
トランスフェクション方法及びそのための組成物
本発明は、Toll様受容体(TLR)を発現する細胞への、分子のターゲットされた送達に関する。本発明の態様は、TLRリガンドに連結した正荷電基を含む化合物を提供する。これらの化合物は、TLR発現細胞のトランスフェクションのインビトロ及びインビボの方法に有用である。本発明の他の態様は、遺伝子発現の抑圧及びDNAワクチン接種アプローチのためのそのような化合物の使用に関する。
【発明の詳細な説明】
【発明の詳細な説明】
【0001】
[発明の分野]
本発明は、一般的に、細胞生物学分野に関する。より具体的には、本発明は、細胞、特に、樹状細胞をトランスフェクションする方法、及び免疫応答を誘発する方法に向けられる。この目的を達成するために、本発明は、Toll様受容体(TLR)ファミリーの1つ又は複数のメンバーと相互作用する能力がある部分に連結した、核酸を結合する能力がある少なくとも1個の正荷電基を含む化合物を提供する。
【0002】
[発明の背景]
本明細書に用いられる場合、用語「由来する」は、特定された完全体(integer)が、特定の供給源から得ることができるが、ただし、必ずしもその供給源から直接というわけではないことを示すものと解釈されるべきである。
【0003】
本明細書を通して、文脈が別な意味を必要としない限り、単語「含む(comprise)」、又は「含む(comprises)」若しくは「含むこと(comprising)」などの語尾変化は、述べられた段階若しくは要素若しくは完全体、又は段階若しくは要素若しくは完全体の群を包含するが、ただし、いかなる他の段階若しくは要素若しくは完全体、又は要素若しくは完全体の群も排除するものではないことを意味するように理解されるだろう。
【0004】
本明細書に記載された本発明が、具体的に記載されたもの以外に変形及び改変を受ける余地があることを当業者は理解しているだろう。本発明が全てのそのような変形及び改変を含むことは理解されるべきである。本発明はまた、本明細書に、個々に又は集合的に、参照された又は示された段階、特徴、組成物、及び化合物の全部、並びに前記段階又は特徴のありとあらゆる組合せ又は任意の2つ以上を含む。
【0005】
本発明は、本明細書に記載された特定の例によって範囲が限定されるべきではない。機能的等価の生成物、組成物、及び方法は明らかに、本明細書に記載された本発明の範囲内である。
【0006】
本出願に引用された全ての参考文献は、本明細書に参照により明確に組み入れられている。
【0007】
本発明は、他に規定がない限り、分子生物学、微生物学、ウイルス学、組換えDNAテクノロジー、溶液中でのペプチド合成、固相ペプチド合成、及び免疫学の通常の技術を用いて、過度の実験なしに実施される。そのような手順は、例えば、参照により組み入れられている以下のテキスト、すなわち、
1.Sambrook、Fritsch、及びManiatis、Molecular Cloning:A Laboratory Manual、Cold Spring Harbor Laboratories、New York、第2版(1989)、I巻、II巻、及びIII巻の全体、
2.DNA Cloning:A Practical Approach、I巻及びII巻(D.N.Glover編、1985)、IRL Press、Oxford、テキストの全体、
3.Oligonucletoide Synthesis:A Practical Approach(M.J.Gait編、1984)IRL Press、Oxford、テキストの全体、並びに特に、Gait、1〜22ページ;Atkinsonら、35〜81ページ;Sproatら、83〜115ページ;及びWuら、135〜151ページによるその中の論文
に記載されている。
【0008】
本明細書中のいかなる先行技術への言及も、この先行技術がオーストラリアにおける共通一般知識の部分を形成するという認識でも、いかなる形の示唆でもなく、そのように解釈されるべきではない。
【0009】
脂質、タンパク質、ペプチド、DNA、多糖、及び/又はそれらの組合せ(例えば、リポ多糖、リポタンパク質)などの分子の、細胞内への送達は、多数の研究及び臨床の目的にとって有用である。例えば、研究者が遺伝子制御及び遺伝子発現、DNA−タンパク質相互作用又はタンパク質−タンパク質相互作用などの細胞内過程を研究するために、分子を細胞へ導入することが不可欠であることが多く、できる限り高効率でそのように行うことが望ましい。現在、研究者は、細胞への分子の送達、すなわち、細胞のトランスフェクションを、様々な効率をもつ様々な手段によって行う。細胞のトランスフェクション効率は、細胞型、細胞分裂の速度及び時期、並びにトランスフェクションされることになっている分子及び(1つ又は複数の)トランスフェクション試薬の両方の個々の性質を含むいくつかの因子に依存している。
【0010】
DNAワクチンは、樹状細胞(DC)などの抗原提示細胞によるDNAの取り込みを介して免疫応答を誘発すると考えられており、樹状細胞が、その後、内部移行したDNAにコードされる抗原を発現し、MHC分子との関連でペプチドとして免疫系に抗原を提示する。小動物モデルにおいて、DNAの投与は、防御免疫応答を誘導することに成功しているが、ただし、ヒト臨床試験において効力の低さが報告されており、免疫応答を誘導するのに高用量のDNAを必要とすることが多い(Kutzler,M.A.及びWeiner,D.B.2004 J Clin Invest、114(9)、1241〜1244)。アデノウイルス及びレトロウイルスは、遺伝子送達のためのベクターとして用いられているが、ヒトに用いるためのこれらのベクターの安全性に関して懸念が存在する(Buckley,R.H.2002 Lancet、360(9341)、1185〜1186)。今までのところ、比較的安全な非ウイルスベクターを用いる、DCのトランスフェクションは困難であることが証明されている。
【0011】
DNAは、正味の負電荷を帯びた分子である。より具体的には、DNAのバックボーン内のリン酸基が負電荷をもつ。したがって、正味の正電荷を有するカチオン性分子は、静電相互作用を介してDNAを吸着することができ、DNAの有望な担体である。そのようなカチオン性分子として、微粒子(Minigo,Gら 2007 Vaccine、25(7)、1316〜1327;Mollenkopf,H.J.ら 2004 Vaccine、22(21〜22)、2690〜2695)、ペプチド(Gratton,J.P.ら 2003 Nat Med、9(3)、357〜362;Riedl,P.ら 2006 Methods Mol Med、127、159〜169)、又はリポソーム(Jiao,Xら 2003 Hepatology、37(2)、452〜460;Ewert,K.ら 2002 J Med Chem、45(23)、5023〜5029)が挙げられる。
【0012】
しかしながら、単にDNAを抗原提示細胞に運ぶだけでは、その結果トランスフェクションが生じるには十分ではなく、抗原特異的応答を作動させるために、DNAの取り込みもまた存在しなければならない。免疫系の抗原提示細胞は、それらの細胞表面上にToll様受容体(TLR)を発現しており、そのTLRは、主として微生物に由来する様々なリガンドに結合する。例えば、TLR−2は細菌リポタンパク質を結合することが知られており、TLR−4は細菌リポ多糖を結合することが知られており、TLR−6は、TLR−1と共同して、脱アシル化細菌脂質を結合することが知られており、TLR−9は、CpG DNAに結合する。樹状細胞サブセットは、9個以上のそのようなTLRを発現することが示されている。DCの表面上の1個又は複数のTLRの会合が、細胞シグナル伝達経路を誘導し、それが、防御免疫の誘導に必要とされる、DCの成熟及び活性化をもたらすことができる。
【0013】
脂質部分、ジパルミトイル−S−グリセリルシステイン(Pam2Cys)は、マイコプラズマ ファーメンタンス(Mycoplasma fermentans)の細胞膜由来のMALP−2として知られた細菌リポタンパク質の合成類似体である。Pam2Cysは、TLR−2及びTLR−6の両方のリガンドである(Okusawa,Tら、Infect Immun 2004、72(3)、1657〜1665)。ペプチドエピトープに結合したPam2Cysを含むワクチンは、強い体液性応答及び細胞性応答を誘導することができる。ペプチドエピトープに結合したPam2CysによるTLR−2の会合の結果として、DC成熟、NF−κBなどの転写因子の活性化と、炎症誘発性サイトカインの分泌と、最終的にはエピトープ特異的ナイーブT細胞を活性化する、流入領域リンパ節へのDCの遊走とが生じる(Jackson,D.C.ら 2004 Proc Natl Acad Sci USA、101(43)、15440〜15445;Zeng,Wら 2002 J Immuol、169(9)、4905〜4912;Chua,B.Y.ら 2007 Vaccine、25(1)、92〜101)。
【0014】
[発明の概要]
本発明に繋がる研究において、本発明者らは、細胞に核酸をトランスフェクションする効率的な手段を生み出そうとした。それはまた、対象において、抗原提示細胞、特に樹状細胞をターゲットにし、体液性免疫応答及び細胞性免疫応答の両方を誘発する、DNAワクチン候補を作製しようとすることでもあった。
【0015】
第1態様において、本発明は、少なくとも1個のTLRリガンドに連結した正荷電基を含む化合物を提供する。
【0016】
第2態様において、本発明は、核酸と、少なくとも1個のTLRリガンドに連結した正荷電基を含む化合物とを含む複合体であって、上記核酸が、核酸と正荷電基との間の静電相互作用によって上記化合物と会合している、複合体を提供する。
【0017】
第3態様において、本発明は、トランスフェクション方法であって、核酸と、少なくとも1個のTLRリガンドに連結した正荷電基を含む化合物とを含む複合体に、TLRを発現する細胞を接触させる段階を含み、上記核酸が、核酸と正荷電基との間の静電相互作用によって上記化合物に会合している、方法を提供する。
【0018】
第4態様において、本発明は、抗原に対する免疫応答を生じさせる方法であって、核酸と、少なくとも1個のTLRリガンドに連結した正荷電基を含む化合物とを含む複合体を対象に投与する段階を含み、上記核酸が、核酸と正荷電基との間の静電相互作用によって上記化合物に会合しており、かつ上記核酸が上記抗原又はそのエピトープをコードする、方法を提供する。
【0019】
第5態様において、本発明は、抗原に対する免疫応答を生じさせる方法であって、核酸と、少なくとも1個のTLRリガンドに連結した正荷電基を含む化合物とを含む複合体をトランスフェクションされた細胞を対象に投与する段階を含み、上記核酸が、核酸と正荷電基との間の静電相互作用によって上記化合物に会合しており、かつ上記核酸が上記抗原又はそのエピトープをコードする、方法を提供する。
【0020】
第6態様において、本発明はまた、TLRを発現する細胞において遺伝子の発現を抑圧する方法であって、核酸と、少なくとも1個のTLRリガンドに連結した正荷電基を含む化合物とを含む複合体を対象に投与する段階を含み、上記核酸が、核酸と正荷電基との間の静電相互作用によって上記化合物に会合しており、上記核酸がsiRNA、shRNA、siRNAをコードするDNA、及びshRNAをコードするDNAからなる群より選択され、かつ上記遺伝子をターゲットとする、方法を提供する。
【0021】
さらには、対象における免疫応答の誘導のためのワクチンの製造、遺伝子異常又は遺伝子欠損をもつ対象の処置のための医薬の製造、遺伝子の異常な発現又は望ましくない発現を患っている対象の処置のための医薬の製造、及び細胞のトランスフェクションのための試薬の製造における、少なくとも1個のTLRリガンドに連結した正荷電基を含む化合物の使用を提供する。
【図面の簡単な説明】
【0022】
【図1】様々な合成された蛍光ペプチド構築物の図表示である。略語;R、アルギニン;K、リシン;L、直鎖;FL、フルオレセイン又はカルボキシルフルオレセイン。
【図2】R4とDNAの相互作用のアガロースゲル分析の写真表示である。様々な量(10μg、1μg、0.5μg、又は0.1μg)のR4を、1μgの(A)pEGFP−N1プラスミドか又は(B)pCI−HAプラスミドのいずれかと37℃で30分間、インキュベートした。その後、DNAをUV光下で可視化することができるようにSYBR−Safe DNAゲル染色を含む1%アガロースゲル上で試料を泳動した。
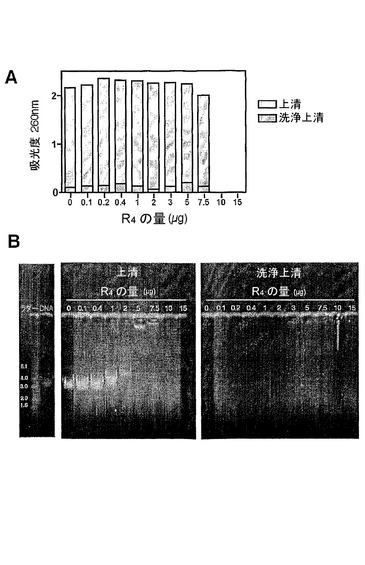
【図3】R4及びDNAを含む溶液の260nmでの吸光度測定値を示す図である。漸増量のR4を10μgのpCI−HAプラスミドと、総容量100μlの水中で37℃、30分間、インキュベートした。(A)試料を9300gで遠心分離して、いかなる不溶性複合体もペレットにし、収集された上清の吸光度測定値を260nmで測定した(薄灰色の棒)。ペレット化試料を100μlの水で再懸濁した後の洗浄上清の吸光度測定値もまた測定した(濃灰色の棒)。(B)上清及び洗浄上清もまた、SYBR−Safe DNAゲル染色を含む1%アガロースゲル上で泳動して、これらの試料中のDNAの存在を検出した。
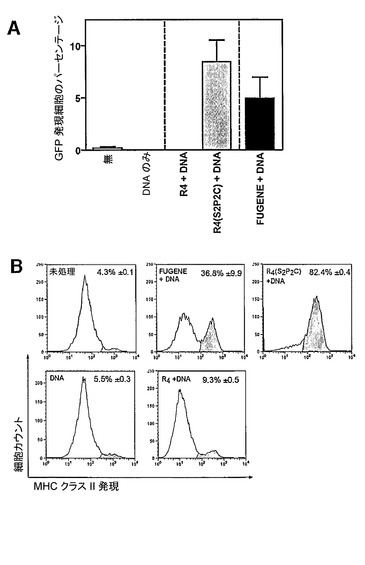
【図4】D1細胞のpEGFP−N1プラスミドでのトランスフェクション、及びD1細胞上のMHCクラスII発現の上方制御のグラフ表示である。3μgのR4若しくはR4(S2Pam2Cys)と37℃で30分間、又はFUGENEのプラスミドDNAに対する比が3:1である混合物と室温で45分間、あらかじめインキュベートした、1μgのpEGFP−N1プラスミドと共にBALB/c脾細胞由来のD1細胞(4×105個)を5%CO2の雰囲気中、37℃で培養した。2日後、細胞を収集し、フローサイトメトリーによって緑色蛍光を測定した(A)。細胞をまた、フローサイトメトリーによる分析の前に、蛍光色素結合抗マウスMHCクラスII(IA/E)抗体で染色した(B)。高レベルのMHCクラスIIを発現する細胞は、成熟であるとみなされ(ヒストグラムの陰影部)、低レベルを発現する細胞は未熟とみなされる(陰影なし)。このカテゴリーにおける細胞のパーセンテージはまた、三連の試料を分析することによって得られた平均及び標準偏差として各パネル内に示されている。
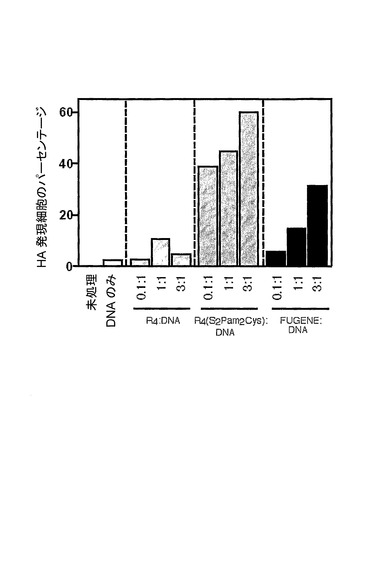
【図5】トランスフェクション試薬対DNA比の異なる混合物を用いたD1細胞のトランスフェクションのグラフ表示である。異なる比率のR4若しくはR4(S2Pam2Cys)と37℃で30分間、又は異なる比率のFUGENEと室温で45分間、あらかじめインキュベートした、1μgのpCI−HAプラスミドと共にBALB/c脾細胞由来のD1細胞(4×105個)を5%CO2中、37℃で48時間、培養した。その後、細胞を収集し、抗HA抗体(クローンE2.6)で染色し、この抗体を、フローサイトメトリーによる分析の前に、FITC結合ヤギ抗マウスIg抗体で検出した。
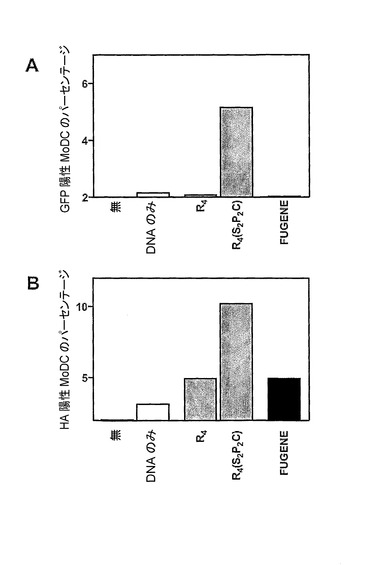
【図6】プラスミド、pEGFP−N1及びpCI−HAでのヒト単球由来樹状細胞(MoDC)のトランスフェクションのグラフ表示である。3μgのR4若しくはR4(S2Pam2Cys)と37℃で30分間、又はFUGENEのプラスミドDNAに対する比が3:1である混合物と室温で45分間、あらかじめインキュベートした、1μgのpEGFP−N1プラスミドと共に単球由来樹状細胞(2×105個)を5%CO2の雰囲気中、37℃で培養した。2日後、細胞を収集し、pEFGP−N1プラスミドをトランスフェクションされた細胞における緑色蛍光をフローサイトメトリーによって測定した(A)。pCI−HAプラスミドをトランスフェクションされた細胞について、蛍光色素結合抗HA抗体を用いてHAタンパク質の表面発現を検出した(B)。
【図7】DNAに対するトランスフェクション試薬の異なる比の混合物を用いる2つの異なる細胞系における緑色蛍光タンパク質のトランスフェクションレベルのグラフ表示である。ヒト肺上皮細胞系A549(A)又はマウス肺胞マクロファージ細胞系MH−S(B)を、異なる希釈度のR4、R4Pam2Cys、又はFUGENEと複合体を形成した、緑色蛍光タンパク質をコードするプラスミドの非存在下又は存在下において2×105個の細胞/mlの濃度で、37℃、5%CO2で48時間、培養した。トランスフェクション効率は、フローサイトメーターで細胞の蛍光を測定することによって決定した。
【図8】非免疫処置マウス、及びR4Pam2CysとDNAの複合体か又はインフルエンザウイルスかのいずれかで免疫処置されたマウスにおけるHA特異的抗体レベルのグラフ表示である。BALB/cマウス(週齢6〜8週間)に、単独か、又は1:3若しくは1:5のモル比でR4(S2Pam2Cys)と複合体を形成したかのいずれかの、インフルエンザ血球凝集素をコードするDNAを含む50μgのDNAプラスミドを0日目及び28日目に尾の付け根に皮下接種した。陽性対照として、マウスをまた、スプリットPR8ウイルス(3μg/マウス)で鼻腔内に免疫処置した。非免疫処置マウスは陰性対照としての役割を果たした。最後の接種後14日目にマウスから血清を採取し、ELISAを行って、インフルエンザPR8ウイルスに特異的な抗体の存在を検出した。
【図9】R4Pam2CysとDNAの複合体で免疫処置されたマウスの脾臓(A)及び鼠径リンパ節(B)における誘導された抗原特異的IFN−γ産生CD8+ T細胞の数のグラフ表示である。BALB/cマウス(週齢6〜8週間)に、単独か、又は1:1、1:2、若しくは1:5のNH3+:PO4−モル比でR4(S2Pam2Cys)と複合体を形成した、インフルエンザ核タンパク質をコードする20μgのDNAプラスミドを尾の付け根に皮下接種した。R4(S2Pam2Cys)を含む溶液の10μlのアリコートをDNAの溶液に2分間ごとに合計1時間、ゆっくり加えることによって、複合体形成を達成した。全ての接種材料を0.7MのNaClに溶解した。免疫処置後7日目又は10日目にマウスから脾臓及び鼠径リンパ節を摘出し、細胞内サイトカイン染色を行って核タンパク質147〜155に特異的なIFN−γ分泌CD8+ T細胞の存在を検出した。それぞれの棒は2匹のマウスからの平均及び標準誤差を表す。
【発明を実施するための形態】
【0023】
驚くべきことに、本発明者らは、本発明の化合物が、DNAと安定な複合体を形成することを見出した。DNAは、上記化合物のカチオン性部分に結合し、この複合体が、上記化合物のTLRリガンド部分を介して、上記リガンドを認識する受容体を発現する細胞、例えば、DCなどの抗原提示細胞にターゲットされる。
【0024】
本発明の化合物によって提供される一つの利点は、それらがTLR発現細胞をターゲットする能力があることである。TLRへの結合の結果として、本発明の化合物は、内部に取り入れられ、そしてまた、TLR媒介性シグナル伝達経路を誘導する。特定の実施形態において、本発明の化合物のPam2Cys部分によるTLR−2の結合は、DC成熟を引き起こし、その結果として、DCのリンパ節への遊走及びTリンパ球への効率的な抗原提示をもたらす。
【0025】
第1態様において、本発明は、少なくとも1個のTLRリガンドに連結した正荷電基を含む化合物を提供する。いくつかの実施形態において、本発明の化合物は、少なくとも1個のTLRリガンドに共有結合した正荷電基を含む。
【0026】
第2態様において、本発明は、核酸と、少なくとも1個のTLRリガンドに連結した正荷電基を含む化合物とを含む複合体であって、上記核酸が、核酸と正荷電基との間の静電相互作用によって上記化合物と会合している、複合体を提供する。
【0027】
第3態様において、本発明は、トランスフェクション方法であって、核酸と、少なくとも1個のTLRリガンドに連結した正荷電基を含む化合物とを含む複合体に、TLRを発現する細胞を接触させる段階を含み、上記核酸が、核酸と正荷電基との間の静電相互作用によって上記化合物に会合している、方法を提供する。
【0028】
第4態様において、本発明は、抗原に対する免疫応答を生じさせる方法であって、核酸と、少なくとも1個のTLRリガンドに連結した正荷電基を含む化合物とを含む複合体を対象に投与する段階を含み、上記核酸が、核酸と正荷電基との間の静電相互作用によって上記化合物に会合しており、かつ上記核酸が上記抗原又はそのエピトープをコードする、方法を提供する。
【0029】
第5態様において、本発明は、抗原に対する免疫応答を生じさせる方法であって、核酸と、少なくとも1個のTLRリガンドに連結した正荷電基を含む化合物とを含む複合体をトランスフェクションされた細胞を対象に投与する段階を含み、上記核酸が、核酸と正荷電基との間の静電相互作用によって上記化合物に会合しており、かつ上記核酸が上記抗原又はそのエピトープをコードする、方法を提供する。
【0030】
第6態様において、本発明はまた、TLRを発現する細胞で遺伝子の発現を抑圧する方法であって、核酸と、少なくとも1個のTLRリガンドに連結した正荷電基を含む化合物とを含む複合体を対象に投与する段階を含み、上記核酸が、核酸と正荷電基との間の静電相互作用によって上記化合物に会合しており、上記核酸がsiRNA、shRNA、siRNAをコードするDNA、及びshRNAをコードするDNAからなる群より選択され、かつ上記遺伝子をターゲットとする、方法を提供する。
【0031】
追加の態様において、本発明はまた、対象における免疫応答の誘導のためのワクチンの製造、遺伝子異常又は遺伝子欠損をもつ対象の処置のための医薬の製造、遺伝子の異常な発現又は別なふうに望ましくない発現を患っている対象の処置のための医薬の製造、及び細胞のトランスフェクションのための試薬の製造における、少なくとも1個のTLRリガンドに連結した正荷電基を含む化合物の使用を提供する。
【0032】
本発明は、TLRリガンド、すなわち、細胞表面TLRに結合する部分に連結した正荷電基を含む化合物を提供する。TLRリガンドのTLRへの結合の結果として、TLRを発現する細胞による上記化合物の取り込み、及び/又はTLR媒介性シグナル伝達経路を介するシグナル伝達が生じる。
【0033】
本明細書に用いられる場合、用語「TLR」は、1個又は複数のToll様受容体を意味し、微生物由来の、構造的に保存された分子に結合する1クラスの膜結合型受容体として定義することができる。これまで、13個のTLR、TLR−1〜TLR−13が同定されており、たいていの哺乳動物種は、10個〜15個の型のToll様受容体を有すると推定されている(Du,X.ら 2000 Eur.Cytokine Netw.11:362〜371;Chuang,T.H.及びUlevitch,R.J.2000 Eur.Cytokine Netw.11:372〜378;Tabeta,K.ら 2004 Proc.Natl.Acad.Sci.USA 101:3516〜3521)。TLRは、一種のパターン認識受容体(PRR)であり、それらのリガンドは、集合的に、病原体関連分子パターン(PAMP)として知られている。
【0034】
本明細書に用いられる場合、「TLRリガンド」は、TLRに選択的又は優先的に結合する分子を意味する。TLRリガンドの例は、病原体における保存された特徴を含み、細菌細胞表面リポ多糖(LPS)、リポタンパク質、リポペプチド、及びリポアラビノマンナン;細菌鞭毛由来のフラゲリンなどのタンパク質;ウイルスの二本鎖RNA、又は細菌及びウイルスのDNAの非メチル化CpGモチーフ;並びに特定の他のRNA及びDNAが挙げられる。TLRの内因性リガンドもまた同定されており、それらには、フィブリノーゲン、熱ショックタンパク質(HSP)、及びDNAが挙げられる。
【0035】
表1は、現在、樹状細胞上に発現していることが知られているTLRを、それらの対応するリガンドと共に列挙する。
【0036】
【表1】

【0037】
本発明の例示的化合物は、リポペプチド「Pam2Cys」に連結したカチオン性ペプチドである。当業者は、用語「リポペプチド」が、コンジュゲートしている、1つ又は複数の脂質部分及び1つ又は複数のアミノ酸配列を含む物質の任意の組成物を意味することを理解するであろう。「Pam2Cys」(ジパルミトイル−S−グルセリル−システイン、又はS−[2,3ビス(パルミトイルオキシ)プロピル]システインとしても知られている)が合成されており(Metzger,J.W.ら 1995 J Pept Sci 1:184)、マイコプラズマ ファーメンタンスから単離されたマクロファージ活性化リポペプチドであるMALP−2の脂質部分に対応する(Sacht,Gら 1998 Eur J Immunol 28:4207:Muhiradt,P.F.ら 1998 Infect Immun 66:4804;Muhiradt,P.F.ら 1997 J Exp Med185:1951)。Pam2CysはTLR−2のリガンドであることが知られている。
【0038】
Pam2Cysは、式(I)すなわち
【化1】
の構造を有する。
【0039】
細胞表面TLRをターゲットするために用いることができる他の脂質部分には、パルミトイル、ミリストイル、ステアロイル、ラウロイル、オクタノイル、又はデカノイルが挙げられる。好ましい基としては、Pam2Cys、Pam3Cys、Ste2Cys、Lau2Cys、及びOct2Cysが挙げられる。
【0040】
本発明の正荷電基として、表2に列挙されたカチオン性化合物、並びに米国特許第6,689,478号明細書及び米国特許第4,035,558号明細書に記載されたポリカチオン性化合物が挙げられるが、それらに限定されるわけではない。
【0041】
【表2】
【0042】
核酸は、核酸のバックボーン内のリン酸基により、正味の負電荷を帯びた分子であるため、それらは、本発明の化合物の正荷電基によって静電相互作用を介して結合され、安定な複合体を形成する。
【0043】
一実施形態において、正荷電基は、アルギニン残基又はリシン残基を含む直鎖又は分岐ペプチドである。
【0044】
「核酸」への言及は、デオキシリボ核酸(DNA)及びリボ核酸(RNA)の両方への言及として理解されるべきであり、それらには、二本鎖DNA、二本鎖RNA、一本鎖DNA、一本鎖RNA、低分子干渉RNA(siRNA)、三重鎖、四重鎖、及び任意の多数本鎖(多重鎖)核酸分子、ペプチド核酸(PNA)などの核酸ハイブリッド、デオキシリボ核酸塩基及びリボ核酸塩基を含む分子、並びに任意のヌクレオチド変異体が挙げられる。
【0045】
一実施形態において、核酸分子は、「プラスミド」として提供される。「プラスミド」への言及は、宿主細胞へ伝達可能であり、かつ宿主細胞において複製を起こすことができる核酸分子への言及として理解されるべきである。しかしながら、核酸分子は、プラスミドに限定されるべきではなく、ウイルスDNA又はウイルスRNAを含む任意の核酸分子であってもよい。
【0046】
当業者は、細胞に核酸をトランスフェクションすることが、多くの研究及び臨床の適用に有用であることを理解しているであろう。「トランスフェクション」への言及は、核酸を含む外因性分子が細胞へ導入される過程への言及として理解されるべきである。
【0047】
多くの研究適用は、細胞のトランスフェクションを必要とする。現在、研究者は、細胞への分子の導入、すなわち、細胞のトランスフェクションを、様々な効率をもつ様々な手段によって行う。いくつかのトランスフェクション試薬を、現在、研究者が利用でき、それらには、FUGENEが挙げられる。しかしながら、細胞のトランスフェクション効率は、例えば、細胞型、細胞分裂の速度及び時期、並びにトランスフェクションされることになっている分子及び(1つ又は複数の)トランスフェクション試薬の両方の個々の性質に依存する。
【0048】
本発明の化合物は、TLRを発現する細胞のトランスフェクションに特に有用である。したがって、TLRを天然で発現する細胞か、又はTLR全体であろうと細胞内シグナル伝達ドメインを欠く改変受容体であろうと、TLRを発現するように安定にトランスフェクションされている細胞のいずれかに、本発明の化合物と複合体を形成した核酸をトランスフェクションすることができる。
【0049】
「細胞」は、核酸分子が本発明の化合物によって送達される任意の細胞として理解されるべきである。細胞は、インビトロの細胞、インビボの細胞、又はエクスビボの細胞であってもよい。細胞は、単離されていてもよく、又は生きている動物の器官若しくは組織の一部を形成していてもよい。細胞はまた、細菌、酵母、真菌、カビ、寄生虫、藻類などの微生物であってもよい。細胞はまた、培養動物細胞若しくは細胞株、又は人工細胞であり得る。
【0050】
本発明の特定の実施形態の細胞に関しての、「単離された」への言及は、その本来の環境(例えば、それが自然発生している場合には、自然環境)から取り出されている物質への言及として理解されるべきである。例えば、生きている生物体の組織に存在する細胞は単離されていないが、その同じ細胞が、自然系内に共存する細胞の一部又は全部から分離されている場合、単離されている。
【0051】
本発明の一態様は、TLRリガンドに連結した正荷電基を含む化合物であって、正荷電基と負荷電核酸との間の静電相互作用によって核酸と安定な複合体を形成することができる、化合物を提供する。本発明の化合物は、TLRを発現する細胞に核酸をターゲットする手段を提供する。
【0052】
一実施形態において、本発明は、TLRリガンドに連結したカチオン性ペプチドを含む化合物であって、カチオン性ペプチドと負荷電DNAとの間の静電相互作用によってDNAと安定な複合体を形成することができる、化合物を提供する。そのような化合物は、TLRを発現する細胞、例えば、抗原提示細胞、より具体的にはDCに、DNAをターゲットする手段を提供する。
【0053】
別の実施形態において、本発明は、トランスフェクション方法であって、核酸と、TLRに結合する少なくとも1個の部分に連結した正荷電基を含む化合物とを含む複合体に、TLRを発現する細胞を接触させる段階を含み、上記核酸が、核酸と正荷電基との間の静電相互作用によって上記化合物に会合している、方法を提供する。
【0054】
さらなる実施形態において、本発明は、抗原に対する免疫応答を生じさせる方法であって、核酸と、TLRに結合する少なくとも1個の部分に連結した正荷電基を含む化合物とを含む複合体を対象に投与する段階を含み、上記核酸が、核酸と正荷電基との間の静電相互作用によって上記化合物に会合しており、かつ上記核酸が上記抗原又はそのエピトープをコードする、方法を提供する。
【0055】
さらに別の態様において、本発明は、抗原に対する免疫応答を生じさせる方法であって、核酸と、TLRに結合する少なくとも1個の部分に連結した正荷電基を含む化合物とを含む複合体をトランスフェクションされた細胞を対象に投与する段階を含み、上記核酸が、核酸と正荷電基との間の静電相互作用によって上記化合物に会合しており、かつ上記核酸が上記抗原又はそのエピトープをコードする、方法を提供する。
【0056】
本発明の化合物をDCに接触させる段階は、結果として、細胞表面MHCクラスII発現の増加によって示されるようなDCの成熟又は活性化を生じさせることができる。したがって、TLRリガンドを含む化合物は、DCをターゲットするためだけでなく、TLR及びMyD88などの下流シグナル伝達分子を介してのDCの活性化のために有用であり得る。
【0057】
それゆえに、本発明の化合物は、DCを活性化するのに有用である。樹状細胞が免疫応答を誘導するために、樹状細胞はまず、それらがリンパ節に遊走し、かつTリンパ球を活性化するために必要な接着分子及び共刺激分子を発現するように、活性化されなければならない。DNAと複合体を形成したTLRリガンドを含む化合物は、DCを活性化するのに特に有用であり、DNAがタンパク質をコードする場合、トランスフェクションされたDCは、翻訳された、DNAによりコードされるタンパク質のペプチド断片を免疫細胞へ提示し、それによって、細胞性及び体液性免疫応答を呼び起こす。換言すれば、本発明の特定の実施形態の化合物は、DNAと複合体を形成した場合、DNAワクチン接種アプローチにとって有用である。
【0058】
本明細書に用いられる場合、「DNAワクチン接種」への言及は、DNAによりコードされるタンパク質に対する免疫応答を誘導するための、対象へのDNAの投与への言及を意味する。本発明の化合物は、限定されるわけではないが、静脈内、鼻腔内、筋肉内、経口、直腸などを含む任意の経路によって対象へ直接、投与されてもよい。
【0059】
本明細書に用いられる場合、「免疫応答」への言及は、侵入した病原体、病原体に感染した細胞若しくは組織、癌性細胞、又は自己免疫若しくは病的炎症の場合には正常なヒト細胞若しくは組織の選択的損傷、破壊若しくはヒト身体からの排除を結果として生じさせるリンパ球、抗原提示細胞、食細胞、顆粒球、及び上記の細胞又は肝臓によって産生される(抗体、サイトカイン、及び補体を含む)可溶性高分子の協調作用への言及を意味する。
【0060】
本明細書に用いられる場合、「Tリンパ球応答」への言及は、Tリンパ球に依存する免疫応答の構成要素(すなわち、ヘルパーTリンパ球、細胞傷害性キラーTリンパ球、又はサプレッサーTリンパ球への、Tリンパ球の増殖及び/又は分化、抗体産生を引き起こすか又は阻止する、Bリンパ球へのヘルパーTリンパ球によるシグナルの供給、細胞傷害性Tリンパ球による特定のターゲット細胞の殺害、並びに他の免疫細胞の機能を調節する、サイトカインなどの可溶性因子の放出)への言及を意味する。
【0061】
「患者」、「対象」、又は「哺乳動物」は、交換可能に用いられ、ヒト患者及び非ヒト霊長類、加えてウサギ、ラット、及びマウスなどの実験動物、並びに他の動物などの哺乳動物を指す。動物には、全ての脊椎動物、例えば、ヒツジ、イヌ、ウシ、ニワトリ、両生類、及び爬虫類などの哺乳動物及び非哺乳動物が挙げられる。
【0062】
本発明の化合物は、DNAワクチン接種に用いることに加えて、いくつかの他の臨床適用に有用である。例えば、自己免疫疾患又は癌の処置は、造血性幹細胞(HSC)又は末梢血単核細胞(PBMC)などの自己細胞のトランスフェクションを必要とすることがある。そのような細胞を、対象から採取し、インビトロで培養し、任意で当技術分野においてよく知られた方法によってDCへ分化させ、かつ/又は核酸をトランスフェクションし、その後、対象へ注入して戻すことができる。いくつかの実施形態において、核酸はワクチン抗原をコードし、トランスフェクションされた細胞は、免疫応答を呼び起こすように意図されてもよい。他の実施形態において、核酸は、サイトカイン又は抗原性寛容原をコードしてもよい。
【0063】
別の態様において、本発明は、TLRを発現する細胞で遺伝子の発現を抑圧する方法であって、核酸と、TLRリガンドに連結した正荷電基を含む化合物とを含む複合体を対象に投与する段階を含み、上記核酸が、核酸と正荷電基との間の静電相互作用によって上記化合物に会合しており、上記核酸がsiRNA、shRNA、siRNAをコードするDNA、及びshRNAをコードするDNAからなる群より選択され、上記siRNA又はshRNAが上記遺伝子をターゲットとする、方法を提供する。
【0064】
「処置すること」又は「処置」は、疾患の症状、合併症、又は生化学的徴候の発生を予防し、又は遅らせて、疾患、状態、若しくは障害(例えば、感染性疾患、炎症、又は自己免疫疾患)の症状を軽減するか、若しくは寛解させ、又はそのさらなる発生を抑止若しくは阻害するための本発明の組成物、化合物、又は薬剤の投与を含む。「処置すること」はさらに、疾患、状態、又は障害(例えば、感染性疾患、炎症、又は自己免疫疾患)の処置又は寛解又は予防における任意の成功のきざしを指し、それらには、軽減、寛解、症状の減少若しくは疾患状態を患者にとってより許容できるものにすること、変性若しくは減退の速度を遅らせること、又は変性の最終点を衰弱がより少ないものにすることなどの任意の客観的又は主観的パラメーターが挙げられる。症状の処置又は寛解は、医師による検査の結果を含む、客観的又は主観的パラメーターに基づくことができる。したがって、用語「処置すること」は、感染性疾患、炎症、又は自己免疫疾患に関連した症状又は状態の発生を予防する若しくは遅延させるための、軽減するための、又は抑止若しくは阻害するための本発明の化合物又は薬剤の投与を含む。用語「治療効果」は、対象における疾患、疾患の症状、又は疾患の副作用の低減、排除、又は予防を指す。本発明の方法を用いる「処置すること」又は「処置」には、感染性疾患、炎症、又は自己免疫疾患のリスクの増加があり得るが、まだ症状を経験していない、又は現していない対象において症状の発生を予防すること、感染性疾患、炎症、又は自己免疫疾患の症状を阻害すること(その発生を遅らせること、又は抑止すること)、感染性疾患、炎症、又は自己免疫疾患の症状又は副作用からの解放を提供すること(対症療法を含む)、及び感染性疾患、炎症、又は自己免疫疾患の症状を軽減すること(退行を引き起こすこと)が挙げられる。処置は、疾患又は状態の顕在化後、症状の予防的(疾患の発生を予防し、若しくは遅延させるための、又はその臨床的若しくは潜在性症状の顕在化を予防するための)又は治療的抑制又は軽減であり得る。
【0065】
追加の態様において、本発明は、対象における免疫応答の誘導のためのワクチンの製造、遺伝子異常又は遺伝子欠損をもつ対象の処置のための医薬の製造、遺伝子の異常な発現又は別なふうに望ましくない発現を患っている対象の処置のための医薬の製造、及び細胞のトランスフェクションのための試薬の製造における、TLRリガンドに連結した正荷電基を含む化合物の使用を提供する。
【0066】
本明細書に示された化合物及び方法は、医学的適用及び獣医学的適用の両方に用いることができる。典型的には、生成物をヒトに用いるが、他の哺乳動物に用いてもよい。用語「患者」及び「対象」は、哺乳動物の個体を示すように意図され、本明細書を通じて、及び特許請求の範囲においてそのように用いられる。本発明の主要な適用は、ヒト患者又は対象に関係しているが、本発明は、実験動物、家畜、動物園動物、野生動物、ペット、又は競技用動物に適用してもよい。
【0067】
当業者が、所望の適用に適するように、正荷電基とTLRリガンドとの任意の組合わせを導入してもよいことは理解されているであろう。
【0068】
これより、以下の非限定的実施例を参照して、本発明をさらに記載する。
【0069】
実施例1
カチオン性ペプチドの合成
分岐ペプチド構築物については、リシン残基を用いて足場鋳型の分岐点を供給することによって合成した。足場内に存在する分岐点の数に依存して、ジカチオン性又はテトラカチオン性構造が生じた(図1)。これらの分岐構築物を、PEG−S RAM樹脂(Rapp Polymere、Tubingen、Germany;置換係数(substitution factor)0.27mmol/g)上で合成した。まず、等モル量のO−ベンゾトリアゾール−N,N,N,N’,N’−テトラメチル−ウロニウム−ヘキサフルオロホスフェート(HBTU;Novabiochem、Darmstadt、Germany)、1−ヒドロキシベンゾトリアゾール(HOBt)、及び1.5倍モル過剰のジイソプロピルエチルアミン(DIPEA;Sigma、Castle Hill、Australia)と共に、Fmoc−リシン(Mtt)−OH(Novabiochem、Laufelfingen、Switzerland)を4倍過剰で、樹脂に結合させた。アシル化を40分間行い、完全な反応をトリニトロベンジルスルホン酸(TNBSA)試験によって確認した(21)。α−アミノ基上のFmoc保護基の除去を2.5%ジアザビシクロ[5.4.0]ウンデカ−7−エン(Sigma、Steinheim、Germany)で達成し、Fmoc−リシン(Fmoc)−OH(Auspep、Melbourne、Australia)を、Fmoc基の除去後、2つの一級アミノ基を露出させて分岐点として働くように、結合させた。ジメチルホルムアミド(DMF;Auspep、Melbourne、Australia)を用いて、それぞれのアシル化段階と脱保護段階の間に樹脂を洗浄した。後続のアミノ酸を4倍過剰で結合させ、アシル化を60分間行った。四価の構築物の組み立てについて、アシル化のさらなるラウンドを、Fmoc−リシン(Fmoc)−OHを用いて60分間行い、4つの分岐点を生じさせた。アミノ酸を含む基の四価アルギニン構築物(R4)への化学選択的ライゲーションを可能にするために、システインを構築物のC末端に挿入し、R4−Cysを生じさせた。分岐ペプチド構築物上へのカルボキシフルオレセイン結合について、最後のアシル化反応後、N末端Fmoc基を除去しなかった。その代わり、C末端リシンのε−アミノ基上に存在するMtt保護基をジクロロメタン中1%TFA(Ajax Finechem、Seven Hills、Australia)での処理によって除去した。その後、等モル量のHOBt、HBTU、及び1.5倍モル過剰のDIPEAの存在下で、5(6)−カルボキシフルオレセイン(Fluka BioChemika、Switzerland)を4倍過剰用いて、暗闇中18時間、露出したε−アミノ基に結合させた。アシル化後、樹脂をDMFで洗浄し、N末端Fmoc基を除去した。全てのペプチドを同時に、固相支持体及び側鎖保護基から切断した。ウォーターズ(Waters)3 HPLCクロマトグラフィーシステムに設置されたバイダック(Vydac)C4カラム(4.6mm×250mm)又はC8カラム(4.6mm×250mm)を用いて逆相分析クロマトグラフィーによってペプチドの純度を評価した。限界溶媒としてH2O中0.1%TFA及びアセトニトリル中0.1%TFAを用いる1ml/分の流速を用いて、クロマトグラムを展開した。必要な場合、セミ分取用ウォーターズ又はGBC HPLCシステム及びセミ分取用バイダックC4カラム(10mm×300mm)を用いて2.5ml/分の流速でペプチドを精製した。ペプチド含有量の推定を、蛍光ペプチドの吸光度を496nmで測定するUV分光光度法によって測定し、83,000M−1cm−1のモル吸光係数を用いて濃度を計算した。合成された蛍光分岐ペプチド構築物の模式図は、図1に示されている。
【0070】
ペプチドの脂質付加について、DMF中50%DCMにおける等価量のHOBt及びDICIの存在下で、4倍過剰のN−フルオレニルメトキシカルボニル−S−(2,3−ジヒドロキシプロピル)−システイン(Fmoc−Dhc−OH)をC末端リシンのε−アミノ基上に結合させることによって脂質部分を構築した。アシル化を40分間行い、結合の成功がトリニトロベンゼンスルホン酸試験を用いて確認されるまで、繰り返した。樹脂結合Fmoc−Dhc−ペプチドの2つのヒドロキシ基の脂質付加を、等モル量のDMAP、10倍過剰の脂肪酸、及び12倍過剰のDICIを含む溶液を用いて一晩、行った。
【0071】
実施例2
R4によるDNA移動の遅延
分岐ペプチドR4とDNAの間の会合を調べるために、緑色蛍光タンパク質及びインフルエンザHAタンパク質をそれぞれコードするDNAプラスミドpEGFP−N1又はpCI−HAの一定量を、様々な量のR4とインキュベートし、アガロースゲル電気泳動によって分析した(図2)。
【0072】
各試料中のR4の量を増加させることによって、DNA移動に影響を及ぼすことができた。臭化エチジウム蛍光によって可視化されたように、陽極へのDNAプラスミド移動の部分的遅延は、0.1〜0.5μgのR4を用いた場合、明らかであり、より多い量のR4とインキュベートされたDNA試料はより遅い速度で動いた。開始点におけるDNAのバンドの出現によって明らかなように、10μgのR4が存在した場合、DNA移動の完全な遅延が達成された。しかしながら、このバンドは、全くかすかにしか見えず、この試料中にほとんどDNAがないことを示唆した。これは、R4が臭化エチジウムのDNAへの結合を阻害すること、及び/又はR4とDNAの間の相互作用の結果として、アガロースゲル中で移動するよりむしろ電気泳動緩衝液中へ拡散する不溶性複合体の形成をもたらすことが原因である可能性がある。これらの結果は、R4の正荷電アミノ基及びグアニジン基がDNAバックボーン内の負荷電リン酸基を中和して、陽極へ移動するその能力を遅延させ得ることを示唆する。
【0073】
実施例3
R4−DNA複合体の形成
R4のDNAへの結合を確認するために、漸増量のR4及びDNAを含む遠心分離した混合物の上清におけるDNAの量を、260nmにおいて吸光度を測定することによって決定した(図3A)。DNAをその電荷の中和によって溶液から沈殿させるR4の能力は、溶液中のDNAの減少を生じさせ、それにしたがって、それら2つの間の会合を示すことになるだろう。遠心分離された上清におけるDNAの量は、0.1μg〜5μgのR4とインキュベートした場合、劇的には変化しなかった。しかしながら、7.5μgのR4を用いた場合、わずかな減少が生じ、これより多い量を含む溶液中において、DNAは検出されなかった。この結果は、存在するR4の量がDNAの溶液からの沈殿と比例することを示す。吸光度の測定値をまた、いかなる遠心分離された不溶性物質も再懸濁するために用いられた洗浄上清についても取った。これらの試料中にDNAがほとんど存在しないことは、R4−DNA沈殿物が安定であること、及びDNAがこの複合体から解離する可能性は低いことを示唆する。
【0074】
アガロースゲル分析(図3B)は、DNAバンドが、0.1〜7.5μgのR4を含む上清試料においては明らかであったが、10μg又は15μgのR4を含む試料においては明らかではなかったことを示す。DNA移動の遅延はまた、1μg又はそれ以上のR4を含む試料において明らかである。
【0075】
実施例4
R4(S2Pam2Cys)−DNAでのDCのトランスフェクション
R4構築物又はR4(S2Pam2Cys)構築物がトランスフェクションされたDNAの発現を誘導する能力があるかどうかを決定するために、マウスDC系、D1にpEGFPプラスミドをR4又はR4(S2Pam2Cys)を用いてトランスフェクションした(図4A)。これらの細胞における緑色蛍光タンパク質(GFP)の発現レベルを、市販されているトランスフェクション試薬FUGENEを用いてトランスフェクションしたD1細胞における緑色蛍光タンパク質の発現レベルと比較した。
【0076】
未処理の細胞、DNAのみで処理された細胞、及びDNAと共にR4で処理された細胞において、蛍光は観察されなかった。しかしながら、R4(S2Pam2Cys)及びDNAをトランスフェクションされた細胞は、pEGFP及びFUGENE試薬をトランスフェクションされた細胞において観察された発現レベルに匹敵するレベルでGFPを発現した。
【0077】
D1細胞培養物はまた、それぞれ未熟細胞及び成熟細胞に対応する、MHCクラスII低か又はMHCクラスII高のいずれかである2つの別々の細胞集団で構成されることが見出された(図4B)。未処理のD1細胞の約4%はMHCクラスII高であった。DNA単独での処理又はR4とのDNAでの処理は、D1細胞に関してMHCクラスIIの発現を感知できるほどには変化させなかった。DNA及びFUGENEで処理されたD1細胞は、約36%のMHCクラスII高細胞を含み、R4(S2Pam2Cys)及びDNAで処理されたD1細胞は、約82%のMHCクラスII高細胞を含んだ。
【0078】
DNAに対するトランスフェクション試薬の比が異なるそれらの混合物を用いるトランスフェクション効率もまた、一定量(1μg)のpCI−HAプラスミドとあらかじめインキュベートした漸増量(0.1μg、1μg、又は3μg)のR4、R4(S2Pam2Cys)、又はFUGENEとD1細胞をインキュベートすることによって調べた(図5)。細胞表面インフルエンザHAタンパク質発現を、HA特異的抗体を用いて測定した。R4(S2Pam2Cys)又はFUGENEを用いた場合、表面HAタンパク質を発現する細胞のパーセンテージによって示されるように、トランスフェクション効率は、用いられるトランスフェクション剤の量と比例した。これの例外は、R4を用いた場合であった。この場合、HA発現レベルは、R4(S2Pam2Cys)又はFUGENEを含むDNA混合物によって誘導されるHA発現レベルより低かったが、DNAに対するR4の重量対重量比が同等である時、最適であるように思われた。しかしながら、より高い比率は、トランスフェクション効率にとって有害であることが見出された。
【0079】
調べられた比率の全部において、R4(S2Pam2Cys)は、抗原の発現を誘導することにおいてFUGENEより優れていることが見出された。R4(S2Pam2Cys)及びDNAを用いてトランスフェクションされた細胞の最大パーセンテージは、3:1比において60%であった。相対的に、DNAに対するEUGENEの等価比率を用いた時、約30%のHA陽性細胞だけが、検出された。
【0080】
実施例5
ヒト単球由来樹状細胞(MoDC)のトランスフェクション
ヒト単球由来樹状細胞(MoDC)のトランスフェクションを増強するR4(S2Pam2Cys)の能力を調べた。MoDCにpEGFPプラスミドをトランスフェクションした実験において(図6A)、R4(S2Pam2Cys)のみが、GFPの発現を誘導することにおいて効率的であった。FUGENEか又はR4のいずれかを加えたpEGFPでの細胞のトランスフェクションは、相対的に非効率であり、これらの細胞におけるトランスフェクションレベルは、プラスミド単独で処理されたものと類似していた。この結果は、MoDCに、各トランスフェクション試薬と共にpCI−HAプラスミドをトランスフェクションした実験において、繰り返された(図6B)。
【0081】
実施例6
R4(S2Pam2Cys)−DNAでの非DC細胞系のトランスフェクション
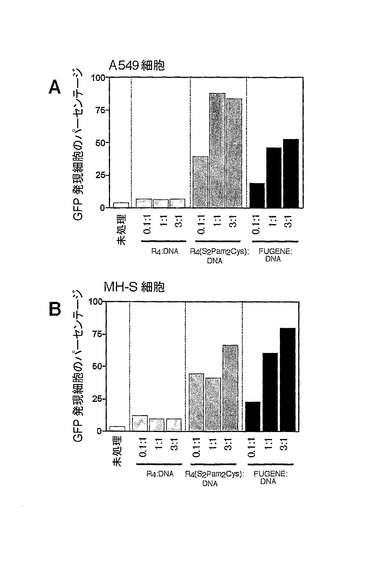
DNAと複合体を形成した、R4構築物又はR4(S2Pam2Cys)構築物が非DC細胞系をトランスフェクションする能力があるかどうかを調べるために、ヒト肺上皮細胞系A549及びマウス肺胞マクロファージ細胞系MH−Sに、R4又はR4(S2Pam2Cys)を用いてpEGFPプラスミドをトランスフェクションした(図7A及び7B)。ヒト肺上皮細胞系A549(A)又はマウス肺胞マクロファージ細胞系MH−S(B)を、異なる希釈度のR4、R4Pam2Cys、又はFUGENEと複合体を形成した、緑色蛍光タンパク質をコードするプラスミドの非存在下又は存在下において2×105個の細胞/mlの濃度で、37℃、5%CO2で48時間、培養した。トランスフェクション効率については、フローサイトメーターで細胞の蛍光を測定することによって決定した。これらの細胞系の両方とも他の研究においてTLR−2を発現することが報告されている(Oshikawa 2003、Slevogt 2007)。未処理のA549又はMH−S細胞、及びDNAと共にR4で処理された細胞において、皆無かそれに近い蛍光が観察された。しかしながら、R4(S2Pam2Cys)及びDNAをトランスフェクションされたA549又はMH−S細胞は、pEGFPプラスミド及びFUGENE試薬をトランスフェクションされた細胞において観察された発現レベルと、より高くはないにしても、匹敵するレベルでGFPを発現した。
【0082】
実施例7
R4(S2Pam2Cys)−DNAで免疫処置されたマウスにおける特異的抗体産生
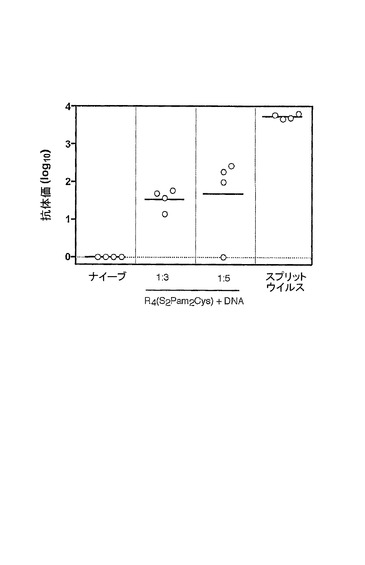
DNAと複合体を形成したR4(S2Pam2Cys)の抗体を誘導する能力を評価した。BALB/cマウス(週齢6〜8週間)に、単独か、又は1:3若しくは1:5のモル比でR4(S2Pam2Cys)と複合体を形成した、インフルエンザ血球凝集素をコードするDNAを含む50μgのDNAプラスミドを0日目及び28日目に尾の付け根に皮下接種した。陽性対照として、マウスをまた、スプリットPR8ウイルス(3μg/マウス)で鼻腔内に免疫処置した。非免疫処置マウスは陰性対照としての役割を果たした。最後の接種後14日目にマウスから血清を採取し、ELISAを行って、インフルエンザPR8ウイルスに特異的な抗体の存在を検出した。血清抗HA抗体をELISAによって測定した。非免疫処置マウス又は「ナイーブ」マウスにおいてHA特異的抗体は検出されなかったが、1:3及び1:5のモル比で混合された2つの用量のR4(S2Pam2Cys)及びDNAで免疫処置されたマウスにおいて、インフルエンザウイルスに結合することができる実質的レベルの抗体が検出された(図8)。
【0083】
実施例8
R4(S2Pam2Cys)−DNAで免疫処置されたマウスにおけるインフルエンザ核タンパク質特異的IFN−γ陽性CD8+T細胞の誘導
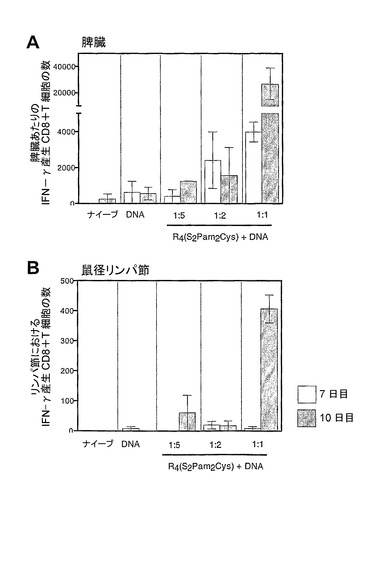
DNAと複合体を形成したR4(S2Pam2Cys)の細胞性免疫応答を誘導する能力を評価した。BALB/cマウス(週齢6〜8週間)に、単独か、又は1:1、1:2、若しくは1:5のNH3+:PO4−モル比でR4(S2Pam2Cys)と複合体を形成した、インフルエンザ核タンパク質をコードする20μgのDNAプラスミドを尾の付け根に皮下接種した。R4(S2Pam2Cys)を含む溶液の10μlのアリコートをDNAの溶液に2分間ごとに合計1時間、ゆっくり加えることによって、複合体形成を達成した。全ての接種材料を0.7MのNaClに溶解した。免疫処置後7日目又は10日目にマウスから脾臓及び鼠径リンパ節を摘出し、細胞内サイトカイン染色を行って核タンパク質147〜155に特異的なIFN−γ分泌CD8+T細胞の存在を検出した。それぞれの棒は2匹のマウスからの平均及び標準誤差を表す。
【0084】
DNA単独で接種されたマウスの脾臓及び鼠径リンパ節では非常に低レベルの活性化CD8+T細胞しか検出されず(それぞれ、図9A及び9B)、ナイーブマウスでは検出されなかった。しかしながら、異なる比率のDNAに対するR4(S2Pam2Cys)を含む複合体の投与は、免疫処置後7日目、脾臓において検出可能なNP特異的IFN−γ産生CD8+T細胞を誘導した(図9A)。特に、1:1比のR4(S2Pam2Cys)とDNAを接種されたマウスのリンパ節及び脾臓の両方において、高レベルの活性化CD8+T細胞が検出された(図9A及び9B)。
【0085】
参考文献
Alexopoulou L., Holt A. C., Medzhitov R. and Flavell R. A. (2001) Recognition of double−stranded RNA and activation of NF−kappaB by Toll−like receptor 3. Nature 413 (6857): 732−738
Asea A., Rehli M., Kabingu E., Boch J. A., Bare O., Auron P. E., Stevenson M. A. and Calderwood S. K. (2002) Novel signal transduction pathway utilized by extracellular HSP70: role of toll−like receptor (TLR) 2 and TLR4. J Biol Chem 277 (17): 15028−15034
Billingham J., Breen C., Rawson J. O., Yarwood J., Mann B.E. (1997) Adsorption of polycations on clays: A comparative in situ study using 133Cs and 23Na solution phase NMR. Journal of Colloid and Interface Science 193:183−189
Bulut Y., Faure E., Thomas L., Karahashi H., Michelsen K. S., Equils O., Morrison S. G., Morrison R. P. and Arditi M. (2002) Chlamydiai heat shock protein 60 activates macrophages and endothelial cells through Toll−like receptor 4 and MD2 in a MyD88−dependent pathway. J.Immunol 168 (3); 1435−1440
Buschle M, Schmidt W, Zauner W, Mechtler K, Trska B, Kirlappos H, et al. (1997) Transloading of tumor antigen−derived peptides into antigen−presenting cells. Proc Natl Acad Sci USA 94(7):3256−3261
Chow J. C., Young D. W., Golenbock D. T., Christ W. J. and Gusovsky F. (1999) Toll−like receptor−4 mediates lipopolysaccharide−induced signal transduction. J−Biol Chem 274 (16): 10689−10692
Christiaens B, Grooten J, Reusens M, Joliot A, Goethals M, Vandekerckhove J, Procbiantz A, Rosseneu M. (2004) Membrane interaction and cellular internalization of penetratin peptides. Eur J Biochem. 271(6);1187−1197
Fawell S, Seery J, Daikh Y, Moore C, Chen LL, Pepinsky B, et al. (1994) Tat−mediated delivery of heterologous proteins into cells. Proc Natl Arad Sci USA 91(2);664− 668
Flacher V, Bouschbacher M, Verronese E, Massacrier C, Sisirak V, Berthier−Vergnes O, de Saint−Vis B, Caux C, Dezutter−Dambuyant C, Lebecque S, Valladeau J. (2006) Hiuman Langerhans cells express a specific TLR profile and differentially respond to viruses and Gram−positive bacteria. J Immunol. 177(11):7959−7967
Futaki S, Suzuki T, Ohashi W, Yagami T, Tanaka S, Ueda K, et al. (2001) Argininerich peptides. An abundant source of membrane−permeable peptides having potential as carriers for intracellular protein delivery, J Biol Chem 276(8):5836−5840
Hayashi F., Smith K. D., Ozinsky A., Hawn T. R., Yi E. C., Goodlett D. R., Eng J. K., Akira S., Underhill D. M. and Aderem A. (2001) The innate immune response to bacterial flagellin is mediated by Toll−like receptor 5. Nature 410 (6832): 1099−1103
Heil F., Hemmi H., Hochrein H., Ampenberger F., Kirschning C., Akira S., Lipford G., Wagner H and Bauer S. (2004) Species−specific recognition of single−stranded RNA via toll−like receptor 7 and 8. Science 303 (5663): 1526−1529
Hemmi H., Kaisho T., Takeuchi O., Sato S., Sanjo H., Hoshino K., Horiuchi T., Tomizawa H., Takeda K. and Akira S. (2002) Small anti−viral compounds activate immune cells via the TLR7 MyD88−dependent signaling pathway. Nat Immunol 3 (2): 196−200
Hemmi H., Takeuchi O., Kawai T., Kaisho T., Sato S., Sanjo H., Matsumoto M., Hoshino K., Wagner H., Takeda K. and Akira S. (2000) A Toll−like receptor recognizes bacterial DNA. Nature 408 (6813): 740−745
Husebye H, Halaas (, Stenmark H, Tunheim G, Sandanger (, Bogen B, Brech A, Latz E, Espevik T. (2006) Endocytic pathways regulate Toll−like receptor 4 signaling and link innate and adaptive immunity. EMBO J. 25(4):683−692
Mitchell DJ, Kim DT, Steinman L, Fathman CG, Rothbard JB. (2000) Polyarginine enters cells more efficiently than other polycationic homopolymers. J Pept Res 56(5):318−325
Morr M., Takeuchi O., Akira S., Simon M. M. and Muhlradt P. F. (2002) Differential recognition of structural details of bacterial lipopeptides by toll−like receptors. Eur J Immunol 32 (12): 3337−3347
Oehlke J, Scheller A, Wiesner B, Krause E, Beyermann M, Klauschenz E, et al. (1998) Cellular uptake of an alpha−helical amphipathic model peptide with the potential to deliver polar compounds into the cell interior non−endocytically. Biochim Biophys Acta 1414(1−2):127−139
Okusawa T., Fujita M., Nakamura J., Into T., Yasuda M., Yoshimura A., Hara Y., Hasebe A., Golenbock D. T., Morita M., Kuroki Y., Ogawa T. and Shibata K. (2004) Relationship between structures and biological activities of mycoplasmal diacylated lip opeptides and their recognition by toll−like receptors 2 and 6. Infect Immun 72(3): 1657−1665
Oshikawa K, Sugiyama Y. (2003) Regulation of toll−like receptor 2 and 4 gene expression in murine alveolar macrophages. Exp Lung Res 29(6):401−412
Otvos L, Jr., Cudic M, Chua BY, Deliyannis G, Jackson DC. (2004) An insect antibacterial peptide−based drug delivery system. Mol Pharm 1(3):220−232
Ozinsky A., Underhill D. M., Fontenot J. D., Hajjar A. M., Smith K. D., Wilson C. B., Schroeder L. and Aderem A. (2000) The repertoire for pattern recognition of pathogens
by the innate immune system is defined by cooperation between toll−like receptors. Proc Natl Acad Sci USA 97 (25): 13766−13771
Pepper M, Dzierszinski F, Wilson E, Tait E, FangQ, Yarovinsky F, Laufer TM, Roos D, Hunter CA. (2008) Plasmacytoid dendritic cells are activated by Toxoplasma gondii to present antigen and produce cytokines. J Immunol. 2008 180(9):6229−6236
Poltorak A., He X, Smirnova L, Liu M. Y., Van Huffel C., Du X., Birdwell D., Alejos E., Silva M., Galanos C., Freudenberg M., Ricciardi−Castagnoli P., Layton B. and Beutler B. (1998) Defective LPS signaling in C3H/HeJ and C57BL/10ScCr mice: mutations in Tlr4 gene. Science 282 (5396): 2085−2088
Pooga M, Hallbrink M, Zorko M, Langel U. (1998) Cell penetration by transportan. Faseb J 12(1):67−77
Schjetne K. W., Thompson K. M., Nilsen N., Flo T. H., Fleckenstein B., Iversen J. G., Espevik T., and Bogen B. (2003) Cutting edge: link between innate and adaptive immunity: Toll−like receptor 2 internalizes antigen for presentation to CD4+ T cells and could be an efficient vaccine target J Immunol 171:32−36
Schwandner R., Dziarski R., Wesche H., Rothe M. and Kirschning C. J. (1999) Peptidoglycan− and lipoteichoic acid−induced cell activation is mediated by toll−like receptor 2. J Biol Chem 274 (25): 17406−17409
Slevogt H, Seybold J, Tiwari KN, Hocke AC, Jonatat C, Dietel S, Hippenstiel S, Singer BB, Bachmann S, Suttorp N, Opitz B. (2007) Moraxella catarrhalis is internalized in respiratory epithelial cells by a trigger like mechanism and initiates a TLR2− and partly NOD1−dependent inflammatory immune response. Cell Microbiol 9(3):694−707
Takeshita F, Leifer CA, Gursel I, Ishii KJ, Takeshita S, Gursel M, Klinman DM. (2001) Cutting edge: Role of Toll−like receptor 9 in CpG DNA−induced activation of human cells. J Immunol. 167(7):3555−3558
Takeuchi O., Kaufmann A., Grote K, Kawai T., Hoshino K., Morr M., Muhlradt P. F. and Akira S. (2000) Cutting edge: preferentially the R−stereoisomer of the mycoplasmal lipopeptide macrophage−activating lipopeptide−2 activates immune cells through a toll−like receptor 2− and MyD88−dependent signaling pathway. J Immunol 164 (2): 554−557
Takeuchi O., Sato S., Horiuchi T., Hoshino K., Takeda K., Dong Z., Modlin R. L. and Akira S. (2002) Cutting edge: role of Toll−like receptor 1 in mediating immune response to microbial lipoproteins. J Immunol 169 (1): 10−14 .
Wagner E., Zenke M., Cotten M., Beug, H., Birnstiel M. L. (1990) Transferrinpolycation conjugates as carriers for DNA uptake into cells. Proc. Natl. Acad. Sci. USA 87:3410−3414
【発明の詳細な説明】
【0001】
[発明の分野]
本発明は、一般的に、細胞生物学分野に関する。より具体的には、本発明は、細胞、特に、樹状細胞をトランスフェクションする方法、及び免疫応答を誘発する方法に向けられる。この目的を達成するために、本発明は、Toll様受容体(TLR)ファミリーの1つ又は複数のメンバーと相互作用する能力がある部分に連結した、核酸を結合する能力がある少なくとも1個の正荷電基を含む化合物を提供する。
【0002】
[発明の背景]
本明細書に用いられる場合、用語「由来する」は、特定された完全体(integer)が、特定の供給源から得ることができるが、ただし、必ずしもその供給源から直接というわけではないことを示すものと解釈されるべきである。
【0003】
本明細書を通して、文脈が別な意味を必要としない限り、単語「含む(comprise)」、又は「含む(comprises)」若しくは「含むこと(comprising)」などの語尾変化は、述べられた段階若しくは要素若しくは完全体、又は段階若しくは要素若しくは完全体の群を包含するが、ただし、いかなる他の段階若しくは要素若しくは完全体、又は要素若しくは完全体の群も排除するものではないことを意味するように理解されるだろう。
【0004】
本明細書に記載された本発明が、具体的に記載されたもの以外に変形及び改変を受ける余地があることを当業者は理解しているだろう。本発明が全てのそのような変形及び改変を含むことは理解されるべきである。本発明はまた、本明細書に、個々に又は集合的に、参照された又は示された段階、特徴、組成物、及び化合物の全部、並びに前記段階又は特徴のありとあらゆる組合せ又は任意の2つ以上を含む。
【0005】
本発明は、本明細書に記載された特定の例によって範囲が限定されるべきではない。機能的等価の生成物、組成物、及び方法は明らかに、本明細書に記載された本発明の範囲内である。
【0006】
本出願に引用された全ての参考文献は、本明細書に参照により明確に組み入れられている。
【0007】
本発明は、他に規定がない限り、分子生物学、微生物学、ウイルス学、組換えDNAテクノロジー、溶液中でのペプチド合成、固相ペプチド合成、及び免疫学の通常の技術を用いて、過度の実験なしに実施される。そのような手順は、例えば、参照により組み入れられている以下のテキスト、すなわち、
1.Sambrook、Fritsch、及びManiatis、Molecular Cloning:A Laboratory Manual、Cold Spring Harbor Laboratories、New York、第2版(1989)、I巻、II巻、及びIII巻の全体、
2.DNA Cloning:A Practical Approach、I巻及びII巻(D.N.Glover編、1985)、IRL Press、Oxford、テキストの全体、
3.Oligonucletoide Synthesis:A Practical Approach(M.J.Gait編、1984)IRL Press、Oxford、テキストの全体、並びに特に、Gait、1〜22ページ;Atkinsonら、35〜81ページ;Sproatら、83〜115ページ;及びWuら、135〜151ページによるその中の論文
に記載されている。
【0008】
本明細書中のいかなる先行技術への言及も、この先行技術がオーストラリアにおける共通一般知識の部分を形成するという認識でも、いかなる形の示唆でもなく、そのように解釈されるべきではない。
【0009】
脂質、タンパク質、ペプチド、DNA、多糖、及び/又はそれらの組合せ(例えば、リポ多糖、リポタンパク質)などの分子の、細胞内への送達は、多数の研究及び臨床の目的にとって有用である。例えば、研究者が遺伝子制御及び遺伝子発現、DNA−タンパク質相互作用又はタンパク質−タンパク質相互作用などの細胞内過程を研究するために、分子を細胞へ導入することが不可欠であることが多く、できる限り高効率でそのように行うことが望ましい。現在、研究者は、細胞への分子の送達、すなわち、細胞のトランスフェクションを、様々な効率をもつ様々な手段によって行う。細胞のトランスフェクション効率は、細胞型、細胞分裂の速度及び時期、並びにトランスフェクションされることになっている分子及び(1つ又は複数の)トランスフェクション試薬の両方の個々の性質を含むいくつかの因子に依存している。
【0010】
DNAワクチンは、樹状細胞(DC)などの抗原提示細胞によるDNAの取り込みを介して免疫応答を誘発すると考えられており、樹状細胞が、その後、内部移行したDNAにコードされる抗原を発現し、MHC分子との関連でペプチドとして免疫系に抗原を提示する。小動物モデルにおいて、DNAの投与は、防御免疫応答を誘導することに成功しているが、ただし、ヒト臨床試験において効力の低さが報告されており、免疫応答を誘導するのに高用量のDNAを必要とすることが多い(Kutzler,M.A.及びWeiner,D.B.2004 J Clin Invest、114(9)、1241〜1244)。アデノウイルス及びレトロウイルスは、遺伝子送達のためのベクターとして用いられているが、ヒトに用いるためのこれらのベクターの安全性に関して懸念が存在する(Buckley,R.H.2002 Lancet、360(9341)、1185〜1186)。今までのところ、比較的安全な非ウイルスベクターを用いる、DCのトランスフェクションは困難であることが証明されている。
【0011】
DNAは、正味の負電荷を帯びた分子である。より具体的には、DNAのバックボーン内のリン酸基が負電荷をもつ。したがって、正味の正電荷を有するカチオン性分子は、静電相互作用を介してDNAを吸着することができ、DNAの有望な担体である。そのようなカチオン性分子として、微粒子(Minigo,Gら 2007 Vaccine、25(7)、1316〜1327;Mollenkopf,H.J.ら 2004 Vaccine、22(21〜22)、2690〜2695)、ペプチド(Gratton,J.P.ら 2003 Nat Med、9(3)、357〜362;Riedl,P.ら 2006 Methods Mol Med、127、159〜169)、又はリポソーム(Jiao,Xら 2003 Hepatology、37(2)、452〜460;Ewert,K.ら 2002 J Med Chem、45(23)、5023〜5029)が挙げられる。
【0012】
しかしながら、単にDNAを抗原提示細胞に運ぶだけでは、その結果トランスフェクションが生じるには十分ではなく、抗原特異的応答を作動させるために、DNAの取り込みもまた存在しなければならない。免疫系の抗原提示細胞は、それらの細胞表面上にToll様受容体(TLR)を発現しており、そのTLRは、主として微生物に由来する様々なリガンドに結合する。例えば、TLR−2は細菌リポタンパク質を結合することが知られており、TLR−4は細菌リポ多糖を結合することが知られており、TLR−6は、TLR−1と共同して、脱アシル化細菌脂質を結合することが知られており、TLR−9は、CpG DNAに結合する。樹状細胞サブセットは、9個以上のそのようなTLRを発現することが示されている。DCの表面上の1個又は複数のTLRの会合が、細胞シグナル伝達経路を誘導し、それが、防御免疫の誘導に必要とされる、DCの成熟及び活性化をもたらすことができる。
【0013】
脂質部分、ジパルミトイル−S−グリセリルシステイン(Pam2Cys)は、マイコプラズマ ファーメンタンス(Mycoplasma fermentans)の細胞膜由来のMALP−2として知られた細菌リポタンパク質の合成類似体である。Pam2Cysは、TLR−2及びTLR−6の両方のリガンドである(Okusawa,Tら、Infect Immun 2004、72(3)、1657〜1665)。ペプチドエピトープに結合したPam2Cysを含むワクチンは、強い体液性応答及び細胞性応答を誘導することができる。ペプチドエピトープに結合したPam2CysによるTLR−2の会合の結果として、DC成熟、NF−κBなどの転写因子の活性化と、炎症誘発性サイトカインの分泌と、最終的にはエピトープ特異的ナイーブT細胞を活性化する、流入領域リンパ節へのDCの遊走とが生じる(Jackson,D.C.ら 2004 Proc Natl Acad Sci USA、101(43)、15440〜15445;Zeng,Wら 2002 J Immuol、169(9)、4905〜4912;Chua,B.Y.ら 2007 Vaccine、25(1)、92〜101)。
【0014】
[発明の概要]
本発明に繋がる研究において、本発明者らは、細胞に核酸をトランスフェクションする効率的な手段を生み出そうとした。それはまた、対象において、抗原提示細胞、特に樹状細胞をターゲットにし、体液性免疫応答及び細胞性免疫応答の両方を誘発する、DNAワクチン候補を作製しようとすることでもあった。
【0015】
第1態様において、本発明は、少なくとも1個のTLRリガンドに連結した正荷電基を含む化合物を提供する。
【0016】
第2態様において、本発明は、核酸と、少なくとも1個のTLRリガンドに連結した正荷電基を含む化合物とを含む複合体であって、上記核酸が、核酸と正荷電基との間の静電相互作用によって上記化合物と会合している、複合体を提供する。
【0017】
第3態様において、本発明は、トランスフェクション方法であって、核酸と、少なくとも1個のTLRリガンドに連結した正荷電基を含む化合物とを含む複合体に、TLRを発現する細胞を接触させる段階を含み、上記核酸が、核酸と正荷電基との間の静電相互作用によって上記化合物に会合している、方法を提供する。
【0018】
第4態様において、本発明は、抗原に対する免疫応答を生じさせる方法であって、核酸と、少なくとも1個のTLRリガンドに連結した正荷電基を含む化合物とを含む複合体を対象に投与する段階を含み、上記核酸が、核酸と正荷電基との間の静電相互作用によって上記化合物に会合しており、かつ上記核酸が上記抗原又はそのエピトープをコードする、方法を提供する。
【0019】
第5態様において、本発明は、抗原に対する免疫応答を生じさせる方法であって、核酸と、少なくとも1個のTLRリガンドに連結した正荷電基を含む化合物とを含む複合体をトランスフェクションされた細胞を対象に投与する段階を含み、上記核酸が、核酸と正荷電基との間の静電相互作用によって上記化合物に会合しており、かつ上記核酸が上記抗原又はそのエピトープをコードする、方法を提供する。
【0020】
第6態様において、本発明はまた、TLRを発現する細胞において遺伝子の発現を抑圧する方法であって、核酸と、少なくとも1個のTLRリガンドに連結した正荷電基を含む化合物とを含む複合体を対象に投与する段階を含み、上記核酸が、核酸と正荷電基との間の静電相互作用によって上記化合物に会合しており、上記核酸がsiRNA、shRNA、siRNAをコードするDNA、及びshRNAをコードするDNAからなる群より選択され、かつ上記遺伝子をターゲットとする、方法を提供する。
【0021】
さらには、対象における免疫応答の誘導のためのワクチンの製造、遺伝子異常又は遺伝子欠損をもつ対象の処置のための医薬の製造、遺伝子の異常な発現又は望ましくない発現を患っている対象の処置のための医薬の製造、及び細胞のトランスフェクションのための試薬の製造における、少なくとも1個のTLRリガンドに連結した正荷電基を含む化合物の使用を提供する。
【図面の簡単な説明】
【0022】
【図1】様々な合成された蛍光ペプチド構築物の図表示である。略語;R、アルギニン;K、リシン;L、直鎖;FL、フルオレセイン又はカルボキシルフルオレセイン。
【図2】R4とDNAの相互作用のアガロースゲル分析の写真表示である。様々な量(10μg、1μg、0.5μg、又は0.1μg)のR4を、1μgの(A)pEGFP−N1プラスミドか又は(B)pCI−HAプラスミドのいずれかと37℃で30分間、インキュベートした。その後、DNAをUV光下で可視化することができるようにSYBR−Safe DNAゲル染色を含む1%アガロースゲル上で試料を泳動した。
【図3】R4及びDNAを含む溶液の260nmでの吸光度測定値を示す図である。漸増量のR4を10μgのpCI−HAプラスミドと、総容量100μlの水中で37℃、30分間、インキュベートした。(A)試料を9300gで遠心分離して、いかなる不溶性複合体もペレットにし、収集された上清の吸光度測定値を260nmで測定した(薄灰色の棒)。ペレット化試料を100μlの水で再懸濁した後の洗浄上清の吸光度測定値もまた測定した(濃灰色の棒)。(B)上清及び洗浄上清もまた、SYBR−Safe DNAゲル染色を含む1%アガロースゲル上で泳動して、これらの試料中のDNAの存在を検出した。
【図4】D1細胞のpEGFP−N1プラスミドでのトランスフェクション、及びD1細胞上のMHCクラスII発現の上方制御のグラフ表示である。3μgのR4若しくはR4(S2Pam2Cys)と37℃で30分間、又はFUGENEのプラスミドDNAに対する比が3:1である混合物と室温で45分間、あらかじめインキュベートした、1μgのpEGFP−N1プラスミドと共にBALB/c脾細胞由来のD1細胞(4×105個)を5%CO2の雰囲気中、37℃で培養した。2日後、細胞を収集し、フローサイトメトリーによって緑色蛍光を測定した(A)。細胞をまた、フローサイトメトリーによる分析の前に、蛍光色素結合抗マウスMHCクラスII(IA/E)抗体で染色した(B)。高レベルのMHCクラスIIを発現する細胞は、成熟であるとみなされ(ヒストグラムの陰影部)、低レベルを発現する細胞は未熟とみなされる(陰影なし)。このカテゴリーにおける細胞のパーセンテージはまた、三連の試料を分析することによって得られた平均及び標準偏差として各パネル内に示されている。
【図5】トランスフェクション試薬対DNA比の異なる混合物を用いたD1細胞のトランスフェクションのグラフ表示である。異なる比率のR4若しくはR4(S2Pam2Cys)と37℃で30分間、又は異なる比率のFUGENEと室温で45分間、あらかじめインキュベートした、1μgのpCI−HAプラスミドと共にBALB/c脾細胞由来のD1細胞(4×105個)を5%CO2中、37℃で48時間、培養した。その後、細胞を収集し、抗HA抗体(クローンE2.6)で染色し、この抗体を、フローサイトメトリーによる分析の前に、FITC結合ヤギ抗マウスIg抗体で検出した。
【図6】プラスミド、pEGFP−N1及びpCI−HAでのヒト単球由来樹状細胞(MoDC)のトランスフェクションのグラフ表示である。3μgのR4若しくはR4(S2Pam2Cys)と37℃で30分間、又はFUGENEのプラスミドDNAに対する比が3:1である混合物と室温で45分間、あらかじめインキュベートした、1μgのpEGFP−N1プラスミドと共に単球由来樹状細胞(2×105個)を5%CO2の雰囲気中、37℃で培養した。2日後、細胞を収集し、pEFGP−N1プラスミドをトランスフェクションされた細胞における緑色蛍光をフローサイトメトリーによって測定した(A)。pCI−HAプラスミドをトランスフェクションされた細胞について、蛍光色素結合抗HA抗体を用いてHAタンパク質の表面発現を検出した(B)。
【図7】DNAに対するトランスフェクション試薬の異なる比の混合物を用いる2つの異なる細胞系における緑色蛍光タンパク質のトランスフェクションレベルのグラフ表示である。ヒト肺上皮細胞系A549(A)又はマウス肺胞マクロファージ細胞系MH−S(B)を、異なる希釈度のR4、R4Pam2Cys、又はFUGENEと複合体を形成した、緑色蛍光タンパク質をコードするプラスミドの非存在下又は存在下において2×105個の細胞/mlの濃度で、37℃、5%CO2で48時間、培養した。トランスフェクション効率は、フローサイトメーターで細胞の蛍光を測定することによって決定した。
【図8】非免疫処置マウス、及びR4Pam2CysとDNAの複合体か又はインフルエンザウイルスかのいずれかで免疫処置されたマウスにおけるHA特異的抗体レベルのグラフ表示である。BALB/cマウス(週齢6〜8週間)に、単独か、又は1:3若しくは1:5のモル比でR4(S2Pam2Cys)と複合体を形成したかのいずれかの、インフルエンザ血球凝集素をコードするDNAを含む50μgのDNAプラスミドを0日目及び28日目に尾の付け根に皮下接種した。陽性対照として、マウスをまた、スプリットPR8ウイルス(3μg/マウス)で鼻腔内に免疫処置した。非免疫処置マウスは陰性対照としての役割を果たした。最後の接種後14日目にマウスから血清を採取し、ELISAを行って、インフルエンザPR8ウイルスに特異的な抗体の存在を検出した。
【図9】R4Pam2CysとDNAの複合体で免疫処置されたマウスの脾臓(A)及び鼠径リンパ節(B)における誘導された抗原特異的IFN−γ産生CD8+ T細胞の数のグラフ表示である。BALB/cマウス(週齢6〜8週間)に、単独か、又は1:1、1:2、若しくは1:5のNH3+:PO4−モル比でR4(S2Pam2Cys)と複合体を形成した、インフルエンザ核タンパク質をコードする20μgのDNAプラスミドを尾の付け根に皮下接種した。R4(S2Pam2Cys)を含む溶液の10μlのアリコートをDNAの溶液に2分間ごとに合計1時間、ゆっくり加えることによって、複合体形成を達成した。全ての接種材料を0.7MのNaClに溶解した。免疫処置後7日目又は10日目にマウスから脾臓及び鼠径リンパ節を摘出し、細胞内サイトカイン染色を行って核タンパク質147〜155に特異的なIFN−γ分泌CD8+ T細胞の存在を検出した。それぞれの棒は2匹のマウスからの平均及び標準誤差を表す。
【発明を実施するための形態】
【0023】
驚くべきことに、本発明者らは、本発明の化合物が、DNAと安定な複合体を形成することを見出した。DNAは、上記化合物のカチオン性部分に結合し、この複合体が、上記化合物のTLRリガンド部分を介して、上記リガンドを認識する受容体を発現する細胞、例えば、DCなどの抗原提示細胞にターゲットされる。
【0024】
本発明の化合物によって提供される一つの利点は、それらがTLR発現細胞をターゲットする能力があることである。TLRへの結合の結果として、本発明の化合物は、内部に取り入れられ、そしてまた、TLR媒介性シグナル伝達経路を誘導する。特定の実施形態において、本発明の化合物のPam2Cys部分によるTLR−2の結合は、DC成熟を引き起こし、その結果として、DCのリンパ節への遊走及びTリンパ球への効率的な抗原提示をもたらす。
【0025】
第1態様において、本発明は、少なくとも1個のTLRリガンドに連結した正荷電基を含む化合物を提供する。いくつかの実施形態において、本発明の化合物は、少なくとも1個のTLRリガンドに共有結合した正荷電基を含む。
【0026】
第2態様において、本発明は、核酸と、少なくとも1個のTLRリガンドに連結した正荷電基を含む化合物とを含む複合体であって、上記核酸が、核酸と正荷電基との間の静電相互作用によって上記化合物と会合している、複合体を提供する。
【0027】
第3態様において、本発明は、トランスフェクション方法であって、核酸と、少なくとも1個のTLRリガンドに連結した正荷電基を含む化合物とを含む複合体に、TLRを発現する細胞を接触させる段階を含み、上記核酸が、核酸と正荷電基との間の静電相互作用によって上記化合物に会合している、方法を提供する。
【0028】
第4態様において、本発明は、抗原に対する免疫応答を生じさせる方法であって、核酸と、少なくとも1個のTLRリガンドに連結した正荷電基を含む化合物とを含む複合体を対象に投与する段階を含み、上記核酸が、核酸と正荷電基との間の静電相互作用によって上記化合物に会合しており、かつ上記核酸が上記抗原又はそのエピトープをコードする、方法を提供する。
【0029】
第5態様において、本発明は、抗原に対する免疫応答を生じさせる方法であって、核酸と、少なくとも1個のTLRリガンドに連結した正荷電基を含む化合物とを含む複合体をトランスフェクションされた細胞を対象に投与する段階を含み、上記核酸が、核酸と正荷電基との間の静電相互作用によって上記化合物に会合しており、かつ上記核酸が上記抗原又はそのエピトープをコードする、方法を提供する。
【0030】
第6態様において、本発明はまた、TLRを発現する細胞で遺伝子の発現を抑圧する方法であって、核酸と、少なくとも1個のTLRリガンドに連結した正荷電基を含む化合物とを含む複合体を対象に投与する段階を含み、上記核酸が、核酸と正荷電基との間の静電相互作用によって上記化合物に会合しており、上記核酸がsiRNA、shRNA、siRNAをコードするDNA、及びshRNAをコードするDNAからなる群より選択され、かつ上記遺伝子をターゲットとする、方法を提供する。
【0031】
追加の態様において、本発明はまた、対象における免疫応答の誘導のためのワクチンの製造、遺伝子異常又は遺伝子欠損をもつ対象の処置のための医薬の製造、遺伝子の異常な発現又は別なふうに望ましくない発現を患っている対象の処置のための医薬の製造、及び細胞のトランスフェクションのための試薬の製造における、少なくとも1個のTLRリガンドに連結した正荷電基を含む化合物の使用を提供する。
【0032】
本発明は、TLRリガンド、すなわち、細胞表面TLRに結合する部分に連結した正荷電基を含む化合物を提供する。TLRリガンドのTLRへの結合の結果として、TLRを発現する細胞による上記化合物の取り込み、及び/又はTLR媒介性シグナル伝達経路を介するシグナル伝達が生じる。
【0033】
本明細書に用いられる場合、用語「TLR」は、1個又は複数のToll様受容体を意味し、微生物由来の、構造的に保存された分子に結合する1クラスの膜結合型受容体として定義することができる。これまで、13個のTLR、TLR−1〜TLR−13が同定されており、たいていの哺乳動物種は、10個〜15個の型のToll様受容体を有すると推定されている(Du,X.ら 2000 Eur.Cytokine Netw.11:362〜371;Chuang,T.H.及びUlevitch,R.J.2000 Eur.Cytokine Netw.11:372〜378;Tabeta,K.ら 2004 Proc.Natl.Acad.Sci.USA 101:3516〜3521)。TLRは、一種のパターン認識受容体(PRR)であり、それらのリガンドは、集合的に、病原体関連分子パターン(PAMP)として知られている。
【0034】
本明細書に用いられる場合、「TLRリガンド」は、TLRに選択的又は優先的に結合する分子を意味する。TLRリガンドの例は、病原体における保存された特徴を含み、細菌細胞表面リポ多糖(LPS)、リポタンパク質、リポペプチド、及びリポアラビノマンナン;細菌鞭毛由来のフラゲリンなどのタンパク質;ウイルスの二本鎖RNA、又は細菌及びウイルスのDNAの非メチル化CpGモチーフ;並びに特定の他のRNA及びDNAが挙げられる。TLRの内因性リガンドもまた同定されており、それらには、フィブリノーゲン、熱ショックタンパク質(HSP)、及びDNAが挙げられる。
【0035】
表1は、現在、樹状細胞上に発現していることが知られているTLRを、それらの対応するリガンドと共に列挙する。
【0036】
【表1】
【0037】
本発明の例示的化合物は、リポペプチド「Pam2Cys」に連結したカチオン性ペプチドである。当業者は、用語「リポペプチド」が、コンジュゲートしている、1つ又は複数の脂質部分及び1つ又は複数のアミノ酸配列を含む物質の任意の組成物を意味することを理解するであろう。「Pam2Cys」(ジパルミトイル−S−グルセリル−システイン、又はS−[2,3ビス(パルミトイルオキシ)プロピル]システインとしても知られている)が合成されており(Metzger,J.W.ら 1995 J Pept Sci 1:184)、マイコプラズマ ファーメンタンスから単離されたマクロファージ活性化リポペプチドであるMALP−2の脂質部分に対応する(Sacht,Gら 1998 Eur J Immunol 28:4207:Muhiradt,P.F.ら 1998 Infect Immun 66:4804;Muhiradt,P.F.ら 1997 J Exp Med185:1951)。Pam2CysはTLR−2のリガンドであることが知られている。
【0038】
Pam2Cysは、式(I)すなわち
【化1】
の構造を有する。
【0039】
細胞表面TLRをターゲットするために用いることができる他の脂質部分には、パルミトイル、ミリストイル、ステアロイル、ラウロイル、オクタノイル、又はデカノイルが挙げられる。好ましい基としては、Pam2Cys、Pam3Cys、Ste2Cys、Lau2Cys、及びOct2Cysが挙げられる。
【0040】
本発明の正荷電基として、表2に列挙されたカチオン性化合物、並びに米国特許第6,689,478号明細書及び米国特許第4,035,558号明細書に記載されたポリカチオン性化合物が挙げられるが、それらに限定されるわけではない。
【0041】
【表2】
【0042】
核酸は、核酸のバックボーン内のリン酸基により、正味の負電荷を帯びた分子であるため、それらは、本発明の化合物の正荷電基によって静電相互作用を介して結合され、安定な複合体を形成する。
【0043】
一実施形態において、正荷電基は、アルギニン残基又はリシン残基を含む直鎖又は分岐ペプチドである。
【0044】
「核酸」への言及は、デオキシリボ核酸(DNA)及びリボ核酸(RNA)の両方への言及として理解されるべきであり、それらには、二本鎖DNA、二本鎖RNA、一本鎖DNA、一本鎖RNA、低分子干渉RNA(siRNA)、三重鎖、四重鎖、及び任意の多数本鎖(多重鎖)核酸分子、ペプチド核酸(PNA)などの核酸ハイブリッド、デオキシリボ核酸塩基及びリボ核酸塩基を含む分子、並びに任意のヌクレオチド変異体が挙げられる。
【0045】
一実施形態において、核酸分子は、「プラスミド」として提供される。「プラスミド」への言及は、宿主細胞へ伝達可能であり、かつ宿主細胞において複製を起こすことができる核酸分子への言及として理解されるべきである。しかしながら、核酸分子は、プラスミドに限定されるべきではなく、ウイルスDNA又はウイルスRNAを含む任意の核酸分子であってもよい。
【0046】
当業者は、細胞に核酸をトランスフェクションすることが、多くの研究及び臨床の適用に有用であることを理解しているであろう。「トランスフェクション」への言及は、核酸を含む外因性分子が細胞へ導入される過程への言及として理解されるべきである。
【0047】
多くの研究適用は、細胞のトランスフェクションを必要とする。現在、研究者は、細胞への分子の導入、すなわち、細胞のトランスフェクションを、様々な効率をもつ様々な手段によって行う。いくつかのトランスフェクション試薬を、現在、研究者が利用でき、それらには、FUGENEが挙げられる。しかしながら、細胞のトランスフェクション効率は、例えば、細胞型、細胞分裂の速度及び時期、並びにトランスフェクションされることになっている分子及び(1つ又は複数の)トランスフェクション試薬の両方の個々の性質に依存する。
【0048】
本発明の化合物は、TLRを発現する細胞のトランスフェクションに特に有用である。したがって、TLRを天然で発現する細胞か、又はTLR全体であろうと細胞内シグナル伝達ドメインを欠く改変受容体であろうと、TLRを発現するように安定にトランスフェクションされている細胞のいずれかに、本発明の化合物と複合体を形成した核酸をトランスフェクションすることができる。
【0049】
「細胞」は、核酸分子が本発明の化合物によって送達される任意の細胞として理解されるべきである。細胞は、インビトロの細胞、インビボの細胞、又はエクスビボの細胞であってもよい。細胞は、単離されていてもよく、又は生きている動物の器官若しくは組織の一部を形成していてもよい。細胞はまた、細菌、酵母、真菌、カビ、寄生虫、藻類などの微生物であってもよい。細胞はまた、培養動物細胞若しくは細胞株、又は人工細胞であり得る。
【0050】
本発明の特定の実施形態の細胞に関しての、「単離された」への言及は、その本来の環境(例えば、それが自然発生している場合には、自然環境)から取り出されている物質への言及として理解されるべきである。例えば、生きている生物体の組織に存在する細胞は単離されていないが、その同じ細胞が、自然系内に共存する細胞の一部又は全部から分離されている場合、単離されている。
【0051】
本発明の一態様は、TLRリガンドに連結した正荷電基を含む化合物であって、正荷電基と負荷電核酸との間の静電相互作用によって核酸と安定な複合体を形成することができる、化合物を提供する。本発明の化合物は、TLRを発現する細胞に核酸をターゲットする手段を提供する。
【0052】
一実施形態において、本発明は、TLRリガンドに連結したカチオン性ペプチドを含む化合物であって、カチオン性ペプチドと負荷電DNAとの間の静電相互作用によってDNAと安定な複合体を形成することができる、化合物を提供する。そのような化合物は、TLRを発現する細胞、例えば、抗原提示細胞、より具体的にはDCに、DNAをターゲットする手段を提供する。
【0053】
別の実施形態において、本発明は、トランスフェクション方法であって、核酸と、TLRに結合する少なくとも1個の部分に連結した正荷電基を含む化合物とを含む複合体に、TLRを発現する細胞を接触させる段階を含み、上記核酸が、核酸と正荷電基との間の静電相互作用によって上記化合物に会合している、方法を提供する。
【0054】
さらなる実施形態において、本発明は、抗原に対する免疫応答を生じさせる方法であって、核酸と、TLRに結合する少なくとも1個の部分に連結した正荷電基を含む化合物とを含む複合体を対象に投与する段階を含み、上記核酸が、核酸と正荷電基との間の静電相互作用によって上記化合物に会合しており、かつ上記核酸が上記抗原又はそのエピトープをコードする、方法を提供する。
【0055】
さらに別の態様において、本発明は、抗原に対する免疫応答を生じさせる方法であって、核酸と、TLRに結合する少なくとも1個の部分に連結した正荷電基を含む化合物とを含む複合体をトランスフェクションされた細胞を対象に投与する段階を含み、上記核酸が、核酸と正荷電基との間の静電相互作用によって上記化合物に会合しており、かつ上記核酸が上記抗原又はそのエピトープをコードする、方法を提供する。
【0056】
本発明の化合物をDCに接触させる段階は、結果として、細胞表面MHCクラスII発現の増加によって示されるようなDCの成熟又は活性化を生じさせることができる。したがって、TLRリガンドを含む化合物は、DCをターゲットするためだけでなく、TLR及びMyD88などの下流シグナル伝達分子を介してのDCの活性化のために有用であり得る。
【0057】
それゆえに、本発明の化合物は、DCを活性化するのに有用である。樹状細胞が免疫応答を誘導するために、樹状細胞はまず、それらがリンパ節に遊走し、かつTリンパ球を活性化するために必要な接着分子及び共刺激分子を発現するように、活性化されなければならない。DNAと複合体を形成したTLRリガンドを含む化合物は、DCを活性化するのに特に有用であり、DNAがタンパク質をコードする場合、トランスフェクションされたDCは、翻訳された、DNAによりコードされるタンパク質のペプチド断片を免疫細胞へ提示し、それによって、細胞性及び体液性免疫応答を呼び起こす。換言すれば、本発明の特定の実施形態の化合物は、DNAと複合体を形成した場合、DNAワクチン接種アプローチにとって有用である。
【0058】
本明細書に用いられる場合、「DNAワクチン接種」への言及は、DNAによりコードされるタンパク質に対する免疫応答を誘導するための、対象へのDNAの投与への言及を意味する。本発明の化合物は、限定されるわけではないが、静脈内、鼻腔内、筋肉内、経口、直腸などを含む任意の経路によって対象へ直接、投与されてもよい。
【0059】
本明細書に用いられる場合、「免疫応答」への言及は、侵入した病原体、病原体に感染した細胞若しくは組織、癌性細胞、又は自己免疫若しくは病的炎症の場合には正常なヒト細胞若しくは組織の選択的損傷、破壊若しくはヒト身体からの排除を結果として生じさせるリンパ球、抗原提示細胞、食細胞、顆粒球、及び上記の細胞又は肝臓によって産生される(抗体、サイトカイン、及び補体を含む)可溶性高分子の協調作用への言及を意味する。
【0060】
本明細書に用いられる場合、「Tリンパ球応答」への言及は、Tリンパ球に依存する免疫応答の構成要素(すなわち、ヘルパーTリンパ球、細胞傷害性キラーTリンパ球、又はサプレッサーTリンパ球への、Tリンパ球の増殖及び/又は分化、抗体産生を引き起こすか又は阻止する、Bリンパ球へのヘルパーTリンパ球によるシグナルの供給、細胞傷害性Tリンパ球による特定のターゲット細胞の殺害、並びに他の免疫細胞の機能を調節する、サイトカインなどの可溶性因子の放出)への言及を意味する。
【0061】
「患者」、「対象」、又は「哺乳動物」は、交換可能に用いられ、ヒト患者及び非ヒト霊長類、加えてウサギ、ラット、及びマウスなどの実験動物、並びに他の動物などの哺乳動物を指す。動物には、全ての脊椎動物、例えば、ヒツジ、イヌ、ウシ、ニワトリ、両生類、及び爬虫類などの哺乳動物及び非哺乳動物が挙げられる。
【0062】
本発明の化合物は、DNAワクチン接種に用いることに加えて、いくつかの他の臨床適用に有用である。例えば、自己免疫疾患又は癌の処置は、造血性幹細胞(HSC)又は末梢血単核細胞(PBMC)などの自己細胞のトランスフェクションを必要とすることがある。そのような細胞を、対象から採取し、インビトロで培養し、任意で当技術分野においてよく知られた方法によってDCへ分化させ、かつ/又は核酸をトランスフェクションし、その後、対象へ注入して戻すことができる。いくつかの実施形態において、核酸はワクチン抗原をコードし、トランスフェクションされた細胞は、免疫応答を呼び起こすように意図されてもよい。他の実施形態において、核酸は、サイトカイン又は抗原性寛容原をコードしてもよい。
【0063】
別の態様において、本発明は、TLRを発現する細胞で遺伝子の発現を抑圧する方法であって、核酸と、TLRリガンドに連結した正荷電基を含む化合物とを含む複合体を対象に投与する段階を含み、上記核酸が、核酸と正荷電基との間の静電相互作用によって上記化合物に会合しており、上記核酸がsiRNA、shRNA、siRNAをコードするDNA、及びshRNAをコードするDNAからなる群より選択され、上記siRNA又はshRNAが上記遺伝子をターゲットとする、方法を提供する。
【0064】
「処置すること」又は「処置」は、疾患の症状、合併症、又は生化学的徴候の発生を予防し、又は遅らせて、疾患、状態、若しくは障害(例えば、感染性疾患、炎症、又は自己免疫疾患)の症状を軽減するか、若しくは寛解させ、又はそのさらなる発生を抑止若しくは阻害するための本発明の組成物、化合物、又は薬剤の投与を含む。「処置すること」はさらに、疾患、状態、又は障害(例えば、感染性疾患、炎症、又は自己免疫疾患)の処置又は寛解又は予防における任意の成功のきざしを指し、それらには、軽減、寛解、症状の減少若しくは疾患状態を患者にとってより許容できるものにすること、変性若しくは減退の速度を遅らせること、又は変性の最終点を衰弱がより少ないものにすることなどの任意の客観的又は主観的パラメーターが挙げられる。症状の処置又は寛解は、医師による検査の結果を含む、客観的又は主観的パラメーターに基づくことができる。したがって、用語「処置すること」は、感染性疾患、炎症、又は自己免疫疾患に関連した症状又は状態の発生を予防する若しくは遅延させるための、軽減するための、又は抑止若しくは阻害するための本発明の化合物又は薬剤の投与を含む。用語「治療効果」は、対象における疾患、疾患の症状、又は疾患の副作用の低減、排除、又は予防を指す。本発明の方法を用いる「処置すること」又は「処置」には、感染性疾患、炎症、又は自己免疫疾患のリスクの増加があり得るが、まだ症状を経験していない、又は現していない対象において症状の発生を予防すること、感染性疾患、炎症、又は自己免疫疾患の症状を阻害すること(その発生を遅らせること、又は抑止すること)、感染性疾患、炎症、又は自己免疫疾患の症状又は副作用からの解放を提供すること(対症療法を含む)、及び感染性疾患、炎症、又は自己免疫疾患の症状を軽減すること(退行を引き起こすこと)が挙げられる。処置は、疾患又は状態の顕在化後、症状の予防的(疾患の発生を予防し、若しくは遅延させるための、又はその臨床的若しくは潜在性症状の顕在化を予防するための)又は治療的抑制又は軽減であり得る。
【0065】
追加の態様において、本発明は、対象における免疫応答の誘導のためのワクチンの製造、遺伝子異常又は遺伝子欠損をもつ対象の処置のための医薬の製造、遺伝子の異常な発現又は別なふうに望ましくない発現を患っている対象の処置のための医薬の製造、及び細胞のトランスフェクションのための試薬の製造における、TLRリガンドに連結した正荷電基を含む化合物の使用を提供する。
【0066】
本明細書に示された化合物及び方法は、医学的適用及び獣医学的適用の両方に用いることができる。典型的には、生成物をヒトに用いるが、他の哺乳動物に用いてもよい。用語「患者」及び「対象」は、哺乳動物の個体を示すように意図され、本明細書を通じて、及び特許請求の範囲においてそのように用いられる。本発明の主要な適用は、ヒト患者又は対象に関係しているが、本発明は、実験動物、家畜、動物園動物、野生動物、ペット、又は競技用動物に適用してもよい。
【0067】
当業者が、所望の適用に適するように、正荷電基とTLRリガンドとの任意の組合わせを導入してもよいことは理解されているであろう。
【0068】
これより、以下の非限定的実施例を参照して、本発明をさらに記載する。
【0069】
実施例1
カチオン性ペプチドの合成
分岐ペプチド構築物については、リシン残基を用いて足場鋳型の分岐点を供給することによって合成した。足場内に存在する分岐点の数に依存して、ジカチオン性又はテトラカチオン性構造が生じた(図1)。これらの分岐構築物を、PEG−S RAM樹脂(Rapp Polymere、Tubingen、Germany;置換係数(substitution factor)0.27mmol/g)上で合成した。まず、等モル量のO−ベンゾトリアゾール−N,N,N,N’,N’−テトラメチル−ウロニウム−ヘキサフルオロホスフェート(HBTU;Novabiochem、Darmstadt、Germany)、1−ヒドロキシベンゾトリアゾール(HOBt)、及び1.5倍モル過剰のジイソプロピルエチルアミン(DIPEA;Sigma、Castle Hill、Australia)と共に、Fmoc−リシン(Mtt)−OH(Novabiochem、Laufelfingen、Switzerland)を4倍過剰で、樹脂に結合させた。アシル化を40分間行い、完全な反応をトリニトロベンジルスルホン酸(TNBSA)試験によって確認した(21)。α−アミノ基上のFmoc保護基の除去を2.5%ジアザビシクロ[5.4.0]ウンデカ−7−エン(Sigma、Steinheim、Germany)で達成し、Fmoc−リシン(Fmoc)−OH(Auspep、Melbourne、Australia)を、Fmoc基の除去後、2つの一級アミノ基を露出させて分岐点として働くように、結合させた。ジメチルホルムアミド(DMF;Auspep、Melbourne、Australia)を用いて、それぞれのアシル化段階と脱保護段階の間に樹脂を洗浄した。後続のアミノ酸を4倍過剰で結合させ、アシル化を60分間行った。四価の構築物の組み立てについて、アシル化のさらなるラウンドを、Fmoc−リシン(Fmoc)−OHを用いて60分間行い、4つの分岐点を生じさせた。アミノ酸を含む基の四価アルギニン構築物(R4)への化学選択的ライゲーションを可能にするために、システインを構築物のC末端に挿入し、R4−Cysを生じさせた。分岐ペプチド構築物上へのカルボキシフルオレセイン結合について、最後のアシル化反応後、N末端Fmoc基を除去しなかった。その代わり、C末端リシンのε−アミノ基上に存在するMtt保護基をジクロロメタン中1%TFA(Ajax Finechem、Seven Hills、Australia)での処理によって除去した。その後、等モル量のHOBt、HBTU、及び1.5倍モル過剰のDIPEAの存在下で、5(6)−カルボキシフルオレセイン(Fluka BioChemika、Switzerland)を4倍過剰用いて、暗闇中18時間、露出したε−アミノ基に結合させた。アシル化後、樹脂をDMFで洗浄し、N末端Fmoc基を除去した。全てのペプチドを同時に、固相支持体及び側鎖保護基から切断した。ウォーターズ(Waters)3 HPLCクロマトグラフィーシステムに設置されたバイダック(Vydac)C4カラム(4.6mm×250mm)又はC8カラム(4.6mm×250mm)を用いて逆相分析クロマトグラフィーによってペプチドの純度を評価した。限界溶媒としてH2O中0.1%TFA及びアセトニトリル中0.1%TFAを用いる1ml/分の流速を用いて、クロマトグラムを展開した。必要な場合、セミ分取用ウォーターズ又はGBC HPLCシステム及びセミ分取用バイダックC4カラム(10mm×300mm)を用いて2.5ml/分の流速でペプチドを精製した。ペプチド含有量の推定を、蛍光ペプチドの吸光度を496nmで測定するUV分光光度法によって測定し、83,000M−1cm−1のモル吸光係数を用いて濃度を計算した。合成された蛍光分岐ペプチド構築物の模式図は、図1に示されている。
【0070】
ペプチドの脂質付加について、DMF中50%DCMにおける等価量のHOBt及びDICIの存在下で、4倍過剰のN−フルオレニルメトキシカルボニル−S−(2,3−ジヒドロキシプロピル)−システイン(Fmoc−Dhc−OH)をC末端リシンのε−アミノ基上に結合させることによって脂質部分を構築した。アシル化を40分間行い、結合の成功がトリニトロベンゼンスルホン酸試験を用いて確認されるまで、繰り返した。樹脂結合Fmoc−Dhc−ペプチドの2つのヒドロキシ基の脂質付加を、等モル量のDMAP、10倍過剰の脂肪酸、及び12倍過剰のDICIを含む溶液を用いて一晩、行った。
【0071】
実施例2
R4によるDNA移動の遅延
分岐ペプチドR4とDNAの間の会合を調べるために、緑色蛍光タンパク質及びインフルエンザHAタンパク質をそれぞれコードするDNAプラスミドpEGFP−N1又はpCI−HAの一定量を、様々な量のR4とインキュベートし、アガロースゲル電気泳動によって分析した(図2)。
【0072】
各試料中のR4の量を増加させることによって、DNA移動に影響を及ぼすことができた。臭化エチジウム蛍光によって可視化されたように、陽極へのDNAプラスミド移動の部分的遅延は、0.1〜0.5μgのR4を用いた場合、明らかであり、より多い量のR4とインキュベートされたDNA試料はより遅い速度で動いた。開始点におけるDNAのバンドの出現によって明らかなように、10μgのR4が存在した場合、DNA移動の完全な遅延が達成された。しかしながら、このバンドは、全くかすかにしか見えず、この試料中にほとんどDNAがないことを示唆した。これは、R4が臭化エチジウムのDNAへの結合を阻害すること、及び/又はR4とDNAの間の相互作用の結果として、アガロースゲル中で移動するよりむしろ電気泳動緩衝液中へ拡散する不溶性複合体の形成をもたらすことが原因である可能性がある。これらの結果は、R4の正荷電アミノ基及びグアニジン基がDNAバックボーン内の負荷電リン酸基を中和して、陽極へ移動するその能力を遅延させ得ることを示唆する。
【0073】
実施例3
R4−DNA複合体の形成
R4のDNAへの結合を確認するために、漸増量のR4及びDNAを含む遠心分離した混合物の上清におけるDNAの量を、260nmにおいて吸光度を測定することによって決定した(図3A)。DNAをその電荷の中和によって溶液から沈殿させるR4の能力は、溶液中のDNAの減少を生じさせ、それにしたがって、それら2つの間の会合を示すことになるだろう。遠心分離された上清におけるDNAの量は、0.1μg〜5μgのR4とインキュベートした場合、劇的には変化しなかった。しかしながら、7.5μgのR4を用いた場合、わずかな減少が生じ、これより多い量を含む溶液中において、DNAは検出されなかった。この結果は、存在するR4の量がDNAの溶液からの沈殿と比例することを示す。吸光度の測定値をまた、いかなる遠心分離された不溶性物質も再懸濁するために用いられた洗浄上清についても取った。これらの試料中にDNAがほとんど存在しないことは、R4−DNA沈殿物が安定であること、及びDNAがこの複合体から解離する可能性は低いことを示唆する。
【0074】
アガロースゲル分析(図3B)は、DNAバンドが、0.1〜7.5μgのR4を含む上清試料においては明らかであったが、10μg又は15μgのR4を含む試料においては明らかではなかったことを示す。DNA移動の遅延はまた、1μg又はそれ以上のR4を含む試料において明らかである。
【0075】
実施例4
R4(S2Pam2Cys)−DNAでのDCのトランスフェクション
R4構築物又はR4(S2Pam2Cys)構築物がトランスフェクションされたDNAの発現を誘導する能力があるかどうかを決定するために、マウスDC系、D1にpEGFPプラスミドをR4又はR4(S2Pam2Cys)を用いてトランスフェクションした(図4A)。これらの細胞における緑色蛍光タンパク質(GFP)の発現レベルを、市販されているトランスフェクション試薬FUGENEを用いてトランスフェクションしたD1細胞における緑色蛍光タンパク質の発現レベルと比較した。
【0076】
未処理の細胞、DNAのみで処理された細胞、及びDNAと共にR4で処理された細胞において、蛍光は観察されなかった。しかしながら、R4(S2Pam2Cys)及びDNAをトランスフェクションされた細胞は、pEGFP及びFUGENE試薬をトランスフェクションされた細胞において観察された発現レベルに匹敵するレベルでGFPを発現した。
【0077】
D1細胞培養物はまた、それぞれ未熟細胞及び成熟細胞に対応する、MHCクラスII低か又はMHCクラスII高のいずれかである2つの別々の細胞集団で構成されることが見出された(図4B)。未処理のD1細胞の約4%はMHCクラスII高であった。DNA単独での処理又はR4とのDNAでの処理は、D1細胞に関してMHCクラスIIの発現を感知できるほどには変化させなかった。DNA及びFUGENEで処理されたD1細胞は、約36%のMHCクラスII高細胞を含み、R4(S2Pam2Cys)及びDNAで処理されたD1細胞は、約82%のMHCクラスII高細胞を含んだ。
【0078】
DNAに対するトランスフェクション試薬の比が異なるそれらの混合物を用いるトランスフェクション効率もまた、一定量(1μg)のpCI−HAプラスミドとあらかじめインキュベートした漸増量(0.1μg、1μg、又は3μg)のR4、R4(S2Pam2Cys)、又はFUGENEとD1細胞をインキュベートすることによって調べた(図5)。細胞表面インフルエンザHAタンパク質発現を、HA特異的抗体を用いて測定した。R4(S2Pam2Cys)又はFUGENEを用いた場合、表面HAタンパク質を発現する細胞のパーセンテージによって示されるように、トランスフェクション効率は、用いられるトランスフェクション剤の量と比例した。これの例外は、R4を用いた場合であった。この場合、HA発現レベルは、R4(S2Pam2Cys)又はFUGENEを含むDNA混合物によって誘導されるHA発現レベルより低かったが、DNAに対するR4の重量対重量比が同等である時、最適であるように思われた。しかしながら、より高い比率は、トランスフェクション効率にとって有害であることが見出された。
【0079】
調べられた比率の全部において、R4(S2Pam2Cys)は、抗原の発現を誘導することにおいてFUGENEより優れていることが見出された。R4(S2Pam2Cys)及びDNAを用いてトランスフェクションされた細胞の最大パーセンテージは、3:1比において60%であった。相対的に、DNAに対するEUGENEの等価比率を用いた時、約30%のHA陽性細胞だけが、検出された。
【0080】
実施例5
ヒト単球由来樹状細胞(MoDC)のトランスフェクション
ヒト単球由来樹状細胞(MoDC)のトランスフェクションを増強するR4(S2Pam2Cys)の能力を調べた。MoDCにpEGFPプラスミドをトランスフェクションした実験において(図6A)、R4(S2Pam2Cys)のみが、GFPの発現を誘導することにおいて効率的であった。FUGENEか又はR4のいずれかを加えたpEGFPでの細胞のトランスフェクションは、相対的に非効率であり、これらの細胞におけるトランスフェクションレベルは、プラスミド単独で処理されたものと類似していた。この結果は、MoDCに、各トランスフェクション試薬と共にpCI−HAプラスミドをトランスフェクションした実験において、繰り返された(図6B)。
【0081】
実施例6
R4(S2Pam2Cys)−DNAでの非DC細胞系のトランスフェクション
DNAと複合体を形成した、R4構築物又はR4(S2Pam2Cys)構築物が非DC細胞系をトランスフェクションする能力があるかどうかを調べるために、ヒト肺上皮細胞系A549及びマウス肺胞マクロファージ細胞系MH−Sに、R4又はR4(S2Pam2Cys)を用いてpEGFPプラスミドをトランスフェクションした(図7A及び7B)。ヒト肺上皮細胞系A549(A)又はマウス肺胞マクロファージ細胞系MH−S(B)を、異なる希釈度のR4、R4Pam2Cys、又はFUGENEと複合体を形成した、緑色蛍光タンパク質をコードするプラスミドの非存在下又は存在下において2×105個の細胞/mlの濃度で、37℃、5%CO2で48時間、培養した。トランスフェクション効率については、フローサイトメーターで細胞の蛍光を測定することによって決定した。これらの細胞系の両方とも他の研究においてTLR−2を発現することが報告されている(Oshikawa 2003、Slevogt 2007)。未処理のA549又はMH−S細胞、及びDNAと共にR4で処理された細胞において、皆無かそれに近い蛍光が観察された。しかしながら、R4(S2Pam2Cys)及びDNAをトランスフェクションされたA549又はMH−S細胞は、pEGFPプラスミド及びFUGENE試薬をトランスフェクションされた細胞において観察された発現レベルと、より高くはないにしても、匹敵するレベルでGFPを発現した。
【0082】
実施例7
R4(S2Pam2Cys)−DNAで免疫処置されたマウスにおける特異的抗体産生
DNAと複合体を形成したR4(S2Pam2Cys)の抗体を誘導する能力を評価した。BALB/cマウス(週齢6〜8週間)に、単独か、又は1:3若しくは1:5のモル比でR4(S2Pam2Cys)と複合体を形成した、インフルエンザ血球凝集素をコードするDNAを含む50μgのDNAプラスミドを0日目及び28日目に尾の付け根に皮下接種した。陽性対照として、マウスをまた、スプリットPR8ウイルス(3μg/マウス)で鼻腔内に免疫処置した。非免疫処置マウスは陰性対照としての役割を果たした。最後の接種後14日目にマウスから血清を採取し、ELISAを行って、インフルエンザPR8ウイルスに特異的な抗体の存在を検出した。血清抗HA抗体をELISAによって測定した。非免疫処置マウス又は「ナイーブ」マウスにおいてHA特異的抗体は検出されなかったが、1:3及び1:5のモル比で混合された2つの用量のR4(S2Pam2Cys)及びDNAで免疫処置されたマウスにおいて、インフルエンザウイルスに結合することができる実質的レベルの抗体が検出された(図8)。
【0083】
実施例8
R4(S2Pam2Cys)−DNAで免疫処置されたマウスにおけるインフルエンザ核タンパク質特異的IFN−γ陽性CD8+T細胞の誘導
DNAと複合体を形成したR4(S2Pam2Cys)の細胞性免疫応答を誘導する能力を評価した。BALB/cマウス(週齢6〜8週間)に、単独か、又は1:1、1:2、若しくは1:5のNH3+:PO4−モル比でR4(S2Pam2Cys)と複合体を形成した、インフルエンザ核タンパク質をコードする20μgのDNAプラスミドを尾の付け根に皮下接種した。R4(S2Pam2Cys)を含む溶液の10μlのアリコートをDNAの溶液に2分間ごとに合計1時間、ゆっくり加えることによって、複合体形成を達成した。全ての接種材料を0.7MのNaClに溶解した。免疫処置後7日目又は10日目にマウスから脾臓及び鼠径リンパ節を摘出し、細胞内サイトカイン染色を行って核タンパク質147〜155に特異的なIFN−γ分泌CD8+T細胞の存在を検出した。それぞれの棒は2匹のマウスからの平均及び標準誤差を表す。
【0084】
DNA単独で接種されたマウスの脾臓及び鼠径リンパ節では非常に低レベルの活性化CD8+T細胞しか検出されず(それぞれ、図9A及び9B)、ナイーブマウスでは検出されなかった。しかしながら、異なる比率のDNAに対するR4(S2Pam2Cys)を含む複合体の投与は、免疫処置後7日目、脾臓において検出可能なNP特異的IFN−γ産生CD8+T細胞を誘導した(図9A)。特に、1:1比のR4(S2Pam2Cys)とDNAを接種されたマウスのリンパ節及び脾臓の両方において、高レベルの活性化CD8+T細胞が検出された(図9A及び9B)。
【0085】
参考文献
Alexopoulou L., Holt A. C., Medzhitov R. and Flavell R. A. (2001) Recognition of double−stranded RNA and activation of NF−kappaB by Toll−like receptor 3. Nature 413 (6857): 732−738
Asea A., Rehli M., Kabingu E., Boch J. A., Bare O., Auron P. E., Stevenson M. A. and Calderwood S. K. (2002) Novel signal transduction pathway utilized by extracellular HSP70: role of toll−like receptor (TLR) 2 and TLR4. J Biol Chem 277 (17): 15028−15034
Billingham J., Breen C., Rawson J. O., Yarwood J., Mann B.E. (1997) Adsorption of polycations on clays: A comparative in situ study using 133Cs and 23Na solution phase NMR. Journal of Colloid and Interface Science 193:183−189
Bulut Y., Faure E., Thomas L., Karahashi H., Michelsen K. S., Equils O., Morrison S. G., Morrison R. P. and Arditi M. (2002) Chlamydiai heat shock protein 60 activates macrophages and endothelial cells through Toll−like receptor 4 and MD2 in a MyD88−dependent pathway. J.Immunol 168 (3); 1435−1440
Buschle M, Schmidt W, Zauner W, Mechtler K, Trska B, Kirlappos H, et al. (1997) Transloading of tumor antigen−derived peptides into antigen−presenting cells. Proc Natl Acad Sci USA 94(7):3256−3261
Chow J. C., Young D. W., Golenbock D. T., Christ W. J. and Gusovsky F. (1999) Toll−like receptor−4 mediates lipopolysaccharide−induced signal transduction. J−Biol Chem 274 (16): 10689−10692
Christiaens B, Grooten J, Reusens M, Joliot A, Goethals M, Vandekerckhove J, Procbiantz A, Rosseneu M. (2004) Membrane interaction and cellular internalization of penetratin peptides. Eur J Biochem. 271(6);1187−1197
Fawell S, Seery J, Daikh Y, Moore C, Chen LL, Pepinsky B, et al. (1994) Tat−mediated delivery of heterologous proteins into cells. Proc Natl Arad Sci USA 91(2);664− 668
Flacher V, Bouschbacher M, Verronese E, Massacrier C, Sisirak V, Berthier−Vergnes O, de Saint−Vis B, Caux C, Dezutter−Dambuyant C, Lebecque S, Valladeau J. (2006) Hiuman Langerhans cells express a specific TLR profile and differentially respond to viruses and Gram−positive bacteria. J Immunol. 177(11):7959−7967
Futaki S, Suzuki T, Ohashi W, Yagami T, Tanaka S, Ueda K, et al. (2001) Argininerich peptides. An abundant source of membrane−permeable peptides having potential as carriers for intracellular protein delivery, J Biol Chem 276(8):5836−5840
Hayashi F., Smith K. D., Ozinsky A., Hawn T. R., Yi E. C., Goodlett D. R., Eng J. K., Akira S., Underhill D. M. and Aderem A. (2001) The innate immune response to bacterial flagellin is mediated by Toll−like receptor 5. Nature 410 (6832): 1099−1103
Heil F., Hemmi H., Hochrein H., Ampenberger F., Kirschning C., Akira S., Lipford G., Wagner H and Bauer S. (2004) Species−specific recognition of single−stranded RNA via toll−like receptor 7 and 8. Science 303 (5663): 1526−1529
Hemmi H., Kaisho T., Takeuchi O., Sato S., Sanjo H., Hoshino K., Horiuchi T., Tomizawa H., Takeda K. and Akira S. (2002) Small anti−viral compounds activate immune cells via the TLR7 MyD88−dependent signaling pathway. Nat Immunol 3 (2): 196−200
Hemmi H., Takeuchi O., Kawai T., Kaisho T., Sato S., Sanjo H., Matsumoto M., Hoshino K., Wagner H., Takeda K. and Akira S. (2000) A Toll−like receptor recognizes bacterial DNA. Nature 408 (6813): 740−745
Husebye H, Halaas (, Stenmark H, Tunheim G, Sandanger (, Bogen B, Brech A, Latz E, Espevik T. (2006) Endocytic pathways regulate Toll−like receptor 4 signaling and link innate and adaptive immunity. EMBO J. 25(4):683−692
Mitchell DJ, Kim DT, Steinman L, Fathman CG, Rothbard JB. (2000) Polyarginine enters cells more efficiently than other polycationic homopolymers. J Pept Res 56(5):318−325
Morr M., Takeuchi O., Akira S., Simon M. M. and Muhlradt P. F. (2002) Differential recognition of structural details of bacterial lipopeptides by toll−like receptors. Eur J Immunol 32 (12): 3337−3347
Oehlke J, Scheller A, Wiesner B, Krause E, Beyermann M, Klauschenz E, et al. (1998) Cellular uptake of an alpha−helical amphipathic model peptide with the potential to deliver polar compounds into the cell interior non−endocytically. Biochim Biophys Acta 1414(1−2):127−139
Okusawa T., Fujita M., Nakamura J., Into T., Yasuda M., Yoshimura A., Hara Y., Hasebe A., Golenbock D. T., Morita M., Kuroki Y., Ogawa T. and Shibata K. (2004) Relationship between structures and biological activities of mycoplasmal diacylated lip opeptides and their recognition by toll−like receptors 2 and 6. Infect Immun 72(3): 1657−1665
Oshikawa K, Sugiyama Y. (2003) Regulation of toll−like receptor 2 and 4 gene expression in murine alveolar macrophages. Exp Lung Res 29(6):401−412
Otvos L, Jr., Cudic M, Chua BY, Deliyannis G, Jackson DC. (2004) An insect antibacterial peptide−based drug delivery system. Mol Pharm 1(3):220−232
Ozinsky A., Underhill D. M., Fontenot J. D., Hajjar A. M., Smith K. D., Wilson C. B., Schroeder L. and Aderem A. (2000) The repertoire for pattern recognition of pathogens
by the innate immune system is defined by cooperation between toll−like receptors. Proc Natl Acad Sci USA 97 (25): 13766−13771
Pepper M, Dzierszinski F, Wilson E, Tait E, FangQ, Yarovinsky F, Laufer TM, Roos D, Hunter CA. (2008) Plasmacytoid dendritic cells are activated by Toxoplasma gondii to present antigen and produce cytokines. J Immunol. 2008 180(9):6229−6236
Poltorak A., He X, Smirnova L, Liu M. Y., Van Huffel C., Du X., Birdwell D., Alejos E., Silva M., Galanos C., Freudenberg M., Ricciardi−Castagnoli P., Layton B. and Beutler B. (1998) Defective LPS signaling in C3H/HeJ and C57BL/10ScCr mice: mutations in Tlr4 gene. Science 282 (5396): 2085−2088
Pooga M, Hallbrink M, Zorko M, Langel U. (1998) Cell penetration by transportan. Faseb J 12(1):67−77
Schjetne K. W., Thompson K. M., Nilsen N., Flo T. H., Fleckenstein B., Iversen J. G., Espevik T., and Bogen B. (2003) Cutting edge: link between innate and adaptive immunity: Toll−like receptor 2 internalizes antigen for presentation to CD4+ T cells and could be an efficient vaccine target J Immunol 171:32−36
Schwandner R., Dziarski R., Wesche H., Rothe M. and Kirschning C. J. (1999) Peptidoglycan− and lipoteichoic acid−induced cell activation is mediated by toll−like receptor 2. J Biol Chem 274 (25): 17406−17409
Slevogt H, Seybold J, Tiwari KN, Hocke AC, Jonatat C, Dietel S, Hippenstiel S, Singer BB, Bachmann S, Suttorp N, Opitz B. (2007) Moraxella catarrhalis is internalized in respiratory epithelial cells by a trigger like mechanism and initiates a TLR2− and partly NOD1−dependent inflammatory immune response. Cell Microbiol 9(3):694−707
Takeshita F, Leifer CA, Gursel I, Ishii KJ, Takeshita S, Gursel M, Klinman DM. (2001) Cutting edge: Role of Toll−like receptor 9 in CpG DNA−induced activation of human cells. J Immunol. 167(7):3555−3558
Takeuchi O., Kaufmann A., Grote K, Kawai T., Hoshino K., Morr M., Muhlradt P. F. and Akira S. (2000) Cutting edge: preferentially the R−stereoisomer of the mycoplasmal lipopeptide macrophage−activating lipopeptide−2 activates immune cells through a toll−like receptor 2− and MyD88−dependent signaling pathway. J Immunol 164 (2): 554−557
Takeuchi O., Sato S., Horiuchi T., Hoshino K., Takeda K., Dong Z., Modlin R. L. and Akira S. (2002) Cutting edge: role of Toll−like receptor 1 in mediating immune response to microbial lipoproteins. J Immunol 169 (1): 10−14 .
Wagner E., Zenke M., Cotten M., Beug, H., Birnstiel M. L. (1990) Transferrinpolycation conjugates as carriers for DNA uptake into cells. Proc. Natl. Acad. Sci. USA 87:3410−3414
【特許請求の範囲】
【請求項1】
少なくとも1個のTLRリガンドに連結した正荷電基を含む化合物。
【請求項2】
前記正荷電基がTLRリガンドに共有結合している、請求項1に記載の化合物。
【請求項3】
前記正荷電基が1個又は複数のアミノ酸を含む、請求項1又は請求項2に記載の化合物。
【請求項4】
前記正荷電基が分岐ペプチド又は直鎖ペプチドである、請求項1〜3のいずれか一項に記載の化合物。
【請求項5】
前記ペプチドが少なくとも1個のアルギニン残基又はリシン残基を含む、請求項4に記載の化合物。
【請求項6】
前記ペプチドが少なくとも4個のアルギニン残基又は少なくとも4個のリシン残基を含む、請求項4又は請求項5に記載の化合物。
【請求項7】
前記TLRリガンドが脂質、又はペプチドグリカン、又はリポタンパク質、又はリポ多糖を含む、請求項1〜6のいずれか一項に記載の化合物。
【請求項8】
前記TLRリガンドがパルミトイル、ミリストイル、ステアロイル、ラウロイル、オクタノイル、又はデカノイルを含む、請求項1〜6のいずれか一項に記載の化合物。
【請求項9】
前記TLRリガンドがPam2Cys、Pam3Cys、Ste2Cys、Lau2Cys、及びOct2Cysからなる群より選択される、請求項1〜6のいずれか一項に記載の化合物。
【請求項10】
前記TLRリガンドがTLR−2か又はTLR−6のいずれかに結合する、請求項1〜9のいずれか一項に記載の化合物。
【請求項11】
前記TLRリガンドがTLR−2に結合する、請求項1〜10のいずれか一項に記載の化合物。
【請求項12】
核酸及び請求項1〜11のいずれか一項に記載の化合物を含む複合体であって、前記核酸と前記正荷電基との間の静電相互作用によって前記核酸が前記化合物と会合している、複合体。
【請求項13】
少なくとも1個のTLRを発現する細胞を、請求項12に記載の複合体に接触させる段階を含む、トランスフェクション方法。
【請求項14】
抗原に対する免疫応答を生じさせる方法であって、請求項12に記載の複合体を対象に投与する段階を含み、前記核酸が前記抗原又はそのエピトープをコードする、方法。
【請求項15】
抗原に対する免疫応答を生じさせる方法であって、請求項12に記載の複合体をトランスフェクションされた細胞を対象に投与する段階を含み、前記核酸が前記抗原又はそのエピトープをコードする方法。
【請求項16】
TLRを発現する細胞で遺伝子の発現を抑圧する方法であって、請求項12に記載の複合体を対象に投与する段階を含み、前記核酸が、siRNA、shRNA、siRNAをコードするDNA、及びshRNAをコードするDNAからなる群より選択され、かつ前記siRNA又はshRNAが前記遺伝子をターゲットとする、方法。
【請求項17】
対象における免疫応答の誘導のためのワクチンの製造における、請求項1〜12のいずれか一項に記載の化合物又は複合体の使用。
【請求項18】
遺伝子異常又は遺伝子欠損をもつ対象の処置のための医薬の製造における、請求項1〜12のいずれか一項に記載の化合物又は複合体の使用。
【請求項19】
遺伝子の異常な発現又は望ましくない発現を患っている対象の処置のための医薬の製造における、請求項1〜12のいずれか一項に記載の化合物又は複合体の使用。
【請求項20】
細胞のトランスフェクションのための試薬の製造における、請求項1〜12のいずれか一項に記載の化合物又は複合体の使用。
【請求項1】
少なくとも1個のTLRリガンドに連結した正荷電基を含む化合物。
【請求項2】
前記正荷電基がTLRリガンドに共有結合している、請求項1に記載の化合物。
【請求項3】
前記正荷電基が1個又は複数のアミノ酸を含む、請求項1又は請求項2に記載の化合物。
【請求項4】
前記正荷電基が分岐ペプチド又は直鎖ペプチドである、請求項1〜3のいずれか一項に記載の化合物。
【請求項5】
前記ペプチドが少なくとも1個のアルギニン残基又はリシン残基を含む、請求項4に記載の化合物。
【請求項6】
前記ペプチドが少なくとも4個のアルギニン残基又は少なくとも4個のリシン残基を含む、請求項4又は請求項5に記載の化合物。
【請求項7】
前記TLRリガンドが脂質、又はペプチドグリカン、又はリポタンパク質、又はリポ多糖を含む、請求項1〜6のいずれか一項に記載の化合物。
【請求項8】
前記TLRリガンドがパルミトイル、ミリストイル、ステアロイル、ラウロイル、オクタノイル、又はデカノイルを含む、請求項1〜6のいずれか一項に記載の化合物。
【請求項9】
前記TLRリガンドがPam2Cys、Pam3Cys、Ste2Cys、Lau2Cys、及びOct2Cysからなる群より選択される、請求項1〜6のいずれか一項に記載の化合物。
【請求項10】
前記TLRリガンドがTLR−2か又はTLR−6のいずれかに結合する、請求項1〜9のいずれか一項に記載の化合物。
【請求項11】
前記TLRリガンドがTLR−2に結合する、請求項1〜10のいずれか一項に記載の化合物。
【請求項12】
核酸及び請求項1〜11のいずれか一項に記載の化合物を含む複合体であって、前記核酸と前記正荷電基との間の静電相互作用によって前記核酸が前記化合物と会合している、複合体。
【請求項13】
少なくとも1個のTLRを発現する細胞を、請求項12に記載の複合体に接触させる段階を含む、トランスフェクション方法。
【請求項14】
抗原に対する免疫応答を生じさせる方法であって、請求項12に記載の複合体を対象に投与する段階を含み、前記核酸が前記抗原又はそのエピトープをコードする、方法。
【請求項15】
抗原に対する免疫応答を生じさせる方法であって、請求項12に記載の複合体をトランスフェクションされた細胞を対象に投与する段階を含み、前記核酸が前記抗原又はそのエピトープをコードする方法。
【請求項16】
TLRを発現する細胞で遺伝子の発現を抑圧する方法であって、請求項12に記載の複合体を対象に投与する段階を含み、前記核酸が、siRNA、shRNA、siRNAをコードするDNA、及びshRNAをコードするDNAからなる群より選択され、かつ前記siRNA又はshRNAが前記遺伝子をターゲットとする、方法。
【請求項17】
対象における免疫応答の誘導のためのワクチンの製造における、請求項1〜12のいずれか一項に記載の化合物又は複合体の使用。
【請求項18】
遺伝子異常又は遺伝子欠損をもつ対象の処置のための医薬の製造における、請求項1〜12のいずれか一項に記載の化合物又は複合体の使用。
【請求項19】
遺伝子の異常な発現又は望ましくない発現を患っている対象の処置のための医薬の製造における、請求項1〜12のいずれか一項に記載の化合物又は複合体の使用。
【請求項20】
細胞のトランスフェクションのための試薬の製造における、請求項1〜12のいずれか一項に記載の化合物又は複合体の使用。
【図1】

【図2】

【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【公表番号】特表2010−539992(P2010−539992A)
【公表日】平成22年12月24日(2010.12.24)
【国際特許分類】
【出願番号】特願2010−528245(P2010−528245)
【出願日】平成20年10月9日(2008.10.9)
【国際出願番号】PCT/AU2008/001501
【国際公開番号】WO2009/046498
【国際公開日】平成21年4月16日(2009.4.16)
【出願人】(500074981)ザ ユニバーシティー オブ メルボルン (5)
【Fターム(参考)】
【公表日】平成22年12月24日(2010.12.24)
【国際特許分類】
【出願日】平成20年10月9日(2008.10.9)
【国際出願番号】PCT/AU2008/001501
【国際公開番号】WO2009/046498
【国際公開日】平成21年4月16日(2009.4.16)
【出願人】(500074981)ザ ユニバーシティー オブ メルボルン (5)
【Fターム(参考)】
[ Back to top ]