの低い条件を使用する手法は、次の通りであ
る(ShiloおよびWeinberg,1981,Proc.Natl.Acad.Sci.USA78:6789-6792も参照)。
すなわち、DNAを含むフィルターを、40℃で6時間、35%のホルムアミド、5X SSC、50m
Mのトリス−HCl(pH7.5)、5mMのEDTA、0.1%のPVP、0.1%のフィコール、1%
のBSAおよび500μg/mlの変性サケ精子DNAを含む溶液で前処理する。ハイブリダイ
ゼーションを同じ溶液で行うが、次の変更を伴う。すなわち、0.02%のPVP、0.02%のフ
ィコール、0.2%のBSA、100μg/mlのサケ精子DNA、10%(wt/vol)の硫酸デキストラ
ンおよび5〜20x 106cpmの32P−標識プローブを使用する。フィルターを40℃で18〜20時
間、ハイブリダイゼーション混合物中でインキュベートした後、55℃で1.5時間、2X SSC
、25mMのトリス−HCl(pH7.4)、5mMのEDTAおよび0.1%のSDSを含む溶液で洗
浄する。洗浄溶液を新しい溶液と入れ換え、60℃でさらに1.5時間インキュベートする。
フィルターを吸い取って乾かし、オートラジオグラフィー用に露光する。必要であれば、
フィルターを65〜68℃で3回目の洗浄にかけ、フィルムに再露光する。使用できるストリ
ンジェンシーの低い他の条件は、当技術分野で周知である(例えば、種の交差したハイブ
リダイゼーションに対して使用されるもの)。
【0031】
別の特定の態様では、ストリンジェンシーの高い条件下で脊椎動物(例えば、哺乳類)
のDelta核酸にハイブリダイズ可能な核酸が提供される。例えば、それに限定されないが
、そのようなストリンジェンシーの高い条件を使用する手法は次の通りである。すなわち
、DNAを含むフィルターの前ハイブリダイゼーションを、65℃で8時間〜一夜、6X SSC
、50mMのトリス−HCl(pH7.5)、1mMのEDTA、0.02%のPVP、0.02%のフィコー
ル、0.02%のBSAおよび500μg/mlの変性サケ精子DNAで構成される緩衝液中で行う。
フィルターを65℃で48時間、100μg/mlの変性サケ精子DNAおよび5〜20x 106cpmの32
P−標識したプローブを含む前ハイブリダイゼーション混合物中でハイブリダイズする。
フィルターの洗浄を、37℃で1時間、2X SSC、0.01%のPVP、0.01%のフィコールおよび
0.01%のBSAを含む溶液中で行う。この後、50℃で45分間、0.1X SSCで洗浄して、オー
トラジオグラフィーにかける。使用できるストリンジェンシーの高い他の条件は、当技術
分野で周知である。
【0032】
脊椎動物のDeltaタンパク質の断片および誘導体をコードする核酸(第5.6節参照)
ならびにDeltaアンチセンス核酸(第5.11節参照)がさらに提供される。すでに明ら
かなように、本明細書で使用する「デルタタンパク質の断片または一部をコードする核酸
」は、デルタタンパク質の列挙した断片または一部のみをコードする核酸を意味するもの
とし、デルタタンパク質の連続配列としての他の隣接した部分は意味しない。
【0033】
他のトポリズミック(toporythmic)タンパク質と相同性のある領域を含む脊椎動物のDel
ta核酸の断片も提供される。他の種のデルタタンパク質のDSL領域(DrosophilaSerra
teおよびDeltaと相同性のある領域)も提供される。図3および8に示すようなDeltaとSe
rrateとの間の保存領域をコードする核酸も提供される。
【0034】
脊椎動物のDelta遺伝子のクローニングに対する特定の態様を特定の例として以下に示
すが、それに限定されるものではない。
【0035】
発現クローニング(一般に当技術分野で公知の技術)に対しては、当技術分野で公知の
方法により、発現ライブラリーを作製する。例えば、mRNA(例えば、ヒト)を単離し
、cDNAを作製して発現ベクター(例えば、バクテリオファージ誘導体)に連結し、後
にその中に導入される宿主細胞によって発現できるようにする。次いで、種々のスクリー
ニングアッセイを使用して、発現したデルタ産物を選択することができる。一つの態様で
は、選択のために抗−デルタ抗体を使用することができる。
【0036】
別の好ましい発明では、選択の前に、PCRを使用してゲノムまたはcDNAライブラ
リーの所望の配列を増幅する。公知Delta配列(好ましくは、脊椎動物の配列)を表すオ
リゴヌクレオチドプライマーがPCRのプライマーとして使用できる。好ましい発明では
、オリゴヌクレオチドプライマーが、SerrateとDeltaとの間の相同性が強いDelta保存セ
グメントの少なくとも一部を表す。合成オリゴヌクレオチドをプライマーとして使用して
、潜在的に興味の対象となる源(RNAまたはDNA)、好ましくはcDNAライブラリ
ー由来の配列をPCRにより増幅してもよい。PCRは、例えば、Perkin-Elmer Cetusサ
ーマルサイクラーおよびTaqポリメラーゼ(Gene Amp)を使用して行うことができる。増
幅されるDNAとしては、真核細胞種由来のmRNAまたはcDNAまたはゲノムDNA
が挙げられる。PCR反応で使用するために、いくつかの異なる縮重プライマーの合成を
選択することができる。また、公知のDeltaヌクレオチド配列と単離される核酸相同体と
の間のヌクレオチド配列の類似性の多少を考慮するために、PCR反応のプライミングに
使用するハイブリダイゼーション条件のストリンジェンシーを変えることもできる。種の
交差したハイブリダイゼーションの場合は、ストリンジェンシーの低い条件が好ましい。
同じ種のハイブリダイゼーションの場合は、ストリンジェンシーが中位の条件が好ましい
。Delta相同体のセグメントの増幅の成功の後、そのセグメントは、分子的にクローン化
して配列決定し、完全なcDNAまたはゲノムクローンを単離するためのプローブとして
利用できる。これにより、次いで、以下に記載するように、遺伝子の完全なヌクレオチド
配列の決定、その発現の分析、および機能的分析のためのそのタンパク質産物の産生が可
能になる。このようにして、デルタタンパク質をコードするさらに別の遺伝子が同定でき
る。そのような手法を、以下の種々の実施例の節で例として示す。
【0037】
上記方法は、Deltaのクローンを得ることができる方法の下記の一般的記載を限定する
ものではない。
【0038】
どの脊椎動物細胞も、潜在的に、Delta遺伝子の分子クローニングに対する核酸源とし
て役立つと考えられる。Deltaをコードする核酸配列は、哺乳類、ヒト、ブタ、ウシ、ネ
コ、トリ、ウマ、イヌおよびそれ以外の霊長類源から単離できる。例えば、Deltaプライ
マーとともにcDNAライブラリーを使用したPCRによって、マウス、ニワトリおよび
ヒトのDelta遺伝子の断片を増幅した。DNAは、クローン化したDNA(例えばDNA
「ライブラリー」)から当技術分野で公知の標準的手法により、化学合成により、cDN
Aクローニングにより、または所望の細胞から精製したゲノムDNAもしくはその断片の
クローニングにより、得ることができる(例えば、Sambrookら、1989,Molecular Clonin
g,A laboratoryManual,第2版、Cold Spring Harbor Laboratory Press,Cold Sprig
Harbor,NewYork; Glover,D.M.(編)、1995,DNA Cloning: A Practical Approach,
MRL Press,Itd.,Oxford,U.K.Vol.I,IIを参照)。ゲノムDNAから誘導されるクロ
ーンは、コード領域以外に、調節およびイントロンDNA領域を含み得る。cDNA由来
のクローンは、エキソン配列のみを含む。源が何であっても、遺伝子は、遺伝子の増殖に
適するベクターに入れて分子的にクローン化すべきである。
【0039】
ゲノムDNA由来の遺伝子の分子クローニングでは、DNA断片が生じ、そのいくつか
は、所望の遺伝子をコードする。DNAは、種々の制限酵素を使用して特定の部位で切断
できる。あるいは、マンガンの存在下でDNAseを使用してDNAを断片化することが
でき、または、DNAを例えば超音波処理などによって物理的に切断することができる。
次いで、直鎖状DNA断片は、それらに限定されないが、アガロースおよびポリアクリル
アミドゲル電気泳動ならびにカラムクロマトグラフィーなどの標準的技術によって、サイ
ズに従って分離することができる。
【0040】
DNA断片がいったん生じると、所望の遺伝子を含む特定のDNA断片の同定は、多く
の方法で行うことができる。例えば、(いずれかの種の)Delta遺伝子もしくはその特異
的RNAの一部またはそれらの断片、例えば細胞外ドメイン(第5.6節参照)のある量
が入手でき、精製・標識することができるならば、生じたDNA断片は、標識したプロー
ブへの核酸ハイブリダイゼーションによってスクリーニングできる(Benton,W.およびDa
vis,R.,1977,Science196:180; Grunstein,M.およびHogness,D.,1975,Proc.Natl
.Acad.Sci.U.S.A.72:3961)。プローブと実質的に相同であるDNA断片はハイブリダ
イズする。また、制限酵素による消化、および既知制限地図が利用できるならばそれに従
って予想されるものとの断片の大きさの比較によって、適切な断片を同定することもでき
る。遺伝子の性質に基づいて、さらに選択を行うことができる。あるいは、遺伝子の有無
は、その発現産物の物理的、化学的または免疫学的性質に基づくアッセイによって検出で
きる。例えば、cDNAクローン、または適切なmRNAをハイブリッド−選択するDN
Aクローンは、電気泳動の移動、等電点挙動、タンパク質分解消化地図、結合活性、in v
itro集合活性(「接着性」)または抗原性がDeltaに対して公知のものと同様または同一
であるものが選択され得る。Deltaに対する抗体が利用できるならば、デルタタンパク質
は、標識した抗体を推定上のDeltaを合成するクローンに結合させることにより、ELI
SA(固相酵素免疫検定法)型手法で同定できる。
【0041】
Delta遺伝子はまた、核酸ハイブリダイゼーションによるmRNA選択、続くinvitro
翻訳によって同定することもできる。この手法では、断片を使用して、相補的mRNAを
ハイブリダイゼーションにより単離する。そのようなDNA断片は、別の種(例えば、Dr
osophila)の利用可能な精製DeltaDNAを表す可能性がある。単離したmRNAの単離
産物のinvitro翻訳産物の免疫沈降分析または機能的分析(例えば、in vitroでの集合力
;受容体への結合;下記を参照)によりmRNA、従って、所望配列を含む相補的DNA
断片が同定される。さらに、特異的mRNAは、細胞から単離したポリソームの、デルタ
タンパク質に対して特異的な固定化抗体への吸着によって選択できる。放射線標識したDe
lta cDNAは、(吸着したポリソームから)選択したmRNAを鋳型として使用して合
成できる。次いで、放射線標識したmRNAまたはcDNAをプローブとして使用して、
他のゲノムDNA断片からDelta DNAを同定することができる。
【0042】
DeltaゲノムDNAを単離する別の方法としては、それらに限定されないが、既知配列
からの遺伝子配列自体の化学的合成またはデルタタンパク質をコードするmRNAに対す
るcDNAの作製が挙げられる。例えば、Delta遺伝子をcDNAクローニングするため
のRNAは、Deltaを発現する細胞から単離できる。他の方法も可能であり、本発明の範
囲内である。
【0043】
同定・単離した遺伝子は、次いで、適切なクローニングベクターに挿入することができ
る。当技術分野で公知の多数のベクター−宿主系が使用できる。可能なベクターとしては
、それらに限定されないが、プラスミドまたは変性ウイルスが挙げられる。しかし、ベク
ター系は、使用する宿主細胞と相溶性でなければならない。そのようなベクターとしては
、それらに限定されないが、λ誘導体などのバクテリオファージ、またはPBR322もしくは
pUCプラスミド誘導体などのプラスミドが挙げられる。クローニングベクターへの挿入は
、例えば、DNA断片を、相補的付着末端を有するクローニングベクターに連結すること
によって行うことができる。しかし、DNAの断片化に使用する相補的制限部位がクロー
ニングベクターに存在しないならば、DNA分子の両端を酵素的に改変することができる
。あるいは、所望するいずれかの部位を、ヌクレオチド配列(リンカー)をDNA末端に
連結することにより作り出すことができる。これらの連結したリンカーは、制限エンドヌ
クレアーゼ認識配列をコードする、化学的に合成した特定のオリゴヌクレオチドを含み得
る。別の方法では、切断したベクターおよびDelta遺伝子を、ホモポリマー末端付着によ
り改変することができる。組換え分子は、形質転換、トランスフェクション、感染、電気
穿孔法などによって宿主細胞に導入することができ、その結果、遺伝子配列の多くのコピ
ーが生じる。
【0044】
別の方法では、適切なクローニングベクターに挿入した後、所望の遺伝子を、「ショッ
トガン」法で同定・単離することができる。所望遺伝子の濃縮は、例えばサイズ分画によ
って、クローニングベクターへの挿入前に行うことができる。
【0045】
特定の態様では、単離したDelta遺伝子、cDNAまたは合成したDNA配列を挿入し
た組換えDNA分子で宿主細胞を形質転換することにより、多コピーの遺伝子を生じるこ
とができる。すなわち、遺伝子は、形質転換体を増殖し、その形質転換体から組換えDN
A分子を単離し、必要であれば、挿入した遺伝子を単離した組換えDNAから回収するこ
とにより、大量に得ることができる。
【0046】
本発明によって提供されるDelta配列としては、天然の脊椎動物デルタタンパク質に見
られるものと実質的に同じアミノ酸配列をコードするヌクレオチド配列および機能的に同
等のアミノ酸を有するコードされたアミノ酸配列が挙げられ、それらは全て、デルタ誘導
体に対する下記の第5.6節に記載されている。
【0047】
5.2.Delta遺伝子の発現
脊椎動物のデルタタンパク質をコードするヌクレオチド配列またはその機能的に活性な
断片もしくは他の誘導体(第5.6節参照)は、適切な発現ベクター、すなわち、挿入さ
れたタンパク質をコードする配列の転写および翻訳に必要な要素を含むベクターに挿入で
きる。また、必要な転写および翻訳シグナルは、天然のDelta遺伝子および/またはその
フランキング領域によって供給され得る。種々の宿主−ベクター系を使用して、タンパク
質をコードする配列を発現することができる。これらには、それらに限定されないが、ウ
イルス(例えば、ワクシニアウイルス、アデノウイルスなど)感染した哺乳類細胞系;ウ
イルス(例えば、バキュロウイルス)感染した昆虫細胞系;酵母ベクターを含む酵母また
はバクテリオファージ、DNA、プラスミドDNAもしくはコスミドDNAで形質転換し
た細菌などの微生物が含まれる。ベクターの発現要素は、その強さおよび特異性が変わる
。使用する宿主−ベクター系によって、多数の適する転写・翻訳要素のいずれか一つを使
用することができる。特定の態様では、Delta遺伝子の付着部分を発現させる。他の特定
の態様では、ヒトDelta遺伝子、またはヒトデルタの機能的に活性な部分をコードする配
列を発現させる。さらに別の態様では、細胞外ドメインを含むデルタの断片、または他の
誘導体、またはデルタの類似体を発現させる。
【0048】
DNA断片のベクターへの挿入に対して先に記載した方法のいずれかを使用して、適切
な転写/翻訳制御シグナルおよびタンパク質をコードする配列から成るキメラ遺伝子を含
む発現ベクターを構築することができる。これらの方法としては、in vitro組換えDNA
および合成法ならびにin vivo組換え法(遺伝子組換え)が挙げられる。デルタタンパク
質またはペプチド断片をコードする核酸配列の発現は、デルタタンパク質またはペプチド
が組換えDNA分子で形質転換された宿主で発現されるように、第二の核酸配列によって
調節することができる。例えば、デルタタンパク質の発現は、当技術分野で公知のプロモ
ーター/エンハンサー要素によって制御できる。Delta遺伝子発現の制御に使用できるプ
ロモーターとしては、それらに限定されないが、SV40初期プロモーター領域(Bernoist
およびChambon,1981,Nature290:304-310)、ラウス肉腫ウイルスの3’の長い末端反復
配列に含まれるプロモーター(Yamamotoら、1980,Cell 22:787-797)、ヘルペスチミジン
キナーゼプロモーター(Wagnerら、1981,proc.Natl.Acad.Sci.U.S.A.78:1441-1445)
、メタロチオネイン遺伝子の調節配列(Brinster ら、1982,Nature 296:39-42);β−ラク
タマーゼプロモーター(Villa-Kamaroffら、1978,Proc.Natl.Acad.Sci.U.S.A.75:37
27-3731)またはtacプロモーター(DeBoerら、1983,Proc.Natl.Acad.Sci.U.S.A.80:2
1-25)などの原核生物発現ベクター;Scientific American,1980 242:74-94の「組換え細
菌由来の有用なタンパク質」も参照; ノパリンシンテターゼプロモーター領域を含む植物
発現ベクター(Herrera-Estrellaら、Nature303:209-213)またはカリフラワーモザイクウ
イルス35SRNAプロモーター(Gardnerら、1981,Nucl.AcidsRes.9:2871)、および光
合成酵素リブロースビスリン酸カルボキシラーゼのプロモーター(Herrera-Estrellaら、1
984,Nature310:115-120);酵素または他の真菌類由来のプロモーター要素、例えばGal4
プロモーター、ADC(アルコールデヒドロゲナーゼ)プロモーター、PGK(ホスホグ
リセロールキナーゼ)プロモーター、アルカリホスファターゼプロモーター、および組織
特異性を示し、トランスジェニック動物に利用されている下記の動物転写制御領域など;
膵小胞細胞において活性なエラスターゼI遺伝子制御領域(Swiftら、1984,Cell 38:639-
646; Ornitzら、1986,ColdSpring Harbor Symp.Quant.Biol.50:399-409; MacDonald
,1987,Hepatology7:425-515); 膵臓β細胞において活性なインスリン遺伝子制御領域(
Hanahan,1985,Nature315:115-122)、リンパ系細胞において活性な免疫グロブリン遺伝
子制御領域(Grosschedlら、1984,Cell38:647-658; Adamesら、1985,Nature 318:533-5
38; Alexanderら、1987,Mol.Cell.Biol.7:1436-1444)、精巣、乳房、リンパ系および
マスト細胞において活性なマウス乳癌ウイルス制御領域(Lederら、1986,Cell 45:485-49
5)、肝臓において活性なアルブミン遺伝子制御領域(Pinkertら、1987,Genesand Devel
.1:268-276)、肝臓において活性なα−フェトプロテイン遺伝子制御領域(Krumlaufら、1
985,Mol.Cell.Biol.5:1639-1648;Hammerら、1987,Science 235:53-58)、肝臓にお
いて活性なα1−抗トリプシン遺伝子制御領域(Kelsey ら、1987,Genes and Devel.1:1
61-171)、骨髄性細胞において活性なβ−グロビン遺伝子制御領域(Mogramら、1985,Natu
re 315:338-340;Kolliasら、1986,Cell 46:89-94); 脳のオリゴデンドロサイト細胞に
おいて活性なミエリン塩基性タンパク質遺伝子制御領域(Readheadら、1987,Cell 48:703
-712); 骨格筋において活性なミオシン軽鎖−2遺伝子制御領域(Sani,1985,Nature314
:283-286)、および視床下部において活性な性腺刺激ホルモン放出ホルモン遺伝子制御領
域(Masonら、1986,Science234:1372-1378)が挙げられる。
【0049】
Delta遺伝子挿入物を含む発現ベクターは、一般的な3つの方法によって同定できる。
すなわち、(a)核酸ハイブリダイゼーション、(b)「マーカー」遺伝子機能の有無、
および(c)挿入配列の発現である。最初の方法では、発現ベクターに挿入した外来遺伝
子の存在を、挿入したトポリズミック遺伝子に相同な配列を含むプローブを使用した核酸
ハイブリダイゼーションにより検出することができる。第二の方法では、組換えベクター
/宿主系を、外来遺伝子のベクターへの挿入によって引き起こされるある種の「マーカー
」遺伝子機能(例えば、チミジンキナーゼ活性、抗生物質に対する耐性、形質転換表現型
、バキュロウイルスにおける閉鎖体の形成など)の有無に基づいて同定し、選択すること
ができる。例えば、Delta遺伝子をベクターのマーカー遺伝子配列内に挿入する場合、Del
ta挿入物を含む組換え体は、マーカー遺伝子機能の不在によって同定できる。第三の方法
では、組換え発現ベクターを、組換え体によって発現される外来遺伝子産物をアッセイす
ることにより同定することができる。そのようなアッセイは、例えば、Delta遺伝子産物i
n vitroアッセイ系の物理的または機能的特性、例えば、ノッチとの集合(結合)、受容
体への結合、抗体との結合に基づくことができる。
【0050】
特定の組換えDNA分子がいったん同定され、単離されると、当技術分野で公知のいく
つかの方法を使用してそれを増殖することができる。適切な宿主系および増殖条件がいっ
たん確立されると、組換え発現ベクターをたくさん増殖し、作製することができる。先に
説明したように、使用できる発現ベクターとしては、それらに限定されないが、次のベク
ターまたはそれらの誘導体が挙げられる。すなわち、ほんの少し挙げると、ワクシニアウ
イルスまたはアデノウイルスなどのヒトまたは動物ウイルス;バキュロウイルスなどの昆
虫ウイルス;酵母ベクター;バクテリオファージベクター(例えば、ラムダ)、ならびに
プラスミドおよびコスミドベクターである。
【0051】
さらに、宿主細胞菌株は、挿入配列の発現を調節し、または遺伝子産物を所望の特定の
様式で改変し、処理するものを選択することができる。ある種のプロモーターからの発現
は、ある種の誘発剤の存在下で高めることかできる。すなわち、遺伝子操作したデルタタ
ンパク質の発現は、制御することができる。さらに、種々の宿主細胞は、タンパク質の翻
訳および翻訳後プロセシングならびに改変(例えば、〔例えば、シグナル配列の〕グリコ
シル化、切断)に対して特徴的かつ特異的な機構を有する。適切な細胞系または宿主系は
、発現された外来タンパク質の所望の改変およびプロセシングが確実に行われるように選
択できる。例えば、細菌系での発現を使用すると、グリコシル化されていないコアタンパ
ク質産物を産生することができる。酵母での発現は、グリコシル化タンパク質を産生する
。哺乳類細胞での発現は、異質哺乳類デルタタンパク質の「天然の」グリコシル化を確実
に行うことができる。さらに、種々のベクター/宿主発現系により、タンパク質分解など
の処理反応を種々の程度に行うことができる。
【0052】
他の特定の態様では、デルタタンパク質、断片、類似体または誘導体を、融合またはキ
メラタンパク質産物((種々のタンパク質の)異質タンパク質配列にペプチド結合によっ
て結合したタンパク質、断片、類似体または誘導体を含む)として発現させることができ
る。そのようなキメラ産物は、所望のアミノ酸配列をコードする適切な核酸配列を、適切
なコードフレームにおいて、当技術分野で公知の方法によって互いに連結し、キメラ産物
を当技術分野で一般に公知の方法によって発現することにより作ることができる。あるい
は、そのようなキメラ産物は、タンパク質合成技術、例えばペプチド合成機を使用するこ
とにより、作ることができる。
【0053】
cDNAおよびゲノム配列は共に、クローン化し、発現させることができる。
【0054】
5.3.Delta遺伝子産物の同定および精製
特定の発明では、本発明は、脊椎動物のデルタ、好ましくはヒトのデルタおよび抗原決
定基を含む(すなわち、抗体によって認識できる)、または、そうでなければ機能的に活
性なそれらの断片および誘導体のアミノ酸配列、ならびにそれらをコードする核酸配列を
提供する。本明細書で使用する「機能的に活性な」物質とは、全長(野生型)のデルタタ
ンパク質に関連する1個以上の公知の機能活性、例えば、ノッチまたはその一部への結合
、他のデルタリガンドへの結合、抗原性(抗−Delta抗体への結合)などを示す物質を意
味する。
【0055】
特定の態様では、本発明は、少なくとも6個のアミノ酸、10個のアミノ酸、25個のアミ
ノ酸、50個のアミノ酸または少なくとも75個のアミノ酸から成るデルタタンパク質の断片
を提供する。該断片を含む分子も提供する。他の態様では、タンパク質は、脊椎動物のデ
ルタタンパク質の細胞外ドメイン、DSLドメイン、上皮増殖因子様反復(ELR)ドメ
イン、ELRのいずれかの組み合わせ、膜貫通ドメイン、もしくは細胞内(細胞質)ドメ
イン、もしくはノッチに結合する部分、またはそれらの組み合わせを含み、あるいは、本
質的にそれらから成る。デルタタンパク質の前記領域のいくつか若しくは全部が欠けた断
片、または断片を含むタンパク質も提供する。前記をコードする核酸を提供する。
【0056】
Delta遺伝子配列を発現する組換え体がいったん同定されると、遺伝子産物を分析する
ことができる。これは、産物の物理的または機能的特性に基づくアッセイ、例えば、産物
の放射能標識および続くゲル電気泳動、イムノアッセイなどによる分析により達成される
。
【0057】
デルタタンパク質がいったん同定されると、クロマトグラフィー(例えば、イオン交換
、アフィニティーおよびサイジングカラムクロマトグラフィー)、遠心分離、示差溶解度
などの標準的方法、またはタンパク質を精製するための他の標準的方法により、単離・精
製することができる。機能的特性は、適切なアッセイを使用して評価できる(第5.7節
参照)。
【0058】
あるいは、組換え体によって産生されるデルタタンパク質がいったん同定されると、そ
のタンパク質のアミノ酸配列は、組換え体に含まれるキメラ遺伝子のヌクレオチド配列か
ら推定できる。その結果、タンパク質は、当技術分野で公知の標準的化学方法によって合
成できる(例えば、Hunkapiller,M.ら、1984,Nature310:105-111)。
【0059】
本発明の特定の態様では、該デルタタンパク質としては、組換えDNA法または化学合
成法のいずれで産生するにしても、それらに限定されないが、基本アミノ酸配列として、
実質的に図2、8、11または14(配列番号2、10、16および39〜65)に示す
アミノ酸配列の全部または一部を含むもの、ならびに断片および他の誘導体、ならびにそ
れらの類似体が挙げられる。
【0060】
5.4.Delta遺伝子およびタンパク質の構造
脊椎動物のDelta遺伝子およびタンパク質の構造は、当技術分野で公知の種々の方法に
よって分析できる。
【0061】
5.4.1.遺伝子分析
Delta遺伝子に対応するクローン化したDNAまたはcDNAは、それらに限定されな
いが、サザンハイブリダイゼーション(Southern,E.M.,1975,J.Mol.Biol.98:503-51
7)、ノーザンハイブリダイゼーション(例えば、Freemanら、1983,Proc.Natl.Acad.
Sci.U.S.A.80:4094-4098を参照)、制限エンドヌクレアーゼマッピング(Maniatis,T.
,1982,MolecularCloning,A Laboratory,Cold Spring Harbor,New York)およびDN
A配列決定分析などの方法によって分析できる。ポリメラーゼ連鎖反応(PCR;米国特
許No.4,683,202、4,683,195および4,889,818;Gyllensteinら、1988,Proc.Natl.Acad
.Sci.U.S.A.85:7652-7656;Ochmanら、1988,Genetics 120:621-623; Lohら、1989,S
cience243:217-220)、およびそれに続くDelta−特異的プローブを用いたサザンハイブリ
ダイゼーションにより、種々の細胞型のDNAにおけるDelta遺伝子の検出が可能である
。PCR以外の増幅法は一般に公知であり、それらも使用できる。一つの態様では、サザ
ンハイブリダイゼーションを使用してDeltaの遺伝子結合を調べることができる。ノーザ
ンハイブリダイゼーション分析を使用すると、Delta遺伝子の発現を調べることができる
。発生または活性の種々の状態にある種々の細胞型のDelta発現を試験することができる
。そのような技法の例およびそれらの結果を、下記第6節に記載する。サザンおよびノー
ザンハイブリダイゼーションのハイブリダイゼーション条件のストリンジェンシーは、使
用する特定のDeltaプローブに対して所望の相関度を有する核酸の検出が確実に行われる
ように操作することができる。
【0062】
制限エンドヌクレアーゼマッピングを使用すると、Delta遺伝子の大体の遺伝子構造を
決定することができる。制限エンドヌクレアーゼ切断によって誘導される制限地図は、D
NA配列決定分析によって確認できる。
【0063】
DNA配列決定分析は、それらに限定されないが、マキサム・ギルバート法(1980,Met
h.Enzymol.65:499-560)、サンガーのジデオキシ法(Sanger,F.,ら、1977,Proc.Natl
.Acad.Sci.U.S.A.74:5463)、T7DNAポリメラーゼの使用(TaborおよびRichardso
n,米国特許No.4,795,699)または自動DNA配列決定装置の使用(例えば、AppliedBios
ystems,Foster City,CA)などの当技術分野で公知の任意の技術によっても行うことがで
きる。
【0064】
5.4.2.タンパク質の分析
デルタタンパク質のアミノ酸配列は、DNA配列からの推定、あるいは、例えば自動ア
ミノ酸配列決定装置を使用する、タンパク質の直接の配列決定により誘導できる。代表的
なデルタタンパク質のアミノ酸配列は、実質的に図2に示し、下記第6節で詳述する配列
を含み、代表的な成熟タンパク質は、アミノ酸番号1〜728によって示される。
【0065】
デルタタンパク質の配列は、親水性分析(Hopp,T.およびWoods,K.,1981,Proc.Nat
l.Acad.Sci.U.S.A.78:3824)によってさらに解析することができる。親水性の特徴は
、デルタタンパク質の疎水性および親水性領域ならびに該領域をコードする遺伝子配列の
対応する領域の同定に使用できる。恐らく、親水性領域が免疫原性であると思われる。
【0066】
二次構造分析(Chou,P.およびFasman,G.,1974,Biochemistry13:222)も、デルタの
特異的二次構造と考えられる領域の同定のために行うことができる。
【0067】
操作、翻訳および二次構造の推定、ならびにオープンリーディングフレームの推定およ
びプロッティングは、当技術分野で利用できるコンピューターソフトウェアプログラムを
使用して行うこともできる。
【0068】
構造分析の他の方法も使用できる。これらには、それらに限定されないが、X−線結晶
学(Engstom,A.,1974,Biochem.Exp.Biol.11:7-13)およびコンピューターによる模型
作成(Fletterick,R.およびZoller,M.編、1986,ComputerGraphics and Molecular M
odeling,inCurrent Communications in Molecular Biology,Cold Spring Harbor Labo
ratory,ColdSpring Harbor,New York)が含まれる。
【0069】
5.5.デルタ(Delta)タンパク質およびその誘導体に対する抗体の生成
本発明に従って、脊椎動物のデルタタンパク質、その断片、その他の誘導体、またはそ
れらのアナログを免疫原として使用してそのような免疫原を認識する抗体を生成すること
ができる。そのような抗体としては、ポリクローナル、モノクローナル、キメラ、単鎖、
Fab断片およびFab発現ライブラリーが包含されるがこれらに限られるわけではない
。特定の態様においてはヒトデルタに対する抗体を産生する。別の態様においてはデルタ
の細胞外ドメインに対する抗体を産生する。また別の態様においてはデルタの細胞内ドメ
インに対する抗体を産生する。
【0070】
デルタタンパク質または誘導体またはアナログに対するポリクローナル抗体を産生する
には業界で公知の各種手法を用いることができる。特定の態様においては、図1a、1b
、7または11に示す配列またはそのサブ配列によってコードされているデルタタンパク
質のエピトープに対するウサギのポリクローナル抗体を得ることができる。抗体の産生に
は、天然のデルタタンパク質、または合成物、またはそれらの誘導体(たとえば断片)を
注射することによって各種宿主動物、たとえばウサギ、マウス、ラットなど(これらに限
られるわけではない)を免疫することができる。免疫応答を高めるために、宿主の種に応
じて各種のアジュバントを用いることができる。たとえば、フロイント(完全および不完
全)、鉱物質ゲルたとえば水酸化アルミニウム、表面活性物質たとえばリゾレシチン、プ
ルロニックポリオール、ポリアニオン、ペプチド、油乳濁液、アオガイヘモシアニン、ジ
ニトロフェノール、ならびに、効力増強に有用なヒトアジュバントたとえばBCG(bacil
le Calmette-Guerin)およびコリネバクテリウム・パルブム(corynebacteriumparvum)が
あるがこれらに限られるわけではない。
【0071】
デルタタンパク質配列またはそのアナログに対するモノクローナル抗体を調製するには
、連続細胞系を培養して抗体分子を産生する技術が使用できる。たとえば、最初Kohlerと
Milsteinによって開発されたハイブリドーマ法(1975,Nature256:495-497)、ならびにト
リオーマ法、ヒトB細胞ハイブリドーマ法(Kozbor et al.,1983,Immunology Today 4:7
2)、およびヒトモノクローナル抗体を産生するEBV‐ハイブリドーマ法(Coleet al.,
1985,inMonoclonal Antibodies and Cancer Therapy,Alan R.Liss,Inc.,pp.77-96
)。本発明の別の態様においては、最近の技術(PCT/US90/02545)を利用して無菌動物でモ
ノクローナル抗体を産生することができる。本発明によると、ヒトハイブリドーマ(Cote
et al.,1983,Proc.Natl.Acad.Sci.U.S.A.80:2026-2030)を使用することによって
、またはヒトB細胞をin vitroでEBVウイルスにより形質転換することによって(Cole
et al.,1985,Monoclonal Antibodies and Cancer Therapy, Alan R.Liss,pp.77-96)
、ヒト抗体を使用してもよいし、また得ることができる。実際、本発明に従って、適当な
生物活性をもつヒト抗体分子由来の遺伝子と共にデルタに特異的なマウス抗体分子由来の
遺伝子をスプライスすることによって「キメラ抗体」を産生するように開発された技術(M
orrison et al.,1984,Proc.Natl.Acad.Sci.U.S.A.81:6851-6855;Neuberger et a
l.,1984,Nature312:604-608; Takeda et al.,1985,Nature 314:452-454)を使用する
ことができる。このような抗体は本発明の範囲内に入る。
【0072】
本発明によると、単鎖抗体の産生について記載された技術(米国特許第4,946,778号)
を、デルタ特異的単鎖抗体の産生に適合させることができる。本発明の別の態様では、F
ab発現ライブラリーの構築に関して記載された技術(Huse et al.,1989,Science 246:
1275-1281)を利用して、デルタタンパク質、誘導体またはアナログに対する所望の特異性
をもつモノクローナルFab断片の迅速で容易な同定が可能になる。
【0073】
分子のイディオタイプを含有する抗体断片は公知の技術によって生成することができる
。そのような断片としては、たとえば、抗体分子のペプシン消化によって生成させること
ができるF(ab′)2断片、このF(ab′)2断片のジスルフィドブリッジを還元す
ることによって生成させることができるFab′断片、およびパパインと還元剤によって
抗体分子を処理することによって生成させることができるFab断片があるがこれらに限
られるわけではない。
【0074】
抗体の産生において、所望の抗体のスクリーニングは業界で公知の技術、たとえばEL
ISA(固相酵素免疫検定法)によって達成することができる。たとえば、脊椎動物のデ
ルタタンパク質の特異的ドメインを認識する抗体を選択するためには、そのようなドメイ
ンを含有するデルタ断片に結合する生成物に対して生成したハイブリドーマを検定すれば
よい。ヒトデルタに対して免疫特異的な抗体を選択するには、ヒトデルタに対する陽性の
結合およびキイロショウジョウバエデルタに対する結合欠如を基にして選択することがで
きる。
【0075】
以上の抗体は、本発明のタンパク質配列の位置決定と活性に関して業界で公知の方法に
、たとえば、これらのタンパク質のイメージング、適当な生理学的サンプル中でのそのレ
ベルの測定、診断法、などに使用することができる(たとえば、後記5.7節参照)。
【0076】
デルタタンパク質の或るドメインに対して特異的な抗体も提供される。特定の態様では
デルタのノッチ結合性断片に結合する抗体が提供される。
【0077】
本発明の別の態様では(後記参照)、抗‐デルタ抗体およびその結合性ドメインを含有
する断片が治療薬である。
【0078】
5.6.デルタタンパク質、誘導体およびアナログ
本発明はさらに、脊椎動物(たとえば哺乳動物)のデルタタンパク質、ならびに脊椎動
物のデルタタンパク質の誘導体(たとえば断片が包含されるがこれに限定はされない)お
よびアナログに係る。デルタタンパク質誘導体およびタンパク質アナログをコードする核
酸も提供される。ひとつの態様では、デルタタンパク質は上記5.1節に記載したデルタ
核酸によってコードされている。特定の態様では、タンパク質、誘導体またはアナログは
マウス、ニワトリ、ラット、ブタ、ウシ、イヌ、サルまたはヒトのデルタタンパク質のも
のである。特定の態様では脊椎動物の完全長成熟デルタタンパク質が提供される。ひとつ
の態様では、シグナル配列(アミノ末端の最初の約17個のアミノ酸)のみを欠く脊椎動
物のデルタタンパク質が提供される。
【0079】
Deltaに関連する誘導体とアナログの産生と用途は本発明の範囲内である。特定の態様
の場合、誘導体またはアナログは機能的に活性である。すなわち、野生型の完全長デルタ
タンパク質に関連する機能活性を1つ以上示すことができる。一例を挙げると、所望の免
疫原性または抗原性を有するそのような誘導体やアナログは、たとえば免疫検定、免疫、
デルタ活性の抑制、などに使用することができる。所望のデルタ特性、たとえばノッチそ
の他のトポリズミックタンパク質との結合、細胞表面受容体との結合を保持または逆に抑
制するそのような分子はそれぞれそのような特性およびその相関物のインデューサーまた
はインヒビターとして使用することができる。ひとつの特定の態様は、抗‐デルタ抗体と
結合することができるがノッチタンパク質またはその他のトポリズミックタンパク質とは
結合することができないデルタ断片に関する。デルタの誘導体またはアナログの所望の活
性は、限定されることはないが5.7節に記載する検定を始めとする業界で公知の手法に
よって試験することができる。
【0080】
特に、デルタ誘導体は、機能的に等価な分子を提供する置換、付加または欠失によりDe
lta配列を変更することによって作成することができる。ヌクレオチドコード配列の縮重
のため、Delta遺伝子と実質的に同じアミノ酸配列をコードする他のDNA配列を本発明
の実施の際に使用してもよい。これらの中には、配列内のアミノ酸残基と機能的に等価な
アミノ酸残基をコードする異なるコドンの置換によって変更されてサイレント変異を生じ
ているDelta遺伝子の全部または一部からなるヌクレオチド配列が包含されるがこれらに
限定されることはない。同様に、本発明のデルタ誘導体には、一次アミノ酸配列として、
配列内の残基が機能的に等価なアミノ酸残基で置換されてサイレント変異を生じている変
更配列を含むデルタタンパク質のアミノ酸配列の全部または一部を含有するものが包含さ
れるがこれらに限られることはない。たとえば、配列内のアミノ酸残基の1つ以上を、機
能的に等価な作用をする類似の極性をもった別のアミノ酸で置換してサイレント変異を生
じさせることができる。配列内のアミノ酸の置換はそのアミノ酸が属するクラスの他の一
員の中から選択すればよい。たとえば、非極性(疎水性)アミノ酸にはアラニン、ロイシ
ン、イソロイシン、バリン、プロリン、フェニルアラニン、トリプトファンおよびメチオ
ニンが含まれる。極性の中性アミノ酸にはグリシン、セリン、スレオニン、システイン、
チロシン、アスパラギンおよびグルタミンが含まれる。正に帯電した(塩基性)アミノ酸
にはアルギニン、リシンおよびヒスチジンが含まれる。負に帯電した(酸性)アミノ酸に
はアスパラギン酸とグルタミン酸が含まれる。
【0081】
本発明の特定の態様では、デルタタンパク質の少なくとも10個の(連続する)アミノ
酸で構成される脊椎動物デルタタンパク質の断片で構成されるかまたはこれを含むタンパ
ク質が提供される。他の態様では、デルタタンパク質の少なくとも20個または50個の
アミノ酸で構成される断片が提供される。特定の態様ではそのような断片が35個、10
0個または200個までのアミノ酸である。デルタの誘導体またはアナログとしては、脊
椎動物のデルタタンパク質またはその断片と実質的に相同であるペプチド(たとえば、同
じサイズのアミノ酸配列と比べてその少なくとも30%、50%、70%または90%が
同一のもの、たとえば、あるドメインを含むもの)、またはDeltaコード配列とハイブリ
ダイズすることができるコード核酸をもつペプチドが包含されるがこれらに限定されるこ
とはない。
【0082】
本発明のデルタ誘導体およびアナログは業界で公知のさまざまな方法によって生成する
ことができる。その生成に至る操作は遺伝子レベルまたはタンパク質レベルで実施するこ
とができる。たとえば、クローン化したDelta遺伝子配列は、業界で公知の多くの方法(Ma
niatis,T.,1990,MolecularCloning,A Laboratory Manual,2d ed.,Cold Spring Ha
rbor Laboratory,ColdSpring Harbor,New York)のいずれかによって修飾することがで
きる。前記配列は、適当な部位で(1種以上の)制限エンドヌクレアーゼによって開裂し
た後所望の場合にはさらに酵素修飾し、単離し、in vitroで連結することができる。デル
タの誘導体またはアナログをコードする遺伝子を生成する際には、そのようにして修飾さ
れた遺伝子がデルタと同じ翻訳リーディングフレーム内にあり、所望のデルタ活性がコー
ドされている遺伝子領域内で翻訳終結シグナルによって中断されないように注意しなけれ
ばならない。
【0083】
さらに、デルタをコードしている核酸配列をin vitroまたはin vivoで突然変異させて
、翻訳配列、開始配列および/または終結配列を創製および/または破壊したり、または
コード領域において変異を起こさせたり、および/または新たな制限エンドヌクレアーゼ
部位を形成するかもしくはすでに存在していたものを破壊したりして、さらなるin vitro
修飾を容易にすることができる。限定するわけではないがin vitro位置指定突然変異誘発
(Hutchinson,C.,etal.,1978,J.Biol.Chem 253:6551)、TAB(登録商標)リンカ
ー(Pharmacia)の利用、などを始めとして業界で公知のいかなる突然変異誘発法も用いる
ことができる。変化した配列を含有するPCRプライマーをPCRに用いて、増幅される
断片中にそのような変化を導入することができる。
【0084】
デルタ配列の操作はまたタンパク質レベルで行なうこともできる。本発明の範囲内に包
含されるものとして、翻訳中またはその後、たとえばグリコシル化、アセチル化、リン酸
化、アミド化、公知の保護基/ブロック基による誘導体化タンパク質分解酵素による開裂
、抗体分子その他の細胞性リガンドとの結合、などによって、さまざまに修飾されたデル
タタンパク質断片またはその他の誘導体もしくはアナログがある。限定するわけではない
が臭化シアン、トリプシン、キモトリプシン、パパイン、V8プロテアーゼ、NaBH4
による特異的化学開裂、アセチル化、ホルミル化、酸化、還元、ツニカマイシンの存在下
での代謝合成、などを始めとする多くの化学修飾のいずれも公知の技術によって実施する
ことができる。
【0085】
さらに、デルタのアナログおよび誘導体は化学的に合成することができる。たとえば、
所望のドメインを含む(5.6.1節参照)か、またはin vitroにおける所望の凝集活性
もしくは受容体との結合を媒介するデルタタンパク質の一部に相当するペプチドを、ペプ
チドシンセサイザーを使用して合成することができる。さらに、所望であれば、非古典的
なアミノ酸または化学的アミノ酸アナログを置換または付加としてデルタ配列中に導入す
ることができる。非古典的なアミノ酸としては、通常のアミノ酸のD‐異性体、α‐アミ
ノイソ酪酸、4‐アミノ酪酸、ヒドロキシプロリン、サルコシン、シトルリン、システイ
ン酸、t‐ブチルグリシン、t‐ブチルアラニン、フェニルグリシン、シクロヘキシルア
ラニン、β‐アラニン、設計用(designer)アミノ酸、たとえばβ‐メチルアミノ酸、Cα
‐メチルアミノ酸、およびNα‐メチルアミノ酸が包含されるがこれらに限定されるわけ
ではない。
【0086】
特定の態様の場合、デルタ誘導体は、脊椎動物のデルタタンパク質またはその断片(好
ましくは、デルタタンパク質の少なくともひとつのあるドメインもしくは一部、またはデ
ルタタンパク質の少なくとも10個のアミノ酸)が、そのアミノまたはカルボキシ末端に
おいてペプチド結合を介して別のタンパク質のアミノ酸配列と結合したキメラタンパク質
すなわち融合タンパク質である。ひとつの態様の場合、そのようなキメラタンパク質は、
そのタンパク質をコードする(別のタンパク質のコード配列と同じフレーム内で結合した
デルタコード配列からなる)核酸の組換え発現によって産生される。そのようなキメラ生
成物は、業界で公知の方法によって所望のアミノ酸配列をコードする適当な核酸配列を適
正なコードフレームで互いに連結し、業界で周知の方法によってそのキメラ生成物を発現
させることにより作成できる。あるいはまた、そのようなキメラ生成物はタンパク質合成
技術によって、たとえばペプチドシンセサイザーを用いて作成できる。特定の態様では、
異種シグナル配列と共に成熟デルタタンパク質をコードするキメラ核酸を発現させて、そ
のキメラタンパク質が細胞によって発現され成熟デルタタンパク質にプロセッシングされ
るようにする。限定するわけではないが別の一例では、Delta遺伝子と別のトポリズミッ
ク遺伝子(たとえばセレート)との両方のコード部分を含む組換え分子を本発明に従って
構築することができる。そのような組換え分子にコードされているタンパク質はセレート
とデルタの両方に関連する特性を発揮し、アゴニストとアンタゴニストを含めて新規なプ
ロフィールの生物学的活性を示すことができるであろう。またデルタとセレートの一次配
列を用いて、コンピューターシミュレーションによってその分子の三次構造を予想するこ
ともでき(Hoppand Woods,1981,Proc.Natl.Acad.Sci.U.S.A.78:3824-3828)、Delt
a/Serrateキメラ組換え遺伝子は三次構造と生物学的機能との相関を考慮に入れて設計す
ることができよう。同様に、任意の異種タンパク質コード配列と融合したDeltaの部分を
含むキメラ遺伝子を構築することができる。ひとつの特定の態様はデルタの少なくとも6
個のアミノ酸の断片を含むキメラタンパク質に関する。
【0087】
別の特定の態様の場合、デルタ誘導体は、別のトポリズミックタンパク質と相同の領域
を含む脊椎動物のデルタの断片である。本明細書で使用する場合、第一のタンパク質の領
域は、その領域内に含まれるアミノ酸の数と等しい数のアミノ酸をもつ第二のタンパク質
内の任意の配列と比較した場合、その領域のアミノ酸配列が少なくとも30%同一である
かまたは少なくとも70%が同一であるかもしくは保存性の変化を含んでいるとき、第二
のタンパク質と「相同である」と考えることとする。たとえば、そのようなデルタ断片は
、限定するわけではないがDSLドメインまたはその一部を含めて、セレートと相同な領
域を1つ以上含んでいることができる。
【0088】
誘導体およびアナログの他の特定の具体例は以下の小節および後記実施例で記載する。
【0089】
5.6.1.本タンパク質の1以上のドメインを含有するデルタ誘導体
ひとつの特定の態様において、本発明は、限定することはないがたとえば細胞外ドメイ
ン、シグナル配列、DSLドメインのアミノ末端領域、DSLドメイン、ELRドメイン
、膜貫通ドメイン、細胞内ドメイン、およびデルタタンパク質の1以上のEGF様反復ド
メイン(ELR)(たとえば、ELR1〜9)、またはこれらの任意の組合せを始めとす
るデルタタンパク質の1以上のドメインを含むかあるいはそれで構成される脊椎動物のデ
ルタ誘導体およびアナログ、特にデルタの断片およびそのような断片の誘導体に係る。ニ
ワトリおよびマウスのデルタタンパク質に関する特定の例の場合、そのようなドメインは
それぞれ後記実施例6および7ならびに図3および9で同定する。すなわち、例として、
脊椎動物デルタの細胞外ドメイン(アミノ酸約1〜545)、シグナル配列(アミノ酸約
1〜17)、DSLドメインのアミノ末端領域(アミノ酸約1〜178)、DSLドメイ
ン(アミノ酸約179〜223)、EGF1(アミノ酸約229〜260)、EGF2(
アミノ酸約261〜292)、EGF3(アミノ酸約293〜332)、EGF4(アミ
ノ酸約333〜370)、EGF5(アミノ酸約371〜409)、EGF6(アミノ酸
約410〜447)、EGF7(アミノ酸約448〜485)、EGF8(アミノ酸約4
86〜523)、膜貫通ドメインおよび細胞内(細胞質)ドメイン(アミノ酸約555〜
728)からなる分子が提供される。
【0090】
特定の態様の場合、脊椎動物のデルタの特定の断片を含む分子は、それぞれのデルタタ
ンパク質の中でキイロショウジョウバエまたはニワトリのデルタタンパク質の特定の断片
と最も相同である断片を含むものである。特定の具体例の場合そのような分子は配列番号
2または16のアミノ酸配列を含むかまたはその配列で構成される。あるいはまた、デル
タ相同体のドメインを含む断片は5.3.2節に記載したタンパク質解析法によって同定
することができる。
【0091】
5.6.2.トポリズミックタンパク質ドメインとの結合を媒介するデルタの誘導体
本発明はまた、トポリズミックタンパク質との結合を媒介する(したがって本明細書で
は「接着性」と名付ける)脊椎動物のデルタ断片およびそのような断片のアナログまたは
誘導体ならびにそれをコードする核酸配列も提供する。
【0092】
ひとつの特定の態様の場合デルタタンパク質の接着性断片はDSLドメインまたはその
一部からなる。DSLドメイン内で、ノッチとの結合を媒介するサブ断片は欠失突然変異
体を発現する構築物の解析によって同定することができる。
【0093】
トポリズミックタンパク質(好ましくはノッチ)と結合する能力は、そのようなトポリ
ズミックタンパク質を発現する細胞およびデルタまたはデルタ誘導体を発現する細胞を用
いるinvitro凝集検定によって立証することができる(5.7節参照)。すなわち、デル
タ断片がノッチタンパク質と結合する能力は、第一の細胞の表面上で発現した場合のデル
タ断片が、第二の細胞の表面上で発現したノッチタンパク質と結合する能力を検出するこ
とによって立証することができる。
【0094】
そのような検定に使用する、局所周期性タンパク質またはその接着性ドメインをコード
する核酸配列は、ヒト、ブタ、ウシ、ネコ、トリ、ウマ、イヌまたは昆虫ならびに霊長類
および公知のトポリズミック遺伝子のホモログが同定され得るその他の任意の種から単離
することができる。
【0095】
5.7.デルタタンパク質、誘導体およびアナログの検定
脊椎動物のデルタタンパク質、誘導体およびアナログの機能活性は各種方法によって検
定することができる。
【0096】
たとえば、ひとつの態様において、抗‐デルタ抗体に結合する能力またはその抗体との
結合に関して野生型デルタと競合する能力を検定する場合、業界で公知の各種免疫検定法
を使用することができる。たとえば、放射線免疫検定法、ELISA(固相酵素免疫検定
法)、「サンドイッチ」免疫検定法、イムノラジオメトリックアッセイ、ゲル核酸沈降反
応(gel diffusionprecipitin reactions)、免疫核酸検定法、(たとえば金コロイド、酵
素または放射性同位体標識を用いる)in situ免疫検定法、ウェスタンブロット、沈降反
応、凝集検定法(たとえばゲル凝集検定法、赤血球凝集検定法)、補体結合検定法、免疫
蛍光検定法、プロテインAアッセイ、および免疫電気泳動検定法、などのような技術を用
いる競合および非競合検定系があるがこれらに限定されることはない。ひとつの態様では
、一次抗体の標識を検出することによって抗体結合を検出する。別の態様では、二次抗体
または試薬と一次抗体の結合を検出することによって一次抗体を検出する。さらに別の態
様では二次抗体を標識する。免疫検定法で結合を検出するための多くの方法が業界で知ら
れており、本発明の範囲内である。
【0097】
別の態様において、トポリズミックタンパク質(たとえばノッチ)との結合を媒介する
能力を検定しようとする場合、in vitro凝集検定法を実施することができる(Fehon et al
.,1990,Cell61:523-534; Rebay et al.,1991,Cell 67:687-699)。 デルタに対する
受容体を同定する別の態様では、たとえば業界で周知の手段によって受容体の結合を検定
することができる。別の態様では、デルタの結合とデルタ受容体を発現する細胞との生理
学的相関(シグナル伝達)を検定することができる。
【0098】
別の態様では、昆虫やその他のモデル系において、遺伝子研究によって、野生型Delta
の誘導体またはアナログであるデルタ突然変異の表現型上の効果を研究することができる
。
【0099】
その他の方法も当業者には知られており、本発明の範囲内である。
【0100】
5.8.治療用途
本発明は、本発明の治療用化合物を投与することによって細胞の運命または分化の障害
を処置する方法を提供する。このような治療用化合物(本明細書中では「治療薬」とする
)としては、デルタタンパク質ならびにそのアナログおよび誘導体(たとえば断片)(た
とえば、上記したようなもの)、それに対する抗体(上記したようなもの)、デルタタン
パク質、アナログまたは誘導体(たとえば上記したようなもの)をコードする核酸、なら
びにDeltaアンチセンス核酸がある。上に述べたように、本発明のアンタゴニスト治療薬
は、デルタ機能および/または(デルタはノッチリガンドであるので)ノッチ機能と拮抗
するかまたはこれを抑制する治療薬である。そのようなアンタゴニスト治療薬は、たとえ
ばデルタと別のタンパク質(たとえばノッチタンパク質)との結合を抑制する当該薬の能
力または好ましくはinvitroでもしくは細胞培養で検定されるように公知のノッチまたは
デルタ機能を抑制する当該薬の能力に基づいた公知の簡便なin vitro検定法を用いて同定
するのが最も好ましい。もっとも、遺伝子検定法(たとえば、キイロショウジョウバエ)
の場合)も使用することができる。好ましい態様の場合、アンタゴニスト治療薬は、ノッ
チとの結合を媒介するデルタの断片のような機能的に活性な断片を含むタンパク質もしく
はその誘導体またはそれに対する抗体である。他の特定の態様では、そのようなアンタゴ
ニスト治療薬がノッチに結合するデルタの断片を含む分子を発現することができる核酸ま
たはDeltaアンチセンス核酸である(5.11節参照)。好ましくは、特定の治療薬の効
果を測定し、かつその投与が冒された組織の処置にとって必要であるかどうかを決定する
には、後述する適切なin vitroまたはin vivo 検定法を用いるべきであることに留意され
たい。その理由は、冒された組織の発生履歴はあるアンタゴニストまたはアゴニスト治療
薬が望まれるかどうかを決定し得るからである。
【0101】
また、デルタタンパク質または誘導体の投与様式、たとえば、可溶性形態で投与するの
か、または細胞内組換え発現用にそれをコードしている核酸を介して投与するのかは、そ
れがアゴニストとして作用するのかまたはアンタゴニストとして作用するのかに影響し得
る。
【0102】
本発明の別の態様においては、Delta遺伝子の一部を含有する核酸をアンタゴニスト治
療薬として用いて相同組換えによるDelta不活化を促進する(Koller and Smithies,1989
,Proc.Natl.Acad.Sci.USA86:8932-8935; Zijlstra et al.,1989,Nature 342:435
-438)。
【0103】
本発明のアゴニスト治療薬は上記したようにデルタ機能を促進する。そのようなアゴニ
スト治療薬としては、デルタとの結合を媒介するノッチの一部を含むタンパク質および誘
導体ならびにそれをコードする核酸(これはそれがコードしている産物をin vivoで発現
するように投与することができる)が包含されるがこれらに限られるわけではない。
【0104】
本発明の治療薬の詳細な説明と起源は本明細書中の5.1〜5.7節に記載した。
【0105】
所望のデルタ特性、たとえばノッチとの結合、細胞内リガンドとの結合を保持するかま
たは抑制する分子は、それぞれそのような特性およびその生理学的関連物のインデューサ
ーまたはインヒビターとして治療上使用することができる。特定の態様においては、デル
タのノッチと結合する部分の配列を含有するペプチド(たとえば、6〜50または15〜
25アミノ酸の範囲、特に約10、15、20または25のアミノ酸)を用いてノッチ機
能に拮抗させる。ひとつの特定の態様の場合、そのようなアンタゴニスト治療薬を用いて
、ノッチ発現の増大に関連するヒトまたはその他の悪性腫瘍(たとえば、頸管癌、結腸癌
、乳癌、扁平腺癌(後述))を処置または予防する。デルタの誘導体またはアナログの望
ましい活性は、限定されることはないが後述の実施例に記載する検定法を始めとする業界
で公知の手法によって試験することができる。たとえば、ノッチEGF‐反復ドメイン(
ELR)11および12に結合しDSLドメインより小さいデルタ断片を含む分子は、欠
失突然変異体を発現させその発現産物とノッチとの結合をいくつかの方法(たとえばin v
itro細胞凝集検定法、相互作用捕獲系。これらのうちのいくつかは後述の実施例で記載す
る)のいずれかで検定することによって取得し選択することができる。ひとつの特定の態
様においては、ペプチドライブラリーをスクリーニングして所望の活性をもつペプチドを
選択することができる。そのようなスクリーニングは、たとえばノッチまたはノッチのE
LR11および12反復ドメインを含有する分子との結合に関して検定することによって
実施することができる。
【0106】
他の治療薬としては脊椎動物のデルタタンパク質と結合する分子がある。したがって、
本発明はそのような分子を同定する方法も提供する。そのような分子は、複数の分子(た
とえば、ペプチドライブラリー、または組合せ化学ライブラリーのもの)を、結合を促進
する条件下でデルタタンパク質と接触させ、デルタタンパク質と結合する分子を回収する
ことからなる方法によって同定することができる。
【0107】
本発明のアゴニストおよびアンタゴニスト治療薬は細胞宿命の障害に対して治療上有用
である。アゴニスト治療薬は、(1)ノッチまたはデルタ機能の欠如またはレベル低下(
正常または望ましいレベルとの比較)が関与する疾患または障害において、たとえば、ノ
ッチまたはデルタタンパク質が欠如しているか、遺伝的に欠損しているか、生物学的に不
活性もしくは活性不全であるか、または発現不足である患者に、また(2)in vitro(ま
たはin vivo)検定(後述)によりデルタアゴニストの投与が有用であることが示された
疾患または障害において、治療的(予防を含む)に投与する。このノッチまたはデルタ機
能の欠如またはレベル低下は、たとえば、患者の組織サンプル(たとえば生検組織由来)
を得、その発現したノッチまたはデルタタンパク質のタンパク質レベル、構造および/ま
たは活性をinvitroで検定することによって容易に検出することができる。すなわち、業
界で標準的な多くの方法を使用することができる。たとえば、免疫検定法によりノッチま
たはデルタタンパク質を検出および/または可視化する(たとえば、ウェスタンブロット
、免疫沈降とそれに続くドデシル硫酸ナトリウムポリアクリルアミドゲル電気泳動、免疫
組織化学、など)か、かつ/またはハイブリダイゼーションアッセイによりノッチまたは
デルタのmRNAを検出および/または可視化することによってそれぞれノッチまたはデ
ルタの発現を検出する(たとえば、ノーザンアッセイ、ドットブロット、in situハイブ
リダイゼーション、など)ことができるが、これらに限定されることはない。
【0108】
特定のアゴニスト治療薬またはアンタゴニスト治療薬の投与が指示されるかどうかを決
定するのに使用することができるin vitro検定法としては、患者の組織サンプルを培養し
て増殖させ、治療薬に暴露するかまたは他の方法でこれを投与し、そのような治療薬の組
織サンプルに対する効果を観察するというin vitro細胞培養検定法がある。患者が悪性腫
瘍であるひとつの態様の場合、そのような悪性腫瘍からの細胞のサンプルを平板培養また
は培養増殖した後その細胞を治療薬に暴露する。(たとえば、末端分化を促進することに
よって)悪性細胞の生存または増殖を抑制する治療薬をin vivoの治療用として選択する
。そのような生存および/または増殖を評価するには業界で標準的な多くの検定法を使用
することができる。たとえば、細胞増殖は3H‐チミジンの取込みを測定することによっ
て、直接の細胞計数によって、原癌遺伝子(たとえばfos、myc)のような公知の遺伝子ま
たは細胞周期マーカーの転写活性の変化を検出することによって検定することができる。
また、細胞生存度はトリパンブルー染色によって評価することができ、分化は細胞形態の
変化に基づいて目視により評価することができる。特定の局面においては、悪性培養細胞
を(1)アゴニスト治療薬および(2)アンタゴニスト治療薬に別々に暴露する。検定結
果により、どちらの治療薬が治療効果をもっているかどうか指示することができる。
【0109】
別の態様の場合、増殖過多または増殖過少障害を有しているかまたは有すると疑われる
組織から得た患者の細胞サンプルに対して所望の効果、すなわちそれぞれ細胞増殖の抑制
または促進効果を示す治療薬を使用することが指示される。そのような増殖過多または増
殖過少障害としては後記5.8.1〜5.8.3節に記載するものがあるがこれらに限ら
れることはない。
【0110】
別の特定の態様の場合、冒された患者タイプの神経細胞からの神経再生/軸索伸長のin
vitro促進を示す治療薬を、神経損傷または神経系変性障害(5.8.2節参照)の処置
に使用することが指示される。
【0111】
さらに、本発明のアンタゴニスト治療薬はまた、ノッチまたはデルタ優性活性化表現型
(「機能の獲得」突然変異)が関与することが決定されているかまたは知られている疾患
または障害にも投与することが指示される。ノッチまたはデルタ優性陰性表現型(「機能
の損失」突然変異)が関与することが決定されているかまたは知られている疾患または障
害にアゴニスト治療薬を投与することが指示される。ノッチタンパク質の各種構造ドメイ
ンの機能は、hsp70熱ショックプロモーターならびに眼に特異的なプロモーターの下
で一連のキイロショウジョウバエNotch欠失突然変異を異所発現させることによってin vi
voで検討されている(Rebayet al.,1993,Cell 74:319-329参照)。2種類の優性表現
型が観察された。1つはNotch機能損失突然変異を示唆しており、他の1つはNotch機能獲
得突然変異を示唆している。優性「活性化」表現型はほとんどの細胞外配列を欠くタンパ
ク質の発現過多の結果であり、優性「陰性」表現型はほとんどの細胞内配列を欠くタンパ
ク質の発現過多の結果であった。これらの結果は、ノッチが受容体(この受容体の細胞外
ドメインはリガンド結合を媒介する)として機能し、細胞質ドメインによって発生シグナ
ルが伝達されることを示している。
【0112】
各種の特定具体例において、患者の障害に関与する細胞型の代表的な細胞を用いてin v
itro検定を実施して、ある治療薬がそのような細胞型に対して所望の効果を有するのか否
か決定することができる。
【0113】
別の態様においては、新生物発生前であることが疑われる患者の組織サンプルの細胞を
同様に平板培養またはin vitroで増殖させ、治療薬に暴露する。より正常な(すなわち、
新生物発生の状態、腫瘍状態、悪性状態、または形質転換表現型の典型ではない)細胞表
現型を生じる治療薬を治療用に選択する。新生物発生前状態、腫瘍状態、または形質転換
もしくは悪性表現型が存在するかどうか評価するには業界で標準的な多くの検定法を使用
することができる。たとえば、形質転換表現型に関連する特性(in vivo腫瘍形成能力と
関連した一組のinvitro特性)としては、より丸くなった細胞形態、より緩い基層付着、
接触阻害の損失、足場依存性の損失、プラスミノーゲンアクチベーターのようなプロテア
ーゼの放出、糖輸送の増大、血清要求性の低下、胎児抗原の発現、250,000ダルトンの表
面タンパク質の消失、などがある(Luria et al.,1978,General Virology,3d Ed.,Jo
hn Wiley &Sons,New York pp.436-446参照)。
【0114】
他の特定の態様においては、本発明に従って、処置すべき特定の患者に由来する細胞サ
ンプルではなく、処置または予防するのが望まれる悪性、腫瘍または腫瘍発現前の障害に
由来する細胞系またはそのような障害に関連する(1つ以上の)特性を示す細胞系、また
は効果が望まれる神経その他の細胞型に由来する細胞系を用いて上記したin vitro検定を
実施することができる。
【0115】
アンタゴニスト治療薬は、(1)(正常または望ましいレベルと比較して)増大したレ
ベルのノッチまたはデルタ機能が関与する疾患または障害において、たとえば、ノッチま
たはデルタタンパク質が発現過多であるかまたは活性過多である場合に、また(2)in v
itro(またはin vivo)検定によりデルタアンタゴニストの投与が有用であることが示さ
れた疾患または障害において、治療的(予防を含む)に投与する。このノッチまたはデル
タ機能の増大したレベルは、上記したような方法によって、タンパク質および/またはR
NAを定量することによって、容易に検出することができる。治療の有用性を決定するた
めに患者の組織サンプルの細胞または適当な細胞系もしくは細胞型を用いてするin vitro
検定は上記したように実施することができる。
【0116】
5.8.1.悪性腫瘍
上記したようにアンタゴニストまたはアゴニスト治療薬の効力を試験することができ、
またそのようにして治療の有用性が観察されたら処置することができる悪性および前新生
物状態としては、5.8.1および5.9.1節において下記するものが包含されるがこ
れらに限定されることはない。
【0117】
その細胞をinvitro(および/またはin vivo)で試験することができ、適当な検定結
果を観察したら本発明に従って処置することができる悪性腫瘍および関連する障害には、
表1に掲げるものがあるがこれらに限られることはない(そのような障害の概論としては
、Fishmanet al.,1985,Medicine,2d Ed.,J.B.Lippincott Co.,Philadelphiaを参
照されたい)。
【表1】
【0118】
特定の態様においては、頸、食道および肺の上皮組織のような上皮組織の悪性腫瘍また
は悪性増殖(dysproliferative)変化(化生および異形成など)を処置または予防する。
【0119】
結腸および頸部の悪性腫瘍ではヒトノッチの発現がこれらの悪性でない組織と比べて増
大している(1994年4月14日に公開されたPCT公開第WO 94/07474号(引用によりそ
の全体が本明細書に含まれているものとする)参照)。したがって、特定の態様において
は、ノッチ機能と拮抗するアンタゴニスト治療薬(たとえばデルタ誘導体)を有効量投与
することによって結腸または頸部の悪性腫瘍または前悪性変化を処置または予防する。結
腸癌や頸管癌でノッチの発現が増大していることは、もっとたくさんの癌性および増殖過
多状態がアップレギュレートされたノッチを示すことを示唆している。したがって、特定
の態様では、各種癌、たとえば、乳癌、扁平腺癌、セミノーム(精上皮腫)、黒色腫(メ
ラノーマ)および肺癌、ならびにそれらの前悪性変化、さらには他の増殖過多障害を、ノ
ッチ機能と拮抗するアンタゴニスト治療薬の投与によって処置または予防することができ
る。
【0120】
5.8.2.神経系障害
上記したようにアンタゴニストまたはアゴニスト治療薬の効力を試験することができ、
またそのようにして治療の有用性が観察されたら処置することができる細胞型を含む神経
系の障害としては、神経系の損傷、ならびに軸索の遮断、ニューロンの減退もしくは退化
、または脱髄に至る疾患または障害が包含されるがこれらに限定されることはない。本発
明に従って(ヒトおよびヒト以外の哺乳類の患者を含めて)患者において処置できる神経
系の損傷としては、中枢神経系(脊髄、脳を含む)または抹消神経系の以下の損傷が包含
されるがこれらに限定されることはない。
【0121】
(i)物理的損傷によって生じるかまたは外科に伴なう損傷を始めとする外傷、たとえば
、神経系の一部を切断する損傷、または圧迫損傷、
(ii)神経系の一部における酸素の欠乏によってニューロンの損傷または壊死に至る虚血
性損傷、たとえば、脳の梗塞もしくは虚血、または脊髄の梗塞もしくは虚血、
(iii)神経系に関連した悪性または神経系組織以外に由来する悪性である悪性組織によ
って神経系の一部が破壊または損傷される悪性損傷、
(iv)感染の結果、たとえば、膿瘍によって、またはヒト免疫不全ウィルス、帯状ヘルペ
スもしくは単純ヘルペスウィルスによる感染に伴なって、またはライム病、結核、梅毒に
伴なって、神経系の一部が破壊または損傷される感染性損傷、
(v)パーキンソン病、アルツハイマー症、ハンチントン舞踏病または筋萎縮性側索硬化
症に伴なう退化を始めとする(以上のものに限定されるわけではない)退化性過程の結果
として神経系の一部が破壊または損傷される退化性損傷、
(vi)ビタミンB12欠乏症、葉酸欠乏症、ヴェルニッケ病、たばこ‐アルコール性弱視
、マルキャファーバ・ビギャミ病(脳梁の一次退化)、およびアルコール性小脳変性を始
めとする(ただし、これらに限定されることはない)代謝の栄養的障害または障害によっ
て神経系の一部が破壊または損傷される栄養上の疾患または障害に伴なう損傷、
(vii)糖尿病(糖尿病性神経障害、ベル麻痺)、全身性エリテマトーデス、癌腫または
サルコイド症を始めとする(ただし限定されない)全身病に伴なう神経損傷、
(viii)アルコール、鉛、または特定の神経毒を始めとする毒性物質によって引き起こさ
れる損傷、
(ix)多発性硬化症、ヒト免疫不全ウィルス関連脊髄障害、横断性脊髄症または各種の病
因、進行性多病巣性白質エンセファロパシー、および中心性橋脱髄症を始めとする(ただ
し限定はされない)脱髄性の疾患によって神経系の一部が破壊または損傷される脱髄性損
傷。
【0122】
本発明に従って神経系の障害を処置するのに有用な治療薬はニューロンの生存または分
化を促進する生物活性を試験することによって選択できる(5.8節も参照のこと)。た
とえば、限定するわけではないが、次のような効果を有する治療薬が本発明で有用であろ
う。
【0123】
(i)培養におけるニューロンの生存時間の延長、
(ii)培養またはinvivoにおけるニューロンの発芽の増大、
(iii)培養またはinvivoにおけるニューロン関連分子、たとえば運動ニューロンに関す
るコリンアセチルトランスフェラーゼまたはアセチルコリンエステラーゼの産生の増大、
または
(iv)invivoにおけるニューロンの機能不全の症状の低減。
【0124】
このような効果は業界で公知のいかなる方法でも測定できる。好ましい非限定具体例の場
合、ニューロンの生存増大はArakawaらにより記載された方法で測定することができ(1990
,J.Neurosci.10:3507-3515)、ニューロンの発芽の増大はPestronkら(1980,Exp.Neur
ol.70:65-82)により、またはBrownら(1981,Ann.Rev.Neurosci.4:17-42)により記載
された方法で検出でき、ニューロン関連分子の産生増大は測定すべき分子に応じて生物検
定、酵素検定、抗体結合、ノーザンブロットアッセイなどによって測定でき、そして運動
ニューロンの機能不全は運動ニューロンの障害の物理的表示、たとえば弱さ、運動ニュー
ロンの伝達速度、または機能欠損を評価することによって測定することができる。
【0125】
特定の態様の場合、本発明に従って処置できる運動ニューロンの障害としては、運動ニ
ューロンおよび神経系の他の成分に影響を与え得る梗塞、感染、毒素に対する暴露、外傷
、外科的損傷、退化症または悪性のような障害ならびにニューロンに選択的に影響を与え
る筋萎縮性側索硬化症のような障害が包含されるがこれに限定されることはない。たとえ
ば、進行性脊髄性筋萎縮症、進行性球麻痺、一次側索硬化症、乳児性および若年性筋萎縮
症、小児の進行性球麻痺(Fazio-Londe症候群)、ポリオおよびポリオ後症候群、ならび
に遺伝性運動感覚性神経障害(Hereditary Motorsensory Neuropathy)(シャルコー・マリ
ー・ツース病)があるがこれらに限られることはない。
【0126】
5.8.3.組織の修復と再生
本発明の別の態様においては、本発明の治療薬を組織の再生と修復の促進に使用する。
たとえば、良性の異常増殖障害の処置が挙げられるがこれに限られることはない。特定の
態様は、肝臓の硬変(瘢痕形成が正常な肝臓再生過程を追越した状態)の処置、ケロイド
(肥大性の瘢痕)形成(瘢痕形成過程が正常な再生を妨げている皮膚の醜悪化)、乾癬(
皮膚の過剰増殖と適正な細胞宿命決定の遅延を特徴とする通常の皮膚の状態)、および禿
頭(最終的に分化した毛包(ノッチに富む組織)が適切に機能していない状態)の処置に
関する。別の態様では、本発明の治療薬を内耳の感覚上皮の退化性または外傷性の障害を
処置するのに使用する。
【0127】
5.9.予防用途
5.9.1.悪性腫瘍
本発明の治療薬を投与することにより、表1に掲げた障害を始めとする(しかし限定さ
れることはない)新生物または悪性状態への進行を予防することができる。そのような投
与は、その治療薬が上記した検定において前記のような障害の処置または予防に有用であ
ることが示された場合に指示される。このような予防用途は、新生物または癌に進行する
ことが知られているかまたは疑われる条件で、特に、過形成、化生、または最も特定的に
は異形成から成る非新生物細胞増殖が起こっている場合に指示されるものである(このよ
うな異常増殖条件の概論についてはRobbins and Angell,1976,Basic Pathology,2d Ed
.,W.B.Saunders Co.,Philadelphia,pp.68-79を参照されたい)。過形成は、構造ま
たは機能を大きく変えることなく、ある組織または器官において細胞数を増大させる調節
された細胞増殖のひとつの形態である。一例として、子宮内膜過形成は子宮内膜癌に先行
することが多い。化生(異形成)は、あるタイプの成体または充分に分化した細胞が別の
タイプの成体細胞の替わりとなる調節された細胞増殖のひとつの形態である。化生(異形
成)は上皮または結合組織細胞で起こり得る。典型的でない化生(異形成)には多少乱れ
た化生上皮が関与する。異形成はしばしば癌の前兆であり、主に上皮で見られる。異形成
は、個々の細胞一様性および細胞の構成的配向に損失がある非新生物細胞増殖の最も乱れ
た形態である。異形成細胞は異常に大きく深く染色した核をもっていることが多く、多形
性を示す。異形成は、慢性過敏状態または炎症が存在する場合に特徴的に起こり、頸管、
呼吸路、口腔および胆嚢で見られることが多い。
【0128】
あるいは、または、過形成、化生もしくは異形成として特徴付けられる異常な細胞増殖
の存在に加えて、患者から得た細胞サンプルがin vivoで示すかまたはin vitroで示す形
質転換表現型または悪性表現型の1つ異常な特徴が存在することは、本発明の治療薬の予
防的/治療的投与の望ましさを示し得る。上記したように、形質転換表現型のそのような
特徴としては、形態変化、より緩い基層付着、接触阻害の損失、足場依存性の損失、プロ
テアーゼの放出、糖輸送の増大、血清要求性の低下、胎児抗原の発現、250,000 ダルトン
の細胞表面タンパク質の消失、などがある(形質転換または悪性表現型に伴なう特性に関
しては前記文献の84〜90頁も参照されたい)。
【0129】
特定の態様において、白斑症、上皮の良性に出現する過形成もしくは異形成損傷、また
はボウエン病、insitu癌腫は予防的介入の望ましいことを示す腫瘍発現前の損傷である
。
【0130】
別の態様においては、線維嚢胞症(嚢胞性過形成、乳房異形成、特に腺疾患(良性上皮
過形成))が予防的介入の望ましいことを示す。
【0131】
他の態様においては、悪性に対する以下の素質的要因の1つ以上を示す患者を有効量の
治療薬の投与によって処置する。悪性に伴なう染色体転座(たとえば、慢性骨髄性白血病
に対するフィラデルフィア染色体、毛包性リンパ腫に対するt(14;18)、など)、
家族性ポリープ症もしくはガードナー症候群(結腸癌の可能な前兆)、良性のモノクロー
ナルγグロブリン異常(多発性骨髄腫の可能な前兆)、ならびにメンデルの(遺伝)遺伝
パターンを示す癌または前癌疾患(たとえば、結腸の家族性ポリープ症、ガードナー症候
群、遺伝性外骨症、多内分泌腺腫症、アミロイドを産生する髄様甲状腺癌および褐色細胞
腫、ポッツ‐ジェガース症候群、フォン・レックリングハウゼンの神経線維腫症、網膜芽
腫、頸動脈小体腫瘍、皮膚黒色癌、眼内黒色癌、色素性乾皮症、毛細管拡張性運動失調、
チェディアック‐東症候群、白子症、ファンコニ無形成貧血、およびブルーム症候群。Ro
bins and Angell,1976,BasicPathology,2d Ed.,W.B.Saunders Co.,Philadelphia
,pp.112-113参照)など)をもつ人との第一級の血族関係。
【0132】
別の特定の態様においては、本発明のアンタゴニスト治療薬をヒトの患者に投与して乳
癌、結腸癌または頸管癌への進行を予防する。
【0133】
5.9.2.他の障害
他の態様においては、本発明の治療薬を投与して、5.8.2節に記載した神経系の障
害または5.8.3節に記載した他の障害(たとえば、肝硬変、乾癬、ケロイド、禿頭)
を予防することができる。
【0134】
5.10.治療または予防上の有用性の立証
本発明の治療薬の所望の治療または予防活性はin vivoで試験することができる。たと
えば、そのような化合物は、ヒトに試験する前に、ラット、マウス、ニワトリ、ウシ、サ
ル、ウサギなどを始めとする(ただしこれらに限られるわけではない)適切な動物モデル
系で試験することができる。ヒトに投与する前のin vivoの試験には業界で公知のいかな
る動物モデル系も使用できる。
【0135】
5.11.Delta発現のアンチセンス調節
本発明は、デルタまたはその一部をコードする遺伝子またはcDNAに対してアンチセ
ンスである少なくとも6個のヌクレオチドの核酸の治療または予防上の用途を提供する。
本明細書中で「アンチセンス」とは、なんらかの配列相補性によってDeltaRNA(好ま
しくはmRNA)の一部とハイブリダイズすることができる核酸を指す。そのようなアン
チセンス核酸は本発明のアンタゴニスト治療薬として有用であり、前記5.8節およびそ
れに続く小節に記載したような障害の処置または予防に使用することができる。
【0136】
本発明のアンチセンス核酸は二本鎖または一本鎖のオリゴヌクレオチド、RNAもしく
はDNAまたはこれらの修飾体もしくは誘導体であることができ、これらは直接細胞に投
与することができるし、あるいは外来の導入された配列の転写によって細胞内で産生する
ことができる。
【0137】
ひとつの特定の態様の場合、本発明によって提供されるDeltaアンチセンス核酸は、そ
の細胞がDelta遺伝子またはNotch遺伝子を発現することを(invitroまたはin vivoで)
立証することができる腫瘍その他の障害の処置に使用することができる。そのような立証
はRNAまたはタンパク質の検出で行なうことができる。
【0138】
本発明はさらに、5.12節で後述するように製剤上許容可能な担体中に有効量の本発
明のDeltaアンチセンス核酸を含む薬剤組成物を提供する。また、本発明の薬剤組成物を
投与することからなる、(5.8および5.9節に記載したような)障害を処置および予
防する方法も提供される。
【0139】
別の態様において、本発明は原核生物または真核生物の細胞中におけるDelta核酸配列
の発現を抑制する方法に関しており、この方法は本発明のアンチセンスDelta核酸を含む
組成物を有効量細胞に与えることからなる。
【0140】
Deltaアンチセンス核酸とその用途は以下に詳細に記載する。
【0141】
5.11.1.Deltaアンチセンス核酸
Deltaアンチセンス核酸はヌクレオチドが少なくとも6個のものであり、オリゴヌクレ
オチド(6〜約50個の範囲のオリゴヌクレオチド)であるのが好ましい。特定の態様の
場合、オリゴヌクレオチドは少なくとも10個のヌクレオチド、少なくとも15個のヌク
レオチド、少なくとも100個のヌクレオチド、または少なくとも200個のヌクレオチ
ドである。オリゴヌクレオチドはDNAもしくはRNAまたはそれらのキメラ混合物もし
くは誘導体もしくは修飾体であり得り、一本鎖または二本鎖であり得る。このオリゴヌク
レオチドはその塩基部分、糖部分またはリン酸基骨格で修飾することができる。またこの
オリゴヌクレオチドはペプチド、または細胞膜を通過する輸送を容易にするもの(たとえ
ば、Letsingeret al.,1989,Proc.Natl.Acad.Sci.U.S.A.86:6553-6556; Lemaitre
et al.,1987,Proc.Natl.Acad.Sci.84:648-652;1988年12月15日に公開さ
れたPCT公開第88/09810号参照)、または血液脳関門(たとえば、1988年4月25日
に公開されたPCT公開第89/10134号参照)、ハイブリダイゼーションで誘発される開裂剤
(たとえば、Krolet al.,1988,BioTechniques 6:958-976参照)、またはインターカレ
ート剤(たとえば、Zon,1988,Pharm.Res.5:539-549参照)のような他の付随する基を
含んでいてもよい。
【0142】
本発明の好ましい態様では、好ましくは一本鎖DNAからなるDeltaアンチセンスオリ
ゴヌクレオチドが提供される。最も好ましい態様の場合、そのようなオリゴヌクレオチド
は、デルタ、最も好ましくはヒトデルタのSH3結合ドメインまたはノッチ結合ドメイン
をコードする配列とアンチセンスの配列からなる。このオリゴヌクレオチドはその構造上
の任意の位置において業界で一般に知られている置換基により修飾されていてもよい。
【0143】
デルタアンチセンスオリゴヌクレオチドは、5‐フルオロウラシル、5‐ブロモウラシ
ル、5‐クロロウラシル、5‐ヨードウラシル、ヒポキサンチン、キサンチン、4‐アセ
チルシトシン、5‐(カルボキシヒドロキシルメチル)ウラシル、5‐カルボキシメチル
アミノメチル‐2‐チオウリジン、5‐カルボキシメチルアミノメチルウラシル、ジヒド
ロウラシル、β‐D‐ガラクトシルキューオシン、イノシン、N6‐イソペンテニルアデ
ニン、1‐メチルグアニン、1‐メチルイノシン、2,2‐ジメチルグアニン、2‐メチ
ルアデニン、2‐メチルグアニン、3‐メチルシトシン、5‐メチルシトシン、N6‐ア
デニン、7‐メチルグアニン、5‐メチルアミノメチルウラシル、5‐メトキシアミノメ
チル‐2‐チオウラシル、β‐D‐マンノシルキューオシン、5′‐メトキシカルボキシ
メチルウラシル、5‐メトキシウラシル、2‐メチルチオ‐N6‐イソペンテニルアデニ
ン、ウラシル‐5‐オキシ酢酸(v)、ワイブトキソシン(wybutoxosine)、シュードウラ
シル、キューオシン、2‐チオシトシン、5‐メチル‐2‐チオウラシル、2‐チオウラ
シル、4‐チオウラシル、5‐メチルウラシル、ウラシル‐5‐オキシ酢酸メチルエステ
ル、ウラシル‐5‐オキシ酢酸(v)、5‐メチル‐2‐チオウラシル、3‐(3‐アミ
ノ‐3‐N‐2‐カルボキシプロピル)ウラシル、(acp3)w、および2,6‐ジア
ミノプリンを包含する(しかし限定されることはない)群の中から選択される少なくとも
1つの修飾された塩基部分を含んでいてもよい。
【0144】
別の態様の場合、オリゴヌクレオチドは、アラビノース、2‐フルオロアラビノース、
キシルロースおよびヘキソースを包含する(しかしこれらに限定されない)群の中から選
択される少なくとも1つの修飾された糖部分を含んでいる。
【0145】
さらに別の態様の場合、オリゴヌクレオチドは、ホスホロチオエート、ホスホロジチオ
エート、ホスホルアミドチオエート、ホスホルアミデート、ホスホルジアミデート、メチ
ルホスホネート、アルキルホスホトリエステルおよびホルムアセタールまたはこれらのア
ナログから成る群の中から選択される少なくとも1つの修飾されたリン酸基骨格を含んで
いる。
【0146】
さらに別の態様ではオリゴヌクレオチドがα‐アノマーオリゴヌクレオチドである。α
‐アノマーオリゴヌクレオチドは相補的RNAと共に特定の二本鎖ハイブリッドを形成す
る。この場合、通常のβ‐単位とは対照的に、ストランドは互いに平行になっている(Gau
tier et al.,1987,Nucl.AcidsRes.15:6625-6641)。
【0147】
オリゴヌクレオチドは別の分子、たとえばペプチド、ハイブリダイゼーションで誘発さ
れる架橋剤、輸送剤、ハイブリダイゼーションで誘発される開裂剤などと結合していても
よい。
【0148】
本発明のオリゴヌクレオチドは業界で公知の標準的な方法によって、たとえば自動DN
Aシンセサイザー(たとえば、Biosearch,Applied Biosystemsなどから市販されている
ものなど)を使用することによって合成できる。例として、ホスホロチオエートオリゴヌ
クレオチドはSteinらの方法(1988,Nucl.AcidsRes.16:3209)によって合成でき、メチ
ルホスホネートオリゴヌクレオチドは調節された多孔ガラスポリマー支持体(Sarin et al
.,1988,Proc.Natl.Acad.Sci.U.S.A.85:7448-7451)等を使用することによって調製
することができる。
【0149】
特定の態様の場合、Deltaアンチセンスオリゴヌクレオチドは触媒RNAすなわちリボ
ザイムを含んでいる(たとえば、1990年10月4日に公開されたPCT公開第90/11364
号、Sarveret al.,1990,Science 247:1222-1225参照)。別の態様ではオリゴヌクレオ
チドが2′‐O‐メチルリボヌクレオチド(Inoue et al.,1987,Nucl.Acids Res.15:6
131-6148)またはキメラRNA−DNAアナログ(Inoueet al.,1987,FEBS Lett.215:3
27-330)である。
【0150】
他の態様の場合、本発明のDeltaアンチセンス核酸は外来配列からの転写によって細胞
内で産生される。たとえば、あるベクターをin vivoで導入して、そのベクターを細胞に
取込ませ、その細胞の中でそのベクターまたはその一部が転写されて本発明のアンチセン
ス核酸(RNA)を産生するようにすることができる。そのようなベクターはDeltaアン
チセンス核酸をコードする配列を含有しているであろう。そのようなベクターは、転写さ
れて所望のアンチセンスRNAを産生することができる限りにおいて、エピソームのまま
でいることもできるし、あるいは染色体に組込まれるようになってもよい。そのようなベ
クターは業界で標準的な組換えDNA技術によって構築することができる。ベクターは哺
乳動物細胞で複製および発現に使われているプラスミド、ウィルスその他業界で公知のも
のでよい。デルタアンチセンスRNAをコードしている配列の発現は哺乳動物、好ましく
はヒトの細胞で作用することが業界で知られている任意のプロモーターによって行なうこ
とができる。そのようなプロモーターは誘導的または構成的のいずれであることもできる
。そのようなプロモーターとしては、SV40の初期プロモーター領域(Bernoist and Ch
ambon,1981,Nature290:304-310)、ラウス肉腫ウィルスの3′の長い末端反復配列に含
まれているプロモーター(Yamamotoet al.,1980,Cell 22:787-797)、ヘルペスチミジン
キナーゼプロモーター(Wagner et al.,1981,Proc.Natl.Acad.Sci.U.S.A.78:1441-
1445)、メタロチオネイン遺伝子の調節配列(Brinsteret al.,1982,Nature 296:39-42)
、などがあるがこれらに限定されることはない。
【0151】
本発明のアンチセンス核酸はDelta遺伝子、好ましくはヒトDelta遺伝子のRNA転写物
の少なくとも一部に相補的な配列を含んでいる。しかし、好ましいとはいっても完全な相
補性は必要とされない。本明細書で「RNAの少なくとも一部に相補的な」配列とは、そ
のRNAとハイブリダイズして安定な二重鎖(duplex)を形成することができるほどに充分
な相補性をもっている配列を意味している。二本鎖のデルタアンチセンス核酸の場合には
その二重鎖DNAの単一のストランドをそのように試験してもよいし、あるいは三重鎖(t
riplex)の形成を検定してもよい。ハイブリダイズする能力は相補性の程度とアンチセン
ス核酸の長さの双方に依存する。一般に、ハイブリダイズする核酸が長くなればなるほど
、それが含有し得るDeltaRNAとの塩基のミスマッチが多くなるがそれでも安定な二重
鎖(または場合によって三重鎖)を形成する可能性が高くなる。当業者は、標準的な手法
を使用して、ハイブリダイズした複合体の融点を決定するミスマッチの許容できる程度を
確認することができる。
【0152】
5.11.2.Deltaアンチセンス核酸の治療上の有用性
Deltaアンチセンス核酸を使用して、DeltaまたはNotchを発現することがわかっている
細胞型の悪性腫瘍またはその他の障害を処置(または予防)することができる。特定の態
様の場合悪性は頸管癌、乳癌もしくは結腸癌または扁平腺癌である。そのような発現を試
験することができる悪性の、腫瘍(新生物)および前新生物細胞としては前記5.8.1
および5.9.1節に記載したものがあるがこれらに限定されることはない。好ましい態
様においては一本鎖のDNAアンチセンスデルタオリゴヌクレオチドを使用する。
【0153】
DeltaまたはNotchRNAを発現する悪性(特に腫瘍)細胞型は業界で公知の各種方法で
同定することができる。そのような方法としては、DeltaまたはNotch特異的核酸とのハイ
ブリダイゼーション(たとえばノーザンハイブリダイゼーション、ドットブロットハイブ
リダイゼーション、insituハイブリダイゼーション)、その細胞型由来のRNAがin vi
troでノッチまたはデルタに翻訳され得る能力を観察すること、免疫検定、などがあるが
これらに限られるわけではない。好ましい態様においては、処置前に、患者から得た一次
腫瘍組織のノッチまたはデルタの発現を、たとえば免疫細胞化学またはin situハイブリ
ダイゼーションによって検定することができる。
【0154】
NotchもしくはDeltaRNAまたはタンパク質を発現するタイプの悪性を有する患者に、
薬剤として許容可能な担体中に有効量のDeltaアンチセンス核酸を含む本発明の薬剤組成
物(5.12節参照)を投与することができる。
【0155】
特定の障害または状態の処置に有効であるDeltaアンチセンス核酸の量はその障害また
は状態の性質に依存しており、標準的な臨床技術によって決定することができる。可能な
場合、ヒトで試験して使用する前に、処置すべき腫瘍タイプのアンチセンス細胞傷害性を
、in vitroで、その後、有用な動物モデル系で、決定するのが望ましい。
【0156】
特定の態様においては、Deltaアンチセンス核酸を含む薬剤組成物をリポソーム、微粒
子、またはマイクロカプセルを介して投与する。本発明のさまざまな態様においては、そ
のような組成物を用いてDeltaアンチセンス核酸の持続放出を達成するのが有用であり得
る。特定の態様の場合、特定の同定可能な腫瘍抗原に対する抗体を介して標的に向かわし
めるリポソームを利用するのが望ましいかもしれない(Leonetti et al.,1990,Proc.Na
tl.Acad.Sci.U.S.A.87:2448-2451;Renneisen et al.,1990,J.Biol.Chem.265:1
6337-16342)。
【0157】
5.12.治療/予防用投与および組成物
本発明は、本発明の治療薬を有効量で被検体に投与することによる処置(および予防)
の方法を提供する。好ましい局面においては、この治療薬は実質的に精製されている。被
検体は、牛、豚、鶏などのような動物をはじめとする(しかしこれらに限定されない)動
物が好ましく、哺乳類が好ましく、ヒトが最も好ましい。
【0158】
各種の送達系が知られており、本発明の治療薬を投与するために使用することができる
。たとえば、リポソーム、微粒子、マイクロカプセルに封入する、組換え細胞によって発
現させる、受容体に媒介されるエンドサイトーシス(たとえば、Wu and Wu,1987,J.Bi
ol.Chem.262:4429-4432参照)、レトロウィルスまたはその他のベクターの一部として
治療薬の核酸を構築する、などである。導入方法としては、皮内、筋肉内、腹膣内、静脈
内、皮下、鼻腔内、硬膜、および経口経路があるがこれらに限定されない。化合物は便利
な任意の経路、たとえば、注入または濃縮塊注射により、上皮もしくは粘膜皮膚ライニン
グ(lining)(たとえば、口腔粘膜、直腸および腸粘膜、など)を通る吸収により投与可能で
、他の生物活性剤と共に投与してもよい。投与は全身的または局所的に行うことができる
。また、本発明の薬剤組成物を適切な経路、たとえば心室内および鞘内注射によって中枢
神経系に導入することが望ましいことがある。心室内注射は、心室内カテーテル、たとえ
ばOmmayaリザーバー(reservoir)のようなリザーバーに接続した心室内カテーテルによっ
て容易にすることができる。また、たとえば吸入器や噴霧器およびエアゾール剤を用いた
製剤を使用することによって、肺投与も使用できる。
【0159】
特定の態様では、処置が必要とされる部分に本発明の薬剤組成物を局所的に投与するの
が望ましいことがある。これは、たとえば(限定ではないが)、手術中の局所注入、たと
えば手術後の創傷包帯と共にする局所適用、注射、カテーテル、座薬、インプラント(こ
れは多孔質、非多孔質またはゼラチン状の材料、たとえばsialastic膜のような膜、また
は線維である)を用いて達成できる。ひとつの態様では、悪性腫瘍または腫瘍もしくは腫
瘍発現前の組織の部位(または前の部位)に直接注射することによって投与することがで
きる。
【0160】
別の態様では、治療薬を小胞、特にリポソームに入れて送達することができる(Langer
,Science249:1527-1533(1990); Treatら,in Liposomes in the Therapy of Infectiou
s Disease andCancer,Lopez-BeresteinおよびFidler(eds.),Liss,New York,pp.353
-365(1989);Lopez-Berestein,ibid.,pp.317-327;参照。概論としてはibid.参照)。
【0161】
さらに別の態様の場合治療薬は制御放出系で送達することができる。ひとつの態様では
ポンプが使用できる(Langer,上掲;Sefton,CRC Crit.Ref.Biomed.Eng.14:201(1987
); Buchwaldら,Surgery88:507(1980); Saudek ら,N.Engl.J.Med.321:574(1989参
照))。別の態様ではポリマー性の材料を使用することができる(Medical Applications
ofControlled Release,LangerおよびWise(eds.),CRC Pres.,Boca Raton,Florida(1
974);Controlled Drug Bioavailability,Drug Product DesignおよびPerformance,Smo
len andBall(eds.),Wiley,New York(1984); RangerおよびPeppas,J.Macromol.Sci
.Rev.Macromol.Chem.23:61(1983)参照。またLevyら,Science228:190(1985); Durin
gら,Ann.Neurol.25:351(1989);Howardら,J.Neurosurg.71:105(1989)も参照のこ
と)。さらに別の態様においては、制御放出系を治療標的、すなわち脳の近くに配置する
ことができ、従って全身投与量の一部しか必要としない(たとえば、Goodson,in Medica
l Applicationsof Controlled Release,上掲,vol.2,pp.115-138(1984)参照)。
【0162】
他の制御放出系がLangerの文献によって議論されている(Science249:1527-1533(1990)
)。
【0163】
治療薬がタンパク質の治療薬をコードしている核酸である特定の態様の場合、その核酸
をin vivoで投与してそれがコードしているタンパク質の発現を促進することができる。
たとえば、レトロウィルスベクター(米国特許第4,980,286号参照)を使用することによ
り、その核酸を適当な核酸発現ベクターの一部として構築し、それを細胞内に組込まれる
ようにして投与する。または直接注射したり、微粒子衝撃(たとえば遺伝子銃(gun)、Bio
listic,Dupont)を使用したり、または脂質もしくは細胞表面受容体もしくはトランスフ
ェクト剤で被覆したり、または核の中に入って行くことが知られているホメオボックス様
ペプチドに結合させて投与したりする(たとえば、Joliotら,1991,Proc.Natl.Acad.
Sci.USA88:1864-1868)。あるいはまた、核酸治療薬を細胞内に導入し、相同組換えに
よって発現のために宿主細胞DNA内部に組込ませることができる。
【0164】
特定の障害の処置または予防に関する特定の態様の場合、次の投与形態を使用するのが
好ましい。
【0165】
【0166】
本発明はまた薬剤組成物も提供する。そのような組成物は、治療上有効な量の治療薬と
、薬剤として許容される担体とを含んでいる。特定の態様の場合、「薬剤として許容され
る」という用語は、動物、特にヒトに使用するものとして、米国の連邦政府もしくは州政
府の規制機関によって認可されているかまたは米国薬局方もしくは他の一般に容認されて
いる薬局方に収載されていることを意味する。「担体」という用語は、治療薬を投与する
ときに一緒に用いる希釈剤、アジュバント、賦形剤またはビヒクルを指す。そのような薬
剤担体は、水および石油、動物、植物もしくは合成起源のものを始めとする油、たとえば
ピーナッツ油、大豆油、鉱物油、ごま油などのような無菌の液体でありうる。薬剤組成物
を静脈内に投与するときには水が好ましい担体である。生理食塩水ならびにデキストロー
スおよびグリセロール水溶液もまた、特に注射用溶液の液体担体として使用することがで
きる。適切な薬剤賦形剤としては、澱粉、グルコース、ラクトース、スクロース、ゼラチ
ン、麦芽、米、小麦粉、チョーク、シリカゲル、ステアリン酸ナトリウム、モノステアリ
ン酸グリセロール、タルク、塩化ナトリウム、乾燥スキムミルク、グリセロール、プロピ
レン、グリコール、水、エタノールなどがある。所望により、該組成物はまた、少量の湿
潤剤もしくは乳化剤、またはpH緩衝剤も含有することができる。これらの組成物は溶液
、懸濁液、エマルション、錠剤、ピル、カプセル、粉末、徐放性製剤などの形態をとるこ
とができる。組成物は通常のバインダーおよびトリグリセリドのような担体と共に座薬と
して調合することができる。経口製剤は薬剤グレードのマンニトール、ラクトース、澱粉
、ステアリン酸マグネシウム、サッカリンナトリウム、セルロース、炭酸マグネシウムな
どのような標準的な担体を含むことができる。適した薬剤担体の例はE.W.Martinによっ
て「Remington´sPharmaceutical Sciences」に記載されている。このような組成物は患
者に適切な投与形態とするのに適した量の担体と共に治療上有効量の治療薬、好ましくは
精製形態の治療薬を含有している。この製剤は投与形態に適していなければならない。
【0167】
好ましい態様の場合、組成物は、通常の手法に従って、人間に静脈内投与するのに適し
た薬剤組成物として調合する。通常静脈内投与用の組成物は無菌の等張水性緩衝液の溶液
である。必要であれば組成物は、可溶化剤および注射部位の痛みを和らげるリグノカイン
のような局所麻酔薬とを含んでいてもよい。一般に、成分は別々に、または単位投与形態
として、たとえば活性剤の量を指示したアンプルまたはサシェットのような気密に密閉し
た容器に入れた凍結乾燥粉末または無水の濃縮物として一緒に供給される。組成物を注入
によって投与しようとする場合、無菌の薬剤グレードの水または生理食塩水を含有する注
入瓶として分配することができる。組成物を注射によって投与する場合、成分が投与前に
混合されるように注射用の無菌水または生理食塩水のアンプルを提供することができる。
【0168】
本発明の治療薬は中性形態または塩形態として調合することができる。薬剤として許容
できる塩には、塩酸、リン酸、酢酸、シュウ酸、酒石酸などから誘導されるもののように
遊離のアミノ基との間で形成されるもの、および水酸化ナトリウム、水酸化カリウム、水
酸化アンモニウム、水酸化カルシウム、水酸化第二鉄、イソプロピルアミン、トリエチル
アミン、2‐エチルアミノエタノール、ヒスチジン、プロカインなどから誘導されるもの
のように遊離のカルボキシル基との間に形成されるものがある。
【0169】
特定の障害または症状の処置に有効な本発明の治療薬の量はその障害または症状の性質
に依存し、標準的な臨床技術によって決定することができる。また、場合によって、in v
itro検定を利用して最適の投与範囲を同定するのに役立てることができる。本製剤中に使
用するべき正確な量もまた投与経路およびその疾患または障害の重さに依存し、医者の判
断および各患者の状況に応じて決定するべきである。しかし、静脈内投与に適した投与範
囲は一般に体重1kgあたり活性化合物約20〜500μgである。鼻腔内投与に適した投
与範囲は一般に体重1kgあたり約0.01pg〜1mgである。有効な投与量はin vitroまた
は動物モデル試験系から得られる用量−反応曲線から推定すればよい。
【0170】
座薬は一般に0.5〜10重量%の範囲で活性成分を含有し、経口製剤は活性成分を1
0〜95%含有するのが好ましい。
【0171】
本発明はまた、本発明の薬剤組成物の成分を1以上充填した1以上の容器を含む薬剤パ
ックまたはキットも提供する。場合によって、そのような容器には、薬剤または生物学的
生成物の製造、使用または販売を規制する政府機関によって規定された形の注意書きを付
けることができる。そのような注意書きは人間への投与に対する製造、使用または販売の
認可を示すものである。
【0172】
5.13.診断用途
デルタタンパク質、類似体、誘導体、およびこれらのサブ配列、Delta核酸(およびそ
れに相補的な配列)、抗デルタ抗体には診断用途がある。このような分子は、免疫検定の
ような検定法で使用して、デルタ発現に影響する各種症状、疾患および障害を検出、予知
、診断、もしくは追跡し、またはその処置を追跡することができる。特に、そのような免
疫検定は、患者から得たサンプルを免疫特異的な結合が起こり得るような条件下で抗デル
タ抗体と接触させ、その抗体による任意の免疫特異的結合の量を測定することからなる方
法によって実施する。特定の局面においては、組織切片における、抗体のそのような結合
を、好ましくは抗ノッチ抗体の結合と組合せて用いて、ある疾患状態における異常なノッ
チおよび/またはデルタ局在化または異常なレベルのノッチ−デルタの同時局在化を検出
することができる。ある特定の態様においては、デルタに対する抗体を用いて、患者の組
織または血清サンプルで、デルタの存在を検定することができ、ここで異常なレベルのデ
ルタは疾患状態の指標となる。内因性のノッチタンパク質におけるデルタ結合能力の異常
なレベル、または内因性のデルタタンパク質におけるノッチ(または他のデルタリガンド
)に対して結合する能力の異常なレベルは、細胞運命の障害(たとえば癌など)の指標と
なり得る。「異常なレベル」とは、その身体の一部から、またはその障害をもたない被検
者から得た類似のサンプル中に存在するレベルまたはこのレベルを表わす標準レベルと比
べて増大したかもしくは低下したレベルを意味する。
【0173】
使用することができる免疫検定には、ウェスタンブロット、放射線免疫検定、ELIS
A(固相酵素免疫検定)、「サンドイッチ」免疫検定、免疫沈降検定、沈降反応、ゲル核
酸沈降反応、免疫核酸検定、凝集検定、補体結合検定、イムノラジオメトリックアッセイ
、免疫蛍光検定、プロテインA免疫検定、などのような技術を用いる競合検定系および非
競合検定系があるがこれらに限定されるわけではない。
【0174】
Delta遺伝子および関連する核酸配列およびサブ配列、たとえば相補的配列、ならびに
その他の局所周期性遺伝子配列もまたハイブリダイゼーションアッセイに使用することが
できる。Delta核酸配列、または少なくとも約8個のヌクレオチドを含むそのサブ配列は
ハイブリダイゼーションプローブとして使用することができる。ハイブリダイゼーション
アッセイは前記したようにデルタの発現および/または活性の異常な変化を伴なう症状、
障害または疾患状態を検出、予知、診断または追跡するのに使用することができる。特に
、そのようなハイブリダイゼーションアッセイは、核酸を含有するサンプルを、ハイブリ
ダイゼーションが起こり得る条件下で、DeltaDNAまたはRNAとハイブリダイズする
ことができる核酸プローブと接触させ、その結果生じるハイブリダイゼーションを検出ま
たは測定することからなる方法によって実施する。
【0175】
さらに、デルタはノッチと結合するので、デルタまたはその結合部分は、サンプル中の
ノッチの存否および/またはその量を検定するのに、たとえば、結腸癌や子宮頸癌のよう
な増大したノッチ発現を示す悪性腫瘍をスクリーニングするのに、使用することができる
。
【実施例】
【0176】
6.ヒヨコのDeltaホモログは予定ニューロンで発現する
本明細書に記載したように、本発明者らはヒヨコのDeltaホモログ、C-Delta-1を単離し
特性決定をした。本発明者らは、C-Delta-1が、神経組織発生の間に新しい細胞が生まれ
、その運命が決定されるときに予定ニューロンで発現されることを示す。ヒヨコにおける
本発明者らのデータは、デルタ/ノッチシグナリングメカニズムと神経組織発生における
その役割とは両方とも脊椎動物で保存されていることを示唆している。
【0177】
6.1.C-Delta-1のクローニング
本発明者らは、ポリメラーゼ連鎖反応(PCR)と縮重したオリゴヌクレオチドプライ
マー(図1a、1bおよび2、配列番号1、2、3および4)を用いてヒヨコのDeltaホ
モログ(C-Delta-1)を同定した。それぞれハエデルタタンパク質配列FGFTWPGT(配列番
号21)およびNGGTCID(配列番号22)に相当する縮重したオリゴヌクレオチドプライ
マーTTCGGITT(C/T)ACITGGCCIGGIAC(配列番号19)およびTCIATGCAIGTICCICC(A/G)TT(
配列番号20)を用いたPCRによってC-Delta-1をクローン化した(Vassinら,1987,EM
BO J.6:3431-3440;Kopczynskiら,1988,Genes Dev.2:17 23-1735)。最初の反応では
、50μlの供給緩衝液(Perkin-Elmer)中で、4〜6胚期に由来する第一鎖オリゴ‐d(
T)でプライムしたcDNA50ng、各プライマー1μg、0.2mMdNTP、2UのT
aqポリメラーゼを使用した。94℃/30秒、50℃/2分、72℃/2分で40サイ
クルの増幅を行なった。増幅したDNA断片をアガロースゲルで分離し、Bluescript pKS
-(Stratagene)でクローン化し、配列決定した。2つのDeltaホモログが同定された。その
うちのひとつ(C-Delta-1)は神経系で発現される。神経系で発現されるホモログで、C-D
elta-1遺伝子の3′端における交互スプライシング事象のために、コードされているタン
パク質のカルボキシ末端が異なる2つの変異体が同定された。コードされているタンパク
質のひとつは、他方のコードされているタンパク質と比べて、カルボキシ末端に12個の
余分なアミノ酸を有する。このコードされている短い方の変異体の配列を配列番号2に示
す。配列番号3によりコードされており、かつ配列番号4のアミノ酸配列で同定される長
い方の変異体は配列番号2のアミノ酸配列と3′端にある12個の余分なアミノ酸(SIPPG
SRTSLGV)から成っている。この長い方の変異体を以下に記載する実験に用いた。RNAを
アフリカツメガエル(Xenopusoocytes)に注入して生物活性を試験した場合変異体のいず
れも同じ生物活性をもっていた。
【0178】
C-Delta-1に対応するDNA断片を用いて、17期の脊髄cDNAライブラリーをスク
リーニングし、いくつかの完全長クローンを得、配列決定した。類似の方法によりヒヨコ
のC-Notch-1遺伝子に由来するDNA断片を増幅した(データは示さない)。部分配列デ
ータと発現パターンは齧歯類のノッチ−1遺伝子との密接な類似性を示している(Weinmas
terら,1991,Development113:199-205; Weinmasterら,1992,Development 116:931-94
1; Lardelli& Lendahl,1993,Exp.Cell Res.204:364-372)。配列はWisconsin GCGセ
ットプログラムを用いて解析した。ヒヨコデルタ1mRNAのGenBank受託番号はU265
90である。C-Delta-1のDNA配列はショウジョウバエデルタと構造的に同族である(
図3、4)がデルタ関連Serrateタンパク質(これも本発明者らがクローン化したもので
あるがデータは示さない)の脊椎動物ホモログとは明らかに異なる722個のアミノ酸の
タンパク質に相当している。C-デルタ-1はその細胞外領域に推定膜貫通ドメイン、シグナ
ル配列および8つのEGF様反復配列を含有している(ショウジョウバエデルタより反復
配列が1つ少ない)。C-デルタ-1のアミノ末端ドメインはノッチに対するin vitro結合に
必要かつ充分であると記載されている(Muskavitch,1994,Dev.Biol.166:415-430)ハエ
のデルタタンパク質中の類似のドメインと密接に関連している。この保持された領域は、
ノッチ様タンパク質の推定リガンドのファミリーの公知のすべてのメンバーにより共有さ
れている(Deltaand Serrate in Drosophila; Lag-2 and Apx-1 in Caenorhabditis eleg
ans)(Hendersonら,1994,Development120:2913-2924; Taxら,1994,Nature 368:150-1
54; Flemingら,1990,GenesDev.4:2188-2201; Thomasら,1991,Development 111:749
-761; Melloら,1994,Cell77:95-106)いわゆるDSLモチーフ(図4)(Hendersonら
,1994,Development120:2913-2924; Tax ら,1994,Nature 368:150-154)を含んでい
る。二番目のシステインに富むN末端領域はハエとヒヨコのタンパク質で保存されている
が、関連するC.elegansタンパク質にはない(図4)。C-デルタ-1と81%同一であり上
記の構造モチーフのすべてを示す(図3)タンパク質をコードしているアフリカツメガエ
ルDelta-1ホモログX-Delta-1もクローン化した。ノッチ結合に重要であると同定されたド
メイン(Mustavitch,1994,Dev.Biol.166:415-430)を含めてヒヨコとハエのデルタタン
パク質間の保存構造は、C-デルタ-1がヒヨコのノッチタンパク質に対するリガンドとして
機能すること、および外側抑制(lateral inhibition)のデルタ/ノッチに媒介されるメカ
ニズムがヒヨコで働き得ることを示唆している。
【0179】
6.2.C-Delta-1とC-Notch-1 の発現は神経組織発生開始と相関している
ショウジョウバエの神経組織発生の間にDeltaは神経前駆体で一時的に発現され、隣接
するNotch発現細胞が同様に神経になるのを抑制する(Haenlinら,1990,Development110
:905-914; Kooh ら,1993,Development117:493-507)。C-Delta-1がヒヨコの神経組織発
生の間に同様に働くとすると、ニューロン前駆体細胞が決定されるようになるにつれてC-
Delta-1が同様にニューロン前駆体細胞でも一時的に発現されるはずである。発生するC
NSにおけるC-Delta-1の発現の解析は、これが実際に起こっていることを示している。
【0180】
要約すると、ホールマウントin situハイブリダイゼーションを実施した。ホルムアル
デヒドで固定した胚をプロテアーゼで処理し、4%ホルムアルデヒド/0.1%グルタル
アルデヒドで再固定した。DIGで標識したRNAプローブとのハイブリダイゼーション
を、0.2%Tween-20と0.5%CHAPSを含有する緩衝液中で、ストリンジェントな
条件(1.5×SSC、50%ホルムアミド、65℃、pH5)下で実施した。洗浄した
胚をBoehringerBlocking Reagent(ベーリンガーブロッキング試薬)で処理し、アルカ
リホスファターゼと結合した抗‐DIG抗体中で一晩インキュベートした。充分に洗浄し
た後胚を30分〜一晩染色した。図5eの胚はハイブリダイゼーション後ワックスで切開
した(wax-sectioned)。
【0181】
神経板におけるC-Delta-1の発現は6〜7期(31時間、0/1体節)に体節形成前の
中胚葉の直前の散乱細胞(scattered cells)で最初に検出される(図5b、5c)。この
領域は中/後部(mid/posterior)後脳になり、そこでは、最も初期に分化したCNSニュ
ーロンが、最初のC-Delta-1発現の6時間後の9期(31時間、7〜9体節)に、神経繊
維抗体により始めて検出される(表2)(Sechrist & Bronner-Fraser,1991,Neuron 7:
947-963)。
【表2】
【0182】
神経組織発生が進むにつれて、C-Delta-1の発現がニューロン分化の時空パターンを予
示し続け(表2)、脊髄に沿って後に、また中脳および前脳に向かって前方に拡大する(
図5d、5e)。たとえば、8期の脊髄における最も後に発現する細胞は将来の6番目の
体節のレベルにあり、これはそのレベルにある最初のニューロンが神経繊維抗原を発現す
る6〜8時間前である(Sechrist & Bronner-Fraser,1991,Neuron 7:947-963)(表2)
。表2に示されているように、C-Delta-1の発現の出現は、最初のニューロン前駆体が分
裂サイクルから離脱したすぐ後で、得られる分化性ニューロンで神経繊維(NF)抗原が
出現する前である。
【0183】
中部後脳は耳の原基のレベルである菱脳の神経小片4〜6を含んでおり、後部後脳は菱
脳の神経小片7および8ならびに体節1〜4を含んでいる。細胞分裂からの離脱のタイミ
ングのデータと神経繊維の発現のデータはSechristら,1991,Neuron7:947-963からとっ
た。いずれの場合も、C-Delta-1は散乱細胞で一様なC-Notch-1発現のドメイン内で発現す
る(図5a)。
【0184】
6.3.C-Delta-1の局在化と経時発現
C-Delta-1の局在化と経時変化は、この遺伝子が神経組織発生の初期の段階で切り替え
られる(switched)こと、およびC-Delta-1を発現する細胞がまだ分化マーカーを示し始め
ていない将来のニューロンであることを示している。この仮説を試験するために、本発明
者らは、将来のニューロンが、初期神経管における非サイクル細胞(non-cycling cells)
のみであるというSechristとBronner-Fraserの観察結果(Sechrist& Bronner-Fraser,19
91,Neuron 7:947-963)を用いた。それらの細胞は神経繊維抗原を発現する11〜15時
間前にその最終S相を終了し(表2)、その核は、最後の有糸分裂の完了後、他の細胞核
すべてがS相にある神経上皮の基底表面付近に特徴的な位置をとる(Sechrist & Bronner-
Fraser,1991,Neuron7:947-963; Martin & Langman,1965,J.Embryol.Exp.Morphol
.14:23-35)(図6a)。本発明者らは7〜9期の胚をブロモデオキシウリジン(Brd
U)で標識し、BrdU取込みとC-Delta-1発現に関して二重に染色した。C-Delta-1を発
現する細胞の95%は未標識であった。それらの核は主として基底表面付近に位置してい
たが、そこではほとんどの他の核がBrdUで標識された(図6b、6c)。PBS中の
0.1mMのBrdU75μlを、insituハイブリダイゼーション用に固定する前に38
℃で2〜4時間インキュベートした7〜9期の胚上に滴下した。、15μmのクリオスタ
ット切片を、ほぼStrahleらの方法(Strahleら,1994,TrendsIn Genet.Sci.10:75-76)
に従って、DIGで標識したRNAプローブとハイブリダイズした。染色後スライドをP
BS中で洗浄し、BrdU免疫検出(immunodetection)用に処理した(Biffoら,1992,His
tochem.Cytochem.40:535-540)。FITCと結合したヤギ抗‐マウス二次抗体(Cappel)
を用いて抗BrdU(1:1000、Sigma)を検出した。C-Delta-1の発現をDIC顕微
鏡法、および慣用の共焦点蛍光顕微鏡法によるBrdU標識化で検査した。これらの結果
は、C-Delta-1が細胞サイクルから離脱しており、実際に将来のニューロンであるに違い
ない細胞で発現されることを示している。二、三のBrdU-/C-Delta-1-細胞は基底ゾ
ーン外にその核を有する。これらは、BrdUに暴露された後すぐに最終S相を終了し、
先端に移動してその最終有糸分裂を完了し、C-Delta-1発現に切り替わる細胞であり得る
。C-Delta-1はまた、後期の神経管および末消神経系でも発現する。ここでも、発現のタ
イミングと発現細胞の位置は、これらの細胞がまだ分化し始めていないニューロン前駆体
であることを示唆している(データは示さない)。このように、C-Delta-1の発現は将来
のニューロンに対する最も早く知られたマーカーであるように思われる。
【0185】
また、C-デルタ-1とC-Serrate-1の転写パターンは胚の多くの領域におけるC-ノッチ-1
の転写パターンとオーバーラップしており(データは示さない)、これはC-ノッチ-1がシ
ョウジョウバエ内のノッチと同様に両者のタンパク質に対する受容体であることを示唆し
ている。特に、3つのすべての遺伝子が発生する中枢神経系の神経原性の領域で発現され
、ここにおいて驚くべき関係が見られる。すなわち、C-Serrate-1とC-デルタ-1の両方の
発現はC-ノッチ-1発現のドメインに限定されるが、このドメイン内でC-Serrate-1とC-デ
ルタ-1の領域は正確に相補的である。オーバーラップする発現パターンは、それらとノッ
チとの機能的相関関係の保存を示唆しており、ヒヨコ、特にその中枢神経系の発生がいろ
いろな位置におけるC-ノッチ-1といろいろなリガンドとの相互作用を含んでいることを暗
示している。
【0186】
6.4.考察
C-デルタ-1のアフリカツメガエルホモログを同様な方法でクローン化した。簡単にいう
と、X-デルタ-1のPCR断片を単離して配列決定した。次にこの断片を用いてX-デルタ-1
の完全長クローンを同定した。X-デルタ-1の発現パターンを検討した。X-デルタ-1は散乱
細胞において、一次ニューロン前駆体が生成している神経板のドメイン内で発現すること
が示された。これは、X-デルタ-1を発現する細胞が将来の一次ニューロンであることを示
唆している。さらに、X-デルタ-1はまた、神経組織発生の他の部位および時期、たとえば
前部神経板および神経原性板ならびに二次ニューロンが生成した場合の神経管発生の後の
段階でも発現する。異所性のX-Delta-1活性は一次ニューロンの産生を抑制し、内因性のX
-Delta-1活性との干渉の結果一次ニューロンの過剰産生が生じた。これらの結果は、X-デ
ルタ-1が将来のニューロンによって隣接細胞に送達される側方抑制を媒介していることを
示している。アフリカツメガエル卵におけるX-デルタ-1の異所性発現は一次神経組織発生
を抑制すること、また、細胞質ドメインのアミノ酸を2つだけ保持している末端を切断し
た(truncated)X-デルタ-1タンパク質の異所性発現が内因性のシグナリングに干渉し、余
分な細胞をニューロン前駆体として発生させることが示された(Chitnisら,Nature(印刷
中))。予備的な証拠は、C-Delta-1がアフリカツメガエルの胚で発現されると類似の抑制
作用をもつことを示している(データは示さない)。本発明者らは、C-Delta-1がショウ
ジョウバエとアフリカツメガエルの対応するタンパク質と同様に神経組織発生の間を通し
て側方抑制を媒介して、任意の時に神経宿命になり得る細胞の割合を制限するということ
を提案する。C-デルタ-1は通常神経組織発生中CNSでもPNSでも多くの他の部位で、
たとえば発生する耳でも発現される。CNSにおいては、C-ノッチはE5ヒヨコ後脳の脳
室ゾーンにおいて神経管の内腔に隣接する分裂細胞で発現されることが示されている。C-
デルタ-1は、分裂を止めてしまっておりニューロン前駆体細胞となろうとしている細胞の
隣接層で発現する。したがって、デルタ/ノッチシグナリングはここにおいて他の神経組
織におけると同様に作用して未決のサイクルニューロン幹細胞の集団を維持することがで
きよう。
【0187】
7.マウスDeltaホモログの単離し特性決定
M-Delta-1と名付けたマウスDeltaホモログを以下のようにして単離した。
【0188】
マウスDelta-1
遺伝子由来組織:8.5日および9.5日齢のマウス胚RNA
単離法:
a)ランダムにプライムした、上記RNAに対するcDNA
b)上記cDNAのPCRには以下のものを使用した。
【0189】
PCRプライマー1:GGITTCACITGGCCIGGIACNTT(配列番号23)[デルタ様である
がSerrate様ではないタンパク質に対して特異的な領域であるGFTWPGTF(配列番号24)
をコードしている]
PCRプライマー2:
GTICCICC(G/A)TT(C/T)TT(G/A)CAIGG(G/A)TT(配列番号25)[EGF様反復配列の多
くの中に存在する配列であるNPCKNGGT(配列番号26)をコードしている]
増幅条件:50ngのcDNA、1μgの各プライマー、0.2mMのdNTP、1.
8UのTaq(Perkin-Elmer)、50μlの供給緩衝液中。94℃/30秒、45℃/2分
、72℃/1分の40サイクル(各サイクル2秒ずつ延長)。
【0190】
増幅された断片は約650塩基対の断片であり、これを部分的に配列決定してC-Delta-
1との相関関係を決定した。
【0191】
c)マウスの11.5日のcDNAライブラリー(Clontech)をスクリーニングした。い
くつかの陽性クローンのうち、ひとつ(pMDL2、インサートのサイズは約4kb)が完全
なタンパク質コード領域を含んでいた。このDNA配列を完全に決定した。
【0192】
図7(配列番号11)はM-Delta-1DNAを含有する単離されたクローンのヌクレオチ
ド配列を示す。
【0193】
図8(配列番号12)はM-デルタ-1の予想されるアミノ酸配列を示す。
【0194】
図9はM-デルタ-1とC-デルタ-1に対して予想されるアミノ酸配列を並べて示す。同一の
アミノ酸を四角で囲って示す。広範囲の配列相同性を示している。コンセンサス配列を下
に示す(配列番号13)。
【0195】
発現パターン:発現パターンは、C-デルタ-1に対して、特に体節形成前の中胚葉、中枢
神経系、末消神経系および腎臓で観察されたものとほぼ同じであることが確認された。
【0196】
8.ヒトデルタホモログの単離と特性決定
H-デルタ-1(HD1)と名付けたヒトデルタ-1ホモログを以下のようにして単離した。
【0197】
インサートのサイズが100〜150kbの範囲であるヒトゲノムライブラリーをマウ
スデルタ-1(M-Delta-1)遺伝子のEcoRI断片で探索した。このライブラリーから、E
coRI断片とハイブリダイズしたゲノムヒトPACクローンを単離した。次に、2つの
縮重しているオリゴヌクレオチドを用いて、ゲノムヒトPACクローンの断片をPCRに
より増幅した。縮重オリゴは次の通り。
【0198】
5′ACIATGAA(C/T)AA(C/T)CTIGCIAA(C/T)TG(配列番号27)[TMNNLANC(配列番号28
)をコード]および
3′AC(A/G)TAIACIGA(C/T)TG(A/G)TA(C/T)TTIGT(配列番号29)[TKYQSVYV(配列番号
30)をコード]または
3′GC(A/G/T)ATIAC(A/G)CA(C/T)TC(A/G)TC(C/T)TT(C/T)TC(配列番号31)[EKDECVIA
(配列番号32)をコード]。
【0199】
ニワトリとマウスのデルタ-1に対するcDNA配列に基づいて、5′オリゴと2つの異
なる3′オリゴを用いて、約354塩基対および387塩基対の断片がそれぞれ単離され
ると予想された。しかし、実際には、525塩基対と予想通りそれより30塩基対小さい
単一の単離物が2つ得られた。大きい方の単離物をジデオキシシークエンシングで配列決
定した。そのヌクレオチド配列を図10に示す(配列番号14)。また、図10には、増
幅したDNA断片の3つの異なるリーディングフレームに対して予想されたアミノ酸配列
(配列番号15、16、17)も示す。シークエンシングエラーのため、両方のプライマ
ー間で中断されてない充分な配列は同定されなかった。その結果、この得られたDNA配
列から直接アミノ酸配列を予想することはできない。しかし、図11に、ヒトデルタ-1(
上)(配列番号18)とヒヨコのデルタ-1(下)との間の肉眼で決定したアミノ酸配列相
同性を示す。シークエンシングエラーのため、この相同性は、相同領域を同定するために
3つの異なるリーディングフレームの間で切り替えることによって得た。
【0200】
完全長のH-デルタ-1(HD1)遺伝子を単離する目的で、大きい方の単離物(配列番号
14)をプローブとして用い、ヒト胎児脳プラスミドライブラリー(Clontech)をスクリー
ニングした。この結果4つの陽性プラークが得られた。これらの陽性プラークのうちの2
つ(HD13およびHD124)が再度のスクリーニングで生き残り、これをサザンブロ
ットで大きいヒトゲノム断片と陽性反応させた。これらの陽性クローンをEcoRIで消
化し、断片をBluescriptKS-ベクターに連結することによってサブクローン化した。T3
およびT7プライマーを用いたジデオキシシークエンスによりインサートのヌクレオチド
配列を得た。結果は、HD124がその両端でニワトリのデルタ-1と相同であるが、HD
13の一端は相同性を示さないことを示していた。一連の酵素による制限消化は2つのク
ローンで非常に類似したパターンを示した。これらはいずれもインサートが約2kbであ
ったがHD13の3′端が異なっていた。
【0201】
HD13とHD124をBstXI、XbaI、HindIIIおよびXhoIで消化
し、制限断片をBluescriptKS-に挿入した後上記のようにして配列決定して内部配列を得
た。得られた配列は3′にHD1の約2000塩基をもっており、3′非コード領域に伸
びていた。HD13はHD124の中に含まれているが、HD13の5′端に付加された
配列はクローニングの際に人工的に入ったものであろう。
【0202】
こうして得られた配列はHD1の5′端を含有していなかったので、T細胞ライブラリ
ーおよび他の胎児脳ライブラリー(Lambda-Zap,Stratagene)でのハイブリダイゼーション
にHD124をプローブとして使用した。T細胞ライブラリーのスクリーニングでは陽性
のものが得られなかった。しかし、Lambda-Zapライブラリーのスクリーニングでは2つの
陽性クローンHD113とHD118が得られた。これらのクローンを、上記のようにE
coRIを用いてBluescriptKS-ベクターに挿入した。HD13およびHD124と比較
するためにインサートを一連の制限酵素で消化し、T3およびT7プライマーを用いて5
′端と3′端を配列決定した。HD113は、配列決定したときに公知のデルタと相同性
を示さないcDNAの小片のみであることが決定された。しかし、HD118は長さが3
kbであり、HD124の全体配列とさらに5′端の追加の配列を含んでいた。重なり欠
失を用いて一組のクローンをHD118から単離し、これらのクローンを自動シークエン
サーを用いてジデオキシシークエンスにかけた。図12Aは、クローンHD118から得
たヒトDeltaの部分ヌクレオチドコンティグ配列(配列番号33)を示す。シークエンシ
ングエラーのため、ヒトDeltaの中断されてない充分なヌクレオチド配列は決定されなか
った。図12Bに、ヒトDeltaの部分ヌクレオチドコンティグ配列(配列番号33)を一
番上に、また、その下に3つの異なるリーディングフレームで予想されるアミノ酸配列を
示す。2番目はリーディングフレーム1(配列番号34)、3番目はリーディングフレー
ム2(配列番号35)、4番目はリーディングフレーム3(配列番号36)である。
【0203】
配列の相同性はマウスデルタ-1アミノ酸配列を用いて肉眼で決定した。マウスアミノ酸
配列と最も相同性の高い配列は図12Bで四角で囲って示し、ヒトデルタ-1の予想された
アミノ酸配列を示す。得られた複合のアミノ酸配列を図14に示す。(図14では、ヒト
デルタ配列の中断されてないさまざまな部分をそれぞれ配列番号39〜65として表示し
てある。)シークエンシングエラーのため最高の相同性をもつリーディングフレームは配
列全体を通して同じではなく、配列中にエラーがある位置でシフトされていることに注意
されたい。
【0204】
また、ニワトリとマウスのデルタに対して肉眼で決定した相同性は、決定されたヒトDe
ltaヌクレオチド配列から推定したアミノ酸配列が、ヒトデルタ-1のN末端の100〜1
50アミノ酸のほぼすべてを含有していることを示している。
【0205】
図13に、マウスDelta-1のヌクレオチド配列(一番上、配列番号37)、図12Aお
よび12Bに示したヒトDelta-1のコーディングヌクレオチド配列(2番目、配列番号3
3)、そしてマウスとヒトのDeltaの間のヌクレオチドコンセンサス配列(3番目、配列
番号38)を示す。
【0206】
図12Aに示したヒトデルタ5′ヌクレオチド配列を含有するプローブを用いて、cD
NAライブラリーを探索してヒトデルタ遺伝子の5′端を単離する。一次陽性クローンを
得た後二次陽性物として確認する。二次陽性物を精製しさらに増殖させる。その後DNA
を単離し配列決定のためにサブクローン化する。
【0207】
本発明の範囲は本明細書に記載した特定の態様に限定されるものではない。実際、以上
の説明と添付した図面から、本明細書に記載したものに加えて本発明のさまざまな修正が
当業者には明らかであろう。そのような修正は添付の請求の範囲内に入るものと考えられ
る。
【0208】
本明細書でさまざまな引用をしたがその開示内容は引用したことによってその全体が本
明細書に含まれているものとする。
【技術分野】
【0001】
本発明は、1995年06月28日に出願された米国仮出願(出願番号60/000,589)について優
先権を主張する。この仮出願は、参考文献としてその全体が本明細書中に取込まれるもの
である。
【0002】
1.はじめに
本出願は、脊椎動物のDelta遺伝子およびそれがコードするタンパク質産物、ならびに
その誘導体および類似体に関する。脊椎動物のデルタタンパク質、誘導体、および抗体の
産生もまた提供される。本発明はさらに、治療用組成物と検査方法および治療方法に関す
る。
【背景技術】
【0003】
2.発明の背景
ショウジョウバエ(Drosophila)における遺伝子の分析は、発生経路の複雑度を詳細に分析し、相互作用している遺伝子座を同定する上で非常に有用であった。しかしながら、遺伝的相互作用の根底にあるプロセスの精確な性質を理解するためには、問題となっている遺伝子のタンパク質産物の知識が必要である。
【0004】
脊椎動物の中枢神経系は、異なる細胞の型の完全な混合物であり、そのほぼすべてが同じ細胞源(神経板およびそれに続く神経管を形成する神経上皮)からつくられている。この細胞のシートにおいて、いくつかが神経細胞になるにもかかわらず他の細胞が非神経細胞のままであるように、神経発生を制御する機構は何なのであろう?この答えは、脊椎動物については、実質的には知られていないが、多くの細胞の相互作用と神経発生の間の細胞の予定運命の決定を制御する遺伝子が、ショウジョウバエにおいて十分に特徴付けられている(Campos-Ortega,1993,J.Neurobiol.24:1305-1327)。神経発生の解剖学的内容の全貌は、昆虫と脊椎動物とでは非常に異なるが、細胞レベルにおいては、類似のイベントが保存された分子機構を介して生じているという可能性が残されている。発生学的、遺伝的そして分子的な証拠から、ショウジョウバエにおける初期段階の外胚葉の分化は、細胞の相互作用に依存するということが示唆された(Doeand Goodman, 1985,Dev.Biol.111:206-219; Technau
Neurogenet.2:291-308;de la Concha et al.,1988,Genetics 118:499-508; Xu et al.,1990,Genes Dev.4:464-475;Artavanis-Tsakonas,1988,Trends Genet.4:95-100)。突然変異の分析より、接合的に作用する小さな群の遺伝子、いわゆる神経遺伝子座が上皮と神経経路との間の外胚葉細胞の選択に影響を与えるということが明らかになった(Poulson,1937,Proc.Natl.Acad.Sci.23:133-137;Lehmann et
Wilhelm Roux'sArch.Dev.Biol.193:283-295; Wieschaus et al.,1984,Wilhelm
Roux's Arch.Dev.Biol.193:267-282)。接合神経遺伝子座(Notch(N),Delta(D),mastermind(mam),Enhancerof Split(E(spl)),neuralized(neu),およびbigbrain(bib)のいずれかにおけるヌル突然変異は、腹部および側面の表皮の構造の損失(expense)における神経系の過栄養を生じさせる。この効果は、表皮前駆細胞を神経系に間違ったルートで送ることによるものであり、そして神経遺伝子の機能が神経の予定運命から表皮の予定運命へと神経領域の範囲内で細胞の流れを変えるために必要であることを暗示する。
【0005】
神経前駆体は連続的な波(successive waves)の神経発生の間、神経上皮からショウジョウバエの胚で生じる(Campos-Ortega &Hartenstein,1985,キイロショウジョウバエの胚の発生(Springer-Verlarg,Berlin;New York);Doe,1992,Development116:855-863)。これらの細胞の産生パターンは、前神経遺伝子および神経遺伝子の活性によって、主として決定される。前神経遺伝子は細胞のクラスターを神経の予定運命に前もって処理するが(Skeath& Carroll,1994,Faseb J.8:714-21でレビューされた)、クラスター中のあるサブセットだけが神経前駆体になる。この制限は、神経遺伝子の作用によるものであり、この神経遺伝子は側面阻害(細胞のシグナリングの阻害のある型)を仲介し、それによって神経の予定運命に拘束される細胞が、これに隣接する細胞を拘束されないままにするかまたは非神経系に入るように強制する(Artavanis-Tsakonas& Simpson,1991,Trends Genet.7:403-408; Doe & Goodman,1985,Dev.BIol.111:206-219)。側面阻害をできないようにする突然変異は、神経細胞、すなわち、「神経の」表現型の過剰産生の原因となる(Lehmannet al.,1981,Roux's Arch.Dev.Biol.190:226-229; Lehmann et al.,Roux's Arch.Dev.Biol.192:62-74)。ショウジョウバエにおいては、阻害シグナルはDelta神経遺伝子によってコードされる膜貫通タンパク質によって送達され、このタンパク質は発生期の神経細胞によって表示される(Heitzler& Simpson,1991,Cell 64:1083-1092)。隣接する細胞は、神経遺伝子Notchによってコードされる、膜貫通受容体タンパク質を発現する(Fortini& Artavanis-Tsakonas,1993,Cell 75:1245-1247)。Delta遺伝子はNotch遺伝子座と相互作用可能な遺伝的単位として同定された(Xuet al.,1990,Genes Dev.4:464-475)。
【0006】
突然変異の分析はまた、神経遺伝子の作用が多面発現性であり、唯一胚発生に限定されるものではないことを明らかにした。例えば、個眼の形成、剛毛の形成および翼の形成は細胞相互作用に依存することもまた知られているが、これらは神経突然変異によって影響される(Morgan et al.,1925,Bibliogr.Genet.2:1-226; Welshons,1956,Dros.Inf.Serv.30:157-158;Preiss et al.,1988,EMBO J.7:3917-3927; Shellenbarger and Mohler,1978,Dev.Biol.62:432-446;Technau and Campos-Ortega,1986,Wilhelm Roux's Dev.Biol.195:445-454; Tomlisonand Ready,1987,Dev.Biol.120:366-376; Cagan and Ready,1989,Genes Dev.3:1099-1112)。神経遺伝子はまた、ハエの筋肉、消化管、外分泌系および再生系の正常な発生に必要とされる(Muskavitch,1994,Dev.Biol.166:415-430)。
【0007】
ノッチおよびデルタはいずれも膜を1回だけ貫く膜貫通タンパク質であり(Wharton et al.,1985,Cell 43:567-581; Kidd and Young,1986,Mol.Cell.Biol.
Genes Dev.2:1723-1735)、その細胞外ドメインに複数のタンデムEGF様反復配列を含む(Muskavitch,1994,Dev.Biol.166:415-430)。Notch遺伝子は、大きなN-末端細胞外ドメインを有する〜300kdタンパク質(我々は「Notch」をこのタンパク質を表すために使用する)をコードする。このタンパク質は、36個の表皮増殖因子(EGF)様タンデム反復配列とそれに続く3個の別のシステインを多く含む反復配列とを含み、これをNotch/lin-12反復配列と命名した(Whartonet al.,1985,Cell 43:567-581; Kidd and Young,1986,Mol.Cell.Biol.6:3094-3108;Yochem et al.,1988,Nature 335:547-550)。分子の研究により、ノッチとデルタとは、初期発生の決定に関与する、細胞のコミュニケーション機構の生化学的に相互作用する要素を構成するという示唆が導かれた(Fehonet al.,1990,Cell 61:523-534)。相同体が線虫(Caenorhabditis elegans)で発見され、ここでもNotch関連遺伝子lin-12とDelta関連遺伝子lag-2とは側面阻害に応答可能であった(Sternberg,1993,CurrentBiol.3:763-765; Henderson et al.,1994,Development 120:2913-2924; Greenwald,1994,Curr.Opin.Genet.Dev.4:556-562)。脊椎動物においては幾つかのノッチ相同体も同定されており(Kopan& Weintraub,1993,J.Cell Biol.121:631-641; Lardelli et al.,1994,Mech.Dev.46:123-136;Lardelli & Lendahl,1993,Exp.Cell Res.204:364-372; Weinmaster et al.,1991,Development113:199-205; Weinmaster et al.,1992,Development 116:931-941; Coffman et al.,1990,Science249:1438-1441; Bierkamp & Campos-Ortega,1993,Mech.Dev.43:87-100)、それらは多くの組織と発生の多くの段階で発現される。Notch-1の欠失は、マウスでは中胚葉節の欠損と胎児の死亡を生じさせたが(Swiateket al.,1994,Genes Dev.8:707-719; Conlon et al.,Rossant,J.Development(J.Dev.)121:1533-1545)、構造的なNotch-1の突然変異活性形は、アフリカツメガエル(Xenopus)および培養哺乳類細胞では細胞の分化を阻害するらしい(Coffmanet al.,1993,Cell 73:659-671; Kopan et al.,1994,Development 120:2385-2396; Nyeet al.,1994,Development 120:2421-2430)。
【0008】
EGF様モチーフが、血液凝固カスケードに関連するタンパク質を含めて、種々のタンパク質中で発見された(Furieand Furie,1988,Cell 53:505-518)。特に、このモチーフは血液凝固第IX因子および第X因子などの細胞外タンパク質(Rees etal.,1988,EMBO J.7:2053-2061; Furie and Furie,1988,Cell 53:505-518)、他のショウジョウバエの遺伝子(Knustet al.,1987,EMBO J.761-766; Rothnberg et al.,1988,Cell 55:1047-1059)、およびトロンボモジュリン(Suzukiet al.,1987,EMBOJ.6:1891-1897)およびLDL受容体(Sudhof et al.,1985,Science 228:815-822)などの幾つかの細胞表面受容体タンパク質において見出された。タンパク結合部位を、トロンボモジュリンおよびウロキナーゼ中のEGF反復配列ドメインにマッピングした(Kurosawaet al.,1988,J.Biol.Chem.263:5993-5996; Appella et al.,1987,J.Biol.Chem.262:4437-4440)。
【0009】
上記の参考文献は、これらが本発明に対する先行技術であるということを認めるものと解釈すべきではない。
【発明の概要】
【課題を解決するための手段】
【0010】
3.発明の概要
本発明は、脊椎動物のDelta遺伝子(ニワトリとマウスのDelta遺伝子、および他の種の
関連遺伝子)のヌクレオチド配列、およびそれらがコードするタンパク質のアミノ酸配列
、並びにその誘導体(例えば、断片)と類似体に関する。前述のヌクレオチド配列にハイ
ブリダイズ可能な核酸またはこれと相補的な核酸もまた提供される。特別な実施態様にお
いては、デルタタンパク質は哺乳類のタンパク質であり、好ましくはヒトのタンパク質で
ある。
【0011】
本発明は、脊椎動物のDelta誘導体および類似体に関し、これらは機能的に活性、すな
わち、全長(野生型の)デルタタンパク質に関連する1以上の公知の機能活性を表示する
ことができる。このような機能活性には、抗原性(抗デルタ抗体に結合する(または結合
についてデルタと拮抗する)能力)、免疫原性(デルタに結合する抗体を産生する能力)
、Notchもしくは他のトポリズミック(toporythmic)タンパク質またはそれらの断片への結
合能(または結合についてデルタと拮抗する能力)(「付着性」)、デルタの受容体への
結合能(または結合についてデルタと拮抗する能力)などが含まれるが、これらに限定さ
れるものではない。ここで使用する「トポリズミックタンパク質」は、Notch遺伝子、Del
ta遺伝子、Serrate遺伝子、Enhancerof split遺伝子、およびDeltex、並びにこの相互作
用する遺伝子のセットの他のメンバーのタンパク質産物をいい、他のメンバーは、それら
の遺伝子配列が、例えば、Delta遺伝子、Serrate遺伝子もしくはNotch遺伝子にハイブリ
ダイズする能力、またはそれらとの相同性、または表現型の相互作用を表示する能力もし
くは生化学的に相互作用するそれらのタンパク質産物の能力によって同定されてもよい。
【0012】
本発明はさらに、脊椎動物のDelta遺伝子の断片(およびその誘導体ならびに類似体)
に関し、これらは、細胞内ドメイン、細胞外ドメイン、膜貫通ドメイン、DSLドメイン、D
SLドメインに対するドメインのアミノ末端を含む1以上のデルタタンパク質のドメイン、
または1以上のデルタタンパク質のEGF様(相同)反復配列、または前述のものの組合わ
せを含むが、これらに限定されるものではない。
【0013】
脊椎動物のデルタタンパク質に対する抗体、その誘導体および類似体が、さらに提供さ
れる。
【0014】
脊椎動物のデルタタンパク質、誘導体および類似体の産生方法、例えば、組換え手段に
よる産生方法もまた提供される。
【0015】
本発明はまた、治療方法、検査方法、およびデルタタンパク質と核酸をベースとする組
成物に関する。本発明は、本発明の組成物を投与することによる細胞の予定運命または分
化の障害の治療を提供する。このような治療用組成物(本明細書中では「治療剤(Therape
utics)」という)は、デルタタンパク質とその類似体および誘導体(断片を含む);それ
らに対する抗体;デルタタンパク質、類似体、もしくは誘導体をコードする核酸;並びに
デルタアンチセンス核酸を含む。好適な実施態様においては、本発明の治療剤を投与して
ガン状態を治療し、または前新生物状態または非腫瘍状態から新生物発生または腫瘍状態
への進行を妨げる。他の特別の実施態様においては、本発明の治療剤を投与して神経系の
障害を治療し、または組織の再生および回復を促進させる。
【0016】
ある実施態様においては、ノッチおよび/またはデルタの機能と拮抗するかまたは阻害
する治療剤(以下「拮抗治療剤」という)を治療効果のために投与する。別の実施態様に
おいては、ノッチおよび/またはデルタの機能を促進させる治療剤(以下「アゴニスト治
療剤」という)を治療効果のために投与する。
【0017】
細胞の予定運命の障害、特に、ノッチおよび/またはデルタタンパク質の異常型もしく
は望ましくないレベルの発現もしくは活性または局在化を含む、過剰増殖障害(例えば、
ガン)または低増殖障害は、このようなレベルの検出によって診断することができ、以下
にさらに詳細に説明する。
【0018】
好適な特徴としては、本発明の治療剤は、少なくとも、ノッチタンパク質またはその断
片への結合を仲介するデルタの断片(以下「付着性断片」という)からなるタンパク質で
ある。
3.1.定義
本明細書中で使用するように、下線をつけてあるかまたは斜体にしてある遺伝子名は遺
伝子を示し、対照的に、それがコードするタンパク質産物は遺伝子名に下線を付していな
い。例えば、「Delta」はDelta遺伝子を意味し、「Delta」はDelta遺伝子のタンパク質産
物を表す。
【図面の簡単な説明】
【0019】
【図1A1】図1A1は、ニワトリのデルタ(C-Delta-1)のDNA配列である(配列番号1)。
【図1A2】図1A2は、ニワトリのデルタ(C-Delta-1)のDNA配列である(配列番号1)。
【図1A3】図1A3は、ニワトリのデルタ(C-Delta-1)のDNA配列である(配列番号1)。
【図1B1】図1B1は、別にスプライスされたニワトリのデルタ(C-Delta-1)のDNA配列である(配列番号3)。
【図1B2】図1B2は、別にスプライスされたニワトリのデルタ(C-Delta-1)のDNA配列である(配列番号3)。
【図2】図2は、ニワトリのデルタ(C-Delta-1)の推定アミノ酸配列である(配列番号2)。
【図3A】図3Aは、X-Delta-1(アフリカツメガエルのDelta;配列番号5)およびショウジョウバエのDelta(配列番号6)とを並べて、保存ドメイン構造、EGF反復配列、DSLドメイン、および膜貫通ドメイン(TM)を示した、C-Delta-1の推定アミノ酸配列である(配列番号2)。保存されたアミノ酸配列は四角で囲み、3つともが揃っておりかつN-末端のシステイン残基が揃っていない部分に、それぞれ●をつけた。C-Delta-1の細胞内ドメインとX-Delta-1の細胞内ドメインとは互いに非常によく似ているが、ショウジョウバエのDeltaの対応する部分に有意な相同性を示さなかった。
【図3B】図3Bは、X-Delta-1(アフリカツメガエルのDelta;配列番号5)およびショウジョウバエのDelta(配列番号6)とを並べて、保存ドメイン構造、EGF反復配列、DSLドメイン、および膜貫通ドメイン(TM)を示した、C-Delta-1の推定アミノ酸配列である(配列番号2)。保存されたアミノ酸配列は四角で囲み、3つともが揃っておりかつN-末端のシステイン残基が揃っていない部分に、それぞれ●をつけた。C-Delta-1の細胞内ドメインとX-Delta-1の細胞内ドメインとは互いに非常によく似ているが、ショウジョウバエのDeltaの対応する部分に有意な相同性を示さなかった。
【図4】図4は、C-Delta-1(配列番号2)、ショウジョウバエのDelta(配列番号6)(Vassinet al.,1987,EMBO J.6:3431-3440; Kopczynski et al.,1988,Genes Dev.2:1723-1735)、ショウジョウバエのSerrate(配列番号7)(Fleminget al.,1990,Genes Dev.4:2188-2201;Thomas et al.,1991,Development 111:749-761)、C-Serrate-1(配列番号8)(Myat,Henrique,Ish-Horowiczand Lewis,準備中)、Apx-1(配列番号9)(Melloet al.,1994,Cell 77:95-106)およびLag-2(配列番号10)(Henderson et al.,1994,Development120:2913-2924; Tax et al.,1994 Nature 368:150-154)からのDSLドメインのアライメントを、保存されたシステインのスペースをあけ、ショウジョウバエと脊椎動物のノッチ様タンパク質のために仮定されたリガンドの間で保存されたアミノ酸、およびC.elegans(線虫)のリガンドでさらに保存されたこれらとを示す(四角で囲んだ)。
【図5A】C-Delta-1の発現とC-Notch-1の発現とは、第1日(El)の神経板における神経発生の開始と相関する。左側が前である。全体をマウントしたin situハイブリダイゼーションの検体を図5A−Dに示す。5Eは断面図である。図5Aは、ステージ7において、神経板の大部分と前中胚葉節の中胚葉の根底にある部分でC-Notch-1が発現された。
【図5B】図5Bは、ステージ7におけるC-Delta-1が神経板、将来の後菱脳、第一の中胚葉節のすぐ前方ですでに検出可能である(白い四角形)。この神経ドメインの後部端は、根底にある前中胚葉節の中胚葉(psm)において、非常に強く発現されたドメインの前部の縁とほぼ水平である。神経板中における初期の発現が生じるかもしれず、根底にある中胚葉中での発現は遮蔽されるかもしれない(未発表の結果)。
【図5C】図5Cは、5B中で四角で囲った場所の拡大図であり、C-Delta-1を発現している神経板中で光散乱された細胞(scattered cells)を示す。
【図5D】図5Dでは、ステージ8において、神経板中におけるC-Delta-1の発現が神経板の発生として後に伸びている。この写真において見ることができる標識された神経板細胞のドメイン(カッコをつけた部分)は、前中胚葉節の中胚葉を超えて後ろに続いている。
【図5E】図5Eは、ステージ8における胚の準矢じり形の断面図であり、神経板(組織の背層;カッコをつけた)の光散乱細胞中でC-Delta-1が発現されており、中胚葉節の中胚葉(腹層)中で幅が広くなっている。断面の平面はわずかに斜めになっており、神経板ドメインの後部が失われている(5D参照)。
【図6A】図6A〜6Cは、C-Delta-1発現細胞がBrdUを取込んでいないことを示す。612個のC-Delta-1細胞のうち、581個がBrdU-(76個の断面;6個の胚)であった。図6Aは、細胞周期中のどの時期が神経上皮の細胞の核の頂部−基底部の位置に関連するかを示す線図である。S期の核は基底にある(Fujita,1963,J.Comp.Neurol.120:37-42; Biffo et al.,1992,Histochem.Cytochem.40:535-540)。核は影をつけて示した。
【図6B】図6A〜6Cは、C-Delta-1発現細胞がBrdUを取込んでいないことを示す。図6Bは、BrdUで2時間標識したステージ9の胚の神経管の断面図であり、C-Delta-1発現細胞(青色の地色上で暗く見える)とBrdU標識核(ピンク)を示す。標識核はDNA合成が生じる基底で優性であり、基底のC-Delta-1発現細胞は未だ標識されていない。
【図6C】図6A〜6Cは、C-Delta-1発現細胞がBrdUを取込んでいないことを示す。図6Cは4時間インキュベートしたステージ9の胚の断面図である。多くの標識核がS期を脱しており、管腔の方へ移動しているが、C-Delta-1発現細胞はまだ基底にあり、BrdUで標識されていない。
【図7A】図7Aは、マウスのDelta遺伝子(M-Delta-1)のDNA配列(配列番号11)である。
【図7B】図7Bは、マウスのDelta遺伝子(M-Delta-1)のDNA配列(配列番号11)である。
【図8】図8は、マウスのDelta遺伝子(M-Delta-1)の推定アミノ酸配列(配列番号12)である。
【図9A】図9Aは、マウスのM-Delta-1(配列番号12)とニワトリのC-Delta-1(配列番号2)の推定アミノ酸配列のアライメントを示す。これらは、それらの多くのアミノ酸配列が同一であることを示す。同一のアミノ酸は四角で囲った。2つの遺伝子の間のコンセンサス配列(配列番号13)は下の方に示した。
【図9B】図9Bは、マウスのM-Delta-1(配列番号12)とニワトリのC-Delta-1(配列番号2)の推定アミノ酸配列のアライメントを示す。これらは、それらの多くのアミノ酸配列が同一であることを示す。同一のアミノ酸は四角で囲った。2つの遺伝子の間のコンセンサス配列(配列番号13)は下の方に示した。
【図10A】ヒトDelta(H-Delta-1)のPCR増幅断片のDNA配列(配列番号14)および3つの可能なオープンリーディングフレームを用いる推定アミノ酸配列、第2行目(配列番号15)、第3行目(配列番号16)、第4行目(配列番号17)。
【図10B】ヒトDelta(H-Delta-1)のPCR増幅断片のDNA配列(配列番号14)および3つの可能なオープンリーディングフレームを用いる推定アミノ酸配列、第2行目(配列番号15)、第3行目(配列番号16)、第4行目(配列番号17)。
【図11】ヒトH-Delta-1(上段)とニワトリC-Delta-1(下段)のアライメント。ヒトデルタの推定アミノ酸配列(配列番号18)を上段に示す。ヒトデルタの配列を「肉眼」で判定し、その場合、C-Delta-1との相同性を最大とすることにより適当なリーディングフレームの配列を決定した。図10に示した単一のリーディングフレームはどれも、リーディングフレームシフトを引き起こした図10のDNA配列のエラーのため、正しい配列を与えなかった。
【図12A1】図12A1はクローンHDl 18からのヒトDelta(H-Delta-1)のコンティグ(contig)DNA配列を表す。
【図12A2】図12A2はクローンHDl 18からのヒトDelta(H-Delta-1)のコンティグ(contig)DNA配列を表す。
【図12A3】図12A3はクローンHDl 18からのヒトDelta(H-Delta-1)のコンティグ(contig)DNA配列を表す。
【図12B1】図12B1は図12Aに示したヌクレオチド配列(上段、配列番号33)および3つの可能なオープンリーディングフレームを用いる推定アミノ酸配列、第2行目(配列番号34)、第3行目(配列番号35)、第4行目(配列番号36)を表す。マウスデルタ−1のアミノ酸配列と最大の相同性を有するアミノ酸配列をボックスで囲ってある。このボックスで囲ったアミノ酸配列はヒトデルタの推定アミノ酸配列である。ここで、リーディングフレームシフトは、この配列中の配列決定エラーが存在する場所を示す。図12Aに示した単一のリーディングフレームはどれも、リーディングフレームのシフトを引き起こしたDNA配列中のエラーのため、中断されないアミノ酸配列を与えなかった。Xは未決定のアミノ酸を示し、Nは未決定のヌクレオチドを示す。
【図12B2】図12B2は図12Aに示したヌクレオチド配列(上段、配列番号33)および3つの可能なオープンリーディングフレームを用いる推定アミノ酸配列、第2行目(配列番号34)、第3行目(配列番号35)、第4行目(配列番号36)を表す。マウスデルタ−1のアミノ酸配列と最大の相同性を有するアミノ酸配列をボックスで囲ってある。このボックスで囲ったアミノ酸配列はヒトデルタの推定アミノ酸配列である。ここで、リーディングフレームシフトは、この配列中の配列決定エラーが存在する場所を示す。図12Aに示した単一のリーディングフレームはどれも、リーディングフレームのシフトを引き起こしたDNA配列中のエラーのため、中断されないアミノ酸配列を与えなかった。Xは未決定のアミノ酸を示し、Nは未決定のヌクレオチドを示す。
【図12B3】図12B3は図12Aに示したヌクレオチド配列(上段、配列番号33)および3つの可能なオープンリーディングフレームを用いる推定アミノ酸配列、第2行目(配列番号34)、第3行目(配列番号35)、第4行目(配列番号36)を表す。マウスデルタ−1のアミノ酸配列と最大の相同性を有するアミノ酸配列をボックスで囲ってある。このボックスで囲ったアミノ酸配列はヒトデルタの推定アミノ酸配列である。ここで、リーディングフレームシフトは、この配列中の配列決定エラーが存在する場所を示す。図12Aに示した単一のリーディングフレームはどれも、リーディングフレームのシフトを引き起こしたDNA配列中のエラーのため、中断されないアミノ酸配列を与えなかった。Xは未決定のアミノ酸を示し、Nは未決定のヌクレオチドを示す。
【図12B4】図12B4は図12Aに示したヌクレオチド配列(上段、配列番号33)および3つの可能なオープンリーディングフレームを用いる推定アミノ酸配列、第2行目(配列番号34)、第3行目(配列番号35)、第4行目(配列番号36)を表す。マウスデルタ−1のアミノ酸配列と最大の相同性を有するアミノ酸配列をボックスで囲ってある。このボックスで囲ったアミノ酸配列はヒトデルタの推定アミノ酸配列である。ここで、リーディングフレームシフトは、この配列中の配列決定エラーが存在する場所を示す。図12Aに示した単一のリーディングフレームはどれも、リーディングフレームのシフトを引き起こしたDNA配列中のエラーのため、中断されないアミノ酸配列を与えなかった。Xは未決定のアミノ酸を示し、Nは未決定のヌクレオチドを示す。
【図12B5】図12B5は図12Aに示したヌクレオチド配列(上段、配列番号33)および3つの可能なオープンリーディングフレームを用いる推定アミノ酸配列、第2行目(配列番号34)、第3行目(配列番号35)、第4行目(配列番号36)を表す。マウスデルタ−1のアミノ酸配列と最大の相同性を有するアミノ酸配列をボックスで囲ってある。このボックスで囲ったアミノ酸配列はヒトデルタの推定アミノ酸配列である。ここで、リーディングフレームシフトは、この配列中の配列決定エラーが存在する場所を示す。図12Aに示した単一のリーディングフレームはどれも、リーディングフレームのシフトを引き起こしたDNA配列中のエラーのため、中断されないアミノ酸配列を与えなかった。Xは未決定のアミノ酸を示し、Nは未決定のヌクレオチドを示す。
【図12B6】図12B6は図12Aに示したヌクレオチド配列(上段、配列番号33)および3つの可能なオープンリーディングフレームを用いる推定アミノ酸配列、第2行目(配列番号34)、第3行目(配列番号35)、第4行目(配列番号36)を表す。マウスデルタ−1のアミノ酸配列と最大の相同性を有するアミノ酸配列をボックスで囲ってある。このボックスで囲ったアミノ酸配列はヒトデルタの推定アミノ酸配列である。ここで、リーディングフレームシフトは、この配列中の配列決定エラーが存在する場所を示す。図12Aに示した単一のリーディングフレームはどれも、リーディングフレームのシフトを引き起こしたDNA配列中のエラーのため、中断されないアミノ酸配列を与えなかった。Xは未決定のアミノ酸を示し、Nは未決定のヌクレオチドを示す。
【図13A】マウスM-Delta-1DNA配列(第1行目、配列番号37)とヒトH-Delta-1DNA配列(第2行目、配列番号33)とそれらの共通配列(第3行目、配列番号38)のアライメント。
【図13B】マウスM-Delta-1DNA配列(第1行目、配列番号37)とヒトH-Delta-1DNA配列(第2行目、配列番号33)とそれらの共通配列(第3行目、配列番号38)のアライメント。
【図13C】マウスM-Delta-1DNA配列(第1行目、配列番号37)とヒトH-Delta-1DNA配列(第2行目、配列番号33)とそれらの共通配列(第3行目、配列番号38)のアライメント。
【図13D】マウスM-Delta-1DNA配列(第1行目、配列番号37)とヒトH-Delta-1DNA配列(第2行目、配列番号33)とそれらの共通配列(第3行目、配列番号38)のアライメント。
【図13E】マウスM-Delta-1DNA配列(第1行目、配列番号37)とヒトH-Delta-1DNA配列(第2行目、配列番号33)とそれらの共通配列(第3行目、配列番号38)のアライメント。
【図13F】マウスM-Delta-1DNA配列(第1行目、配列番号37)とヒトH-Delta-1DNA配列(第2行目、配列番号33)とそれらの共通配列(第3行目、配列番号38)のアライメント。
【図13G】マウスM-Delta-1DNA配列(第1行目、配列番号37)とヒトH-Delta-1DNA配列(第2行目、配列番号33)とそれらの共通配列(第3行目、配列番号38)のアライメント。
【図14A】図12Bからのボックスで囲ったアミノ酸配列を表す、コンポサイト・ヒトデルタ(H-Delta-1)アミノ酸配列(それぞれ配列番号39〜65)を示す。「>」は配列が下の行に続くことを示す。「*」は配列の中断を示す。
【図14B】図12Bからのボックスで囲ったアミノ酸配列を表す、コンポサイト・ヒトデルタ(H-Delta-1)アミノ酸配列(それぞれ配列番号39〜65)を示す。「>」は配列が下の行に続くことを示す。「*」は配列の中断を示す。
【発明を実施するための形態】
【0020】
5.発明の詳細な説明
本発明は、脊椎動物Delta遺伝子のヌクレオチド配列およびそれらのコード化タンパク
質のアミノ酸配列に関する。本発明はさらに、脊椎動物デルタタンパク質の断片、並びに
他の誘導体および類似体に関する。このような断片または誘導体をコードする核酸も本発
明の範囲内である。本発明は多くの異なる脊椎動物種のDelta遺伝子とそれらのコード化
タンパク質を提供する。本発明のDelta遺伝子にはニワトリ、マウス、ヒトのDelta、およ
び他の脊椎動物種の関連遺伝子(相同体)が含まれる。特定の実施態様において、Delta
遺伝子およびタンパク質は脊椎動物、特に哺乳動物に由来するものである。本発明の好ま
しい実施態様では、デルタタンパク質がヒトタンパク質である。前記タンパク質および誘
導体の、例えば組換え法による、産生も提供される。
【0021】
本発明は、機能的に活性な本発明のデルタ誘導体および類似体に関し、すなわち、それ
らは全長(野生型)タンパク質と関連した既知の機能的活性を1以上示すことができるも
のである。このような機能的活性として、抗原性〔抗デルタ抗体と結合する(または該結
合に関してデルタと競合する)能力〕、免疫原性〔デルタと結合する抗体を誘導する能力
〕、ノッチ(Notch)または他のトポリズミック(toporythmic)タンパク質もしくはその断片
と結合する(または該結合に関してデルタと競合する)能力(「接着性」)、デルタの受
容体と結合する(または該結合に関してデルタと競合する)能力、細胞の運命または分化
に影響を与える能力、および治療活性が挙げられるが、これらに限らない。本明細書中で
用いる「トポリズミックタンパク質」 とは、Notch、Delta、Serrate、Enhancer of spli
t、およびDeltex、並びにこの相互作用遺伝子ファミリーの他のメンバー(例えば、Delta
、SerrateまたはNotchとハイブリダイズするその遺伝子配列の能力、またはDelta、Serr
ateまたはNotchに対するその相同性、または表現型相互作用を示すその遺伝子の能力によ
って同定しうる)のタンパク質産物を指す。
【0022】
本発明はさらに、デルタタンパク質の細胞内ドメイン、細胞外ドメイン、DSLドメイ
ン、DSLドメインのアミノ末端側の領域、膜貫通領域、膜結合領域、もしくは1以上の
EGF様(相同)反復ドメイン、またはこれらの任意の組合せを含むがこれらに限らない
、デルタタンパク質の1以上のドメインを含むデルタの断片(並びにその誘導体および類
似体)に関する。
【0023】
さらに、脊椎動物デルタ、その誘導体および類似体に対する抗体も提供される。
【0024】
以下で実証するように、デルタは細胞の運命(分化)の決定に関与する発生および他の
生理学的プロセスにおいて、特にノッチのリガンドとして、重要な役割を果たしている。
とりわけ、デルタは中枢神経系の細胞の運命を決定づける上で主要な役割を担っていると
考えられる。本発明の核酸およびアミノ酸配列並びに抗体は、ヒトと他の生物種のデルタ
mRNAおよびタンパク質の検出・定量に、その発現の研究に、デルタとその断片、他の
誘導体および類似体の産生に、分化および他の生理学的プロセスの研究・操作に用いるこ
とができる。本発明はまた、デルタタンパク質および核酸に基づく治療および診断法、並
びに医薬組成物に関する。本発明は、本発明の治療用化合物を投与することによる細胞運
命または分化の障害の治療を提供する。このような治療用化合物(本明細書中では「治療
薬」という)としては、デルタタンパク質、その類似体および誘導体(断片を含む)、そ
れらに対する抗体、デルタタンパク質、類似体または誘導体をコードする核酸、およびデ
ルタ・アンチセンス核酸が挙げられる。好ましい実施態様では、本発明の治療薬は癌状態
を治療するために、あるいは前新生物または非悪性状態から新生物または悪性状態への進
行を防ぐために投与される。他の特定の実施態様では、本発明の治療薬は神経系の障害を
治療するために、または組織の再生および修復を促進するために投与される。
【0025】
一実施態様において、ノッチおよび/またはデルタ機能と拮抗するまたはそれらを阻害
する治療薬(以後「アンタゴニスト治療薬」という)が治療効果のために投与される。他
の実施態様において、ノッチおよび/またはデルタ機能を促進する治療薬(以後「アゴニ
スト治療薬」という)が治療効果のために投与される。
【0026】
細胞運命の疾患、特にノッチおよび/またはデルタタンパク質の発現、活性または局在
化の異常なもしくは望ましくないレベルを伴う過増殖性(例えば、癌)または低増殖性疾
患は、以下で詳述するように、このようなレベルを検出することにより診断することがで
きる。
【0027】
好ましい態様において、本発明の治療薬は、ノッチタンパク質またはその断片への結合
を媒介するデルタの少なくとも断片(本明細書中では「接着性断片」という)からなるタ
ンパク質である。
【0028】
本発明は、特に、ニワトリDelta相同体のクローニング(第6節)、マウスDelta相同体
のクローニング(第7節)、およびヒトDelta相同体のクローニング(第8節)を開示す
る以下の実施例により例示される。
【0029】
制限ではなく、開示を明瞭にするために、本発明の詳細な説明を以下の分節に大別する
。
【0030】
5.1.Delta遺伝子の単離
本発明は、脊椎動物のDelta核酸のヌクレオチド配列に関する。特定の態様では、ヒト
のDelta核酸は、図10(配列番号14)もしくは図12A(配列番号33)に示すcD
NA配列、またはそのコード領域、または脊椎動物のデルタ(Delta)タンパク質をコード
する核酸(例えば、配列番号1、3、11、14または33の配列を有する)を含む。本
発明は、脊椎動物のDelta配列の少なくとも8個のヌクレオチド(すなわち、ハイブリダ
イズ可能な部分)から成る核酸を提供し、他の態様では、核酸が、Delta配列の少なくと
も25個の(連続)ヌクレオチド、50個のヌクレオチド、100個のヌクレオチド、150個のヌ
クレオチド、もしくは200個のヌクレオチド、またはDeltaの全長をコードする配列から成
る。本発明はまた、前記配列もしくはその相補体にハイブリダイズ可能な、または相補的
な核酸にも関する。特定の発明では、脊椎動物のDelta遺伝子の少なくとも10、25、50、1
00もしくは200ヌクレオチド、または全体をコードする領域に相補的な配列を含む核酸が
提供される。特定の態様では、ストリンジェンシーの低い条件下で、脊椎動物(例えば、
哺乳類)のDelta核酸(例えば、配列番号14もしくは配列番号33またはそれらの少な
くとも10、25、50、100もしくは200個のヌクレオチド部分を有する)、またはデルタ誘導
体をコードする核酸にハイブリダイズ可能な核酸が提供される。例としては、それに限定
されないが、そのようなストリンジェンシーの低い条件を使用する手法は、次の通りであ
る(ShiloおよびWeinberg,1981,Proc.Natl.Acad.Sci.USA78:6789-6792も参照)。
すなわち、DNAを含むフィルターを、40℃で6時間、35%のホルムアミド、5X SSC、50m
Mのトリス−HCl(pH7.5)、5mMのEDTA、0.1%のPVP、0.1%のフィコール、1%
のBSAおよび500μg/mlの変性サケ精子DNAを含む溶液で前処理する。ハイブリダイ
ゼーションを同じ溶液で行うが、次の変更を伴う。すなわち、0.02%のPVP、0.02%のフ
ィコール、0.2%のBSA、100μg/mlのサケ精子DNA、10%(wt/vol)の硫酸デキストラ
ンおよび5〜20x 106cpmの32P−標識プローブを使用する。フィルターを40℃で18〜20時
間、ハイブリダイゼーション混合物中でインキュベートした後、55℃で1.5時間、2X SSC
、25mMのトリス−HCl(pH7.4)、5mMのEDTAおよび0.1%のSDSを含む溶液で洗
浄する。洗浄溶液を新しい溶液と入れ換え、60℃でさらに1.5時間インキュベートする。
フィルターを吸い取って乾かし、オートラジオグラフィー用に露光する。必要であれば、
フィルターを65〜68℃で3回目の洗浄にかけ、フィルムに再露光する。使用できるストリ
ンジェンシーの低い他の条件は、当技術分野で周知である(例えば、種の交差したハイブ
リダイゼーションに対して使用されるもの)。
【0031】
別の特定の態様では、ストリンジェンシーの高い条件下で脊椎動物(例えば、哺乳類)
のDelta核酸にハイブリダイズ可能な核酸が提供される。例えば、それに限定されないが
、そのようなストリンジェンシーの高い条件を使用する手法は次の通りである。すなわち
、DNAを含むフィルターの前ハイブリダイゼーションを、65℃で8時間〜一夜、6X SSC
、50mMのトリス−HCl(pH7.5)、1mMのEDTA、0.02%のPVP、0.02%のフィコー
ル、0.02%のBSAおよび500μg/mlの変性サケ精子DNAで構成される緩衝液中で行う。
フィルターを65℃で48時間、100μg/mlの変性サケ精子DNAおよび5〜20x 106cpmの32
P−標識したプローブを含む前ハイブリダイゼーション混合物中でハイブリダイズする。
フィルターの洗浄を、37℃で1時間、2X SSC、0.01%のPVP、0.01%のフィコールおよび
0.01%のBSAを含む溶液中で行う。この後、50℃で45分間、0.1X SSCで洗浄して、オー
トラジオグラフィーにかける。使用できるストリンジェンシーの高い他の条件は、当技術
分野で周知である。
【0032】
脊椎動物のDeltaタンパク質の断片および誘導体をコードする核酸(第5.6節参照)
ならびにDeltaアンチセンス核酸(第5.11節参照)がさらに提供される。すでに明ら
かなように、本明細書で使用する「デルタタンパク質の断片または一部をコードする核酸
」は、デルタタンパク質の列挙した断片または一部のみをコードする核酸を意味するもの
とし、デルタタンパク質の連続配列としての他の隣接した部分は意味しない。
【0033】
他のトポリズミック(toporythmic)タンパク質と相同性のある領域を含む脊椎動物のDel
ta核酸の断片も提供される。他の種のデルタタンパク質のDSL領域(DrosophilaSerra
teおよびDeltaと相同性のある領域)も提供される。図3および8に示すようなDeltaとSe
rrateとの間の保存領域をコードする核酸も提供される。
【0034】
脊椎動物のDelta遺伝子のクローニングに対する特定の態様を特定の例として以下に示
すが、それに限定されるものではない。
【0035】
発現クローニング(一般に当技術分野で公知の技術)に対しては、当技術分野で公知の
方法により、発現ライブラリーを作製する。例えば、mRNA(例えば、ヒト)を単離し
、cDNAを作製して発現ベクター(例えば、バクテリオファージ誘導体)に連結し、後
にその中に導入される宿主細胞によって発現できるようにする。次いで、種々のスクリー
ニングアッセイを使用して、発現したデルタ産物を選択することができる。一つの態様で
は、選択のために抗−デルタ抗体を使用することができる。
【0036】
別の好ましい発明では、選択の前に、PCRを使用してゲノムまたはcDNAライブラ
リーの所望の配列を増幅する。公知Delta配列(好ましくは、脊椎動物の配列)を表すオ
リゴヌクレオチドプライマーがPCRのプライマーとして使用できる。好ましい発明では
、オリゴヌクレオチドプライマーが、SerrateとDeltaとの間の相同性が強いDelta保存セ
グメントの少なくとも一部を表す。合成オリゴヌクレオチドをプライマーとして使用して
、潜在的に興味の対象となる源(RNAまたはDNA)、好ましくはcDNAライブラリ
ー由来の配列をPCRにより増幅してもよい。PCRは、例えば、Perkin-Elmer Cetusサ
ーマルサイクラーおよびTaqポリメラーゼ(Gene Amp)を使用して行うことができる。増
幅されるDNAとしては、真核細胞種由来のmRNAまたはcDNAまたはゲノムDNA
が挙げられる。PCR反応で使用するために、いくつかの異なる縮重プライマーの合成を
選択することができる。また、公知のDeltaヌクレオチド配列と単離される核酸相同体と
の間のヌクレオチド配列の類似性の多少を考慮するために、PCR反応のプライミングに
使用するハイブリダイゼーション条件のストリンジェンシーを変えることもできる。種の
交差したハイブリダイゼーションの場合は、ストリンジェンシーの低い条件が好ましい。
同じ種のハイブリダイゼーションの場合は、ストリンジェンシーが中位の条件が好ましい
。Delta相同体のセグメントの増幅の成功の後、そのセグメントは、分子的にクローン化
して配列決定し、完全なcDNAまたはゲノムクローンを単離するためのプローブとして
利用できる。これにより、次いで、以下に記載するように、遺伝子の完全なヌクレオチド
配列の決定、その発現の分析、および機能的分析のためのそのタンパク質産物の産生が可
能になる。このようにして、デルタタンパク質をコードするさらに別の遺伝子が同定でき
る。そのような手法を、以下の種々の実施例の節で例として示す。
【0037】
上記方法は、Deltaのクローンを得ることができる方法の下記の一般的記載を限定する
ものではない。
【0038】
どの脊椎動物細胞も、潜在的に、Delta遺伝子の分子クローニングに対する核酸源とし
て役立つと考えられる。Deltaをコードする核酸配列は、哺乳類、ヒト、ブタ、ウシ、ネ
コ、トリ、ウマ、イヌおよびそれ以外の霊長類源から単離できる。例えば、Deltaプライ
マーとともにcDNAライブラリーを使用したPCRによって、マウス、ニワトリおよび
ヒトのDelta遺伝子の断片を増幅した。DNAは、クローン化したDNA(例えばDNA
「ライブラリー」)から当技術分野で公知の標準的手法により、化学合成により、cDN
Aクローニングにより、または所望の細胞から精製したゲノムDNAもしくはその断片の
クローニングにより、得ることができる(例えば、Sambrookら、1989,Molecular Clonin
g,A laboratoryManual,第2版、Cold Spring Harbor Laboratory Press,Cold Sprig
Harbor,NewYork; Glover,D.M.(編)、1995,DNA Cloning: A Practical Approach,
MRL Press,Itd.,Oxford,U.K.Vol.I,IIを参照)。ゲノムDNAから誘導されるクロ
ーンは、コード領域以外に、調節およびイントロンDNA領域を含み得る。cDNA由来
のクローンは、エキソン配列のみを含む。源が何であっても、遺伝子は、遺伝子の増殖に
適するベクターに入れて分子的にクローン化すべきである。
【0039】
ゲノムDNA由来の遺伝子の分子クローニングでは、DNA断片が生じ、そのいくつか
は、所望の遺伝子をコードする。DNAは、種々の制限酵素を使用して特定の部位で切断
できる。あるいは、マンガンの存在下でDNAseを使用してDNAを断片化することが
でき、または、DNAを例えば超音波処理などによって物理的に切断することができる。
次いで、直鎖状DNA断片は、それらに限定されないが、アガロースおよびポリアクリル
アミドゲル電気泳動ならびにカラムクロマトグラフィーなどの標準的技術によって、サイ
ズに従って分離することができる。
【0040】
DNA断片がいったん生じると、所望の遺伝子を含む特定のDNA断片の同定は、多く
の方法で行うことができる。例えば、(いずれかの種の)Delta遺伝子もしくはその特異
的RNAの一部またはそれらの断片、例えば細胞外ドメイン(第5.6節参照)のある量
が入手でき、精製・標識することができるならば、生じたDNA断片は、標識したプロー
ブへの核酸ハイブリダイゼーションによってスクリーニングできる(Benton,W.およびDa
vis,R.,1977,Science196:180; Grunstein,M.およびHogness,D.,1975,Proc.Natl
.Acad.Sci.U.S.A.72:3961)。プローブと実質的に相同であるDNA断片はハイブリダ
イズする。また、制限酵素による消化、および既知制限地図が利用できるならばそれに従
って予想されるものとの断片の大きさの比較によって、適切な断片を同定することもでき
る。遺伝子の性質に基づいて、さらに選択を行うことができる。あるいは、遺伝子の有無
は、その発現産物の物理的、化学的または免疫学的性質に基づくアッセイによって検出で
きる。例えば、cDNAクローン、または適切なmRNAをハイブリッド−選択するDN
Aクローンは、電気泳動の移動、等電点挙動、タンパク質分解消化地図、結合活性、in v
itro集合活性(「接着性」)または抗原性がDeltaに対して公知のものと同様または同一
であるものが選択され得る。Deltaに対する抗体が利用できるならば、デルタタンパク質
は、標識した抗体を推定上のDeltaを合成するクローンに結合させることにより、ELI
SA(固相酵素免疫検定法)型手法で同定できる。
【0041】
Delta遺伝子はまた、核酸ハイブリダイゼーションによるmRNA選択、続くinvitro
翻訳によって同定することもできる。この手法では、断片を使用して、相補的mRNAを
ハイブリダイゼーションにより単離する。そのようなDNA断片は、別の種(例えば、Dr
osophila)の利用可能な精製DeltaDNAを表す可能性がある。単離したmRNAの単離
産物のinvitro翻訳産物の免疫沈降分析または機能的分析(例えば、in vitroでの集合力
;受容体への結合;下記を参照)によりmRNA、従って、所望配列を含む相補的DNA
断片が同定される。さらに、特異的mRNAは、細胞から単離したポリソームの、デルタ
タンパク質に対して特異的な固定化抗体への吸着によって選択できる。放射線標識したDe
lta cDNAは、(吸着したポリソームから)選択したmRNAを鋳型として使用して合
成できる。次いで、放射線標識したmRNAまたはcDNAをプローブとして使用して、
他のゲノムDNA断片からDelta DNAを同定することができる。
【0042】
DeltaゲノムDNAを単離する別の方法としては、それらに限定されないが、既知配列
からの遺伝子配列自体の化学的合成またはデルタタンパク質をコードするmRNAに対す
るcDNAの作製が挙げられる。例えば、Delta遺伝子をcDNAクローニングするため
のRNAは、Deltaを発現する細胞から単離できる。他の方法も可能であり、本発明の範
囲内である。
【0043】
同定・単離した遺伝子は、次いで、適切なクローニングベクターに挿入することができ
る。当技術分野で公知の多数のベクター−宿主系が使用できる。可能なベクターとしては
、それらに限定されないが、プラスミドまたは変性ウイルスが挙げられる。しかし、ベク
ター系は、使用する宿主細胞と相溶性でなければならない。そのようなベクターとしては
、それらに限定されないが、λ誘導体などのバクテリオファージ、またはPBR322もしくは
pUCプラスミド誘導体などのプラスミドが挙げられる。クローニングベクターへの挿入は
、例えば、DNA断片を、相補的付着末端を有するクローニングベクターに連結すること
によって行うことができる。しかし、DNAの断片化に使用する相補的制限部位がクロー
ニングベクターに存在しないならば、DNA分子の両端を酵素的に改変することができる
。あるいは、所望するいずれかの部位を、ヌクレオチド配列(リンカー)をDNA末端に
連結することにより作り出すことができる。これらの連結したリンカーは、制限エンドヌ
クレアーゼ認識配列をコードする、化学的に合成した特定のオリゴヌクレオチドを含み得
る。別の方法では、切断したベクターおよびDelta遺伝子を、ホモポリマー末端付着によ
り改変することができる。組換え分子は、形質転換、トランスフェクション、感染、電気
穿孔法などによって宿主細胞に導入することができ、その結果、遺伝子配列の多くのコピ
ーが生じる。
【0044】
別の方法では、適切なクローニングベクターに挿入した後、所望の遺伝子を、「ショッ
トガン」法で同定・単離することができる。所望遺伝子の濃縮は、例えばサイズ分画によ
って、クローニングベクターへの挿入前に行うことができる。
【0045】
特定の態様では、単離したDelta遺伝子、cDNAまたは合成したDNA配列を挿入し
た組換えDNA分子で宿主細胞を形質転換することにより、多コピーの遺伝子を生じるこ
とができる。すなわち、遺伝子は、形質転換体を増殖し、その形質転換体から組換えDN
A分子を単離し、必要であれば、挿入した遺伝子を単離した組換えDNAから回収するこ
とにより、大量に得ることができる。
【0046】
本発明によって提供されるDelta配列としては、天然の脊椎動物デルタタンパク質に見
られるものと実質的に同じアミノ酸配列をコードするヌクレオチド配列および機能的に同
等のアミノ酸を有するコードされたアミノ酸配列が挙げられ、それらは全て、デルタ誘導
体に対する下記の第5.6節に記載されている。
【0047】
5.2.Delta遺伝子の発現
脊椎動物のデルタタンパク質をコードするヌクレオチド配列またはその機能的に活性な
断片もしくは他の誘導体(第5.6節参照)は、適切な発現ベクター、すなわち、挿入さ
れたタンパク質をコードする配列の転写および翻訳に必要な要素を含むベクターに挿入で
きる。また、必要な転写および翻訳シグナルは、天然のDelta遺伝子および/またはその
フランキング領域によって供給され得る。種々の宿主−ベクター系を使用して、タンパク
質をコードする配列を発現することができる。これらには、それらに限定されないが、ウ
イルス(例えば、ワクシニアウイルス、アデノウイルスなど)感染した哺乳類細胞系;ウ
イルス(例えば、バキュロウイルス)感染した昆虫細胞系;酵母ベクターを含む酵母また
はバクテリオファージ、DNA、プラスミドDNAもしくはコスミドDNAで形質転換し
た細菌などの微生物が含まれる。ベクターの発現要素は、その強さおよび特異性が変わる
。使用する宿主−ベクター系によって、多数の適する転写・翻訳要素のいずれか一つを使
用することができる。特定の態様では、Delta遺伝子の付着部分を発現させる。他の特定
の態様では、ヒトDelta遺伝子、またはヒトデルタの機能的に活性な部分をコードする配
列を発現させる。さらに別の態様では、細胞外ドメインを含むデルタの断片、または他の
誘導体、またはデルタの類似体を発現させる。
【0048】
DNA断片のベクターへの挿入に対して先に記載した方法のいずれかを使用して、適切
な転写/翻訳制御シグナルおよびタンパク質をコードする配列から成るキメラ遺伝子を含
む発現ベクターを構築することができる。これらの方法としては、in vitro組換えDNA
および合成法ならびにin vivo組換え法(遺伝子組換え)が挙げられる。デルタタンパク
質またはペプチド断片をコードする核酸配列の発現は、デルタタンパク質またはペプチド
が組換えDNA分子で形質転換された宿主で発現されるように、第二の核酸配列によって
調節することができる。例えば、デルタタンパク質の発現は、当技術分野で公知のプロモ
ーター/エンハンサー要素によって制御できる。Delta遺伝子発現の制御に使用できるプ
ロモーターとしては、それらに限定されないが、SV40初期プロモーター領域(Bernoist
およびChambon,1981,Nature290:304-310)、ラウス肉腫ウイルスの3’の長い末端反復
配列に含まれるプロモーター(Yamamotoら、1980,Cell 22:787-797)、ヘルペスチミジン
キナーゼプロモーター(Wagnerら、1981,proc.Natl.Acad.Sci.U.S.A.78:1441-1445)
、メタロチオネイン遺伝子の調節配列(Brinster ら、1982,Nature 296:39-42);β−ラク
タマーゼプロモーター(Villa-Kamaroffら、1978,Proc.Natl.Acad.Sci.U.S.A.75:37
27-3731)またはtacプロモーター(DeBoerら、1983,Proc.Natl.Acad.Sci.U.S.A.80:2
1-25)などの原核生物発現ベクター;Scientific American,1980 242:74-94の「組換え細
菌由来の有用なタンパク質」も参照; ノパリンシンテターゼプロモーター領域を含む植物
発現ベクター(Herrera-Estrellaら、Nature303:209-213)またはカリフラワーモザイクウ
イルス35SRNAプロモーター(Gardnerら、1981,Nucl.AcidsRes.9:2871)、および光
合成酵素リブロースビスリン酸カルボキシラーゼのプロモーター(Herrera-Estrellaら、1
984,Nature310:115-120);酵素または他の真菌類由来のプロモーター要素、例えばGal4
プロモーター、ADC(アルコールデヒドロゲナーゼ)プロモーター、PGK(ホスホグ
リセロールキナーゼ)プロモーター、アルカリホスファターゼプロモーター、および組織
特異性を示し、トランスジェニック動物に利用されている下記の動物転写制御領域など;
膵小胞細胞において活性なエラスターゼI遺伝子制御領域(Swiftら、1984,Cell 38:639-
646; Ornitzら、1986,ColdSpring Harbor Symp.Quant.Biol.50:399-409; MacDonald
,1987,Hepatology7:425-515); 膵臓β細胞において活性なインスリン遺伝子制御領域(
Hanahan,1985,Nature315:115-122)、リンパ系細胞において活性な免疫グロブリン遺伝
子制御領域(Grosschedlら、1984,Cell38:647-658; Adamesら、1985,Nature 318:533-5
38; Alexanderら、1987,Mol.Cell.Biol.7:1436-1444)、精巣、乳房、リンパ系および
マスト細胞において活性なマウス乳癌ウイルス制御領域(Lederら、1986,Cell 45:485-49
5)、肝臓において活性なアルブミン遺伝子制御領域(Pinkertら、1987,Genesand Devel
.1:268-276)、肝臓において活性なα−フェトプロテイン遺伝子制御領域(Krumlaufら、1
985,Mol.Cell.Biol.5:1639-1648;Hammerら、1987,Science 235:53-58)、肝臓にお
いて活性なα1−抗トリプシン遺伝子制御領域(Kelsey ら、1987,Genes and Devel.1:1
61-171)、骨髄性細胞において活性なβ−グロビン遺伝子制御領域(Mogramら、1985,Natu
re 315:338-340;Kolliasら、1986,Cell 46:89-94); 脳のオリゴデンドロサイト細胞に
おいて活性なミエリン塩基性タンパク質遺伝子制御領域(Readheadら、1987,Cell 48:703
-712); 骨格筋において活性なミオシン軽鎖−2遺伝子制御領域(Sani,1985,Nature314
:283-286)、および視床下部において活性な性腺刺激ホルモン放出ホルモン遺伝子制御領
域(Masonら、1986,Science234:1372-1378)が挙げられる。
【0049】
Delta遺伝子挿入物を含む発現ベクターは、一般的な3つの方法によって同定できる。
すなわち、(a)核酸ハイブリダイゼーション、(b)「マーカー」遺伝子機能の有無、
および(c)挿入配列の発現である。最初の方法では、発現ベクターに挿入した外来遺伝
子の存在を、挿入したトポリズミック遺伝子に相同な配列を含むプローブを使用した核酸
ハイブリダイゼーションにより検出することができる。第二の方法では、組換えベクター
/宿主系を、外来遺伝子のベクターへの挿入によって引き起こされるある種の「マーカー
」遺伝子機能(例えば、チミジンキナーゼ活性、抗生物質に対する耐性、形質転換表現型
、バキュロウイルスにおける閉鎖体の形成など)の有無に基づいて同定し、選択すること
ができる。例えば、Delta遺伝子をベクターのマーカー遺伝子配列内に挿入する場合、Del
ta挿入物を含む組換え体は、マーカー遺伝子機能の不在によって同定できる。第三の方法
では、組換え発現ベクターを、組換え体によって発現される外来遺伝子産物をアッセイす
ることにより同定することができる。そのようなアッセイは、例えば、Delta遺伝子産物i
n vitroアッセイ系の物理的または機能的特性、例えば、ノッチとの集合(結合)、受容
体への結合、抗体との結合に基づくことができる。
【0050】
特定の組換えDNA分子がいったん同定され、単離されると、当技術分野で公知のいく
つかの方法を使用してそれを増殖することができる。適切な宿主系および増殖条件がいっ
たん確立されると、組換え発現ベクターをたくさん増殖し、作製することができる。先に
説明したように、使用できる発現ベクターとしては、それらに限定されないが、次のベク
ターまたはそれらの誘導体が挙げられる。すなわち、ほんの少し挙げると、ワクシニアウ
イルスまたはアデノウイルスなどのヒトまたは動物ウイルス;バキュロウイルスなどの昆
虫ウイルス;酵母ベクター;バクテリオファージベクター(例えば、ラムダ)、ならびに
プラスミドおよびコスミドベクターである。
【0051】
さらに、宿主細胞菌株は、挿入配列の発現を調節し、または遺伝子産物を所望の特定の
様式で改変し、処理するものを選択することができる。ある種のプロモーターからの発現
は、ある種の誘発剤の存在下で高めることかできる。すなわち、遺伝子操作したデルタタ
ンパク質の発現は、制御することができる。さらに、種々の宿主細胞は、タンパク質の翻
訳および翻訳後プロセシングならびに改変(例えば、〔例えば、シグナル配列の〕グリコ
シル化、切断)に対して特徴的かつ特異的な機構を有する。適切な細胞系または宿主系は
、発現された外来タンパク質の所望の改変およびプロセシングが確実に行われるように選
択できる。例えば、細菌系での発現を使用すると、グリコシル化されていないコアタンパ
ク質産物を産生することができる。酵母での発現は、グリコシル化タンパク質を産生する
。哺乳類細胞での発現は、異質哺乳類デルタタンパク質の「天然の」グリコシル化を確実
に行うことができる。さらに、種々のベクター/宿主発現系により、タンパク質分解など
の処理反応を種々の程度に行うことができる。
【0052】
他の特定の態様では、デルタタンパク質、断片、類似体または誘導体を、融合またはキ
メラタンパク質産物((種々のタンパク質の)異質タンパク質配列にペプチド結合によっ
て結合したタンパク質、断片、類似体または誘導体を含む)として発現させることができ
る。そのようなキメラ産物は、所望のアミノ酸配列をコードする適切な核酸配列を、適切
なコードフレームにおいて、当技術分野で公知の方法によって互いに連結し、キメラ産物
を当技術分野で一般に公知の方法によって発現することにより作ることができる。あるい
は、そのようなキメラ産物は、タンパク質合成技術、例えばペプチド合成機を使用するこ
とにより、作ることができる。
【0053】
cDNAおよびゲノム配列は共に、クローン化し、発現させることができる。
【0054】
5.3.Delta遺伝子産物の同定および精製
特定の発明では、本発明は、脊椎動物のデルタ、好ましくはヒトのデルタおよび抗原決
定基を含む(すなわち、抗体によって認識できる)、または、そうでなければ機能的に活
性なそれらの断片および誘導体のアミノ酸配列、ならびにそれらをコードする核酸配列を
提供する。本明細書で使用する「機能的に活性な」物質とは、全長(野生型)のデルタタ
ンパク質に関連する1個以上の公知の機能活性、例えば、ノッチまたはその一部への結合
、他のデルタリガンドへの結合、抗原性(抗−Delta抗体への結合)などを示す物質を意
味する。
【0055】
特定の態様では、本発明は、少なくとも6個のアミノ酸、10個のアミノ酸、25個のアミ
ノ酸、50個のアミノ酸または少なくとも75個のアミノ酸から成るデルタタンパク質の断片
を提供する。該断片を含む分子も提供する。他の態様では、タンパク質は、脊椎動物のデ
ルタタンパク質の細胞外ドメイン、DSLドメイン、上皮増殖因子様反復(ELR)ドメ
イン、ELRのいずれかの組み合わせ、膜貫通ドメイン、もしくは細胞内(細胞質)ドメ
イン、もしくはノッチに結合する部分、またはそれらの組み合わせを含み、あるいは、本
質的にそれらから成る。デルタタンパク質の前記領域のいくつか若しくは全部が欠けた断
片、または断片を含むタンパク質も提供する。前記をコードする核酸を提供する。
【0056】
Delta遺伝子配列を発現する組換え体がいったん同定されると、遺伝子産物を分析する
ことができる。これは、産物の物理的または機能的特性に基づくアッセイ、例えば、産物
の放射能標識および続くゲル電気泳動、イムノアッセイなどによる分析により達成される
。
【0057】
デルタタンパク質がいったん同定されると、クロマトグラフィー(例えば、イオン交換
、アフィニティーおよびサイジングカラムクロマトグラフィー)、遠心分離、示差溶解度
などの標準的方法、またはタンパク質を精製するための他の標準的方法により、単離・精
製することができる。機能的特性は、適切なアッセイを使用して評価できる(第5.7節
参照)。
【0058】
あるいは、組換え体によって産生されるデルタタンパク質がいったん同定されると、そ
のタンパク質のアミノ酸配列は、組換え体に含まれるキメラ遺伝子のヌクレオチド配列か
ら推定できる。その結果、タンパク質は、当技術分野で公知の標準的化学方法によって合
成できる(例えば、Hunkapiller,M.ら、1984,Nature310:105-111)。
【0059】
本発明の特定の態様では、該デルタタンパク質としては、組換えDNA法または化学合
成法のいずれで産生するにしても、それらに限定されないが、基本アミノ酸配列として、
実質的に図2、8、11または14(配列番号2、10、16および39〜65)に示す
アミノ酸配列の全部または一部を含むもの、ならびに断片および他の誘導体、ならびにそ
れらの類似体が挙げられる。
【0060】
5.4.Delta遺伝子およびタンパク質の構造
脊椎動物のDelta遺伝子およびタンパク質の構造は、当技術分野で公知の種々の方法に
よって分析できる。
【0061】
5.4.1.遺伝子分析
Delta遺伝子に対応するクローン化したDNAまたはcDNAは、それらに限定されな
いが、サザンハイブリダイゼーション(Southern,E.M.,1975,J.Mol.Biol.98:503-51
7)、ノーザンハイブリダイゼーション(例えば、Freemanら、1983,Proc.Natl.Acad.
Sci.U.S.A.80:4094-4098を参照)、制限エンドヌクレアーゼマッピング(Maniatis,T.
,1982,MolecularCloning,A Laboratory,Cold Spring Harbor,New York)およびDN
A配列決定分析などの方法によって分析できる。ポリメラーゼ連鎖反応(PCR;米国特
許No.4,683,202、4,683,195および4,889,818;Gyllensteinら、1988,Proc.Natl.Acad
.Sci.U.S.A.85:7652-7656;Ochmanら、1988,Genetics 120:621-623; Lohら、1989,S
cience243:217-220)、およびそれに続くDelta−特異的プローブを用いたサザンハイブリ
ダイゼーションにより、種々の細胞型のDNAにおけるDelta遺伝子の検出が可能である
。PCR以外の増幅法は一般に公知であり、それらも使用できる。一つの態様では、サザ
ンハイブリダイゼーションを使用してDeltaの遺伝子結合を調べることができる。ノーザ
ンハイブリダイゼーション分析を使用すると、Delta遺伝子の発現を調べることができる
。発生または活性の種々の状態にある種々の細胞型のDelta発現を試験することができる
。そのような技法の例およびそれらの結果を、下記第6節に記載する。サザンおよびノー
ザンハイブリダイゼーションのハイブリダイゼーション条件のストリンジェンシーは、使
用する特定のDeltaプローブに対して所望の相関度を有する核酸の検出が確実に行われる
ように操作することができる。
【0062】
制限エンドヌクレアーゼマッピングを使用すると、Delta遺伝子の大体の遺伝子構造を
決定することができる。制限エンドヌクレアーゼ切断によって誘導される制限地図は、D
NA配列決定分析によって確認できる。
【0063】
DNA配列決定分析は、それらに限定されないが、マキサム・ギルバート法(1980,Met
h.Enzymol.65:499-560)、サンガーのジデオキシ法(Sanger,F.,ら、1977,Proc.Natl
.Acad.Sci.U.S.A.74:5463)、T7DNAポリメラーゼの使用(TaborおよびRichardso
n,米国特許No.4,795,699)または自動DNA配列決定装置の使用(例えば、AppliedBios
ystems,Foster City,CA)などの当技術分野で公知の任意の技術によっても行うことがで
きる。
【0064】
5.4.2.タンパク質の分析
デルタタンパク質のアミノ酸配列は、DNA配列からの推定、あるいは、例えば自動ア
ミノ酸配列決定装置を使用する、タンパク質の直接の配列決定により誘導できる。代表的
なデルタタンパク質のアミノ酸配列は、実質的に図2に示し、下記第6節で詳述する配列
を含み、代表的な成熟タンパク質は、アミノ酸番号1〜728によって示される。
【0065】
デルタタンパク質の配列は、親水性分析(Hopp,T.およびWoods,K.,1981,Proc.Nat
l.Acad.Sci.U.S.A.78:3824)によってさらに解析することができる。親水性の特徴は
、デルタタンパク質の疎水性および親水性領域ならびに該領域をコードする遺伝子配列の
対応する領域の同定に使用できる。恐らく、親水性領域が免疫原性であると思われる。
【0066】
二次構造分析(Chou,P.およびFasman,G.,1974,Biochemistry13:222)も、デルタの
特異的二次構造と考えられる領域の同定のために行うことができる。
【0067】
操作、翻訳および二次構造の推定、ならびにオープンリーディングフレームの推定およ
びプロッティングは、当技術分野で利用できるコンピューターソフトウェアプログラムを
使用して行うこともできる。
【0068】
構造分析の他の方法も使用できる。これらには、それらに限定されないが、X−線結晶
学(Engstom,A.,1974,Biochem.Exp.Biol.11:7-13)およびコンピューターによる模型
作成(Fletterick,R.およびZoller,M.編、1986,ComputerGraphics and Molecular M
odeling,inCurrent Communications in Molecular Biology,Cold Spring Harbor Labo
ratory,ColdSpring Harbor,New York)が含まれる。
【0069】
5.5.デルタ(Delta)タンパク質およびその誘導体に対する抗体の生成
本発明に従って、脊椎動物のデルタタンパク質、その断片、その他の誘導体、またはそ
れらのアナログを免疫原として使用してそのような免疫原を認識する抗体を生成すること
ができる。そのような抗体としては、ポリクローナル、モノクローナル、キメラ、単鎖、
Fab断片およびFab発現ライブラリーが包含されるがこれらに限られるわけではない
。特定の態様においてはヒトデルタに対する抗体を産生する。別の態様においてはデルタ
の細胞外ドメインに対する抗体を産生する。また別の態様においてはデルタの細胞内ドメ
インに対する抗体を産生する。
【0070】
デルタタンパク質または誘導体またはアナログに対するポリクローナル抗体を産生する
には業界で公知の各種手法を用いることができる。特定の態様においては、図1a、1b
、7または11に示す配列またはそのサブ配列によってコードされているデルタタンパク
質のエピトープに対するウサギのポリクローナル抗体を得ることができる。抗体の産生に
は、天然のデルタタンパク質、または合成物、またはそれらの誘導体(たとえば断片)を
注射することによって各種宿主動物、たとえばウサギ、マウス、ラットなど(これらに限
られるわけではない)を免疫することができる。免疫応答を高めるために、宿主の種に応
じて各種のアジュバントを用いることができる。たとえば、フロイント(完全および不完
全)、鉱物質ゲルたとえば水酸化アルミニウム、表面活性物質たとえばリゾレシチン、プ
ルロニックポリオール、ポリアニオン、ペプチド、油乳濁液、アオガイヘモシアニン、ジ
ニトロフェノール、ならびに、効力増強に有用なヒトアジュバントたとえばBCG(bacil
le Calmette-Guerin)およびコリネバクテリウム・パルブム(corynebacteriumparvum)が
あるがこれらに限られるわけではない。
【0071】
デルタタンパク質配列またはそのアナログに対するモノクローナル抗体を調製するには
、連続細胞系を培養して抗体分子を産生する技術が使用できる。たとえば、最初Kohlerと
Milsteinによって開発されたハイブリドーマ法(1975,Nature256:495-497)、ならびにト
リオーマ法、ヒトB細胞ハイブリドーマ法(Kozbor et al.,1983,Immunology Today 4:7
2)、およびヒトモノクローナル抗体を産生するEBV‐ハイブリドーマ法(Coleet al.,
1985,inMonoclonal Antibodies and Cancer Therapy,Alan R.Liss,Inc.,pp.77-96
)。本発明の別の態様においては、最近の技術(PCT/US90/02545)を利用して無菌動物でモ
ノクローナル抗体を産生することができる。本発明によると、ヒトハイブリドーマ(Cote
et al.,1983,Proc.Natl.Acad.Sci.U.S.A.80:2026-2030)を使用することによって
、またはヒトB細胞をin vitroでEBVウイルスにより形質転換することによって(Cole
et al.,1985,Monoclonal Antibodies and Cancer Therapy, Alan R.Liss,pp.77-96)
、ヒト抗体を使用してもよいし、また得ることができる。実際、本発明に従って、適当な
生物活性をもつヒト抗体分子由来の遺伝子と共にデルタに特異的なマウス抗体分子由来の
遺伝子をスプライスすることによって「キメラ抗体」を産生するように開発された技術(M
orrison et al.,1984,Proc.Natl.Acad.Sci.U.S.A.81:6851-6855;Neuberger et a
l.,1984,Nature312:604-608; Takeda et al.,1985,Nature 314:452-454)を使用する
ことができる。このような抗体は本発明の範囲内に入る。
【0072】
本発明によると、単鎖抗体の産生について記載された技術(米国特許第4,946,778号)
を、デルタ特異的単鎖抗体の産生に適合させることができる。本発明の別の態様では、F
ab発現ライブラリーの構築に関して記載された技術(Huse et al.,1989,Science 246:
1275-1281)を利用して、デルタタンパク質、誘導体またはアナログに対する所望の特異性
をもつモノクローナルFab断片の迅速で容易な同定が可能になる。
【0073】
分子のイディオタイプを含有する抗体断片は公知の技術によって生成することができる
。そのような断片としては、たとえば、抗体分子のペプシン消化によって生成させること
ができるF(ab′)2断片、このF(ab′)2断片のジスルフィドブリッジを還元す
ることによって生成させることができるFab′断片、およびパパインと還元剤によって
抗体分子を処理することによって生成させることができるFab断片があるがこれらに限
られるわけではない。
【0074】
抗体の産生において、所望の抗体のスクリーニングは業界で公知の技術、たとえばEL
ISA(固相酵素免疫検定法)によって達成することができる。たとえば、脊椎動物のデ
ルタタンパク質の特異的ドメインを認識する抗体を選択するためには、そのようなドメイ
ンを含有するデルタ断片に結合する生成物に対して生成したハイブリドーマを検定すれば
よい。ヒトデルタに対して免疫特異的な抗体を選択するには、ヒトデルタに対する陽性の
結合およびキイロショウジョウバエデルタに対する結合欠如を基にして選択することがで
きる。
【0075】
以上の抗体は、本発明のタンパク質配列の位置決定と活性に関して業界で公知の方法に
、たとえば、これらのタンパク質のイメージング、適当な生理学的サンプル中でのそのレ
ベルの測定、診断法、などに使用することができる(たとえば、後記5.7節参照)。
【0076】
デルタタンパク質の或るドメインに対して特異的な抗体も提供される。特定の態様では
デルタのノッチ結合性断片に結合する抗体が提供される。
【0077】
本発明の別の態様では(後記参照)、抗‐デルタ抗体およびその結合性ドメインを含有
する断片が治療薬である。
【0078】
5.6.デルタタンパク質、誘導体およびアナログ
本発明はさらに、脊椎動物(たとえば哺乳動物)のデルタタンパク質、ならびに脊椎動
物のデルタタンパク質の誘導体(たとえば断片が包含されるがこれに限定はされない)お
よびアナログに係る。デルタタンパク質誘導体およびタンパク質アナログをコードする核
酸も提供される。ひとつの態様では、デルタタンパク質は上記5.1節に記載したデルタ
核酸によってコードされている。特定の態様では、タンパク質、誘導体またはアナログは
マウス、ニワトリ、ラット、ブタ、ウシ、イヌ、サルまたはヒトのデルタタンパク質のも
のである。特定の態様では脊椎動物の完全長成熟デルタタンパク質が提供される。ひとつ
の態様では、シグナル配列(アミノ末端の最初の約17個のアミノ酸)のみを欠く脊椎動
物のデルタタンパク質が提供される。
【0079】
Deltaに関連する誘導体とアナログの産生と用途は本発明の範囲内である。特定の態様
の場合、誘導体またはアナログは機能的に活性である。すなわち、野生型の完全長デルタ
タンパク質に関連する機能活性を1つ以上示すことができる。一例を挙げると、所望の免
疫原性または抗原性を有するそのような誘導体やアナログは、たとえば免疫検定、免疫、
デルタ活性の抑制、などに使用することができる。所望のデルタ特性、たとえばノッチそ
の他のトポリズミックタンパク質との結合、細胞表面受容体との結合を保持または逆に抑
制するそのような分子はそれぞれそのような特性およびその相関物のインデューサーまた
はインヒビターとして使用することができる。ひとつの特定の態様は、抗‐デルタ抗体と
結合することができるがノッチタンパク質またはその他のトポリズミックタンパク質とは
結合することができないデルタ断片に関する。デルタの誘導体またはアナログの所望の活
性は、限定されることはないが5.7節に記載する検定を始めとする業界で公知の手法に
よって試験することができる。
【0080】
特に、デルタ誘導体は、機能的に等価な分子を提供する置換、付加または欠失によりDe
lta配列を変更することによって作成することができる。ヌクレオチドコード配列の縮重
のため、Delta遺伝子と実質的に同じアミノ酸配列をコードする他のDNA配列を本発明
の実施の際に使用してもよい。これらの中には、配列内のアミノ酸残基と機能的に等価な
アミノ酸残基をコードする異なるコドンの置換によって変更されてサイレント変異を生じ
ているDelta遺伝子の全部または一部からなるヌクレオチド配列が包含されるがこれらに
限定されることはない。同様に、本発明のデルタ誘導体には、一次アミノ酸配列として、
配列内の残基が機能的に等価なアミノ酸残基で置換されてサイレント変異を生じている変
更配列を含むデルタタンパク質のアミノ酸配列の全部または一部を含有するものが包含さ
れるがこれらに限られることはない。たとえば、配列内のアミノ酸残基の1つ以上を、機
能的に等価な作用をする類似の極性をもった別のアミノ酸で置換してサイレント変異を生
じさせることができる。配列内のアミノ酸の置換はそのアミノ酸が属するクラスの他の一
員の中から選択すればよい。たとえば、非極性(疎水性)アミノ酸にはアラニン、ロイシ
ン、イソロイシン、バリン、プロリン、フェニルアラニン、トリプトファンおよびメチオ
ニンが含まれる。極性の中性アミノ酸にはグリシン、セリン、スレオニン、システイン、
チロシン、アスパラギンおよびグルタミンが含まれる。正に帯電した(塩基性)アミノ酸
にはアルギニン、リシンおよびヒスチジンが含まれる。負に帯電した(酸性)アミノ酸に
はアスパラギン酸とグルタミン酸が含まれる。
【0081】
本発明の特定の態様では、デルタタンパク質の少なくとも10個の(連続する)アミノ
酸で構成される脊椎動物デルタタンパク質の断片で構成されるかまたはこれを含むタンパ
ク質が提供される。他の態様では、デルタタンパク質の少なくとも20個または50個の
アミノ酸で構成される断片が提供される。特定の態様ではそのような断片が35個、10
0個または200個までのアミノ酸である。デルタの誘導体またはアナログとしては、脊
椎動物のデルタタンパク質またはその断片と実質的に相同であるペプチド(たとえば、同
じサイズのアミノ酸配列と比べてその少なくとも30%、50%、70%または90%が
同一のもの、たとえば、あるドメインを含むもの)、またはDeltaコード配列とハイブリ
ダイズすることができるコード核酸をもつペプチドが包含されるがこれらに限定されるこ
とはない。
【0082】
本発明のデルタ誘導体およびアナログは業界で公知のさまざまな方法によって生成する
ことができる。その生成に至る操作は遺伝子レベルまたはタンパク質レベルで実施するこ
とができる。たとえば、クローン化したDelta遺伝子配列は、業界で公知の多くの方法(Ma
niatis,T.,1990,MolecularCloning,A Laboratory Manual,2d ed.,Cold Spring Ha
rbor Laboratory,ColdSpring Harbor,New York)のいずれかによって修飾することがで
きる。前記配列は、適当な部位で(1種以上の)制限エンドヌクレアーゼによって開裂し
た後所望の場合にはさらに酵素修飾し、単離し、in vitroで連結することができる。デル
タの誘導体またはアナログをコードする遺伝子を生成する際には、そのようにして修飾さ
れた遺伝子がデルタと同じ翻訳リーディングフレーム内にあり、所望のデルタ活性がコー
ドされている遺伝子領域内で翻訳終結シグナルによって中断されないように注意しなけれ
ばならない。
【0083】
さらに、デルタをコードしている核酸配列をin vitroまたはin vivoで突然変異させて
、翻訳配列、開始配列および/または終結配列を創製および/または破壊したり、または
コード領域において変異を起こさせたり、および/または新たな制限エンドヌクレアーゼ
部位を形成するかもしくはすでに存在していたものを破壊したりして、さらなるin vitro
修飾を容易にすることができる。限定するわけではないがin vitro位置指定突然変異誘発
(Hutchinson,C.,etal.,1978,J.Biol.Chem 253:6551)、TAB(登録商標)リンカ
ー(Pharmacia)の利用、などを始めとして業界で公知のいかなる突然変異誘発法も用いる
ことができる。変化した配列を含有するPCRプライマーをPCRに用いて、増幅される
断片中にそのような変化を導入することができる。
【0084】
デルタ配列の操作はまたタンパク質レベルで行なうこともできる。本発明の範囲内に包
含されるものとして、翻訳中またはその後、たとえばグリコシル化、アセチル化、リン酸
化、アミド化、公知の保護基/ブロック基による誘導体化タンパク質分解酵素による開裂
、抗体分子その他の細胞性リガンドとの結合、などによって、さまざまに修飾されたデル
タタンパク質断片またはその他の誘導体もしくはアナログがある。限定するわけではない
が臭化シアン、トリプシン、キモトリプシン、パパイン、V8プロテアーゼ、NaBH4
による特異的化学開裂、アセチル化、ホルミル化、酸化、還元、ツニカマイシンの存在下
での代謝合成、などを始めとする多くの化学修飾のいずれも公知の技術によって実施する
ことができる。
【0085】
さらに、デルタのアナログおよび誘導体は化学的に合成することができる。たとえば、
所望のドメインを含む(5.6.1節参照)か、またはin vitroにおける所望の凝集活性
もしくは受容体との結合を媒介するデルタタンパク質の一部に相当するペプチドを、ペプ
チドシンセサイザーを使用して合成することができる。さらに、所望であれば、非古典的
なアミノ酸または化学的アミノ酸アナログを置換または付加としてデルタ配列中に導入す
ることができる。非古典的なアミノ酸としては、通常のアミノ酸のD‐異性体、α‐アミ
ノイソ酪酸、4‐アミノ酪酸、ヒドロキシプロリン、サルコシン、シトルリン、システイ
ン酸、t‐ブチルグリシン、t‐ブチルアラニン、フェニルグリシン、シクロヘキシルア
ラニン、β‐アラニン、設計用(designer)アミノ酸、たとえばβ‐メチルアミノ酸、Cα
‐メチルアミノ酸、およびNα‐メチルアミノ酸が包含されるがこれらに限定されるわけ
ではない。
【0086】
特定の態様の場合、デルタ誘導体は、脊椎動物のデルタタンパク質またはその断片(好
ましくは、デルタタンパク質の少なくともひとつのあるドメインもしくは一部、またはデ
ルタタンパク質の少なくとも10個のアミノ酸)が、そのアミノまたはカルボキシ末端に
おいてペプチド結合を介して別のタンパク質のアミノ酸配列と結合したキメラタンパク質
すなわち融合タンパク質である。ひとつの態様の場合、そのようなキメラタンパク質は、
そのタンパク質をコードする(別のタンパク質のコード配列と同じフレーム内で結合した
デルタコード配列からなる)核酸の組換え発現によって産生される。そのようなキメラ生
成物は、業界で公知の方法によって所望のアミノ酸配列をコードする適当な核酸配列を適
正なコードフレームで互いに連結し、業界で周知の方法によってそのキメラ生成物を発現
させることにより作成できる。あるいはまた、そのようなキメラ生成物はタンパク質合成
技術によって、たとえばペプチドシンセサイザーを用いて作成できる。特定の態様では、
異種シグナル配列と共に成熟デルタタンパク質をコードするキメラ核酸を発現させて、そ
のキメラタンパク質が細胞によって発現され成熟デルタタンパク質にプロセッシングされ
るようにする。限定するわけではないが別の一例では、Delta遺伝子と別のトポリズミッ
ク遺伝子(たとえばセレート)との両方のコード部分を含む組換え分子を本発明に従って
構築することができる。そのような組換え分子にコードされているタンパク質はセレート
とデルタの両方に関連する特性を発揮し、アゴニストとアンタゴニストを含めて新規なプ
ロフィールの生物学的活性を示すことができるであろう。またデルタとセレートの一次配
列を用いて、コンピューターシミュレーションによってその分子の三次構造を予想するこ
ともでき(Hoppand Woods,1981,Proc.Natl.Acad.Sci.U.S.A.78:3824-3828)、Delt
a/Serrateキメラ組換え遺伝子は三次構造と生物学的機能との相関を考慮に入れて設計す
ることができよう。同様に、任意の異種タンパク質コード配列と融合したDeltaの部分を
含むキメラ遺伝子を構築することができる。ひとつの特定の態様はデルタの少なくとも6
個のアミノ酸の断片を含むキメラタンパク質に関する。
【0087】
別の特定の態様の場合、デルタ誘導体は、別のトポリズミックタンパク質と相同の領域
を含む脊椎動物のデルタの断片である。本明細書で使用する場合、第一のタンパク質の領
域は、その領域内に含まれるアミノ酸の数と等しい数のアミノ酸をもつ第二のタンパク質
内の任意の配列と比較した場合、その領域のアミノ酸配列が少なくとも30%同一である
かまたは少なくとも70%が同一であるかもしくは保存性の変化を含んでいるとき、第二
のタンパク質と「相同である」と考えることとする。たとえば、そのようなデルタ断片は
、限定するわけではないがDSLドメインまたはその一部を含めて、セレートと相同な領
域を1つ以上含んでいることができる。
【0088】
誘導体およびアナログの他の特定の具体例は以下の小節および後記実施例で記載する。
【0089】
5.6.1.本タンパク質の1以上のドメインを含有するデルタ誘導体
ひとつの特定の態様において、本発明は、限定することはないがたとえば細胞外ドメイ
ン、シグナル配列、DSLドメインのアミノ末端領域、DSLドメイン、ELRドメイン
、膜貫通ドメイン、細胞内ドメイン、およびデルタタンパク質の1以上のEGF様反復ド
メイン(ELR)(たとえば、ELR1〜9)、またはこれらの任意の組合せを始めとす
るデルタタンパク質の1以上のドメインを含むかあるいはそれで構成される脊椎動物のデ
ルタ誘導体およびアナログ、特にデルタの断片およびそのような断片の誘導体に係る。ニ
ワトリおよびマウスのデルタタンパク質に関する特定の例の場合、そのようなドメインは
それぞれ後記実施例6および7ならびに図3および9で同定する。すなわち、例として、
脊椎動物デルタの細胞外ドメイン(アミノ酸約1〜545)、シグナル配列(アミノ酸約
1〜17)、DSLドメインのアミノ末端領域(アミノ酸約1〜178)、DSLドメイ
ン(アミノ酸約179〜223)、EGF1(アミノ酸約229〜260)、EGF2(
アミノ酸約261〜292)、EGF3(アミノ酸約293〜332)、EGF4(アミ
ノ酸約333〜370)、EGF5(アミノ酸約371〜409)、EGF6(アミノ酸
約410〜447)、EGF7(アミノ酸約448〜485)、EGF8(アミノ酸約4
86〜523)、膜貫通ドメインおよび細胞内(細胞質)ドメイン(アミノ酸約555〜
728)からなる分子が提供される。
【0090】
特定の態様の場合、脊椎動物のデルタの特定の断片を含む分子は、それぞれのデルタタ
ンパク質の中でキイロショウジョウバエまたはニワトリのデルタタンパク質の特定の断片
と最も相同である断片を含むものである。特定の具体例の場合そのような分子は配列番号
2または16のアミノ酸配列を含むかまたはその配列で構成される。あるいはまた、デル
タ相同体のドメインを含む断片は5.3.2節に記載したタンパク質解析法によって同定
することができる。
【0091】
5.6.2.トポリズミックタンパク質ドメインとの結合を媒介するデルタの誘導体
本発明はまた、トポリズミックタンパク質との結合を媒介する(したがって本明細書で
は「接着性」と名付ける)脊椎動物のデルタ断片およびそのような断片のアナログまたは
誘導体ならびにそれをコードする核酸配列も提供する。
【0092】
ひとつの特定の態様の場合デルタタンパク質の接着性断片はDSLドメインまたはその
一部からなる。DSLドメイン内で、ノッチとの結合を媒介するサブ断片は欠失突然変異
体を発現する構築物の解析によって同定することができる。
【0093】
トポリズミックタンパク質(好ましくはノッチ)と結合する能力は、そのようなトポリ
ズミックタンパク質を発現する細胞およびデルタまたはデルタ誘導体を発現する細胞を用
いるinvitro凝集検定によって立証することができる(5.7節参照)。すなわち、デル
タ断片がノッチタンパク質と結合する能力は、第一の細胞の表面上で発現した場合のデル
タ断片が、第二の細胞の表面上で発現したノッチタンパク質と結合する能力を検出するこ
とによって立証することができる。
【0094】
そのような検定に使用する、局所周期性タンパク質またはその接着性ドメインをコード
する核酸配列は、ヒト、ブタ、ウシ、ネコ、トリ、ウマ、イヌまたは昆虫ならびに霊長類
および公知のトポリズミック遺伝子のホモログが同定され得るその他の任意の種から単離
することができる。
【0095】
5.7.デルタタンパク質、誘導体およびアナログの検定
脊椎動物のデルタタンパク質、誘導体およびアナログの機能活性は各種方法によって検
定することができる。
【0096】
たとえば、ひとつの態様において、抗‐デルタ抗体に結合する能力またはその抗体との
結合に関して野生型デルタと競合する能力を検定する場合、業界で公知の各種免疫検定法
を使用することができる。たとえば、放射線免疫検定法、ELISA(固相酵素免疫検定
法)、「サンドイッチ」免疫検定法、イムノラジオメトリックアッセイ、ゲル核酸沈降反
応(gel diffusionprecipitin reactions)、免疫核酸検定法、(たとえば金コロイド、酵
素または放射性同位体標識を用いる)in situ免疫検定法、ウェスタンブロット、沈降反
応、凝集検定法(たとえばゲル凝集検定法、赤血球凝集検定法)、補体結合検定法、免疫
蛍光検定法、プロテインAアッセイ、および免疫電気泳動検定法、などのような技術を用
いる競合および非競合検定系があるがこれらに限定されることはない。ひとつの態様では
、一次抗体の標識を検出することによって抗体結合を検出する。別の態様では、二次抗体
または試薬と一次抗体の結合を検出することによって一次抗体を検出する。さらに別の態
様では二次抗体を標識する。免疫検定法で結合を検出するための多くの方法が業界で知ら
れており、本発明の範囲内である。
【0097】
別の態様において、トポリズミックタンパク質(たとえばノッチ)との結合を媒介する
能力を検定しようとする場合、in vitro凝集検定法を実施することができる(Fehon et al
.,1990,Cell61:523-534; Rebay et al.,1991,Cell 67:687-699)。 デルタに対する
受容体を同定する別の態様では、たとえば業界で周知の手段によって受容体の結合を検定
することができる。別の態様では、デルタの結合とデルタ受容体を発現する細胞との生理
学的相関(シグナル伝達)を検定することができる。
【0098】
別の態様では、昆虫やその他のモデル系において、遺伝子研究によって、野生型Delta
の誘導体またはアナログであるデルタ突然変異の表現型上の効果を研究することができる
。
【0099】
その他の方法も当業者には知られており、本発明の範囲内である。
【0100】
5.8.治療用途
本発明は、本発明の治療用化合物を投与することによって細胞の運命または分化の障害
を処置する方法を提供する。このような治療用化合物(本明細書中では「治療薬」とする
)としては、デルタタンパク質ならびにそのアナログおよび誘導体(たとえば断片)(た
とえば、上記したようなもの)、それに対する抗体(上記したようなもの)、デルタタン
パク質、アナログまたは誘導体(たとえば上記したようなもの)をコードする核酸、なら
びにDeltaアンチセンス核酸がある。上に述べたように、本発明のアンタゴニスト治療薬
は、デルタ機能および/または(デルタはノッチリガンドであるので)ノッチ機能と拮抗
するかまたはこれを抑制する治療薬である。そのようなアンタゴニスト治療薬は、たとえ
ばデルタと別のタンパク質(たとえばノッチタンパク質)との結合を抑制する当該薬の能
力または好ましくはinvitroでもしくは細胞培養で検定されるように公知のノッチまたは
デルタ機能を抑制する当該薬の能力に基づいた公知の簡便なin vitro検定法を用いて同定
するのが最も好ましい。もっとも、遺伝子検定法(たとえば、キイロショウジョウバエ)
の場合)も使用することができる。好ましい態様の場合、アンタゴニスト治療薬は、ノッ
チとの結合を媒介するデルタの断片のような機能的に活性な断片を含むタンパク質もしく
はその誘導体またはそれに対する抗体である。他の特定の態様では、そのようなアンタゴ
ニスト治療薬がノッチに結合するデルタの断片を含む分子を発現することができる核酸ま
たはDeltaアンチセンス核酸である(5.11節参照)。好ましくは、特定の治療薬の効
果を測定し、かつその投与が冒された組織の処置にとって必要であるかどうかを決定する
には、後述する適切なin vitroまたはin vivo 検定法を用いるべきであることに留意され
たい。その理由は、冒された組織の発生履歴はあるアンタゴニストまたはアゴニスト治療
薬が望まれるかどうかを決定し得るからである。
【0101】
また、デルタタンパク質または誘導体の投与様式、たとえば、可溶性形態で投与するの
か、または細胞内組換え発現用にそれをコードしている核酸を介して投与するのかは、そ
れがアゴニストとして作用するのかまたはアンタゴニストとして作用するのかに影響し得
る。
【0102】
本発明の別の態様においては、Delta遺伝子の一部を含有する核酸をアンタゴニスト治
療薬として用いて相同組換えによるDelta不活化を促進する(Koller and Smithies,1989
,Proc.Natl.Acad.Sci.USA86:8932-8935; Zijlstra et al.,1989,Nature 342:435
-438)。
【0103】
本発明のアゴニスト治療薬は上記したようにデルタ機能を促進する。そのようなアゴニ
スト治療薬としては、デルタとの結合を媒介するノッチの一部を含むタンパク質および誘
導体ならびにそれをコードする核酸(これはそれがコードしている産物をin vivoで発現
するように投与することができる)が包含されるがこれらに限られるわけではない。
【0104】
本発明の治療薬の詳細な説明と起源は本明細書中の5.1〜5.7節に記載した。
【0105】
所望のデルタ特性、たとえばノッチとの結合、細胞内リガンドとの結合を保持するかま
たは抑制する分子は、それぞれそのような特性およびその生理学的関連物のインデューサ
ーまたはインヒビターとして治療上使用することができる。特定の態様においては、デル
タのノッチと結合する部分の配列を含有するペプチド(たとえば、6〜50または15〜
25アミノ酸の範囲、特に約10、15、20または25のアミノ酸)を用いてノッチ機
能に拮抗させる。ひとつの特定の態様の場合、そのようなアンタゴニスト治療薬を用いて
、ノッチ発現の増大に関連するヒトまたはその他の悪性腫瘍(たとえば、頸管癌、結腸癌
、乳癌、扁平腺癌(後述))を処置または予防する。デルタの誘導体またはアナログの望
ましい活性は、限定されることはないが後述の実施例に記載する検定法を始めとする業界
で公知の手法によって試験することができる。たとえば、ノッチEGF‐反復ドメイン(
ELR)11および12に結合しDSLドメインより小さいデルタ断片を含む分子は、欠
失突然変異体を発現させその発現産物とノッチとの結合をいくつかの方法(たとえばin v
itro細胞凝集検定法、相互作用捕獲系。これらのうちのいくつかは後述の実施例で記載す
る)のいずれかで検定することによって取得し選択することができる。ひとつの特定の態
様においては、ペプチドライブラリーをスクリーニングして所望の活性をもつペプチドを
選択することができる。そのようなスクリーニングは、たとえばノッチまたはノッチのE
LR11および12反復ドメインを含有する分子との結合に関して検定することによって
実施することができる。
【0106】
他の治療薬としては脊椎動物のデルタタンパク質と結合する分子がある。したがって、
本発明はそのような分子を同定する方法も提供する。そのような分子は、複数の分子(た
とえば、ペプチドライブラリー、または組合せ化学ライブラリーのもの)を、結合を促進
する条件下でデルタタンパク質と接触させ、デルタタンパク質と結合する分子を回収する
ことからなる方法によって同定することができる。
【0107】
本発明のアゴニストおよびアンタゴニスト治療薬は細胞宿命の障害に対して治療上有用
である。アゴニスト治療薬は、(1)ノッチまたはデルタ機能の欠如またはレベル低下(
正常または望ましいレベルとの比較)が関与する疾患または障害において、たとえば、ノ
ッチまたはデルタタンパク質が欠如しているか、遺伝的に欠損しているか、生物学的に不
活性もしくは活性不全であるか、または発現不足である患者に、また(2)in vitro(ま
たはin vivo)検定(後述)によりデルタアゴニストの投与が有用であることが示された
疾患または障害において、治療的(予防を含む)に投与する。このノッチまたはデルタ機
能の欠如またはレベル低下は、たとえば、患者の組織サンプル(たとえば生検組織由来)
を得、その発現したノッチまたはデルタタンパク質のタンパク質レベル、構造および/ま
たは活性をinvitroで検定することによって容易に検出することができる。すなわち、業
界で標準的な多くの方法を使用することができる。たとえば、免疫検定法によりノッチま
たはデルタタンパク質を検出および/または可視化する(たとえば、ウェスタンブロット
、免疫沈降とそれに続くドデシル硫酸ナトリウムポリアクリルアミドゲル電気泳動、免疫
組織化学、など)か、かつ/またはハイブリダイゼーションアッセイによりノッチまたは
デルタのmRNAを検出および/または可視化することによってそれぞれノッチまたはデ
ルタの発現を検出する(たとえば、ノーザンアッセイ、ドットブロット、in situハイブ
リダイゼーション、など)ことができるが、これらに限定されることはない。
【0108】
特定のアゴニスト治療薬またはアンタゴニスト治療薬の投与が指示されるかどうかを決
定するのに使用することができるin vitro検定法としては、患者の組織サンプルを培養し
て増殖させ、治療薬に暴露するかまたは他の方法でこれを投与し、そのような治療薬の組
織サンプルに対する効果を観察するというin vitro細胞培養検定法がある。患者が悪性腫
瘍であるひとつの態様の場合、そのような悪性腫瘍からの細胞のサンプルを平板培養また
は培養増殖した後その細胞を治療薬に暴露する。(たとえば、末端分化を促進することに
よって)悪性細胞の生存または増殖を抑制する治療薬をin vivoの治療用として選択する
。そのような生存および/または増殖を評価するには業界で標準的な多くの検定法を使用
することができる。たとえば、細胞増殖は3H‐チミジンの取込みを測定することによっ
て、直接の細胞計数によって、原癌遺伝子(たとえばfos、myc)のような公知の遺伝子ま
たは細胞周期マーカーの転写活性の変化を検出することによって検定することができる。
また、細胞生存度はトリパンブルー染色によって評価することができ、分化は細胞形態の
変化に基づいて目視により評価することができる。特定の局面においては、悪性培養細胞
を(1)アゴニスト治療薬および(2)アンタゴニスト治療薬に別々に暴露する。検定結
果により、どちらの治療薬が治療効果をもっているかどうか指示することができる。
【0109】
別の態様の場合、増殖過多または増殖過少障害を有しているかまたは有すると疑われる
組織から得た患者の細胞サンプルに対して所望の効果、すなわちそれぞれ細胞増殖の抑制
または促進効果を示す治療薬を使用することが指示される。そのような増殖過多または増
殖過少障害としては後記5.8.1〜5.8.3節に記載するものがあるがこれらに限ら
れることはない。
【0110】
別の特定の態様の場合、冒された患者タイプの神経細胞からの神経再生/軸索伸長のin
vitro促進を示す治療薬を、神経損傷または神経系変性障害(5.8.2節参照)の処置
に使用することが指示される。
【0111】
さらに、本発明のアンタゴニスト治療薬はまた、ノッチまたはデルタ優性活性化表現型
(「機能の獲得」突然変異)が関与することが決定されているかまたは知られている疾患
または障害にも投与することが指示される。ノッチまたはデルタ優性陰性表現型(「機能
の損失」突然変異)が関与することが決定されているかまたは知られている疾患または障
害にアゴニスト治療薬を投与することが指示される。ノッチタンパク質の各種構造ドメイ
ンの機能は、hsp70熱ショックプロモーターならびに眼に特異的なプロモーターの下
で一連のキイロショウジョウバエNotch欠失突然変異を異所発現させることによってin vi
voで検討されている(Rebayet al.,1993,Cell 74:319-329参照)。2種類の優性表現
型が観察された。1つはNotch機能損失突然変異を示唆しており、他の1つはNotch機能獲
得突然変異を示唆している。優性「活性化」表現型はほとんどの細胞外配列を欠くタンパ
ク質の発現過多の結果であり、優性「陰性」表現型はほとんどの細胞内配列を欠くタンパ
ク質の発現過多の結果であった。これらの結果は、ノッチが受容体(この受容体の細胞外
ドメインはリガンド結合を媒介する)として機能し、細胞質ドメインによって発生シグナ
ルが伝達されることを示している。
【0112】
各種の特定具体例において、患者の障害に関与する細胞型の代表的な細胞を用いてin v
itro検定を実施して、ある治療薬がそのような細胞型に対して所望の効果を有するのか否
か決定することができる。
【0113】
別の態様においては、新生物発生前であることが疑われる患者の組織サンプルの細胞を
同様に平板培養またはin vitroで増殖させ、治療薬に暴露する。より正常な(すなわち、
新生物発生の状態、腫瘍状態、悪性状態、または形質転換表現型の典型ではない)細胞表
現型を生じる治療薬を治療用に選択する。新生物発生前状態、腫瘍状態、または形質転換
もしくは悪性表現型が存在するかどうか評価するには業界で標準的な多くの検定法を使用
することができる。たとえば、形質転換表現型に関連する特性(in vivo腫瘍形成能力と
関連した一組のinvitro特性)としては、より丸くなった細胞形態、より緩い基層付着、
接触阻害の損失、足場依存性の損失、プラスミノーゲンアクチベーターのようなプロテア
ーゼの放出、糖輸送の増大、血清要求性の低下、胎児抗原の発現、250,000ダルトンの表
面タンパク質の消失、などがある(Luria et al.,1978,General Virology,3d Ed.,Jo
hn Wiley &Sons,New York pp.436-446参照)。
【0114】
他の特定の態様においては、本発明に従って、処置すべき特定の患者に由来する細胞サ
ンプルではなく、処置または予防するのが望まれる悪性、腫瘍または腫瘍発現前の障害に
由来する細胞系またはそのような障害に関連する(1つ以上の)特性を示す細胞系、また
は効果が望まれる神経その他の細胞型に由来する細胞系を用いて上記したin vitro検定を
実施することができる。
【0115】
アンタゴニスト治療薬は、(1)(正常または望ましいレベルと比較して)増大したレ
ベルのノッチまたはデルタ機能が関与する疾患または障害において、たとえば、ノッチま
たはデルタタンパク質が発現過多であるかまたは活性過多である場合に、また(2)in v
itro(またはin vivo)検定によりデルタアンタゴニストの投与が有用であることが示さ
れた疾患または障害において、治療的(予防を含む)に投与する。このノッチまたはデル
タ機能の増大したレベルは、上記したような方法によって、タンパク質および/またはR
NAを定量することによって、容易に検出することができる。治療の有用性を決定するた
めに患者の組織サンプルの細胞または適当な細胞系もしくは細胞型を用いてするin vitro
検定は上記したように実施することができる。
【0116】
5.8.1.悪性腫瘍
上記したようにアンタゴニストまたはアゴニスト治療薬の効力を試験することができ、
またそのようにして治療の有用性が観察されたら処置することができる悪性および前新生
物状態としては、5.8.1および5.9.1節において下記するものが包含されるがこ
れらに限定されることはない。
【0117】
その細胞をinvitro(および/またはin vivo)で試験することができ、適当な検定結
果を観察したら本発明に従って処置することができる悪性腫瘍および関連する障害には、
表1に掲げるものがあるがこれらに限られることはない(そのような障害の概論としては
、Fishmanet al.,1985,Medicine,2d Ed.,J.B.Lippincott Co.,Philadelphiaを参
照されたい)。
【表1】
【0118】
特定の態様においては、頸、食道および肺の上皮組織のような上皮組織の悪性腫瘍また
は悪性増殖(dysproliferative)変化(化生および異形成など)を処置または予防する。
【0119】
結腸および頸部の悪性腫瘍ではヒトノッチの発現がこれらの悪性でない組織と比べて増
大している(1994年4月14日に公開されたPCT公開第WO 94/07474号(引用によりそ
の全体が本明細書に含まれているものとする)参照)。したがって、特定の態様において
は、ノッチ機能と拮抗するアンタゴニスト治療薬(たとえばデルタ誘導体)を有効量投与
することによって結腸または頸部の悪性腫瘍または前悪性変化を処置または予防する。結
腸癌や頸管癌でノッチの発現が増大していることは、もっとたくさんの癌性および増殖過
多状態がアップレギュレートされたノッチを示すことを示唆している。したがって、特定
の態様では、各種癌、たとえば、乳癌、扁平腺癌、セミノーム(精上皮腫)、黒色腫(メ
ラノーマ)および肺癌、ならびにそれらの前悪性変化、さらには他の増殖過多障害を、ノ
ッチ機能と拮抗するアンタゴニスト治療薬の投与によって処置または予防することができ
る。
【0120】
5.8.2.神経系障害
上記したようにアンタゴニストまたはアゴニスト治療薬の効力を試験することができ、
またそのようにして治療の有用性が観察されたら処置することができる細胞型を含む神経
系の障害としては、神経系の損傷、ならびに軸索の遮断、ニューロンの減退もしくは退化
、または脱髄に至る疾患または障害が包含されるがこれらに限定されることはない。本発
明に従って(ヒトおよびヒト以外の哺乳類の患者を含めて)患者において処置できる神経
系の損傷としては、中枢神経系(脊髄、脳を含む)または抹消神経系の以下の損傷が包含
されるがこれらに限定されることはない。
【0121】
(i)物理的損傷によって生じるかまたは外科に伴なう損傷を始めとする外傷、たとえば
、神経系の一部を切断する損傷、または圧迫損傷、
(ii)神経系の一部における酸素の欠乏によってニューロンの損傷または壊死に至る虚血
性損傷、たとえば、脳の梗塞もしくは虚血、または脊髄の梗塞もしくは虚血、
(iii)神経系に関連した悪性または神経系組織以外に由来する悪性である悪性組織によ
って神経系の一部が破壊または損傷される悪性損傷、
(iv)感染の結果、たとえば、膿瘍によって、またはヒト免疫不全ウィルス、帯状ヘルペ
スもしくは単純ヘルペスウィルスによる感染に伴なって、またはライム病、結核、梅毒に
伴なって、神経系の一部が破壊または損傷される感染性損傷、
(v)パーキンソン病、アルツハイマー症、ハンチントン舞踏病または筋萎縮性側索硬化
症に伴なう退化を始めとする(以上のものに限定されるわけではない)退化性過程の結果
として神経系の一部が破壊または損傷される退化性損傷、
(vi)ビタミンB12欠乏症、葉酸欠乏症、ヴェルニッケ病、たばこ‐アルコール性弱視
、マルキャファーバ・ビギャミ病(脳梁の一次退化)、およびアルコール性小脳変性を始
めとする(ただし、これらに限定されることはない)代謝の栄養的障害または障害によっ
て神経系の一部が破壊または損傷される栄養上の疾患または障害に伴なう損傷、
(vii)糖尿病(糖尿病性神経障害、ベル麻痺)、全身性エリテマトーデス、癌腫または
サルコイド症を始めとする(ただし限定されない)全身病に伴なう神経損傷、
(viii)アルコール、鉛、または特定の神経毒を始めとする毒性物質によって引き起こさ
れる損傷、
(ix)多発性硬化症、ヒト免疫不全ウィルス関連脊髄障害、横断性脊髄症または各種の病
因、進行性多病巣性白質エンセファロパシー、および中心性橋脱髄症を始めとする(ただ
し限定はされない)脱髄性の疾患によって神経系の一部が破壊または損傷される脱髄性損
傷。
【0122】
本発明に従って神経系の障害を処置するのに有用な治療薬はニューロンの生存または分
化を促進する生物活性を試験することによって選択できる(5.8節も参照のこと)。た
とえば、限定するわけではないが、次のような効果を有する治療薬が本発明で有用であろ
う。
【0123】
(i)培養におけるニューロンの生存時間の延長、
(ii)培養またはinvivoにおけるニューロンの発芽の増大、
(iii)培養またはinvivoにおけるニューロン関連分子、たとえば運動ニューロンに関す
るコリンアセチルトランスフェラーゼまたはアセチルコリンエステラーゼの産生の増大、
または
(iv)invivoにおけるニューロンの機能不全の症状の低減。
【0124】
このような効果は業界で公知のいかなる方法でも測定できる。好ましい非限定具体例の場
合、ニューロンの生存増大はArakawaらにより記載された方法で測定することができ(1990
,J.Neurosci.10:3507-3515)、ニューロンの発芽の増大はPestronkら(1980,Exp.Neur
ol.70:65-82)により、またはBrownら(1981,Ann.Rev.Neurosci.4:17-42)により記載
された方法で検出でき、ニューロン関連分子の産生増大は測定すべき分子に応じて生物検
定、酵素検定、抗体結合、ノーザンブロットアッセイなどによって測定でき、そして運動
ニューロンの機能不全は運動ニューロンの障害の物理的表示、たとえば弱さ、運動ニュー
ロンの伝達速度、または機能欠損を評価することによって測定することができる。
【0125】
特定の態様の場合、本発明に従って処置できる運動ニューロンの障害としては、運動ニ
ューロンおよび神経系の他の成分に影響を与え得る梗塞、感染、毒素に対する暴露、外傷
、外科的損傷、退化症または悪性のような障害ならびにニューロンに選択的に影響を与え
る筋萎縮性側索硬化症のような障害が包含されるがこれに限定されることはない。たとえ
ば、進行性脊髄性筋萎縮症、進行性球麻痺、一次側索硬化症、乳児性および若年性筋萎縮
症、小児の進行性球麻痺(Fazio-Londe症候群)、ポリオおよびポリオ後症候群、ならび
に遺伝性運動感覚性神経障害(Hereditary Motorsensory Neuropathy)(シャルコー・マリ
ー・ツース病)があるがこれらに限られることはない。
【0126】
5.8.3.組織の修復と再生
本発明の別の態様においては、本発明の治療薬を組織の再生と修復の促進に使用する。
たとえば、良性の異常増殖障害の処置が挙げられるがこれに限られることはない。特定の
態様は、肝臓の硬変(瘢痕形成が正常な肝臓再生過程を追越した状態)の処置、ケロイド
(肥大性の瘢痕)形成(瘢痕形成過程が正常な再生を妨げている皮膚の醜悪化)、乾癬(
皮膚の過剰増殖と適正な細胞宿命決定の遅延を特徴とする通常の皮膚の状態)、および禿
頭(最終的に分化した毛包(ノッチに富む組織)が適切に機能していない状態)の処置に
関する。別の態様では、本発明の治療薬を内耳の感覚上皮の退化性または外傷性の障害を
処置するのに使用する。
【0127】
5.9.予防用途
5.9.1.悪性腫瘍
本発明の治療薬を投与することにより、表1に掲げた障害を始めとする(しかし限定さ
れることはない)新生物または悪性状態への進行を予防することができる。そのような投
与は、その治療薬が上記した検定において前記のような障害の処置または予防に有用であ
ることが示された場合に指示される。このような予防用途は、新生物または癌に進行する
ことが知られているかまたは疑われる条件で、特に、過形成、化生、または最も特定的に
は異形成から成る非新生物細胞増殖が起こっている場合に指示されるものである(このよ
うな異常増殖条件の概論についてはRobbins and Angell,1976,Basic Pathology,2d Ed
.,W.B.Saunders Co.,Philadelphia,pp.68-79を参照されたい)。過形成は、構造ま
たは機能を大きく変えることなく、ある組織または器官において細胞数を増大させる調節
された細胞増殖のひとつの形態である。一例として、子宮内膜過形成は子宮内膜癌に先行
することが多い。化生(異形成)は、あるタイプの成体または充分に分化した細胞が別の
タイプの成体細胞の替わりとなる調節された細胞増殖のひとつの形態である。化生(異形
成)は上皮または結合組織細胞で起こり得る。典型的でない化生(異形成)には多少乱れ
た化生上皮が関与する。異形成はしばしば癌の前兆であり、主に上皮で見られる。異形成
は、個々の細胞一様性および細胞の構成的配向に損失がある非新生物細胞増殖の最も乱れ
た形態である。異形成細胞は異常に大きく深く染色した核をもっていることが多く、多形
性を示す。異形成は、慢性過敏状態または炎症が存在する場合に特徴的に起こり、頸管、
呼吸路、口腔および胆嚢で見られることが多い。
【0128】
あるいは、または、過形成、化生もしくは異形成として特徴付けられる異常な細胞増殖
の存在に加えて、患者から得た細胞サンプルがin vivoで示すかまたはin vitroで示す形
質転換表現型または悪性表現型の1つ異常な特徴が存在することは、本発明の治療薬の予
防的/治療的投与の望ましさを示し得る。上記したように、形質転換表現型のそのような
特徴としては、形態変化、より緩い基層付着、接触阻害の損失、足場依存性の損失、プロ
テアーゼの放出、糖輸送の増大、血清要求性の低下、胎児抗原の発現、250,000 ダルトン
の細胞表面タンパク質の消失、などがある(形質転換または悪性表現型に伴なう特性に関
しては前記文献の84〜90頁も参照されたい)。
【0129】
特定の態様において、白斑症、上皮の良性に出現する過形成もしくは異形成損傷、また
はボウエン病、insitu癌腫は予防的介入の望ましいことを示す腫瘍発現前の損傷である
。
【0130】
別の態様においては、線維嚢胞症(嚢胞性過形成、乳房異形成、特に腺疾患(良性上皮
過形成))が予防的介入の望ましいことを示す。
【0131】
他の態様においては、悪性に対する以下の素質的要因の1つ以上を示す患者を有効量の
治療薬の投与によって処置する。悪性に伴なう染色体転座(たとえば、慢性骨髄性白血病
に対するフィラデルフィア染色体、毛包性リンパ腫に対するt(14;18)、など)、
家族性ポリープ症もしくはガードナー症候群(結腸癌の可能な前兆)、良性のモノクロー
ナルγグロブリン異常(多発性骨髄腫の可能な前兆)、ならびにメンデルの(遺伝)遺伝
パターンを示す癌または前癌疾患(たとえば、結腸の家族性ポリープ症、ガードナー症候
群、遺伝性外骨症、多内分泌腺腫症、アミロイドを産生する髄様甲状腺癌および褐色細胞
腫、ポッツ‐ジェガース症候群、フォン・レックリングハウゼンの神経線維腫症、網膜芽
腫、頸動脈小体腫瘍、皮膚黒色癌、眼内黒色癌、色素性乾皮症、毛細管拡張性運動失調、
チェディアック‐東症候群、白子症、ファンコニ無形成貧血、およびブルーム症候群。Ro
bins and Angell,1976,BasicPathology,2d Ed.,W.B.Saunders Co.,Philadelphia
,pp.112-113参照)など)をもつ人との第一級の血族関係。
【0132】
別の特定の態様においては、本発明のアンタゴニスト治療薬をヒトの患者に投与して乳
癌、結腸癌または頸管癌への進行を予防する。
【0133】
5.9.2.他の障害
他の態様においては、本発明の治療薬を投与して、5.8.2節に記載した神経系の障
害または5.8.3節に記載した他の障害(たとえば、肝硬変、乾癬、ケロイド、禿頭)
を予防することができる。
【0134】
5.10.治療または予防上の有用性の立証
本発明の治療薬の所望の治療または予防活性はin vivoで試験することができる。たと
えば、そのような化合物は、ヒトに試験する前に、ラット、マウス、ニワトリ、ウシ、サ
ル、ウサギなどを始めとする(ただしこれらに限られるわけではない)適切な動物モデル
系で試験することができる。ヒトに投与する前のin vivoの試験には業界で公知のいかな
る動物モデル系も使用できる。
【0135】
5.11.Delta発現のアンチセンス調節
本発明は、デルタまたはその一部をコードする遺伝子またはcDNAに対してアンチセ
ンスである少なくとも6個のヌクレオチドの核酸の治療または予防上の用途を提供する。
本明細書中で「アンチセンス」とは、なんらかの配列相補性によってDeltaRNA(好ま
しくはmRNA)の一部とハイブリダイズすることができる核酸を指す。そのようなアン
チセンス核酸は本発明のアンタゴニスト治療薬として有用であり、前記5.8節およびそ
れに続く小節に記載したような障害の処置または予防に使用することができる。
【0136】
本発明のアンチセンス核酸は二本鎖または一本鎖のオリゴヌクレオチド、RNAもしく
はDNAまたはこれらの修飾体もしくは誘導体であることができ、これらは直接細胞に投
与することができるし、あるいは外来の導入された配列の転写によって細胞内で産生する
ことができる。
【0137】
ひとつの特定の態様の場合、本発明によって提供されるDeltaアンチセンス核酸は、そ
の細胞がDelta遺伝子またはNotch遺伝子を発現することを(invitroまたはin vivoで)
立証することができる腫瘍その他の障害の処置に使用することができる。そのような立証
はRNAまたはタンパク質の検出で行なうことができる。
【0138】
本発明はさらに、5.12節で後述するように製剤上許容可能な担体中に有効量の本発
明のDeltaアンチセンス核酸を含む薬剤組成物を提供する。また、本発明の薬剤組成物を
投与することからなる、(5.8および5.9節に記載したような)障害を処置および予
防する方法も提供される。
【0139】
別の態様において、本発明は原核生物または真核生物の細胞中におけるDelta核酸配列
の発現を抑制する方法に関しており、この方法は本発明のアンチセンスDelta核酸を含む
組成物を有効量細胞に与えることからなる。
【0140】
Deltaアンチセンス核酸とその用途は以下に詳細に記載する。
【0141】
5.11.1.Deltaアンチセンス核酸
Deltaアンチセンス核酸はヌクレオチドが少なくとも6個のものであり、オリゴヌクレ
オチド(6〜約50個の範囲のオリゴヌクレオチド)であるのが好ましい。特定の態様の
場合、オリゴヌクレオチドは少なくとも10個のヌクレオチド、少なくとも15個のヌク
レオチド、少なくとも100個のヌクレオチド、または少なくとも200個のヌクレオチ
ドである。オリゴヌクレオチドはDNAもしくはRNAまたはそれらのキメラ混合物もし
くは誘導体もしくは修飾体であり得り、一本鎖または二本鎖であり得る。このオリゴヌク
レオチドはその塩基部分、糖部分またはリン酸基骨格で修飾することができる。またこの
オリゴヌクレオチドはペプチド、または細胞膜を通過する輸送を容易にするもの(たとえ
ば、Letsingeret al.,1989,Proc.Natl.Acad.Sci.U.S.A.86:6553-6556; Lemaitre
et al.,1987,Proc.Natl.Acad.Sci.84:648-652;1988年12月15日に公開さ
れたPCT公開第88/09810号参照)、または血液脳関門(たとえば、1988年4月25日
に公開されたPCT公開第89/10134号参照)、ハイブリダイゼーションで誘発される開裂剤
(たとえば、Krolet al.,1988,BioTechniques 6:958-976参照)、またはインターカレ
ート剤(たとえば、Zon,1988,Pharm.Res.5:539-549参照)のような他の付随する基を
含んでいてもよい。
【0142】
本発明の好ましい態様では、好ましくは一本鎖DNAからなるDeltaアンチセンスオリ
ゴヌクレオチドが提供される。最も好ましい態様の場合、そのようなオリゴヌクレオチド
は、デルタ、最も好ましくはヒトデルタのSH3結合ドメインまたはノッチ結合ドメイン
をコードする配列とアンチセンスの配列からなる。このオリゴヌクレオチドはその構造上
の任意の位置において業界で一般に知られている置換基により修飾されていてもよい。
【0143】
デルタアンチセンスオリゴヌクレオチドは、5‐フルオロウラシル、5‐ブロモウラシ
ル、5‐クロロウラシル、5‐ヨードウラシル、ヒポキサンチン、キサンチン、4‐アセ
チルシトシン、5‐(カルボキシヒドロキシルメチル)ウラシル、5‐カルボキシメチル
アミノメチル‐2‐チオウリジン、5‐カルボキシメチルアミノメチルウラシル、ジヒド
ロウラシル、β‐D‐ガラクトシルキューオシン、イノシン、N6‐イソペンテニルアデ
ニン、1‐メチルグアニン、1‐メチルイノシン、2,2‐ジメチルグアニン、2‐メチ
ルアデニン、2‐メチルグアニン、3‐メチルシトシン、5‐メチルシトシン、N6‐ア
デニン、7‐メチルグアニン、5‐メチルアミノメチルウラシル、5‐メトキシアミノメ
チル‐2‐チオウラシル、β‐D‐マンノシルキューオシン、5′‐メトキシカルボキシ
メチルウラシル、5‐メトキシウラシル、2‐メチルチオ‐N6‐イソペンテニルアデニ
ン、ウラシル‐5‐オキシ酢酸(v)、ワイブトキソシン(wybutoxosine)、シュードウラ
シル、キューオシン、2‐チオシトシン、5‐メチル‐2‐チオウラシル、2‐チオウラ
シル、4‐チオウラシル、5‐メチルウラシル、ウラシル‐5‐オキシ酢酸メチルエステ
ル、ウラシル‐5‐オキシ酢酸(v)、5‐メチル‐2‐チオウラシル、3‐(3‐アミ
ノ‐3‐N‐2‐カルボキシプロピル)ウラシル、(acp3)w、および2,6‐ジア
ミノプリンを包含する(しかし限定されることはない)群の中から選択される少なくとも
1つの修飾された塩基部分を含んでいてもよい。
【0144】
別の態様の場合、オリゴヌクレオチドは、アラビノース、2‐フルオロアラビノース、
キシルロースおよびヘキソースを包含する(しかしこれらに限定されない)群の中から選
択される少なくとも1つの修飾された糖部分を含んでいる。
【0145】
さらに別の態様の場合、オリゴヌクレオチドは、ホスホロチオエート、ホスホロジチオ
エート、ホスホルアミドチオエート、ホスホルアミデート、ホスホルジアミデート、メチ
ルホスホネート、アルキルホスホトリエステルおよびホルムアセタールまたはこれらのア
ナログから成る群の中から選択される少なくとも1つの修飾されたリン酸基骨格を含んで
いる。
【0146】
さらに別の態様ではオリゴヌクレオチドがα‐アノマーオリゴヌクレオチドである。α
‐アノマーオリゴヌクレオチドは相補的RNAと共に特定の二本鎖ハイブリッドを形成す
る。この場合、通常のβ‐単位とは対照的に、ストランドは互いに平行になっている(Gau
tier et al.,1987,Nucl.AcidsRes.15:6625-6641)。
【0147】
オリゴヌクレオチドは別の分子、たとえばペプチド、ハイブリダイゼーションで誘発さ
れる架橋剤、輸送剤、ハイブリダイゼーションで誘発される開裂剤などと結合していても
よい。
【0148】
本発明のオリゴヌクレオチドは業界で公知の標準的な方法によって、たとえば自動DN
Aシンセサイザー(たとえば、Biosearch,Applied Biosystemsなどから市販されている
ものなど)を使用することによって合成できる。例として、ホスホロチオエートオリゴヌ
クレオチドはSteinらの方法(1988,Nucl.AcidsRes.16:3209)によって合成でき、メチ
ルホスホネートオリゴヌクレオチドは調節された多孔ガラスポリマー支持体(Sarin et al
.,1988,Proc.Natl.Acad.Sci.U.S.A.85:7448-7451)等を使用することによって調製
することができる。
【0149】
特定の態様の場合、Deltaアンチセンスオリゴヌクレオチドは触媒RNAすなわちリボ
ザイムを含んでいる(たとえば、1990年10月4日に公開されたPCT公開第90/11364
号、Sarveret al.,1990,Science 247:1222-1225参照)。別の態様ではオリゴヌクレオ
チドが2′‐O‐メチルリボヌクレオチド(Inoue et al.,1987,Nucl.Acids Res.15:6
131-6148)またはキメラRNA−DNAアナログ(Inoueet al.,1987,FEBS Lett.215:3
27-330)である。
【0150】
他の態様の場合、本発明のDeltaアンチセンス核酸は外来配列からの転写によって細胞
内で産生される。たとえば、あるベクターをin vivoで導入して、そのベクターを細胞に
取込ませ、その細胞の中でそのベクターまたはその一部が転写されて本発明のアンチセン
ス核酸(RNA)を産生するようにすることができる。そのようなベクターはDeltaアン
チセンス核酸をコードする配列を含有しているであろう。そのようなベクターは、転写さ
れて所望のアンチセンスRNAを産生することができる限りにおいて、エピソームのまま
でいることもできるし、あるいは染色体に組込まれるようになってもよい。そのようなベ
クターは業界で標準的な組換えDNA技術によって構築することができる。ベクターは哺
乳動物細胞で複製および発現に使われているプラスミド、ウィルスその他業界で公知のも
のでよい。デルタアンチセンスRNAをコードしている配列の発現は哺乳動物、好ましく
はヒトの細胞で作用することが業界で知られている任意のプロモーターによって行なうこ
とができる。そのようなプロモーターは誘導的または構成的のいずれであることもできる
。そのようなプロモーターとしては、SV40の初期プロモーター領域(Bernoist and Ch
ambon,1981,Nature290:304-310)、ラウス肉腫ウィルスの3′の長い末端反復配列に含
まれているプロモーター(Yamamotoet al.,1980,Cell 22:787-797)、ヘルペスチミジン
キナーゼプロモーター(Wagner et al.,1981,Proc.Natl.Acad.Sci.U.S.A.78:1441-
1445)、メタロチオネイン遺伝子の調節配列(Brinsteret al.,1982,Nature 296:39-42)
、などがあるがこれらに限定されることはない。
【0151】
本発明のアンチセンス核酸はDelta遺伝子、好ましくはヒトDelta遺伝子のRNA転写物
の少なくとも一部に相補的な配列を含んでいる。しかし、好ましいとはいっても完全な相
補性は必要とされない。本明細書で「RNAの少なくとも一部に相補的な」配列とは、そ
のRNAとハイブリダイズして安定な二重鎖(duplex)を形成することができるほどに充分
な相補性をもっている配列を意味している。二本鎖のデルタアンチセンス核酸の場合には
その二重鎖DNAの単一のストランドをそのように試験してもよいし、あるいは三重鎖(t
riplex)の形成を検定してもよい。ハイブリダイズする能力は相補性の程度とアンチセン
ス核酸の長さの双方に依存する。一般に、ハイブリダイズする核酸が長くなればなるほど
、それが含有し得るDeltaRNAとの塩基のミスマッチが多くなるがそれでも安定な二重
鎖(または場合によって三重鎖)を形成する可能性が高くなる。当業者は、標準的な手法
を使用して、ハイブリダイズした複合体の融点を決定するミスマッチの許容できる程度を
確認することができる。
【0152】
5.11.2.Deltaアンチセンス核酸の治療上の有用性
Deltaアンチセンス核酸を使用して、DeltaまたはNotchを発現することがわかっている
細胞型の悪性腫瘍またはその他の障害を処置(または予防)することができる。特定の態
様の場合悪性は頸管癌、乳癌もしくは結腸癌または扁平腺癌である。そのような発現を試
験することができる悪性の、腫瘍(新生物)および前新生物細胞としては前記5.8.1
および5.9.1節に記載したものがあるがこれらに限定されることはない。好ましい態
様においては一本鎖のDNAアンチセンスデルタオリゴヌクレオチドを使用する。
【0153】
DeltaまたはNotchRNAを発現する悪性(特に腫瘍)細胞型は業界で公知の各種方法で
同定することができる。そのような方法としては、DeltaまたはNotch特異的核酸とのハイ
ブリダイゼーション(たとえばノーザンハイブリダイゼーション、ドットブロットハイブ
リダイゼーション、insituハイブリダイゼーション)、その細胞型由来のRNAがin vi
troでノッチまたはデルタに翻訳され得る能力を観察すること、免疫検定、などがあるが
これらに限られるわけではない。好ましい態様においては、処置前に、患者から得た一次
腫瘍組織のノッチまたはデルタの発現を、たとえば免疫細胞化学またはin situハイブリ
ダイゼーションによって検定することができる。
【0154】
NotchもしくはDeltaRNAまたはタンパク質を発現するタイプの悪性を有する患者に、
薬剤として許容可能な担体中に有効量のDeltaアンチセンス核酸を含む本発明の薬剤組成
物(5.12節参照)を投与することができる。
【0155】
特定の障害または状態の処置に有効であるDeltaアンチセンス核酸の量はその障害また
は状態の性質に依存しており、標準的な臨床技術によって決定することができる。可能な
場合、ヒトで試験して使用する前に、処置すべき腫瘍タイプのアンチセンス細胞傷害性を
、in vitroで、その後、有用な動物モデル系で、決定するのが望ましい。
【0156】
特定の態様においては、Deltaアンチセンス核酸を含む薬剤組成物をリポソーム、微粒
子、またはマイクロカプセルを介して投与する。本発明のさまざまな態様においては、そ
のような組成物を用いてDeltaアンチセンス核酸の持続放出を達成するのが有用であり得
る。特定の態様の場合、特定の同定可能な腫瘍抗原に対する抗体を介して標的に向かわし
めるリポソームを利用するのが望ましいかもしれない(Leonetti et al.,1990,Proc.Na
tl.Acad.Sci.U.S.A.87:2448-2451;Renneisen et al.,1990,J.Biol.Chem.265:1
6337-16342)。
【0157】
5.12.治療/予防用投与および組成物
本発明は、本発明の治療薬を有効量で被検体に投与することによる処置(および予防)
の方法を提供する。好ましい局面においては、この治療薬は実質的に精製されている。被
検体は、牛、豚、鶏などのような動物をはじめとする(しかしこれらに限定されない)動
物が好ましく、哺乳類が好ましく、ヒトが最も好ましい。
【0158】
各種の送達系が知られており、本発明の治療薬を投与するために使用することができる
。たとえば、リポソーム、微粒子、マイクロカプセルに封入する、組換え細胞によって発
現させる、受容体に媒介されるエンドサイトーシス(たとえば、Wu and Wu,1987,J.Bi
ol.Chem.262:4429-4432参照)、レトロウィルスまたはその他のベクターの一部として
治療薬の核酸を構築する、などである。導入方法としては、皮内、筋肉内、腹膣内、静脈
内、皮下、鼻腔内、硬膜、および経口経路があるがこれらに限定されない。化合物は便利
な任意の経路、たとえば、注入または濃縮塊注射により、上皮もしくは粘膜皮膚ライニン
グ(lining)(たとえば、口腔粘膜、直腸および腸粘膜、など)を通る吸収により投与可能で
、他の生物活性剤と共に投与してもよい。投与は全身的または局所的に行うことができる
。また、本発明の薬剤組成物を適切な経路、たとえば心室内および鞘内注射によって中枢
神経系に導入することが望ましいことがある。心室内注射は、心室内カテーテル、たとえ
ばOmmayaリザーバー(reservoir)のようなリザーバーに接続した心室内カテーテルによっ
て容易にすることができる。また、たとえば吸入器や噴霧器およびエアゾール剤を用いた
製剤を使用することによって、肺投与も使用できる。
【0159】
特定の態様では、処置が必要とされる部分に本発明の薬剤組成物を局所的に投与するの
が望ましいことがある。これは、たとえば(限定ではないが)、手術中の局所注入、たと
えば手術後の創傷包帯と共にする局所適用、注射、カテーテル、座薬、インプラント(こ
れは多孔質、非多孔質またはゼラチン状の材料、たとえばsialastic膜のような膜、また
は線維である)を用いて達成できる。ひとつの態様では、悪性腫瘍または腫瘍もしくは腫
瘍発現前の組織の部位(または前の部位)に直接注射することによって投与することがで
きる。
【0160】
別の態様では、治療薬を小胞、特にリポソームに入れて送達することができる(Langer
,Science249:1527-1533(1990); Treatら,in Liposomes in the Therapy of Infectiou
s Disease andCancer,Lopez-BeresteinおよびFidler(eds.),Liss,New York,pp.353
-365(1989);Lopez-Berestein,ibid.,pp.317-327;参照。概論としてはibid.参照)。
【0161】
さらに別の態様の場合治療薬は制御放出系で送達することができる。ひとつの態様では
ポンプが使用できる(Langer,上掲;Sefton,CRC Crit.Ref.Biomed.Eng.14:201(1987
); Buchwaldら,Surgery88:507(1980); Saudek ら,N.Engl.J.Med.321:574(1989参
照))。別の態様ではポリマー性の材料を使用することができる(Medical Applications
ofControlled Release,LangerおよびWise(eds.),CRC Pres.,Boca Raton,Florida(1
974);Controlled Drug Bioavailability,Drug Product DesignおよびPerformance,Smo
len andBall(eds.),Wiley,New York(1984); RangerおよびPeppas,J.Macromol.Sci
.Rev.Macromol.Chem.23:61(1983)参照。またLevyら,Science228:190(1985); Durin
gら,Ann.Neurol.25:351(1989);Howardら,J.Neurosurg.71:105(1989)も参照のこ
と)。さらに別の態様においては、制御放出系を治療標的、すなわち脳の近くに配置する
ことができ、従って全身投与量の一部しか必要としない(たとえば、Goodson,in Medica
l Applicationsof Controlled Release,上掲,vol.2,pp.115-138(1984)参照)。
【0162】
他の制御放出系がLangerの文献によって議論されている(Science249:1527-1533(1990)
)。
【0163】
治療薬がタンパク質の治療薬をコードしている核酸である特定の態様の場合、その核酸
をin vivoで投与してそれがコードしているタンパク質の発現を促進することができる。
たとえば、レトロウィルスベクター(米国特許第4,980,286号参照)を使用することによ
り、その核酸を適当な核酸発現ベクターの一部として構築し、それを細胞内に組込まれる
ようにして投与する。または直接注射したり、微粒子衝撃(たとえば遺伝子銃(gun)、Bio
listic,Dupont)を使用したり、または脂質もしくは細胞表面受容体もしくはトランスフ
ェクト剤で被覆したり、または核の中に入って行くことが知られているホメオボックス様
ペプチドに結合させて投与したりする(たとえば、Joliotら,1991,Proc.Natl.Acad.
Sci.USA88:1864-1868)。あるいはまた、核酸治療薬を細胞内に導入し、相同組換えに
よって発現のために宿主細胞DNA内部に組込ませることができる。
【0164】
特定の障害の処置または予防に関する特定の態様の場合、次の投与形態を使用するのが
好ましい。
【0165】
【0166】
本発明はまた薬剤組成物も提供する。そのような組成物は、治療上有効な量の治療薬と
、薬剤として許容される担体とを含んでいる。特定の態様の場合、「薬剤として許容され
る」という用語は、動物、特にヒトに使用するものとして、米国の連邦政府もしくは州政
府の規制機関によって認可されているかまたは米国薬局方もしくは他の一般に容認されて
いる薬局方に収載されていることを意味する。「担体」という用語は、治療薬を投与する
ときに一緒に用いる希釈剤、アジュバント、賦形剤またはビヒクルを指す。そのような薬
剤担体は、水および石油、動物、植物もしくは合成起源のものを始めとする油、たとえば
ピーナッツ油、大豆油、鉱物油、ごま油などのような無菌の液体でありうる。薬剤組成物
を静脈内に投与するときには水が好ましい担体である。生理食塩水ならびにデキストロー
スおよびグリセロール水溶液もまた、特に注射用溶液の液体担体として使用することがで
きる。適切な薬剤賦形剤としては、澱粉、グルコース、ラクトース、スクロース、ゼラチ
ン、麦芽、米、小麦粉、チョーク、シリカゲル、ステアリン酸ナトリウム、モノステアリ
ン酸グリセロール、タルク、塩化ナトリウム、乾燥スキムミルク、グリセロール、プロピ
レン、グリコール、水、エタノールなどがある。所望により、該組成物はまた、少量の湿
潤剤もしくは乳化剤、またはpH緩衝剤も含有することができる。これらの組成物は溶液
、懸濁液、エマルション、錠剤、ピル、カプセル、粉末、徐放性製剤などの形態をとるこ
とができる。組成物は通常のバインダーおよびトリグリセリドのような担体と共に座薬と
して調合することができる。経口製剤は薬剤グレードのマンニトール、ラクトース、澱粉
、ステアリン酸マグネシウム、サッカリンナトリウム、セルロース、炭酸マグネシウムな
どのような標準的な担体を含むことができる。適した薬剤担体の例はE.W.Martinによっ
て「Remington´sPharmaceutical Sciences」に記載されている。このような組成物は患
者に適切な投与形態とするのに適した量の担体と共に治療上有効量の治療薬、好ましくは
精製形態の治療薬を含有している。この製剤は投与形態に適していなければならない。
【0167】
好ましい態様の場合、組成物は、通常の手法に従って、人間に静脈内投与するのに適し
た薬剤組成物として調合する。通常静脈内投与用の組成物は無菌の等張水性緩衝液の溶液
である。必要であれば組成物は、可溶化剤および注射部位の痛みを和らげるリグノカイン
のような局所麻酔薬とを含んでいてもよい。一般に、成分は別々に、または単位投与形態
として、たとえば活性剤の量を指示したアンプルまたはサシェットのような気密に密閉し
た容器に入れた凍結乾燥粉末または無水の濃縮物として一緒に供給される。組成物を注入
によって投与しようとする場合、無菌の薬剤グレードの水または生理食塩水を含有する注
入瓶として分配することができる。組成物を注射によって投与する場合、成分が投与前に
混合されるように注射用の無菌水または生理食塩水のアンプルを提供することができる。
【0168】
本発明の治療薬は中性形態または塩形態として調合することができる。薬剤として許容
できる塩には、塩酸、リン酸、酢酸、シュウ酸、酒石酸などから誘導されるもののように
遊離のアミノ基との間で形成されるもの、および水酸化ナトリウム、水酸化カリウム、水
酸化アンモニウム、水酸化カルシウム、水酸化第二鉄、イソプロピルアミン、トリエチル
アミン、2‐エチルアミノエタノール、ヒスチジン、プロカインなどから誘導されるもの
のように遊離のカルボキシル基との間に形成されるものがある。
【0169】
特定の障害または症状の処置に有効な本発明の治療薬の量はその障害または症状の性質
に依存し、標準的な臨床技術によって決定することができる。また、場合によって、in v
itro検定を利用して最適の投与範囲を同定するのに役立てることができる。本製剤中に使
用するべき正確な量もまた投与経路およびその疾患または障害の重さに依存し、医者の判
断および各患者の状況に応じて決定するべきである。しかし、静脈内投与に適した投与範
囲は一般に体重1kgあたり活性化合物約20〜500μgである。鼻腔内投与に適した投
与範囲は一般に体重1kgあたり約0.01pg〜1mgである。有効な投与量はin vitroまた
は動物モデル試験系から得られる用量−反応曲線から推定すればよい。
【0170】
座薬は一般に0.5〜10重量%の範囲で活性成分を含有し、経口製剤は活性成分を1
0〜95%含有するのが好ましい。
【0171】
本発明はまた、本発明の薬剤組成物の成分を1以上充填した1以上の容器を含む薬剤パ
ックまたはキットも提供する。場合によって、そのような容器には、薬剤または生物学的
生成物の製造、使用または販売を規制する政府機関によって規定された形の注意書きを付
けることができる。そのような注意書きは人間への投与に対する製造、使用または販売の
認可を示すものである。
【0172】
5.13.診断用途
デルタタンパク質、類似体、誘導体、およびこれらのサブ配列、Delta核酸(およびそ
れに相補的な配列)、抗デルタ抗体には診断用途がある。このような分子は、免疫検定の
ような検定法で使用して、デルタ発現に影響する各種症状、疾患および障害を検出、予知
、診断、もしくは追跡し、またはその処置を追跡することができる。特に、そのような免
疫検定は、患者から得たサンプルを免疫特異的な結合が起こり得るような条件下で抗デル
タ抗体と接触させ、その抗体による任意の免疫特異的結合の量を測定することからなる方
法によって実施する。特定の局面においては、組織切片における、抗体のそのような結合
を、好ましくは抗ノッチ抗体の結合と組合せて用いて、ある疾患状態における異常なノッ
チおよび/またはデルタ局在化または異常なレベルのノッチ−デルタの同時局在化を検出
することができる。ある特定の態様においては、デルタに対する抗体を用いて、患者の組
織または血清サンプルで、デルタの存在を検定することができ、ここで異常なレベルのデ
ルタは疾患状態の指標となる。内因性のノッチタンパク質におけるデルタ結合能力の異常
なレベル、または内因性のデルタタンパク質におけるノッチ(または他のデルタリガンド
)に対して結合する能力の異常なレベルは、細胞運命の障害(たとえば癌など)の指標と
なり得る。「異常なレベル」とは、その身体の一部から、またはその障害をもたない被検
者から得た類似のサンプル中に存在するレベルまたはこのレベルを表わす標準レベルと比
べて増大したかもしくは低下したレベルを意味する。
【0173】
使用することができる免疫検定には、ウェスタンブロット、放射線免疫検定、ELIS
A(固相酵素免疫検定)、「サンドイッチ」免疫検定、免疫沈降検定、沈降反応、ゲル核
酸沈降反応、免疫核酸検定、凝集検定、補体結合検定、イムノラジオメトリックアッセイ
、免疫蛍光検定、プロテインA免疫検定、などのような技術を用いる競合検定系および非
競合検定系があるがこれらに限定されるわけではない。
【0174】
Delta遺伝子および関連する核酸配列およびサブ配列、たとえば相補的配列、ならびに
その他の局所周期性遺伝子配列もまたハイブリダイゼーションアッセイに使用することが
できる。Delta核酸配列、または少なくとも約8個のヌクレオチドを含むそのサブ配列は
ハイブリダイゼーションプローブとして使用することができる。ハイブリダイゼーション
アッセイは前記したようにデルタの発現および/または活性の異常な変化を伴なう症状、
障害または疾患状態を検出、予知、診断または追跡するのに使用することができる。特に
、そのようなハイブリダイゼーションアッセイは、核酸を含有するサンプルを、ハイブリ
ダイゼーションが起こり得る条件下で、DeltaDNAまたはRNAとハイブリダイズする
ことができる核酸プローブと接触させ、その結果生じるハイブリダイゼーションを検出ま
たは測定することからなる方法によって実施する。
【0175】
さらに、デルタはノッチと結合するので、デルタまたはその結合部分は、サンプル中の
ノッチの存否および/またはその量を検定するのに、たとえば、結腸癌や子宮頸癌のよう
な増大したノッチ発現を示す悪性腫瘍をスクリーニングするのに、使用することができる
。
【実施例】
【0176】
6.ヒヨコのDeltaホモログは予定ニューロンで発現する
本明細書に記載したように、本発明者らはヒヨコのDeltaホモログ、C-Delta-1を単離し
特性決定をした。本発明者らは、C-Delta-1が、神経組織発生の間に新しい細胞が生まれ
、その運命が決定されるときに予定ニューロンで発現されることを示す。ヒヨコにおける
本発明者らのデータは、デルタ/ノッチシグナリングメカニズムと神経組織発生における
その役割とは両方とも脊椎動物で保存されていることを示唆している。
【0177】
6.1.C-Delta-1のクローニング
本発明者らは、ポリメラーゼ連鎖反応(PCR)と縮重したオリゴヌクレオチドプライ
マー(図1a、1bおよび2、配列番号1、2、3および4)を用いてヒヨコのDeltaホ
モログ(C-Delta-1)を同定した。それぞれハエデルタタンパク質配列FGFTWPGT(配列番
号21)およびNGGTCID(配列番号22)に相当する縮重したオリゴヌクレオチドプライ
マーTTCGGITT(C/T)ACITGGCCIGGIAC(配列番号19)およびTCIATGCAIGTICCICC(A/G)TT(
配列番号20)を用いたPCRによってC-Delta-1をクローン化した(Vassinら,1987,EM
BO J.6:3431-3440;Kopczynskiら,1988,Genes Dev.2:17 23-1735)。最初の反応では
、50μlの供給緩衝液(Perkin-Elmer)中で、4〜6胚期に由来する第一鎖オリゴ‐d(
T)でプライムしたcDNA50ng、各プライマー1μg、0.2mMdNTP、2UのT
aqポリメラーゼを使用した。94℃/30秒、50℃/2分、72℃/2分で40サイ
クルの増幅を行なった。増幅したDNA断片をアガロースゲルで分離し、Bluescript pKS
-(Stratagene)でクローン化し、配列決定した。2つのDeltaホモログが同定された。その
うちのひとつ(C-Delta-1)は神経系で発現される。神経系で発現されるホモログで、C-D
elta-1遺伝子の3′端における交互スプライシング事象のために、コードされているタン
パク質のカルボキシ末端が異なる2つの変異体が同定された。コードされているタンパク
質のひとつは、他方のコードされているタンパク質と比べて、カルボキシ末端に12個の
余分なアミノ酸を有する。このコードされている短い方の変異体の配列を配列番号2に示
す。配列番号3によりコードされており、かつ配列番号4のアミノ酸配列で同定される長
い方の変異体は配列番号2のアミノ酸配列と3′端にある12個の余分なアミノ酸(SIPPG
SRTSLGV)から成っている。この長い方の変異体を以下に記載する実験に用いた。RNAを
アフリカツメガエル(Xenopusoocytes)に注入して生物活性を試験した場合変異体のいず
れも同じ生物活性をもっていた。
【0178】
C-Delta-1に対応するDNA断片を用いて、17期の脊髄cDNAライブラリーをスク
リーニングし、いくつかの完全長クローンを得、配列決定した。類似の方法によりヒヨコ
のC-Notch-1遺伝子に由来するDNA断片を増幅した(データは示さない)。部分配列デ
ータと発現パターンは齧歯類のノッチ−1遺伝子との密接な類似性を示している(Weinmas
terら,1991,Development113:199-205; Weinmasterら,1992,Development 116:931-94
1; Lardelli& Lendahl,1993,Exp.Cell Res.204:364-372)。配列はWisconsin GCGセ
ットプログラムを用いて解析した。ヒヨコデルタ1mRNAのGenBank受託番号はU265
90である。C-Delta-1のDNA配列はショウジョウバエデルタと構造的に同族である(
図3、4)がデルタ関連Serrateタンパク質(これも本発明者らがクローン化したもので
あるがデータは示さない)の脊椎動物ホモログとは明らかに異なる722個のアミノ酸の
タンパク質に相当している。C-デルタ-1はその細胞外領域に推定膜貫通ドメイン、シグナ
ル配列および8つのEGF様反復配列を含有している(ショウジョウバエデルタより反復
配列が1つ少ない)。C-デルタ-1のアミノ末端ドメインはノッチに対するin vitro結合に
必要かつ充分であると記載されている(Muskavitch,1994,Dev.Biol.166:415-430)ハエ
のデルタタンパク質中の類似のドメインと密接に関連している。この保持された領域は、
ノッチ様タンパク質の推定リガンドのファミリーの公知のすべてのメンバーにより共有さ
れている(Deltaand Serrate in Drosophila; Lag-2 and Apx-1 in Caenorhabditis eleg
ans)(Hendersonら,1994,Development120:2913-2924; Taxら,1994,Nature 368:150-1
54; Flemingら,1990,GenesDev.4:2188-2201; Thomasら,1991,Development 111:749
-761; Melloら,1994,Cell77:95-106)いわゆるDSLモチーフ(図4)(Hendersonら
,1994,Development120:2913-2924; Tax ら,1994,Nature 368:150-154)を含んでい
る。二番目のシステインに富むN末端領域はハエとヒヨコのタンパク質で保存されている
が、関連するC.elegansタンパク質にはない(図4)。C-デルタ-1と81%同一であり上
記の構造モチーフのすべてを示す(図3)タンパク質をコードしているアフリカツメガエ
ルDelta-1ホモログX-Delta-1もクローン化した。ノッチ結合に重要であると同定されたド
メイン(Mustavitch,1994,Dev.Biol.166:415-430)を含めてヒヨコとハエのデルタタン
パク質間の保存構造は、C-デルタ-1がヒヨコのノッチタンパク質に対するリガンドとして
機能すること、および外側抑制(lateral inhibition)のデルタ/ノッチに媒介されるメカ
ニズムがヒヨコで働き得ることを示唆している。
【0179】
6.2.C-Delta-1とC-Notch-1 の発現は神経組織発生開始と相関している
ショウジョウバエの神経組織発生の間にDeltaは神経前駆体で一時的に発現され、隣接
するNotch発現細胞が同様に神経になるのを抑制する(Haenlinら,1990,Development110
:905-914; Kooh ら,1993,Development117:493-507)。C-Delta-1がヒヨコの神経組織発
生の間に同様に働くとすると、ニューロン前駆体細胞が決定されるようになるにつれてC-
Delta-1が同様にニューロン前駆体細胞でも一時的に発現されるはずである。発生するC
NSにおけるC-Delta-1の発現の解析は、これが実際に起こっていることを示している。
【0180】
要約すると、ホールマウントin situハイブリダイゼーションを実施した。ホルムアル
デヒドで固定した胚をプロテアーゼで処理し、4%ホルムアルデヒド/0.1%グルタル
アルデヒドで再固定した。DIGで標識したRNAプローブとのハイブリダイゼーション
を、0.2%Tween-20と0.5%CHAPSを含有する緩衝液中で、ストリンジェントな
条件(1.5×SSC、50%ホルムアミド、65℃、pH5)下で実施した。洗浄した
胚をBoehringerBlocking Reagent(ベーリンガーブロッキング試薬)で処理し、アルカ
リホスファターゼと結合した抗‐DIG抗体中で一晩インキュベートした。充分に洗浄し
た後胚を30分〜一晩染色した。図5eの胚はハイブリダイゼーション後ワックスで切開
した(wax-sectioned)。
【0181】
神経板におけるC-Delta-1の発現は6〜7期(31時間、0/1体節)に体節形成前の
中胚葉の直前の散乱細胞(scattered cells)で最初に検出される(図5b、5c)。この
領域は中/後部(mid/posterior)後脳になり、そこでは、最も初期に分化したCNSニュ
ーロンが、最初のC-Delta-1発現の6時間後の9期(31時間、7〜9体節)に、神経繊
維抗体により始めて検出される(表2)(Sechrist & Bronner-Fraser,1991,Neuron 7:
947-963)。
【表2】
【0182】
神経組織発生が進むにつれて、C-Delta-1の発現がニューロン分化の時空パターンを予
示し続け(表2)、脊髄に沿って後に、また中脳および前脳に向かって前方に拡大する(
図5d、5e)。たとえば、8期の脊髄における最も後に発現する細胞は将来の6番目の
体節のレベルにあり、これはそのレベルにある最初のニューロンが神経繊維抗原を発現す
る6〜8時間前である(Sechrist & Bronner-Fraser,1991,Neuron 7:947-963)(表2)
。表2に示されているように、C-Delta-1の発現の出現は、最初のニューロン前駆体が分
裂サイクルから離脱したすぐ後で、得られる分化性ニューロンで神経繊維(NF)抗原が
出現する前である。
【0183】
中部後脳は耳の原基のレベルである菱脳の神経小片4〜6を含んでおり、後部後脳は菱
脳の神経小片7および8ならびに体節1〜4を含んでいる。細胞分裂からの離脱のタイミ
ングのデータと神経繊維の発現のデータはSechristら,1991,Neuron7:947-963からとっ
た。いずれの場合も、C-Delta-1は散乱細胞で一様なC-Notch-1発現のドメイン内で発現す
る(図5a)。
【0184】
6.3.C-Delta-1の局在化と経時発現
C-Delta-1の局在化と経時変化は、この遺伝子が神経組織発生の初期の段階で切り替え
られる(switched)こと、およびC-Delta-1を発現する細胞がまだ分化マーカーを示し始め
ていない将来のニューロンであることを示している。この仮説を試験するために、本発明
者らは、将来のニューロンが、初期神経管における非サイクル細胞(non-cycling cells)
のみであるというSechristとBronner-Fraserの観察結果(Sechrist& Bronner-Fraser,19
91,Neuron 7:947-963)を用いた。それらの細胞は神経繊維抗原を発現する11〜15時
間前にその最終S相を終了し(表2)、その核は、最後の有糸分裂の完了後、他の細胞核
すべてがS相にある神経上皮の基底表面付近に特徴的な位置をとる(Sechrist & Bronner-
Fraser,1991,Neuron7:947-963; Martin & Langman,1965,J.Embryol.Exp.Morphol
.14:23-35)(図6a)。本発明者らは7〜9期の胚をブロモデオキシウリジン(Brd
U)で標識し、BrdU取込みとC-Delta-1発現に関して二重に染色した。C-Delta-1を発
現する細胞の95%は未標識であった。それらの核は主として基底表面付近に位置してい
たが、そこではほとんどの他の核がBrdUで標識された(図6b、6c)。PBS中の
0.1mMのBrdU75μlを、insituハイブリダイゼーション用に固定する前に38
℃で2〜4時間インキュベートした7〜9期の胚上に滴下した。、15μmのクリオスタ
ット切片を、ほぼStrahleらの方法(Strahleら,1994,TrendsIn Genet.Sci.10:75-76)
に従って、DIGで標識したRNAプローブとハイブリダイズした。染色後スライドをP
BS中で洗浄し、BrdU免疫検出(immunodetection)用に処理した(Biffoら,1992,His
tochem.Cytochem.40:535-540)。FITCと結合したヤギ抗‐マウス二次抗体(Cappel)
を用いて抗BrdU(1:1000、Sigma)を検出した。C-Delta-1の発現をDIC顕微
鏡法、および慣用の共焦点蛍光顕微鏡法によるBrdU標識化で検査した。これらの結果
は、C-Delta-1が細胞サイクルから離脱しており、実際に将来のニューロンであるに違い
ない細胞で発現されることを示している。二、三のBrdU-/C-Delta-1-細胞は基底ゾ
ーン外にその核を有する。これらは、BrdUに暴露された後すぐに最終S相を終了し、
先端に移動してその最終有糸分裂を完了し、C-Delta-1発現に切り替わる細胞であり得る
。C-Delta-1はまた、後期の神経管および末消神経系でも発現する。ここでも、発現のタ
イミングと発現細胞の位置は、これらの細胞がまだ分化し始めていないニューロン前駆体
であることを示唆している(データは示さない)。このように、C-Delta-1の発現は将来
のニューロンに対する最も早く知られたマーカーであるように思われる。
【0185】
また、C-デルタ-1とC-Serrate-1の転写パターンは胚の多くの領域におけるC-ノッチ-1
の転写パターンとオーバーラップしており(データは示さない)、これはC-ノッチ-1がシ
ョウジョウバエ内のノッチと同様に両者のタンパク質に対する受容体であることを示唆し
ている。特に、3つのすべての遺伝子が発生する中枢神経系の神経原性の領域で発現され
、ここにおいて驚くべき関係が見られる。すなわち、C-Serrate-1とC-デルタ-1の両方の
発現はC-ノッチ-1発現のドメインに限定されるが、このドメイン内でC-Serrate-1とC-デ
ルタ-1の領域は正確に相補的である。オーバーラップする発現パターンは、それらとノッ
チとの機能的相関関係の保存を示唆しており、ヒヨコ、特にその中枢神経系の発生がいろ
いろな位置におけるC-ノッチ-1といろいろなリガンドとの相互作用を含んでいることを暗
示している。
【0186】
6.4.考察
C-デルタ-1のアフリカツメガエルホモログを同様な方法でクローン化した。簡単にいう
と、X-デルタ-1のPCR断片を単離して配列決定した。次にこの断片を用いてX-デルタ-1
の完全長クローンを同定した。X-デルタ-1の発現パターンを検討した。X-デルタ-1は散乱
細胞において、一次ニューロン前駆体が生成している神経板のドメイン内で発現すること
が示された。これは、X-デルタ-1を発現する細胞が将来の一次ニューロンであることを示
唆している。さらに、X-デルタ-1はまた、神経組織発生の他の部位および時期、たとえば
前部神経板および神経原性板ならびに二次ニューロンが生成した場合の神経管発生の後の
段階でも発現する。異所性のX-Delta-1活性は一次ニューロンの産生を抑制し、内因性のX
-Delta-1活性との干渉の結果一次ニューロンの過剰産生が生じた。これらの結果は、X-デ
ルタ-1が将来のニューロンによって隣接細胞に送達される側方抑制を媒介していることを
示している。アフリカツメガエル卵におけるX-デルタ-1の異所性発現は一次神経組織発生
を抑制すること、また、細胞質ドメインのアミノ酸を2つだけ保持している末端を切断し
た(truncated)X-デルタ-1タンパク質の異所性発現が内因性のシグナリングに干渉し、余
分な細胞をニューロン前駆体として発生させることが示された(Chitnisら,Nature(印刷
中))。予備的な証拠は、C-Delta-1がアフリカツメガエルの胚で発現されると類似の抑制
作用をもつことを示している(データは示さない)。本発明者らは、C-Delta-1がショウ
ジョウバエとアフリカツメガエルの対応するタンパク質と同様に神経組織発生の間を通し
て側方抑制を媒介して、任意の時に神経宿命になり得る細胞の割合を制限するということ
を提案する。C-デルタ-1は通常神経組織発生中CNSでもPNSでも多くの他の部位で、
たとえば発生する耳でも発現される。CNSにおいては、C-ノッチはE5ヒヨコ後脳の脳
室ゾーンにおいて神経管の内腔に隣接する分裂細胞で発現されることが示されている。C-
デルタ-1は、分裂を止めてしまっておりニューロン前駆体細胞となろうとしている細胞の
隣接層で発現する。したがって、デルタ/ノッチシグナリングはここにおいて他の神経組
織におけると同様に作用して未決のサイクルニューロン幹細胞の集団を維持することがで
きよう。
【0187】
7.マウスDeltaホモログの単離し特性決定
M-Delta-1と名付けたマウスDeltaホモログを以下のようにして単離した。
【0188】
マウスDelta-1
遺伝子由来組織:8.5日および9.5日齢のマウス胚RNA
単離法:
a)ランダムにプライムした、上記RNAに対するcDNA
b)上記cDNAのPCRには以下のものを使用した。
【0189】
PCRプライマー1:GGITTCACITGGCCIGGIACNTT(配列番号23)[デルタ様である
がSerrate様ではないタンパク質に対して特異的な領域であるGFTWPGTF(配列番号24)
をコードしている]
PCRプライマー2:
GTICCICC(G/A)TT(C/T)TT(G/A)CAIGG(G/A)TT(配列番号25)[EGF様反復配列の多
くの中に存在する配列であるNPCKNGGT(配列番号26)をコードしている]
増幅条件:50ngのcDNA、1μgの各プライマー、0.2mMのdNTP、1.
8UのTaq(Perkin-Elmer)、50μlの供給緩衝液中。94℃/30秒、45℃/2分
、72℃/1分の40サイクル(各サイクル2秒ずつ延長)。
【0190】
増幅された断片は約650塩基対の断片であり、これを部分的に配列決定してC-Delta-
1との相関関係を決定した。
【0191】
c)マウスの11.5日のcDNAライブラリー(Clontech)をスクリーニングした。い
くつかの陽性クローンのうち、ひとつ(pMDL2、インサートのサイズは約4kb)が完全
なタンパク質コード領域を含んでいた。このDNA配列を完全に決定した。
【0192】
図7(配列番号11)はM-Delta-1DNAを含有する単離されたクローンのヌクレオチ
ド配列を示す。
【0193】
図8(配列番号12)はM-デルタ-1の予想されるアミノ酸配列を示す。
【0194】
図9はM-デルタ-1とC-デルタ-1に対して予想されるアミノ酸配列を並べて示す。同一の
アミノ酸を四角で囲って示す。広範囲の配列相同性を示している。コンセンサス配列を下
に示す(配列番号13)。
【0195】
発現パターン:発現パターンは、C-デルタ-1に対して、特に体節形成前の中胚葉、中枢
神経系、末消神経系および腎臓で観察されたものとほぼ同じであることが確認された。
【0196】
8.ヒトデルタホモログの単離と特性決定
H-デルタ-1(HD1)と名付けたヒトデルタ-1ホモログを以下のようにして単離した。
【0197】
インサートのサイズが100〜150kbの範囲であるヒトゲノムライブラリーをマウ
スデルタ-1(M-Delta-1)遺伝子のEcoRI断片で探索した。このライブラリーから、E
coRI断片とハイブリダイズしたゲノムヒトPACクローンを単離した。次に、2つの
縮重しているオリゴヌクレオチドを用いて、ゲノムヒトPACクローンの断片をPCRに
より増幅した。縮重オリゴは次の通り。
【0198】
5′ACIATGAA(C/T)AA(C/T)CTIGCIAA(C/T)TG(配列番号27)[TMNNLANC(配列番号28
)をコード]および
3′AC(A/G)TAIACIGA(C/T)TG(A/G)TA(C/T)TTIGT(配列番号29)[TKYQSVYV(配列番号
30)をコード]または
3′GC(A/G/T)ATIAC(A/G)CA(C/T)TC(A/G)TC(C/T)TT(C/T)TC(配列番号31)[EKDECVIA
(配列番号32)をコード]。
【0199】
ニワトリとマウスのデルタ-1に対するcDNA配列に基づいて、5′オリゴと2つの異
なる3′オリゴを用いて、約354塩基対および387塩基対の断片がそれぞれ単離され
ると予想された。しかし、実際には、525塩基対と予想通りそれより30塩基対小さい
単一の単離物が2つ得られた。大きい方の単離物をジデオキシシークエンシングで配列決
定した。そのヌクレオチド配列を図10に示す(配列番号14)。また、図10には、増
幅したDNA断片の3つの異なるリーディングフレームに対して予想されたアミノ酸配列
(配列番号15、16、17)も示す。シークエンシングエラーのため、両方のプライマ
ー間で中断されてない充分な配列は同定されなかった。その結果、この得られたDNA配
列から直接アミノ酸配列を予想することはできない。しかし、図11に、ヒトデルタ-1(
上)(配列番号18)とヒヨコのデルタ-1(下)との間の肉眼で決定したアミノ酸配列相
同性を示す。シークエンシングエラーのため、この相同性は、相同領域を同定するために
3つの異なるリーディングフレームの間で切り替えることによって得た。
【0200】
完全長のH-デルタ-1(HD1)遺伝子を単離する目的で、大きい方の単離物(配列番号
14)をプローブとして用い、ヒト胎児脳プラスミドライブラリー(Clontech)をスクリー
ニングした。この結果4つの陽性プラークが得られた。これらの陽性プラークのうちの2
つ(HD13およびHD124)が再度のスクリーニングで生き残り、これをサザンブロ
ットで大きいヒトゲノム断片と陽性反応させた。これらの陽性クローンをEcoRIで消
化し、断片をBluescriptKS-ベクターに連結することによってサブクローン化した。T3
およびT7プライマーを用いたジデオキシシークエンスによりインサートのヌクレオチド
配列を得た。結果は、HD124がその両端でニワトリのデルタ-1と相同であるが、HD
13の一端は相同性を示さないことを示していた。一連の酵素による制限消化は2つのク
ローンで非常に類似したパターンを示した。これらはいずれもインサートが約2kbであ
ったがHD13の3′端が異なっていた。
【0201】
HD13とHD124をBstXI、XbaI、HindIIIおよびXhoIで消化
し、制限断片をBluescriptKS-に挿入した後上記のようにして配列決定して内部配列を得
た。得られた配列は3′にHD1の約2000塩基をもっており、3′非コード領域に伸
びていた。HD13はHD124の中に含まれているが、HD13の5′端に付加された
配列はクローニングの際に人工的に入ったものであろう。
【0202】
こうして得られた配列はHD1の5′端を含有していなかったので、T細胞ライブラリ
ーおよび他の胎児脳ライブラリー(Lambda-Zap,Stratagene)でのハイブリダイゼーション
にHD124をプローブとして使用した。T細胞ライブラリーのスクリーニングでは陽性
のものが得られなかった。しかし、Lambda-Zapライブラリーのスクリーニングでは2つの
陽性クローンHD113とHD118が得られた。これらのクローンを、上記のようにE
coRIを用いてBluescriptKS-ベクターに挿入した。HD13およびHD124と比較
するためにインサートを一連の制限酵素で消化し、T3およびT7プライマーを用いて5
′端と3′端を配列決定した。HD113は、配列決定したときに公知のデルタと相同性
を示さないcDNAの小片のみであることが決定された。しかし、HD118は長さが3
kbであり、HD124の全体配列とさらに5′端の追加の配列を含んでいた。重なり欠
失を用いて一組のクローンをHD118から単離し、これらのクローンを自動シークエン
サーを用いてジデオキシシークエンスにかけた。図12Aは、クローンHD118から得
たヒトDeltaの部分ヌクレオチドコンティグ配列(配列番号33)を示す。シークエンシ
ングエラーのため、ヒトDeltaの中断されてない充分なヌクレオチド配列は決定されなか
った。図12Bに、ヒトDeltaの部分ヌクレオチドコンティグ配列(配列番号33)を一
番上に、また、その下に3つの異なるリーディングフレームで予想されるアミノ酸配列を
示す。2番目はリーディングフレーム1(配列番号34)、3番目はリーディングフレー
ム2(配列番号35)、4番目はリーディングフレーム3(配列番号36)である。
【0203】
配列の相同性はマウスデルタ-1アミノ酸配列を用いて肉眼で決定した。マウスアミノ酸
配列と最も相同性の高い配列は図12Bで四角で囲って示し、ヒトデルタ-1の予想された
アミノ酸配列を示す。得られた複合のアミノ酸配列を図14に示す。(図14では、ヒト
デルタ配列の中断されてないさまざまな部分をそれぞれ配列番号39〜65として表示し
てある。)シークエンシングエラーのため最高の相同性をもつリーディングフレームは配
列全体を通して同じではなく、配列中にエラーがある位置でシフトされていることに注意
されたい。
【0204】
また、ニワトリとマウスのデルタに対して肉眼で決定した相同性は、決定されたヒトDe
ltaヌクレオチド配列から推定したアミノ酸配列が、ヒトデルタ-1のN末端の100〜1
50アミノ酸のほぼすべてを含有していることを示している。
【0205】
図13に、マウスDelta-1のヌクレオチド配列(一番上、配列番号37)、図12Aお
よび12Bに示したヒトDelta-1のコーディングヌクレオチド配列(2番目、配列番号3
3)、そしてマウスとヒトのDeltaの間のヌクレオチドコンセンサス配列(3番目、配列
番号38)を示す。
【0206】
図12Aに示したヒトデルタ5′ヌクレオチド配列を含有するプローブを用いて、cD
NAライブラリーを探索してヒトデルタ遺伝子の5′端を単離する。一次陽性クローンを
得た後二次陽性物として確認する。二次陽性物を精製しさらに増殖させる。その後DNA
を単離し配列決定のためにサブクローン化する。
【0207】
本発明の範囲は本明細書に記載した特定の態様に限定されるものではない。実際、以上
の説明と添付した図面から、本明細書に記載したものに加えて本発明のさまざまな修正が
当業者には明らかであろう。そのような修正は添付の請求の範囲内に入るものと考えられ
る。
【0208】
本明細書でさまざまな引用をしたがその開示内容は引用したことによってその全体が本
明細書に含まれているものとする。
【特許請求の範囲】
【請求項1】
明細書に記載の発明。
【請求項1】
明細書に記載の発明。
【図1A1】


【図1A2】

【図1A3】

【図1B1】

【図1B2】

【図2】

【図3A】

【図3B】

【図4】
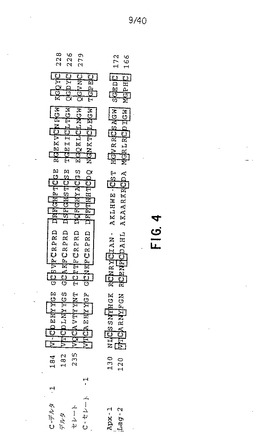
【図5A】

【図5B】

【図5C】

【図5D】

【図5E】
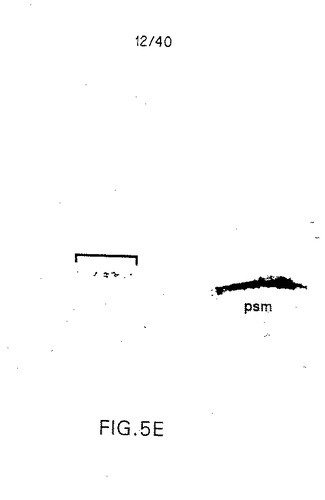
【図6A】
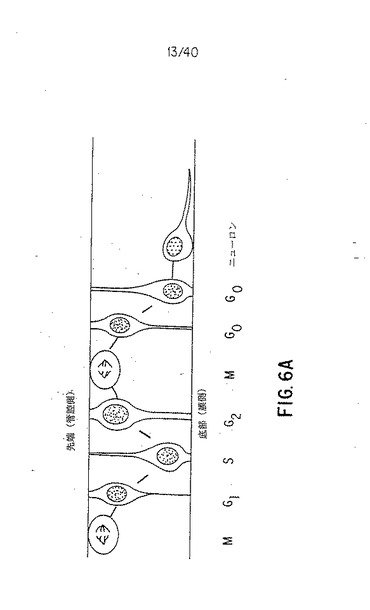
【図6B】

【図6C】

【図7A】

【図7B】

【図8】

【図9A】

【図9B】

【図10A】

【図10B】
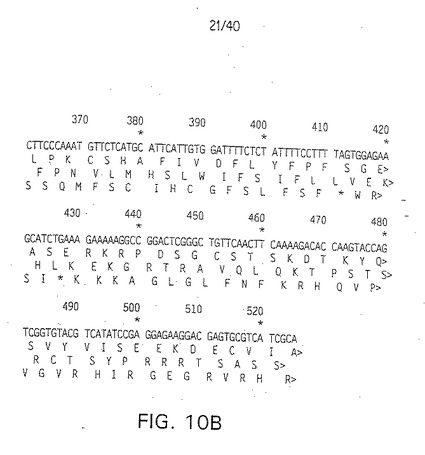
【図11】
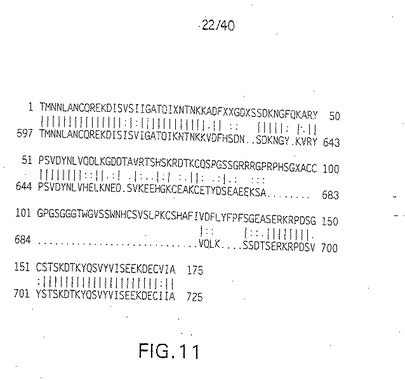
【図12A1】
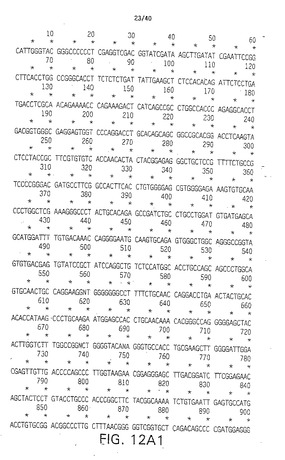
【図12A2】

【図12A3】
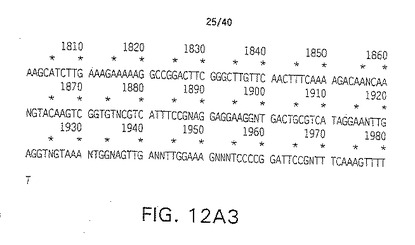
【図12B1】

【図12B2】

【図12B3】

【図12B4】
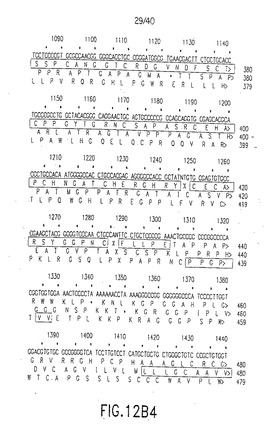
【図12B5】

【図12B6】
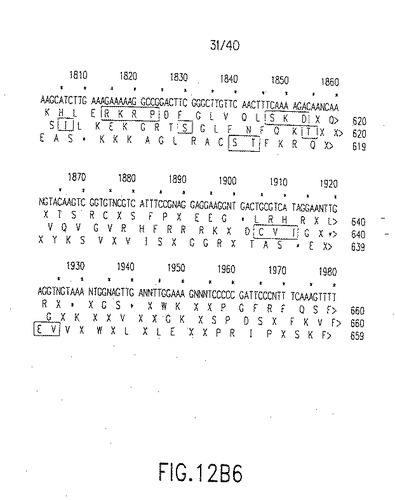
【図13A】

【図13B】

【図13C】

【図13D】

【図13E】

【図13F】

【図13G】

【図14A】

【図14B】

【図1A2】
【図1A3】
【図1B1】
【図1B2】
【図2】
【図3A】
【図3B】
【図4】
【図5A】
【図5B】
【図5C】
【図5D】
【図5E】
【図6A】
【図6B】
【図6C】
【図7A】
【図7B】
【図8】
【図9A】
【図9B】
【図10A】
【図10B】
【図11】
【図12A1】
【図12A2】
【図12A3】
【図12B1】
【図12B2】
【図12B3】
【図12B4】
【図12B5】
【図12B6】
【図13A】
【図13B】
【図13C】
【図13D】
【図13E】
【図13F】
【図13G】
【図14A】
【図14B】
【公開番号】特開2012−70735(P2012−70735A)
【公開日】平成24年4月12日(2012.4.12)
【国際特許分類】
【出願番号】特願2011−220536(P2011−220536)
【出願日】平成23年10月4日(2011.10.4)
【分割の表示】特願2008−179448(P2008−179448)の分割
【原出願日】平成8年6月28日(1996.6.28)
【出願人】(592210108)インペリアル キャンサー リサーチ テクノロジー リミテッド (5)
【出願人】(392019352)イェール ユニバーシティー (38)
【氏名又は名称原語表記】YALE UNIVERSITY
【Fターム(参考)】
【公開日】平成24年4月12日(2012.4.12)
【国際特許分類】
【出願日】平成23年10月4日(2011.10.4)
【分割の表示】特願2008−179448(P2008−179448)の分割
【原出願日】平成8年6月28日(1996.6.28)
【出願人】(592210108)インペリアル キャンサー リサーチ テクノロジー リミテッド (5)
【出願人】(392019352)イェール ユニバーシティー (38)
【氏名又は名称原語表記】YALE UNIVERSITY
【Fターム(参考)】
[ Back to top ]