
ネトリン−1による内皮細胞死の誘導物質選択方法およびその適用
本発明は、内皮細胞、好ましくは血管または新生血管由来の内皮細胞の死を誘発することができる化合物を選択するインビトロでの方法に関する。本発明は、更に内皮細胞、好ましくはネトリン−1を発現する腫瘍の血管または新生血管由来の内皮細胞の死を誘発することができる化合物としてのネトリン−1の機能阻害剤の使用を含んでなる。最後に、本発明は、内皮細胞の死を誘発することができる化合物を選択するためのキットに関する。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、内皮細胞、好ましくは血管または新生血管由来の内皮細胞の死を誘発することができる化合物を選択するためのインビトロ(in vitro)の方法に関する。本発明は更に、内皮細胞、好ましくはネトリン−1を発現する腫瘍の血管または新生血管由来の内皮細胞の死を誘発することができる化合物としてのネトリン−1の機能阻害剤の使用も含んでなる。最後に、本発明は、内皮細胞の死を誘発することができる化合物を選択するためのキットに関する。
【背景技術】
【0002】
軸索誘導分子として最初に報告されたネトリン−1は、分枝器官の形態発生における役割から血管形成における機能までの範囲の神経外過程においても機能することが最近示されている(Cirulli and Yebra, 2007; Liu et al., 2004; Lu et al., 2004; Wilson et al., 2006)。興味深いことには、血管形成中のネトリン−1の役割に関して矛盾する結果が報告された。しかしながら、以下の2つの明らかな発生学的結果は、最初はちぐはぐであると思われた:Eichmann、Tessier−Lavigneと共同研究者らは、マウスにおけるUNC5Bの発生学的不活性化は血管形成の増加と関係しており、それによってネトリン−1の抗血管形成活性が示唆されることをあざやかに立証したが、Liと共同研究者らは、UNC5Bのリガンドをコードするネトリン−1aの不活性化は、ゼブラフィッシュ発生中の血管の喪失と関係していることを示した(Lu et al., 2004; Wilson et al., 2006)。
【0003】
血管形成は、多数の様々な疾病状態の開始および/または拡散における重要な要因であるので、信頼性のあるやり方で血管形成を制御しまたは阻害することができる治療組成物が必要である。実際に、血管形成の過程は、多数の疾病状態で変更されることが分かっており、多くの場合に、疾病と関連する病理学的損傷は抑制されていない血管形成に関係している。例えば、腫瘍は成長するためには新たな毛細血管の成長を刺激し続けなければならないので、血管形成は腫瘍成長における一因子である。従って、血管または新生血管の内皮細胞の死を促進しまたは誘発することができる化合物は、新たな毛細血管の成長を阻害することができ、これらの疾患の治療に有用なことがある。
【発明の開示】
【0004】
従って、本発明の目的は、内皮細胞死の誘導物質としての活性を示す化合物および医薬組成物を提供することである。
【0005】
本発明者らは、報告された、ネトリン−1が血管形成促進性または抗血管形成性であるという矛盾した結論を生じた明らかに対立する観察結果は、ネトリン−1が内皮細胞の生存因子として作用し、一方、その未結合UNC5B受容体が細胞のアポトーシスを誘発するという事実から生じていることを明らかにした。血管発生に対するネトリン−1の効果は、エクスビボ(ex vivo)アッセイにおけるカスパーゼ阻害剤に似ており、カスパーゼ活性の阻害またはUNC5B受容体のようなネトリン−1受容体のサイレンシングは、いずれもゼブラフィッシュにおけるネトリン−1サイレンシングによって誘発される血管発芽欠損を救済するのに十分である。
【0006】
従って、未結合UNC5Bのアポトーシス促進効果と内皮細胞上のネトリン−1に結合したUNC5Bとの生存効果は、血管形成過程中に協同するものと思われる。
【0007】
第一の態様では、本発明は、ネトリン−1が血管形成中の内皮細胞アポトーシスを阻害することを見出したことに基づく内皮細胞の死を誘発することができる化合物を選択するインビトロでの方法に関する。
【0008】
従って、本発明は、内皮細胞死を誘発することができる化合物を選択するインビトロでの方法であって、
a)ネトリン−1、またはその断片、および内皮細胞を含む培地を有し、
b)前記培地を、試験する化合物と接触させ、
c)前記内皮細胞の死を測定し、および
d)段階c)における測定が前記化合物の存在下にて内皮細胞死の有意な誘発を示しているときに、前記化合物を選択する
段階を含んでなる、方法に関する。
【0009】
特定態様では、試験する化合物の存在によって誘発される細胞死は、下記の材料および方法「細胞死分析」の項に示されているトリパンブルー染色法を用いることによって分析しまたは評価することができる(Mehlen et al., Nature, 1998も参照されたい)。
【0010】
もう一つの態様では、本発明は、内皮細胞死を誘発することができる化合物を選択する方法であって、
a)少なくとも1種類のネトリン−1受容体を発現する内皮細胞を含む培地を有し、
b)前記培地を、試験する化合物と接触させ、
c)前記の試験する化合物のこのネトリン−1受容体のアポトーシス促進活性を促進する能力を測定し、および
d)段階c)における測定が、前記化合物が、このネトリン−1受容体の、好ましくは更にネトリン−1の存在下にてこのネトリン−1受容体の、アポトーシス促進活性を有意に促進することを示しているときに、前記化合物を内皮細胞死の誘導物質として選択する
段階を含んでなる、方法に関する。
【0011】
もう一つの態様では、本発明は、
段階c)の測定が、試験する化合物の細胞死関連タンパク質キナーゼ(DAPK)のキナーゼ活性の活性化を誘発する能力を測定することにあり、および
d)段階c)における測定が、前記化合物の存在下にて、好ましくは更にネトリン−1の存在下にて細胞死関連タンパク質キナーゼ(DAPK)のキナーゼ活性の有意な活性化を示しているときに、前記化合物を内皮細胞死の誘導物質インデューサーとして選択する、
本発明による内皮細胞の死を誘発することができる化合物の選択方法に関する。
【0012】
もう一つの態様では、本発明は、
段階c)の測定が、試験する化合物のDAPKリン酸化を阻害する能力を測定することにあり、および
d)段階c)における測定が、前記化合物の存在下にて、好ましくは更にネトリン−1の存在下にてDAPKリン酸化の有意な阻害を示しているときに、前記化合物を内皮細胞死の誘導物質として選択する、
請求項2に記載の内皮細胞死を誘発することができる化合物の選択方法に関する。
【0013】
もう一つの態様では、本発明は、内皮細胞死を誘発することができる化合物の選択方法であって、
a)少なくとも1種類のネトリン−1受容体を発現する内皮細胞を含む培地を有し、かつ前記ネトリン−1受容体細胞内ドメインがネトリン−1の存在下にて二量体化または多量体化することができ、
b)前記培地を、試験する化合物と接触させ、培地が更にネトリン−1、またはネトリン−1受容体の細胞外ドメインと相互作用することができるその断片を含み、
c)前記ネトリン−1受容体細胞内ドメインの二量体化または多量体化が上記の試験する化合物の存在下で阻害されるかどうかを決定し、および
d)段階c)における決定が、前記内皮細胞の前記ネトリン−1受容体の細胞内ドメインの二量体化または多量体化を有意に阻害することを示しているときに、前記化合物を内皮細胞死の誘導物質として選択する
段階を含んでなる、方法に関する。
【0014】
試験する化合物の存在下における前記ネトリン−1受容体細胞内ドメインの二量体化または多量体化の阻害の決定分析は、例えば、2007年9月7日に公表された特許文献Mehlen et al., WO2007/099133号に見出すことができる。
【0015】
好ましい態様では、段階a)の培地はネトリン−1を更に含む。
【0016】
もう一つの好ましい態様では、本発明による内皮細胞の死を誘発することができる化合物を選択する方法において、前記化合物は血管または新生血管の死を誘発することができる。
【0017】
好ましい態様では、本発明による方法の段階a)において、前記内皮細胞は、DCC(Deleted in Colorectal Cancer)、UNC5A、UNC5B、UNC5CおよびUNC5D、ネオゲニンおよびアデノシンA2bの群から選択される少なくとも1種類のネトリン−1受容体、好ましくはUNC5Bを発現する内皮細胞である。
【0018】
ヒトネトリン−1、またはDCC、UNC5A、UNC5B、UNC5CおよびUNC5D、ネオゲニンおよびアデノシンA2b(Unc−5ホモログ1、2および3はUnc−5ホモログA、BおよびCと同義である)のようなヒトネトリン受容体のアミノ酸配列は、当業者に周知である。特定ドメインの局在化を有するこれらのアミノ酸配列の例は、例えば、ヒトネトリン−1についてはGenbankに受入番号AAD09221またはNP004813、ヒトネトリン受容体DCC前駆体についてはNP_005206、ヒトネトリン受容体Unc−5ホモログ1についてはNP588610、ネトリン受容体Unc−5ホモログ2についてはQ8IZJ1、およびUnc−5ホモログ3についてはO95185、およびヒトUNC5DについてはAAQ88514により見出すことができる。
【0019】
更に好ましくは、ネトリン−1受容体UNC5Bを発現する内皮細胞である。
【0020】
もう一つの好ましい態様では、本発明による方法において、段階a)の前記内皮細胞は、ヒトまたはニワトリの内皮細胞からなる群、好ましくはヒトまたはニワトリ血管または新生血管、更に好ましくはヒト臍帯静脈内皮細胞(HUVEC)およびヒト臍帯動脈内皮細胞(HUAEC)またはヒナドリ漿尿膜から選択される内皮細胞である。
【0021】
第二の態様では、本発明は、
ネトリン−1と上記ネトリン−1受容体との相互作用を特異的に阻害することができるネトリン−1受容体またはその断片の細胞外ドメインを含んでなる化合物、
ネトリン−1またはネトリン−1受容体に対して特異的な、特に前記ネトリン−1受容体の細胞外ドメインまたは前記ネトリン−1受容体の細胞外ドメインと相互作用することができるネトリン−1断片に対するモノクローナルまたはポリクローナル抗体、および
ネトリン−1タンパク質をコードする核酸に特異的なアンチセンスおよびRNAi(干渉RNA)オリゴヌクレオチドからなる群から選択されるネトリン−1発現を阻害することができる化合物
からなる群から選択される、内皮細胞死の誘導物質としての、化合物を含んでなる。
【0022】
好ましくは、本発明は、血管または新生血管の死を誘発する内皮細胞死の誘導物質としての化合物に関する。
【0023】
更に好ましい態様では、本発明は、腫瘍の血管または新生血管の死の誘導物質としての化合物に関する。
【0024】
もう一つのより好ましい態様では、前記腫瘍の腫瘍細胞はネトリン−1を発現または過剰発現する。
【0025】
「ネトリン−1と前記ネトリン−1受容体との間の相互作用を特異的に阻害することができる」という表現におけるネトリン−1とそのネトリン−1受容体との間の相互作用という用語は、本明細書では、一般にリガンドとその受容体との間の特異的結合による相互作用を表すことを指している。この相互作用の阻害は、一般的には、特に競合的リガンド(前記ネトリン−1受容体のこの細胞外膜ドメインに対する抗体など)の存在下、またはネトリン−1(そのネトリン−1受容体の可溶性細胞外膜ドメイン、またはその部分など)と特異的複合体を形成することができる化合物の存在下にて、ネトリン−1のその受容体への結合の完全または部分的阻害によって得ることができる。
【0026】
前記ネトリン−1受容体断片は、ネトリン−1受容体の細胞外ドメイン、またはネトリン−1と特異的に相互作用することができるその部分を含んでなるか、またはそれらである。
【0027】
前記ネトリン−1および/または前記ネトリン−1受容体は、好ましくは哺乳類由来であり、特にマウス、ラットまたはヒト由来であり、更に好ましくはヒト由来である。前記ネトリン−1は、組換えネトリン−1であることができる。前記ネトリン−1は、ニワトリ由来であることもできる。
【0028】
本明細書で用いられる「抗体」という用語は、免疫グロブリン分子および免疫グロブリン分子の免疫学的に活性な部分、すなわちネトリン−1タンパク質またはその受容体と特異的に結合する(免疫反応する)抗原結合部位を含む分子を表す。
【0029】
「抗体」という用語は、モノクローナルまたはポリクローナル抗体だけでなく、キメラまたはヒト化抗体をも含んでなる。
【0030】
単離したネトリン−1タンパク質またはネトリン−1受容体タンパク質、またはそれらの特異的断片は、ポリクローナルおよびモノクローナル抗体調製の標準的手法を用いてこのようなタンパク質を結合する抗体を生成するための免疫原として用いることができる。また、これらの特異抗体を生成するのに用いてもよい少なくとも1種類の抗原決定基を含むこれらのタンパク質の任意の断片を用いてもよい。
【0031】
タンパク質免疫原は、典型的には、適当な被検体(例えば、ウサギ、ヤギ、マウスまたは他の哺乳類)を免疫原で免疫することによって抗体を調製するのに用いられる。適当な免疫原製剤は、前記タンパク質またはその断片を含むことができ、更にフロイントの完全または不完全アジュバントのようなアジュバント、または同様な免疫刺激剤を含むこともできる。
【0032】
従って、本発明によって用いられる抗体としては、ポリクローナル、モノクローナルキメラまたはヒト化抗体が挙げられる。ネトリン−1タンパク質またはその受容体のアミノ酸配列の少なくとも8〜10個のアミノ酸の連続スパンを含んでなるポリペプチドを含むエピトープに選択的に結合することができるまたは選択的に結合している抗体。インタクト(intact)抗体またはその機能性断片(例えば、FabまたはF(ab’)2、scFv)を用いることができる。
【0033】
例えば、候補オリゴヌクレオチドの検出のためのインビトロ手法としては、ノーザンまたはサザンブロットハイブリダイゼーションおよびインサイチュー(in situ)ハイブリダイゼーションが挙げられる。候補タンパク質の検出のためのインビトロ手法としては、酵素免疫測定法(ELISA)、ウェスタンブロット、免疫沈澱および免疫蛍光が挙げられる。
【0034】
アンチセンスは、ネトリン−1コード配列のmRNAまたはcDNA、またはその相補性配列に特異的にハイブリダイズする長さが少なくとも10、15または20ヌクレオチドのオリゴヌクレオチドであることができる。ネトリン−1コード配列の対応する断片と少なくとも95%、98%、99%および100%の同一性を有するオリゴヌクレオチドが好ましい。
【0035】
干渉RNA(RNAi)は、二本鎖RNA(dsRNA)がその相補性配列を有する遺伝子の発現を特異的に抑制する現象である。RNAiは、従って多くの生物の有用な研究手段となっている。dsRNAにより遺伝子発現を抑制する機構は完全には理解されていないが、実験データは重要な識見を提供している。この手法は、哺乳類細胞における遺伝子機能を研究する手段として大きな可能性を有しており、siRNA(低分子干渉RNA)に基づく薬剤の開発へと導くことができる。
【0036】
好ましくは、本発明の化合物において、ネトリン−1受容体またはその断片の前記細胞外ドメインは、DCC、UNC5A、UNC5B、UNC5CおよびUNC5D、ネオゲニンおよびアデノシンA2bからなる群から選択され、更に好ましくはDCC、UNC5A、UNC5BおよびUNC5Cの群から選択される。UNC5Bが、さらにより好ましい。
【0037】
もう一つの態様では、本発明は、ネトリン−1を発現または過剰発現する腫瘍細胞を含む腫瘍に罹っている患者を治療するための癌の進行を阻害することを目的とする医薬組成物の調製のための本発明による化合物に関する。
【0038】
好ましくは、前記癌は、乳癌、結直腸癌、肺癌、神経芽腫、神経膠腫、急性脊髄性白血病、肉腫、黒色腫、卵巣腺癌、腎腺癌、膵腺癌、子宮腺癌、胃腺癌、肝腺癌および直腸腺癌からなる群から選択される。
【0039】
好ましくは、前記癌は転移性または侵攻性癌である。
【0040】
本発明の化合物を患者に投与するときには、場合によっては薬学上許容可能なビヒクルを含んでなる組成物の成分として投与するのが好ましい。この組成物は、経口または任意の他の好都合な経路によって投与することができ、別の生物活性剤と共に投与してもよい。投与は、全身または局所的であることができる。例えば、リポソームでのカプセル化、微小粒子、マイクロカプセル、カプセルなどの様々な送達系が知られており、選択された本発明の化合物またはその薬学上許容可能な塩を投与するのに用いることができる。
【0041】
投与方法としては、皮内、筋肉内、腹腔内、静脈内、皮下、鼻内、硬膜外、経口、舌下、鼻内、脳内、膣内、経皮、直腸、吸入、または局所的投与が挙げられるが、これらに限定されない。投与様式は、現場実務者の裁量に任される。ほとんどの場合に、投与によって、化合物は血流中にまたは直接原発性腫瘍に放出される。
【0042】
本発明による、または本発明による方法によって選択された化合物を含んでなる組成物も、本発明の部分を形成する。これらの組成物は、薬学上許容可能なビヒクルの適量をさらに含んでなり、患者に適正に投与するための形態を提供するようにすることもできる。「薬学上許容可能な」という用語は、規制当局によって認可され、または動物、哺乳類、更に詳細にはヒトで使用するための国定または承認された薬局方に記載されていることを意味する。「ビヒクル」という用語は、希釈剤、アジュバント、賦形剤またはキャリヤーであって、これによって本発明の化合物を投与するものを表す。このような薬学ビヒクルは、水および油のような液体であり、ピーナッツ油、大豆油、鉱油、ゴマ油など石油、動物、植物または合成起源のものであることができる。薬学ビヒクルは、塩水、ゼラチン、澱粉などであることができる。更に、補助剤、安定剤、増粘剤、滑沢剤および着色料を用いてもよい。食塩溶液、水性デキストロースおよびグリセロール溶液を、特に注射溶液の液体ビヒクルとして用いることもできる。適当な薬学ビヒクルとしては、澱粉、グルコース、ラクトース、スクロース、ゼラチン、ステアリン酸ナトリウム、モノステアリン酸グリセロール、塩化ナトリウム、乾燥スキムミルク、グリセロール、プロピレン、グリコール、水などの賦形剤も挙げられる。試験化合物の組成物は、所望ならば、少量の湿潤剤または乳化剤、またはpH緩衝剤を含むこともできる。本発明の組成物は、溶液、懸濁液、エマルション、錠剤、丸薬、ペレット、カプセル、液体を含むカプセル、散剤、徐放性処方物、座薬、エマルション、エアゾール、スプレー、懸濁液の形態、または使用に適当な任意の他の形態を採ることができる。前記組成物は、通常は経口投与または静脈内投与の目的でヒトに適合させた医薬組成物として常法に準じて処方される。活性化合物の量または治療に有効となる量は、標準的臨床手法によって決定することができる。更に、インビトロまたはインビボ(in vivo)アッセイを場合によって用いて、最適用量範囲を確認しやすくすることができる。用いられる正確な用量は投与経路および疾病の重篤度によって変化し、現場実務者の判断およびそれぞれの患者の環境によって決定されるべきである。しかしながら、経口、鼻内、皮内または静脈内投与は、通常は約0.01〜約75mg/kg体重/日、更に好ましくは約0.5〜5mg/kg体重/日である。
【0043】
第三の態様では、本発明は、内皮細胞、好ましくは血管または新生血管由来の内皮細胞の死を誘発することができる化合物を選択するためのキットであって、
ネトリン−1タンパク質またはネトリン−1受容体タンパク質と特異的に相互作用して結合対を形成することができるその断片、および
ネトリン−1受容体、好ましくはDCC、UNC5A、UNC5B、UNC5CおよびUNC5D、ネオゲニンおよびアデノシンA2bからなる群から選択され、更に好ましくは受容体UNC5Bを発現する内皮細胞
を含んでなる、キットに関する。
【0044】
好ましい態様では、前記キットは、内皮細胞HUVECまたはHUAEC細胞を含んでなる。
【0045】
本発明を上記態様に関して説明してきたが、上記説明および下記の実施例は本発明を例示するためのものであり、本発明の範囲を限定するためのものではないことを理解すべきである。本発明の範囲内における他の態様、利点および変更は、本発明が関係する当業者には明らかになるであろう。
【図面の簡単な説明】
【0046】
【図1A】図1A〜1H:ネトリン−1は、恐らくはUNC5Bによって引き起こされるアポトーシスを阻害することにより内皮細胞アポトーシスを防止する。 図1A:ヒト臍帯静脈/動脈内皮細胞(HUVEC/HUAEC)はUNC5AおよびUNC5B依存性受容体を発現するが、ネトリン−1を発現しない。定量RT−PCRは、実験手順に記載の通りに行った。
【図1B】図1B:ネトリン−1投与は、トリパンブルー排除によって測定される血清飢餓時に見られるHUVEC/HUAEC細胞死を阻害する。少なくとも100個の細胞/条件を計数した。相対指数は、平均値+/−s.e.m(n=3)として示す。*:p値<0.005;**:p値<0.0001(Student t検定)。
【図1C】図1C:ネトリン−1は、TUNEL染色によって測定したHUVECにおけるアポトーシスを防止する。TUNEL陽性細胞の定量は、平均値+/−s.e.mである(Student t検定)。
【図1D】図1D:ネトリン−1投与により、HUVECおよびHUAECにおけるカスパーゼ−3活性が減少する。カスパーゼ−3活性の相対指数は、平均値+/−s.e.m(n=3)として表される。開裂したカスパーゼ−3陽性細胞の免疫染色および相対指数(平均値+/−s.e.m)が示される(Student t検定)。
【図1E】図1E:ネトリン−1投与により、HUVECおよびHUAECにおけるカスパーゼ−3活性が減少する。カスパーゼ−3活性の相対指数は、平均値+/−s.e.m(n=3)として表される。開裂したカスパーゼ−3陽性細胞の免疫染色および相対指数(平均値+/−s.e.m)が示される(Student t検定)。
【図1F】図1F:siRNAによる、UNC5Aではなく、UNC5Bサイレンシングは、カスパーゼ−3活性の減少と関連している。値は、平均値およびs.e.m(n=3)である。総て、p<0.05(コントロール条件におけるレベルと比較したStudent t検定)。
【図1G】図1G:DAPキナーゼは、HUVECおよびHUAECにおけるUNC5Bによって引き起こされるアポトーシスを介在する。DAPキナーゼΔCaM(DAPキナーゼの構成性の活性突然変異体型)およびDAPキナーゼDD(DAPキナーゼのドミナントネガティブ型)の強制発現後の内皮細胞における細胞死(Toxilight)。値は平均値およびs.e.m.である。総て、p<0.001(コントロール条件におけるレベルと比較したStudent t検定)。
【図1H】図1H:ネトリン−1投与により、DAPキナーゼリン酸化が誘発される。抗ホスホ−DAPキナーゼを用いて、酵素活性のレベルを測定した。
【図2A】図2A〜2F:カスパーゼ阻害剤およびUNC5Bサイレンシングは、新生血管形成におけるネトリン−1効果に似ている。 図2A:ネトリン−1またはカスパーゼ阻害剤z−VAD−fmkおよびBAFは、エクスビボでのネズミ大動脈リングマトリゲルアッセイにおける微小血管形成を促進する。少なくとも8種類のマウスから切除された大動脈リングを、条件毎に示す。微小血管全長(ピクセル単位)の定量は、実験手順に記載の通りに行った。総てのp値を示す(χ2検定)。
【図2B】図2B:ネトリン−1またはカスパーゼ阻害剤z−VAD−fmkおよびBAFは、エクスビボでのネズミ大動脈リングマトリゲルアッセイにおける微小血管形成を促進する。少なくとも8種類のマウスから切除された大動脈リングを、条件毎に示す。微小血管全長(ピクセル単位)の定量は、実験手順に記載の通りに行った。総てのp値を示す(χ2検定)。
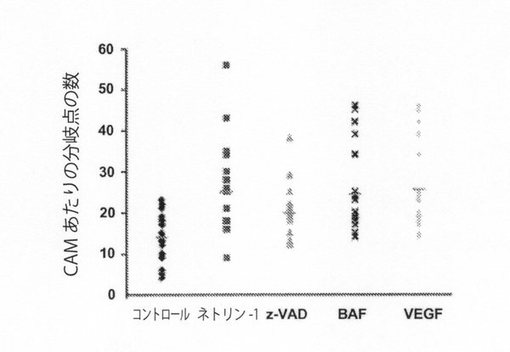
【図2C】図2C:ネトリン−1およびカスパーゼ阻害剤は、CAMアッセイにおける新生血管形成を誘発する。CAMモデルは、材料および方法に記載の通りに処理した8日齢のヒナドリ胚(n≧10)を用いて調製した。メチルセルロースディスクに輪郭を描く。デノボ(de novo)血管形成は、分岐点の数として表す(n≧13フィールド/条件)。平均値は、直線によって示す。*:p値<0.02;**:p値<0.0005(Mann−Whitney U検定)。
【図2D】図2D:ネトリン−1およびカスパーゼ阻害剤は、CAMアッセイにおける新生血管形成を誘発する。CAMモデルは、材料および方法に記載の通りに処理した8日齢のヒナドリ胚(n≧10)を用いて調製した。メチルセルロースディスクに輪郭を描く。デノボ血管形成は、分岐点の数として表す(n≧13フィールド/条件)。平均値は、直線によって示す。*:p値<0.02;**:p値<0.0005(Mann−Whitney U検定)。
【図2E】図2E:siRNAによるUNC5Bサイレンシングは、CAMアッセイにおける新生血管形成を増加する。実験手順に記載の通りに3回のsiRNA注射をE.5とE.9との間に行った。スクランブルsiRNAを、コントロールとして用いた。デノボ血管形成は、分岐点の数として表す(n>30異なるフィールド/条件)。平均値は、直線によって示す。P値=0.009(Mann−Whitney U検定)。
【図2F】図2F:siRNAによるUNC5Bサイレンシングは、CAMアッセイにおける新生血管形成を増加する。実験手順に記載の通りに3回のsiRNA注射をE.5とE.9との間に行った。スクランブルsiRNAを、コントロールとして用いた。デノボ血管形成は、分岐点の数として表す(n>30異なるフィールド/条件)。平均値は、直線によって示す。P値=0.009(Mann−Whitney U検定)。
【図3A】図3A〜3C:ノックダウンネトリン−1aゼブラフィッシュ胚の表現型は、アポトーシスの阻害によって救済される。 fli:egfpゼブラフィッシュ胚を、1〜4細胞期でコントロール(ctl)、スプライス(spl)または翻訳(tsl)遮断ネトリン1a、UNC5BおよびDAPキナーゼモルホリノと共に単独でまたは組み合わせて投与した。表現型を、24〜54hpfに分析した。主要血管構造の典型的なイメージを示す。左が先。カスパーゼ阻害剤BAFは、ネトリン−1a splモルホリノを投与したゼブラフィッシュ胚では傍脊索血管形成(星印)を回復し(図3A)、ネトリン−1a tslモルホリノを投与したゼブラフィッシュ胚ではISV形成を回復する(図3B)。同様の表現型の救済は、ゼブラフィッシュ胚におけるUNC5BまたはDAPキナーゼおよびネトリン1a tsl遮断モルホリノの同時投与によって観察された(図3B)。 図3A:定量を示す。p値を示す(χ2検定)。
【図3B】図3B:定量を示す。p値を示す(χ2検定)。
【図3C】図3C:BAF処理またはUNC5B遮断モルホリノ投与は、TUNEL染色(左パネル)またはカスパーゼ−3活性(右パネル)によって示されるようにゼブラフィッシュ胚におけるネトリン1a tsl遮断モルホリノ投与後に観察されるアポトーシスの増加を阻害する。カスパーゼ−3活性は、ネトリン−1a tslモルホリノのみを投与した胚に関する相対指数として表される。平均値およびs.e.m.は、3回の独立した実験から計算される。p値を示す(Student t検定)。
【図4A】図4A〜4G:ネトリン−1は、UNC5Bによって引き起こされる内皮細胞アポトーシスを防止する。 図4A〜4B:HUVECおよびHUAEC細胞におけるネトリン−1発現の評価。 図4A:特異的ネトリン−1抗体を用いる免疫蛍光を用いた。ネトリン−1標識は、MRC−5肺繊維芽細胞系とは反対に、HUVECおよびHUAEC細胞では検出されなかった。
【図4B】図4B:培地に分泌されたネトリン−1の定量は、Apotech Corporation製のネトリン−1(ヒト)ELISAキットを用いて行った。
【図4C】図4C:ネトリン−1処理により、HUVECおよびHUAECにおけるカスパーゼ−3活性は用量依存的に減少する。カスパーゼ−3活性の相対指数は、平均値+/−s.e.mと表される(n=3)。*:p<0.05;**p<0.0025(コントロール条件におけるレベルと比較したStudent t検定)。
【図4D】図4D:ネトリン−1処理により、50μMエトポシドで一晩処理したときに見られたHUVEC/HUAEC細胞死が阻害される。細胞死は、実験手順に記載のToxilightアッセイを用いて評価する。相対指数は、平均値およびs.e.mである(n=3)(p<0.05、Student t検定)。
【図4E】図4E:ラットUNC5H2の細胞外ドメインに対応する組換えタンパク質の添加により、ネトリン−1によって介在される細胞死阻害が防止される。結果は、それぞれの場合にコントロールに対する相対指数として表される(平均値+/−s.e.m,n=3、t検定)。
【図4F】図4F:スクランブルsiRNAでトランスフェクションしたコントロール細胞と比較したUNC5BsiRNAでトランスフェクションした細胞におけるUNC5B発現の定量RT−PCR分析。トランスフェクションの効率を示す、siRNAでトランスフェクションした細胞におけるGFP発現の典型的画像。
【図4G】図4G:スクランブルsiRNAでトランスフェクションしたコントロール細胞と比較したUNC5AsiRNAでトランスフェクションした細胞におけるUNC5A発現の定量RT−PCR分析。
【図5A】図5A〜5C:エクスビボセッティングにおけるUNC5Bの発現。 図5A:大動脈リングでのUNC5B発現の定量RT−PCR分析。
【図5B】図5B:CAMでの定量RT−PCRによるネトリン−1、UNC5H2(マウスUNC5B)およびDAPキナーゼ発現の分析。ネトリン−1とは反対に、UNC5H2およびその下流エフェクターが発現する。
【図5C】図5C:CAMでのUNC5B発現の定量RT−PCR分析、およびスクランブルsiRNAを用いてトランスフェクションしたコントロールと比較したCAMでのUNC5B発現に対する血液循環に投与したUNC5BsiRNAの効果。
【図6A】図6A〜6D:ゼブラフィッシュ胚におけるネトリン−1aおよびUNC5Bモルホリノの効果。 図6A:ネトリン−1bは、ゼブラフィッシュにおける発生上の血管形成に関与しているとは思われない。翻訳遮断ネトリン−1bモルホリノは、発生上の血管形成に対して重大な欠損を持っていない。欠損のない胚の割合および分析した胚の総数を示す。ネトリン−1オーソログ(otholog)遺伝子の系統発生的研究。100ブートストラップ複製を用いるJTTモデルでのタンパク質配列に関するML。ゼブラフィッシュにおけるネトリン−1オーソログであるネトリン−1aおよびネトリン−1bには、下線を付けている。
【図6B】図6B:ゼブラフィッシュ胚におけるネトリン−1aおよびUNC5B発現のRT−PCR分析。
【図6C】図6C:第三のネトリン−1a遮断モルホリノは、図3A〜3Cに記載のものと同様な血管の欠損を生じる。ISV欠損を有する胚の割合および分析した胚の総数を示す。
【図6D】図6D:ネトリン−1aサイレンシングは、全ゼブラフィッシュ胚のカスパーゼ−3活性を増加させる。カスパーゼ−3活性は、コントロールMOを投与した胚に関する相対指数として表される。平均値およびs.e.m.は、3回の独立した実験から計算される。
【図7】DCC−5Fbnデコイペプチドは、内皮細胞に対するネトリン生存効果を逆転させる。HUVECにおけるネトリン−1によるカスパーゼ−3活性化の減少は、DCCの第五の細胞外免疫グロブリンドメインに対応するデコイペプチドDCC−5Fbnによる細胞の同時処理によって減少する。カスパーゼ−3活性の相対的減少割合は、平均値+/−s.e.mとして表される(n=3)。Student t検定:p<0.015。
【0047】
材料および方法
A)内皮細胞培養およびトランスフェクション
内皮細胞培養およびトランスフェクション:
ヒト臍帯静脈および動脈細胞(Promocell,継代数<8)を、2%血清(キット、Promocell)を補足した内皮細胞成長培地(ECGM,Promocell)で80%コンフルエンスに達するまで培養した。HUVECを、AMAXA Nucleofectorシステムを用いて製造業者の指示に従ってトランスフェクションした。簡単に説明すれば、500,000個の細胞を100μlのHUVEC Nucleofector溶液に再懸濁し、1μgsiRNAまたは1.5μgプラスミドDNAおよび0.5μgGFPコードコントロールプラスミドを用いてトランスフェクションした。UNC5A、UNC5BおよびスクランブルsiRNAは、Sigma−Proligoによってデザインされた。突然変異体DAPキナーゼ(DAPキナーゼΔCamおよびDAPキナーゼDD)をコードするプラスミドは、以前に記載された(Llambi et al.,2005)。トランスフェクション後、細胞を1%成長培地で培養し、24時間後に回収した。
【0048】
B)細胞死アッセイ
細胞死アッセイは、上記に記載の通りに行った。簡単に説明すれば、内皮細胞を、10,000個/cm2を完全培地に16時間播種した。細胞死は、無血清培地(内皮細胞基礎培地、Promocell)での飢餓によりまたは50μMエトポシドに暴露することにより誘発した。ネトリン−1は、Axxora−Apotechから購入した。細胞死は、上記の通りにトリパンブルー排除アッセイによって評価した。DNA断片化を検出するため、カバーグラス上で培養した処理済み細胞を4%パラホルムアルデヒド(PFA)に20分間固定し、ターミナルデオキシヌクレオチジルトランスフェラーゼ介在dUTP−ビオチンニック末端標識(TUNEL)を、300U/mlTUNEL酵素(300U/ml)および6μMビオチニル化dUTP(Roche Diagnostics)で行った。カスパーゼ−3活性を、Clontech(米国)製ApoAlert CPP32キットを用いて、文献記載の抗活性カスパーゼ−3抗体(Cell signaling)を用いる固定細胞での免疫染色によって測定した(Llambi et al.,2005; Tauszig-Delamasure et al.,2007)。Toxilightアッセイは、製造業者の指示に従って行った(Lonza)。ネトリン−1滴定実験のため、ラット組換えUNC5H2−Fc(R&D)を1μg/mlで用いた。
【0049】
C)ウェスタンブロット分析および免疫蛍光
10,000個/cm2で播種し、無血清培地(内皮細胞基礎培地,Promocell)中でネトリン−1と共に24時間処理したまたは処理しなかった内皮細胞を回収して、プロテアーゼ阻害剤の存在下にて50mMHEPES pH7.6、125mMNaCl、5mMEDTAおよび0.1%NP−40緩衝液中で溶解した。タンパク質抽出物を、抗DAP−キナーゼおよび抗ホスホ−DAPキナーゼ抗体(Sigma)で免疫ブロットした。免疫蛍光分析のため、細胞をLabTekチャンバースライド上に播種し、4%PFAで15分間固定した。抗ネトリン−1ラットモノクローナル抗体(R&D)を1/150の希釈倍率で用い、特異的二次抗体(Alexa488 Donkey anti-Rat,Molecular Probes-Invitrogen)を用いて可視化した。
【0050】
D)定量RT−PCR
HUVEC/HUAEC、大動脈リング、CAMおよびゼブラフィッシュ胚におけるネトリン−1、依存性受容体、DCCおよびUNC5A−D、およびDAPキナーゼ発現を評価するため、総RNAをNucleospin RNAIIキット(Macherey-Nagel)を用いて抽出し、1μgをiScript cDNA Synthesisキット(BioRad)を用いて逆転写した。リアルタイム定量RT−PCRを、LightCyclerFastStartDNAMasterSYBERGreenIキット(Roche)を用いてLightCycler2.0装置(Roche)上で行った。総ての最適増幅のための反応条件並びにプライマー選択は、上記の通りに決定した。プライマーの配列は、必要であれば入手可能である。
【0051】
E)大動脈リングアッセイ
大動脈リングアッセイは、Nicosiaと共同研究者らによって記載されている方法によって行った(Nicosia and Ottinetti,1990)。大動脈は6〜8週齢のBalb/Cマウスから切除し、外膜周囲の脂肪および結合組織を除去し、ECGMで2回洗浄し、1〜1.5mmの長さのリングに切断した。成長因子減少マトリゲル(Growth factor reduced Matrigel; BD Biosciences)100μlを、48穴組織培養プレート(Costar)のそれぞれのウェルに添加し、37°Cで30分間固化させた。大動脈リングをマトリックス層に置き、Matrigel100μlに埋設し、完全成長培地(ECGM,Promocell)で24時間回復させた。次いで、それらをネトリン−1(150ng/ml)、z−VAD−fmk(Tebu;20nM)、BAF(Sigma;20nM)またはVEGF(Sigma;13ng/ml)で処理しまたは処理せずに、4日目に位相差顕微鏡で写真を撮った。微小血管生長の全長は、AxioVisionRelease4.6ソフトウェアを用いて計算した。
【0052】
F)CAMアッセイ
ヒナドリ漿尿膜(CAM)アッセイは、上記に記載の通りに行った。白色レグホンの卵を、加湿雰囲気下にて37°Cでインキュベーションした。72時間インキュベーションした後、3mlの卵白を除去し、四角い窓を殻に開けた。開口部は、8日目までセロテープで閉じた。次いで、ネトリン−1(150ng/ml)、z−VAD−fmk(Tebu;20nM)、BAF(Sigma;20nM)またはVEGF(Sigma)をPBSで希釈し、等容のオートクレーブ処理した1%メチルセルロース溶液と混合した。コントロールは、ビヒクルのみで調製した。それぞれの試料溶液(10μl)をパラフィルムに滴下して乾燥した。メチルセルロースディスクをパラフィルムから外して、8日齢ヒナドリ胚のCAM上に置いた。3日後、CAMを撮影して、血管形成応答をAxioVisionRelease4.6ソフトウェアを用いてメチルセルロースディスク下で二次異所性血管芽の数を測定することによって評価した。CAMにおけるUNC5Bサイレンシングは、E.5、E.7およびE.9でのCAM血液循環中でSigma−Proligoによってデザインされた特異的siRNAを投与することによって行った。分析は、E.11で行った。
【0053】
G)ゼブラフィッシュ胚のモルホリノノックダウン
トランスジェニックfli:egfpゼブラフィッシュをZebrafish International Resource Center(オレゴン大学)から入手し、28°Cで標準的飼育条件下に保持した(Lawson and Weinstein,2002)。胚をプロナーゼ酵素処理を用いてコリオン除去し、6ngのスプライス/翻訳遮断ネトリン−1、UNC5Bおよび予めデザインして特性決定したコントロールモルホリノを、単独でまたは組み合わせて1〜4細胞期の胚の高卵黄に投与した。第三のスプライス遮断モルホリノ特異ターゲッティングネトリン−1aをデザインし(GeneTools)、同じ用量で胚に投与した。最後に、DAPキナーゼ(GeneTools)の翻訳遮断モルホリノターゲッティングゼブラフィッシュオーソログ3ngを投与し、コントロールと比較してノックダウン胚におけるDAPキナーゼmRNA発現を35%減少させた。モルホリノの配列は、必要であれば入手可能である。胚を0.003%フェニルチオ尿素を補足したE3培地で培養し、色素形成を阻害した。ネトリン−1aサイレンシングによって誘発される表現型の救済を、モルホリノ投与の4時間後にE3培地に40nMBAFを加えることによって試験した。TUNEL染色のため、胚を4%PFA中で48hpfで4°Cにて一晩固定した。次いで、それらをPBS−Tween0.1%(PBST)で10分間洗浄し、連続的メタノール槽(25、50、75および100%)で累進的に脱水した。再水和(75、50および25%メタノール系)の後、胚をPBSTで洗浄し、PBS中で10μg/mlプロテイナーゼK(Roche)で5分間処理し、PBSTで再度洗浄し、PBS−BSA1%中で30分間インキュベーションした。TUNEL標識は、予め上記したとおりに行い、Cy3結合−ストレプトアビジン(1/400;Jackson Immunoresearch)を用いて可視化した。カスパーゼ−3活性アッセイは、24時間発生時に胚全体について上記の通りに行った。微速度撮影実験のため、ゼブラフィッシュ胚を0.016%トリカインを含む3%メチルセルロース溶液に埋設し、運動を抑制した(Isogai et al.,Development and Disease,2003)。透過ビデオ顕微鏡画像化を、Timelapse Axiovert100M(PLATIM,ENS Lyon)を用いて行った。
【0054】
DCC−5Fbn産生:DCC−5Fbn産生は、標準的手順を用いて行うことができる。簡単に説明すれば、BL21細胞をイミダゾールに応じてDCC−5Fbnを強制的に発現させ、BL21溶解物をFlag−アガロース(Sigma)を用いてアフィニティークロマトグラフィーにかけた。
【実施例】
【0055】
本発明者らは、本明細書において、これらの2つの明らかに対立する観察を、ネトリン−1の未結合UNC5B受容体によって誘発されるアポトーシスを遮断することにより、血管形成中に内皮細胞の生存を促進するネトリン−1の能力を説明することができるかどうかを検討した。実際に、ネトリン−1は、いわゆる依存性受容体DCCおよびUNC5A−Dについてのリガンドとして作用することによって上皮細胞死を予防し、それらのリガンドの存在下および非存在下のいずれにおいても活性な機能特性を共有しており、リガンド結合は正のシグナル伝達を誘発するが、依存性受容体は別のシグナル伝達カスケードを誘発し、それらの栄養リガンドから解放されたときに細胞死を生じる(Llambi et al.,2001; Mehlen et al.,1998; Tanikawa et al.,2003)(Bredesen et al.,2005; Mehlen and Bredesen,2004)。このスキームでは、UNC5Bの喪失、すなわちアポトーシス促進シグナル伝達の喪失は、実際に血管数の増加と関連している(Lu et al.,2004)。一方、ネトリン−1の喪失、すなわちアポトーシス促進シグナル伝達の増加は、血管消失と関係していると予想される(Wilson et al.,2006)。このような内皮細胞生存とアポトーシスとの間の動的バランスは、血管の完全性の保持を助長する可能性があり、これはまた、以前に報告されているように血管発生および病理学的血管形成の際の血管ホメオスタシスにも重要である(Alavi et al.,2003; Carmeliet,2005; Duval et al.,2003; Santoro et al.,2007; Winn et al.,2005)。
【0056】
例1:ネトリン−1はその未結合UNC5B受容体によって誘発される内皮細胞アポトーシスを遮断する
ネトリン−1がその未結合UNCB受容体によって誘発されるアポトーシスを遮断するかどうかを決定するため、どのネトリン−1依存性受容体、すなわちDCC、UNC5A、UNC5B、UNC5CおよびUNC5Dが、ヒト臍帯静脈/動脈内皮細胞(HUVEC/HUAEC)で発現するかを最初に決定した。定量RT−PCRによって示されているように、UNC5AおよびUNC5Bは、いずれもHUVECおよびHUAECで発現するが(図1A)、UNC5C、UNC5DおよびDCCを検出することはできなかった(データは示さず)。ネトリン−1は、これらの細胞では、RNA(図1A)またはタンパク質レベル(図4A〜4B)でも発現しなかった。次いで、ネトリン−1が内皮細胞の生存因子として働くかどうかを、HUVECおよびHUAECが血清飢餓下で経験する細胞死の自発的傾向を遮断するその能力を分析することによって検討した。ネトリンは、50ng/mlの用量が、生理学的かつ機能的であることが以前に報告されているので、総てのインビトロ実験でこの用量を用いた((Serafini et al.,1996; Serafini et al.,1994)および図4C)。図1Bに示されているように、ネトリン−1は、HUVECおよびHUAEC両方の細胞死を阻害した(p<0.005)。同様な結果は、エトポシドによって引き起こされる細胞死についても得られた(図4D)。ネトリン−1は、TUNEL陽性細胞数の減少(図1C)およびカスパーゼ−3活性の減少(図1D〜1E)によって示されるように、内皮細胞のアポトーシスを防止した(HUVECについては30%までおよびHUAECについては40%まで;総てp<0.005)。UNC5H2の細胞外ドメインに対応する組換えタンパク質の添加により、ネトリン−1によって介在される細胞死の阻害が防止され、生存効果はネトリン−1に特異的であるという結論を支持していた(図4E)。従って、ネトリン−1は、内皮細胞に対して生存因子として働く。
【0057】
UNC5Bおよび/またはUNC5Aが内皮細胞のアポトーシスを誘発するかどうかを決定するため、siRNA法を用いてそれらの発現を一過的にサイレンシングした(図1Fおよび図4F〜4G)。UNC5Bサイレンシングにより、カスパーゼ−3活性は減少したが、UNC5A siRNAはカスパーゼ−3活性に対して全く効果を示さなかった(図1F;総てp<0.05)。更に、UNC5Bサイレンシングと一緒のネトリン−1の添加は、追加的生存効果と関連しなかった(図1F)。従って、内皮細胞に対するネトリン−1生存効果は、UNC5B発現、および別の状況では未結合UNC5Bによって誘発されるアポトーシスの阻害からの同様な結果によって変化する。UNC5Bによって引き起こされるアポトーシスはDAPキナーゼによって介在されることが示されているので、最初にHUVECおよびHUAEC細胞で構成性の活性DAPキナーゼDAPKΔCaMを発現させた(Shohat et al.,2001)。図1Gに示されるように、DAPキナーゼの構成性の活性形態の発現により、細胞死が引き起こされた(総てp<0.0001)。次に、DAPキナーゼのドミナントネガティブ突然変異体DAPK DDを強制発現させた(Llambi et al.,2005)。ネトリン−1処理と同様に、DAPK DDは細胞死誘発を防止した。更に、UNC5Bアポトーシス促進シグナル伝達は、DAPキナーゼ自動リン酸化の阻害を介するDAPキナーゼの活性化によって介在されるので(Llambi et al.,2005)、次にDAPキナーゼリン酸化レベルを測定した。図1Hに示されるように、ネトリン−1処理により、恐らくはUNC5Bアポトーシス促進シグナル伝達カスケードの遮断によってHUVECにおけるDAPキナーゼリン酸化が増加した。UNC5B突然変異体胚における血管数の増加が見られたことと一緒に(Lu et al.,2004)、これらのインビトロデータは、ネトリン−1が血管形成の際にUNC5Bによって引き起こされるアポトーシスを遮断することができるという考察を支持している。
【0058】
例2:ネトリン−1の血管形成促進効果は、カスパーゼ阻害およびUNC5Bサイレンシングに似ている
次に、ネトリン−1の血管形成促進活性が、細胞生存を促進するその能力に関するという概念を、新血管形成のエクスビボモデルを用いて検討した。最初に、マウス大動脈リングアッセイを用いて、大動脈外植片におけるUNC5BのネズミオーソログであるUNC5H2の発現を評価した(図5A)。ネトリン−1は、外植片において発現されることは見られなかった(データは示さず)。VEGFで観察されたのと同様に、ネトリン−1処理では、コントロール条件と比較してマトリゲルに埋設した大動脈リングから毛細血管の出芽が増大し(図2A〜2B;p<0.001)、ネトリン−1の血管形成促進活性を支持していた。興味深いことには、カスパーゼ阻害薬z−VAD−fmkおよびBAFも新毛細血管形成を誘発し、ネトリン−1の効果に似ていた(図2A〜2B;p<0.002)。同じ結果は、ヒナドリ漿尿膜(CAM)アッセイで得られた(図2C〜2D、および発現分析についての図5B)。実際に、VEGF、ネトリン−1、z−VAD−fmkまたはBAFで浸漬したメチルセルロースディスク付近では、新血管形成の有意な増加が観察された(図2C〜2D;p<0.02)。従って、これらのエクスビボ生理学的モデルでは、アポトーシスの阻害はネトリン−1の効果に十分似ていると思われる。更に、CAMでのsiRNAによるUNC5Bサイレンシングにより新血管分枝が増加し(図5C)、ネトリン−1およびその受容体UNC5Bは、血管形成過程中に拮抗的に作用する(図2E〜2F;p=0.009)。
【0059】
例3:ゼブラフィッシュ発生中のネトリン−1aサイレンシングは、カスパーゼ阻害およびUNC5B/DAPキナーゼサイレンシングによって救済される血管欠損を生じる
ネトリン−1の血管形成促進および抗アポトーシス活性の間の原因と結果の関係をインビボで更に調査するため、抗アポトーシス処理がゼブラフィッシュ発生中にネトリン−1崩壊によって誘発される血管欠損を救済することができるかどうかを検討した。2種類のネトリン−1遺伝子オーソログであるネトリン−1aおよびネトリン−1bは、ゼブラフィッシュで特性決定されている(Lauderdale et al.,1997; Strahle et al.,1997)。ネトリン−1bサイレンシングは血管発生に対して何ら明らかな効果を持たないが(図6A)、EichmanのグループおよびLiのグループはそれぞれ異なるネトリン−1aモルホリノを用いて、ネトリン−1aのサイレンシングが血管発生欠損と関連していることを示した(Lu et al.,2004; Wilson et al.,2006)。スプライス遮断モルホリノ(spl−ネトリン−1a)を用いるネトリン−1aサイレンシングは、傍脊索血管(parachordal vessels:PAV)の形成を防止することが示されている(Wilson et al.,2006)。fli:egfpトランスジェニックゼブラフィッシュ、血管において特異的にGFPの発現をターゲッティングするトランスジェニックモデル(Lawson and Weinstein,2002)を用いて、受精から50〜54時間後(hpf)に、このようなspl−ネトリン−1aモルホリノを投与した胚の57.3%がPAVを欠いており(n=75)、節間血管(intersegmental vessels:ISV)および背側縦吻合血管(dorsal longitudinal anastomotic vessels:DLAV)の発生は正常である(図3A)ことを観察した。spl−ネトリン−1aを投与した胚を汎用カスパーゼ阻害剤BAFで処理すると、PAV形成は、未処理胚での42.7%と比較して処理済み胚の92.6%(n=54)で見られたので、この表現型を良好に逆転した(図3A;p<0.0001)。以前に報告されているように(Wilson et al.,2006)、翻訳遮断モルホリノ(tsl−ネトリン−1a、図6B)並びに第三のネトリン−1a遮断モルホリノ(図6C)を用いるネトリン−1aサイレンシングにより、ISVの組織崩壊、背側吻合血管(DLAV)の非存在(図3B)および発生上の欠損を生じた。ISV発生の微速度撮影分析により、ISVの非存在または未熟成長停止(premature growth arrest)を確かめた(例4参照)。同様の欠損は、アポトーシスタンパク質スルビビンの小阻害剤についてノックダウンしたゼブラフィッシュ胚に以前に報告されていた(Ma et al.,2007)。更に、カスパーゼ−3活性の増加が、ネトリン−1aノックダウン胚で観察された(図6D)。これらの胚のBAFによる処理は、コントロールでの23.0%と比較して処理済み胚の54.5%が少なくとも部分的ISVを示したので(図3B;p<0.001)、ISV形成の救済に十分であった。依存性受容体概念に従って、UNC5Bモルホリノの同時投与(図6B)は、胚の79.6%(n=54)においてtsl−ネトリン−1aモルホリノ投与によって誘発される欠損を修正することもできた(図3B;p<0.0001)。同様の救済は、tsl−ネトリン−1a遮断モルホリノとモルホリノターゲッティングDAPキナーゼの共投与によって得られた(図3B;p<0.0001)。
【0060】
BAF処理またはUNC5Bモルホリノ投与は、TUNEL免疫染色アッセイおよびカスパーゼ−3活性アッセイ(図3C)によって測定されるように、ネトリン−1aサイレンシング後の胚で観察される細胞死の増加を救済し、ネトリン−1aによって阻害された内皮細胞死は、血管発生中にUNC5Bによって特異的に介在されるという考えを更に支持する。
【0061】
例4:fli−EGFPゼブラフィッシュ胚における主要血管発生に対するネトリン−1aモルホリノ効果の微速度撮影分析
fli−EGFPトランスジェニックゼブラフィッシュにおける血管動力学の微速度撮影画像を得ることができる。従って、節間血管(ISV)の最初の出現は、3分間の時点間隔で収集した時点によって示すことができる。
【0062】
これらの微速度撮影画像(データは示さず)は、ISVおよび背側縦吻合血管(DLAV)形成は、通常はコントロールモルホリノを投与したfli−EGFPゼブラフィッシュ胚で起こることを示している。翻訳遮断ネトリン−1aモルホリノを投与したfli−EGFPゼブラフィッシュ胚では、ISVは未熟成長停止(premature growth arrest)を形成または提示することができず、DLAVは存在しない。
【0063】
本発明者らは、本明細書において、ネトリン−1が内皮細胞の生存を制御し、かつ、その未結合UNC5B受容体によって誘発されるアポトーシスを遮断することによって少なくとも部分的に血管形成を促進することを示した。この機能の調節緩和(deregulation)は、本発明者らおよび第三者が観察した血管形成の調節を説明することができる(Lu et al.,2004; Wilson et al.,2006)。このことは、発生および病理学的血管形成中の細胞死調節の重要性を記載している証拠の成長線と一致する(Birdsey et al.,2008; O'Connor et al.,2000; Santoro et al.,2007)。
【0064】
ネトリン−1依存性受容体は、別の状況では栄養リガンドの利用できないことのセッティングにおいて発生する上皮細胞のアポトーシスを誘発することによって腫瘍サプレッサーとして作用することが最近提案された(Grady,2007; Mehlen and Puisieux,2006)。従って、その依存性受容体のアポトーシス促進活性の喪失は、結直腸癌におけるDCCおよびUNC5C発現の頻繁な喪失によって例示されるように、腫瘍細胞についての選択的利点を表している(Bernet et al.,2007; Mazelin et al.,2004)。相補的に、ネトリン−1発現の利得は、腫瘍細胞についての同様な選択的利点を表しており、この現象は、転移性乳癌や肺癌などの幾つかの種類の侵攻性癌で実際に観察される(Delloye et al.,2008; Fitamant et al.,2008)。本明細書に示された結果を考慮して、本発明者らは、腫瘍上皮細胞における発現のこのオートクラインネトリン−1利得が2つの付加的効果を有することがあることを提案する。第一には、報告されているように、依存性受容体によって引き起こされる細胞死を阻害することによって上皮腫瘍細胞についての選択的利点を付与する(Fitamant et al.,2008)。第二には、それは、潜在的に血管保持および/または発生にも有利であり、従って癌の進行を促進する可能性がある。従って、ネトリン−1受容体との相互作用の滴定または阻害によるネトリン−1の機能の崩壊に基づく抗癌法は、腫瘍上皮細胞だけでなく腫瘍血管形成血管をも根絶するものであり、従って癌における決定的なターゲットとしてのネトリン−1の位置を更に強化する。
【0065】
例5:DCC−5Fbnデコイペプチドは、内皮細胞に対するネトリン−1生存効果を防止する
DCCの第五の免疫グロブリンドメインに対応するデコイペプチドDCC−5Fbnでの処理は、ネトリン−1受容体多量体化を阻害することによってネトリン−1生存効果を逆転することが以前に示された(Mille et al.,2009)。
【0066】
このようなペプチドが内皮細胞に対するネトリン−1効果を遮断できるかどうかを決定するため、内皮細胞をネトリン−1のみまたはネトリン−1およびDCC−5Fbnで処理したときのカスパーゼ−3活性の相対的減少を比較した。DCC−5Fbnを添加すると、カスパーゼ−3活性に対するネトリン−1効果は1/2に減少し、従って内皮細胞に対するその生存効果を強力に減少する(図7,p<0.015)。
【0067】
文献
Alavi et al.(2003). Science 301,94-96.
Bernet et al.(2007). Gastroenterology 133,1840-1848.
Birdsey et al.(2008). Blood 111,3498.
Bredesen et al.(2005). Cell Death Differ 12,1031-1043.
Carmeliet,P.(2005). Nature 438,932-936.
Cirulli et al.(2007). Nat Rev Mol Cell Biol 8,296-306.
Delloye et al.(2008). J Natl Cancer Ins in press.
Duval et al.(2003). Angiogenesis 6,171-183.
Fitamant et al.(2008). Proc Natl Acad Sci USA,105,4850-4855.
Grady,W.M.(2007). Gastroenterology 133,2045-2049.
Lauderdale et al.(1997). Mol Cell Neurosci 9,293-313.
Lawson et al.(2002). Dev Biol 248,307-318.
Liu et al.(2004). Curr Biol 14,897-905.
Llambi et al.(2001). Embo J 20,2715-2722.
Llambi et al.(2005). Embo J 24,1192-1201.
Lu et al.(2004). Nature 432,179-186.
Ma et al.(2007). BMC Dev Biol 7,50.
Mazelin et al.(2004). Nature 431,80-84.
Mehlen et al.(2004). Apoptosis 9,37-49.
Mehlen et al.(2006). Nat Rev Cancer 6,449-458.
Mehlen et al.(1998). Nature 395,801-804.
Mille et al.(2009). Cell Death Differ. 16(10):1344-51.
Nicosia et al.(1990). Lab Invest 63,115-122.
O'Connor et al.(2000). Am J Pathol 156,393-398.
Santoro et al.(2007). Nat Genet 39,1397-1402.
Serafini et al.(1996). Cell 87,1001-1014.
Serafini et al.(1994). Cell 78,409-424.
Shohat et al.(2001). J Biol Chem 276,47460-47467.
Strahle et al.(1997). Mech Dev 62,147-160.
Tanikawa et al.(2003). Nat Cell Biol 5,216-223.
Tauszig-Delamasure et al.(2007). Proc Natl Acad Sci USA,104,13361-13366.
Wilson et al.(2006). Science 313,640.
Winn et al.(2005). J Thromb Haemost 3,1815-1824.
【技術分野】
【0001】
本発明は、内皮細胞、好ましくは血管または新生血管由来の内皮細胞の死を誘発することができる化合物を選択するためのインビトロ(in vitro)の方法に関する。本発明は更に、内皮細胞、好ましくはネトリン−1を発現する腫瘍の血管または新生血管由来の内皮細胞の死を誘発することができる化合物としてのネトリン−1の機能阻害剤の使用も含んでなる。最後に、本発明は、内皮細胞の死を誘発することができる化合物を選択するためのキットに関する。
【背景技術】
【0002】
軸索誘導分子として最初に報告されたネトリン−1は、分枝器官の形態発生における役割から血管形成における機能までの範囲の神経外過程においても機能することが最近示されている(Cirulli and Yebra, 2007; Liu et al., 2004; Lu et al., 2004; Wilson et al., 2006)。興味深いことには、血管形成中のネトリン−1の役割に関して矛盾する結果が報告された。しかしながら、以下の2つの明らかな発生学的結果は、最初はちぐはぐであると思われた:Eichmann、Tessier−Lavigneと共同研究者らは、マウスにおけるUNC5Bの発生学的不活性化は血管形成の増加と関係しており、それによってネトリン−1の抗血管形成活性が示唆されることをあざやかに立証したが、Liと共同研究者らは、UNC5Bのリガンドをコードするネトリン−1aの不活性化は、ゼブラフィッシュ発生中の血管の喪失と関係していることを示した(Lu et al., 2004; Wilson et al., 2006)。
【0003】
血管形成は、多数の様々な疾病状態の開始および/または拡散における重要な要因であるので、信頼性のあるやり方で血管形成を制御しまたは阻害することができる治療組成物が必要である。実際に、血管形成の過程は、多数の疾病状態で変更されることが分かっており、多くの場合に、疾病と関連する病理学的損傷は抑制されていない血管形成に関係している。例えば、腫瘍は成長するためには新たな毛細血管の成長を刺激し続けなければならないので、血管形成は腫瘍成長における一因子である。従って、血管または新生血管の内皮細胞の死を促進しまたは誘発することができる化合物は、新たな毛細血管の成長を阻害することができ、これらの疾患の治療に有用なことがある。
【発明の開示】
【0004】
従って、本発明の目的は、内皮細胞死の誘導物質としての活性を示す化合物および医薬組成物を提供することである。
【0005】
本発明者らは、報告された、ネトリン−1が血管形成促進性または抗血管形成性であるという矛盾した結論を生じた明らかに対立する観察結果は、ネトリン−1が内皮細胞の生存因子として作用し、一方、その未結合UNC5B受容体が細胞のアポトーシスを誘発するという事実から生じていることを明らかにした。血管発生に対するネトリン−1の効果は、エクスビボ(ex vivo)アッセイにおけるカスパーゼ阻害剤に似ており、カスパーゼ活性の阻害またはUNC5B受容体のようなネトリン−1受容体のサイレンシングは、いずれもゼブラフィッシュにおけるネトリン−1サイレンシングによって誘発される血管発芽欠損を救済するのに十分である。
【0006】
従って、未結合UNC5Bのアポトーシス促進効果と内皮細胞上のネトリン−1に結合したUNC5Bとの生存効果は、血管形成過程中に協同するものと思われる。
【0007】
第一の態様では、本発明は、ネトリン−1が血管形成中の内皮細胞アポトーシスを阻害することを見出したことに基づく内皮細胞の死を誘発することができる化合物を選択するインビトロでの方法に関する。
【0008】
従って、本発明は、内皮細胞死を誘発することができる化合物を選択するインビトロでの方法であって、
a)ネトリン−1、またはその断片、および内皮細胞を含む培地を有し、
b)前記培地を、試験する化合物と接触させ、
c)前記内皮細胞の死を測定し、および
d)段階c)における測定が前記化合物の存在下にて内皮細胞死の有意な誘発を示しているときに、前記化合物を選択する
段階を含んでなる、方法に関する。
【0009】
特定態様では、試験する化合物の存在によって誘発される細胞死は、下記の材料および方法「細胞死分析」の項に示されているトリパンブルー染色法を用いることによって分析しまたは評価することができる(Mehlen et al., Nature, 1998も参照されたい)。
【0010】
もう一つの態様では、本発明は、内皮細胞死を誘発することができる化合物を選択する方法であって、
a)少なくとも1種類のネトリン−1受容体を発現する内皮細胞を含む培地を有し、
b)前記培地を、試験する化合物と接触させ、
c)前記の試験する化合物のこのネトリン−1受容体のアポトーシス促進活性を促進する能力を測定し、および
d)段階c)における測定が、前記化合物が、このネトリン−1受容体の、好ましくは更にネトリン−1の存在下にてこのネトリン−1受容体の、アポトーシス促進活性を有意に促進することを示しているときに、前記化合物を内皮細胞死の誘導物質として選択する
段階を含んでなる、方法に関する。
【0011】
もう一つの態様では、本発明は、
段階c)の測定が、試験する化合物の細胞死関連タンパク質キナーゼ(DAPK)のキナーゼ活性の活性化を誘発する能力を測定することにあり、および
d)段階c)における測定が、前記化合物の存在下にて、好ましくは更にネトリン−1の存在下にて細胞死関連タンパク質キナーゼ(DAPK)のキナーゼ活性の有意な活性化を示しているときに、前記化合物を内皮細胞死の誘導物質インデューサーとして選択する、
本発明による内皮細胞の死を誘発することができる化合物の選択方法に関する。
【0012】
もう一つの態様では、本発明は、
段階c)の測定が、試験する化合物のDAPKリン酸化を阻害する能力を測定することにあり、および
d)段階c)における測定が、前記化合物の存在下にて、好ましくは更にネトリン−1の存在下にてDAPKリン酸化の有意な阻害を示しているときに、前記化合物を内皮細胞死の誘導物質として選択する、
請求項2に記載の内皮細胞死を誘発することができる化合物の選択方法に関する。
【0013】
もう一つの態様では、本発明は、内皮細胞死を誘発することができる化合物の選択方法であって、
a)少なくとも1種類のネトリン−1受容体を発現する内皮細胞を含む培地を有し、かつ前記ネトリン−1受容体細胞内ドメインがネトリン−1の存在下にて二量体化または多量体化することができ、
b)前記培地を、試験する化合物と接触させ、培地が更にネトリン−1、またはネトリン−1受容体の細胞外ドメインと相互作用することができるその断片を含み、
c)前記ネトリン−1受容体細胞内ドメインの二量体化または多量体化が上記の試験する化合物の存在下で阻害されるかどうかを決定し、および
d)段階c)における決定が、前記内皮細胞の前記ネトリン−1受容体の細胞内ドメインの二量体化または多量体化を有意に阻害することを示しているときに、前記化合物を内皮細胞死の誘導物質として選択する
段階を含んでなる、方法に関する。
【0014】
試験する化合物の存在下における前記ネトリン−1受容体細胞内ドメインの二量体化または多量体化の阻害の決定分析は、例えば、2007年9月7日に公表された特許文献Mehlen et al., WO2007/099133号に見出すことができる。
【0015】
好ましい態様では、段階a)の培地はネトリン−1を更に含む。
【0016】
もう一つの好ましい態様では、本発明による内皮細胞の死を誘発することができる化合物を選択する方法において、前記化合物は血管または新生血管の死を誘発することができる。
【0017】
好ましい態様では、本発明による方法の段階a)において、前記内皮細胞は、DCC(Deleted in Colorectal Cancer)、UNC5A、UNC5B、UNC5CおよびUNC5D、ネオゲニンおよびアデノシンA2bの群から選択される少なくとも1種類のネトリン−1受容体、好ましくはUNC5Bを発現する内皮細胞である。
【0018】
ヒトネトリン−1、またはDCC、UNC5A、UNC5B、UNC5CおよびUNC5D、ネオゲニンおよびアデノシンA2b(Unc−5ホモログ1、2および3はUnc−5ホモログA、BおよびCと同義である)のようなヒトネトリン受容体のアミノ酸配列は、当業者に周知である。特定ドメインの局在化を有するこれらのアミノ酸配列の例は、例えば、ヒトネトリン−1についてはGenbankに受入番号AAD09221またはNP004813、ヒトネトリン受容体DCC前駆体についてはNP_005206、ヒトネトリン受容体Unc−5ホモログ1についてはNP588610、ネトリン受容体Unc−5ホモログ2についてはQ8IZJ1、およびUnc−5ホモログ3についてはO95185、およびヒトUNC5DについてはAAQ88514により見出すことができる。
【0019】
更に好ましくは、ネトリン−1受容体UNC5Bを発現する内皮細胞である。
【0020】
もう一つの好ましい態様では、本発明による方法において、段階a)の前記内皮細胞は、ヒトまたはニワトリの内皮細胞からなる群、好ましくはヒトまたはニワトリ血管または新生血管、更に好ましくはヒト臍帯静脈内皮細胞(HUVEC)およびヒト臍帯動脈内皮細胞(HUAEC)またはヒナドリ漿尿膜から選択される内皮細胞である。
【0021】
第二の態様では、本発明は、
ネトリン−1と上記ネトリン−1受容体との相互作用を特異的に阻害することができるネトリン−1受容体またはその断片の細胞外ドメインを含んでなる化合物、
ネトリン−1またはネトリン−1受容体に対して特異的な、特に前記ネトリン−1受容体の細胞外ドメインまたは前記ネトリン−1受容体の細胞外ドメインと相互作用することができるネトリン−1断片に対するモノクローナルまたはポリクローナル抗体、および
ネトリン−1タンパク質をコードする核酸に特異的なアンチセンスおよびRNAi(干渉RNA)オリゴヌクレオチドからなる群から選択されるネトリン−1発現を阻害することができる化合物
からなる群から選択される、内皮細胞死の誘導物質としての、化合物を含んでなる。
【0022】
好ましくは、本発明は、血管または新生血管の死を誘発する内皮細胞死の誘導物質としての化合物に関する。
【0023】
更に好ましい態様では、本発明は、腫瘍の血管または新生血管の死の誘導物質としての化合物に関する。
【0024】
もう一つのより好ましい態様では、前記腫瘍の腫瘍細胞はネトリン−1を発現または過剰発現する。
【0025】
「ネトリン−1と前記ネトリン−1受容体との間の相互作用を特異的に阻害することができる」という表現におけるネトリン−1とそのネトリン−1受容体との間の相互作用という用語は、本明細書では、一般にリガンドとその受容体との間の特異的結合による相互作用を表すことを指している。この相互作用の阻害は、一般的には、特に競合的リガンド(前記ネトリン−1受容体のこの細胞外膜ドメインに対する抗体など)の存在下、またはネトリン−1(そのネトリン−1受容体の可溶性細胞外膜ドメイン、またはその部分など)と特異的複合体を形成することができる化合物の存在下にて、ネトリン−1のその受容体への結合の完全または部分的阻害によって得ることができる。
【0026】
前記ネトリン−1受容体断片は、ネトリン−1受容体の細胞外ドメイン、またはネトリン−1と特異的に相互作用することができるその部分を含んでなるか、またはそれらである。
【0027】
前記ネトリン−1および/または前記ネトリン−1受容体は、好ましくは哺乳類由来であり、特にマウス、ラットまたはヒト由来であり、更に好ましくはヒト由来である。前記ネトリン−1は、組換えネトリン−1であることができる。前記ネトリン−1は、ニワトリ由来であることもできる。
【0028】
本明細書で用いられる「抗体」という用語は、免疫グロブリン分子および免疫グロブリン分子の免疫学的に活性な部分、すなわちネトリン−1タンパク質またはその受容体と特異的に結合する(免疫反応する)抗原結合部位を含む分子を表す。
【0029】
「抗体」という用語は、モノクローナルまたはポリクローナル抗体だけでなく、キメラまたはヒト化抗体をも含んでなる。
【0030】
単離したネトリン−1タンパク質またはネトリン−1受容体タンパク質、またはそれらの特異的断片は、ポリクローナルおよびモノクローナル抗体調製の標準的手法を用いてこのようなタンパク質を結合する抗体を生成するための免疫原として用いることができる。また、これらの特異抗体を生成するのに用いてもよい少なくとも1種類の抗原決定基を含むこれらのタンパク質の任意の断片を用いてもよい。
【0031】
タンパク質免疫原は、典型的には、適当な被検体(例えば、ウサギ、ヤギ、マウスまたは他の哺乳類)を免疫原で免疫することによって抗体を調製するのに用いられる。適当な免疫原製剤は、前記タンパク質またはその断片を含むことができ、更にフロイントの完全または不完全アジュバントのようなアジュバント、または同様な免疫刺激剤を含むこともできる。
【0032】
従って、本発明によって用いられる抗体としては、ポリクローナル、モノクローナルキメラまたはヒト化抗体が挙げられる。ネトリン−1タンパク質またはその受容体のアミノ酸配列の少なくとも8〜10個のアミノ酸の連続スパンを含んでなるポリペプチドを含むエピトープに選択的に結合することができるまたは選択的に結合している抗体。インタクト(intact)抗体またはその機能性断片(例えば、FabまたはF(ab’)2、scFv)を用いることができる。
【0033】
例えば、候補オリゴヌクレオチドの検出のためのインビトロ手法としては、ノーザンまたはサザンブロットハイブリダイゼーションおよびインサイチュー(in situ)ハイブリダイゼーションが挙げられる。候補タンパク質の検出のためのインビトロ手法としては、酵素免疫測定法(ELISA)、ウェスタンブロット、免疫沈澱および免疫蛍光が挙げられる。
【0034】
アンチセンスは、ネトリン−1コード配列のmRNAまたはcDNA、またはその相補性配列に特異的にハイブリダイズする長さが少なくとも10、15または20ヌクレオチドのオリゴヌクレオチドであることができる。ネトリン−1コード配列の対応する断片と少なくとも95%、98%、99%および100%の同一性を有するオリゴヌクレオチドが好ましい。
【0035】
干渉RNA(RNAi)は、二本鎖RNA(dsRNA)がその相補性配列を有する遺伝子の発現を特異的に抑制する現象である。RNAiは、従って多くの生物の有用な研究手段となっている。dsRNAにより遺伝子発現を抑制する機構は完全には理解されていないが、実験データは重要な識見を提供している。この手法は、哺乳類細胞における遺伝子機能を研究する手段として大きな可能性を有しており、siRNA(低分子干渉RNA)に基づく薬剤の開発へと導くことができる。
【0036】
好ましくは、本発明の化合物において、ネトリン−1受容体またはその断片の前記細胞外ドメインは、DCC、UNC5A、UNC5B、UNC5CおよびUNC5D、ネオゲニンおよびアデノシンA2bからなる群から選択され、更に好ましくはDCC、UNC5A、UNC5BおよびUNC5Cの群から選択される。UNC5Bが、さらにより好ましい。
【0037】
もう一つの態様では、本発明は、ネトリン−1を発現または過剰発現する腫瘍細胞を含む腫瘍に罹っている患者を治療するための癌の進行を阻害することを目的とする医薬組成物の調製のための本発明による化合物に関する。
【0038】
好ましくは、前記癌は、乳癌、結直腸癌、肺癌、神経芽腫、神経膠腫、急性脊髄性白血病、肉腫、黒色腫、卵巣腺癌、腎腺癌、膵腺癌、子宮腺癌、胃腺癌、肝腺癌および直腸腺癌からなる群から選択される。
【0039】
好ましくは、前記癌は転移性または侵攻性癌である。
【0040】
本発明の化合物を患者に投与するときには、場合によっては薬学上許容可能なビヒクルを含んでなる組成物の成分として投与するのが好ましい。この組成物は、経口または任意の他の好都合な経路によって投与することができ、別の生物活性剤と共に投与してもよい。投与は、全身または局所的であることができる。例えば、リポソームでのカプセル化、微小粒子、マイクロカプセル、カプセルなどの様々な送達系が知られており、選択された本発明の化合物またはその薬学上許容可能な塩を投与するのに用いることができる。
【0041】
投与方法としては、皮内、筋肉内、腹腔内、静脈内、皮下、鼻内、硬膜外、経口、舌下、鼻内、脳内、膣内、経皮、直腸、吸入、または局所的投与が挙げられるが、これらに限定されない。投与様式は、現場実務者の裁量に任される。ほとんどの場合に、投与によって、化合物は血流中にまたは直接原発性腫瘍に放出される。
【0042】
本発明による、または本発明による方法によって選択された化合物を含んでなる組成物も、本発明の部分を形成する。これらの組成物は、薬学上許容可能なビヒクルの適量をさらに含んでなり、患者に適正に投与するための形態を提供するようにすることもできる。「薬学上許容可能な」という用語は、規制当局によって認可され、または動物、哺乳類、更に詳細にはヒトで使用するための国定または承認された薬局方に記載されていることを意味する。「ビヒクル」という用語は、希釈剤、アジュバント、賦形剤またはキャリヤーであって、これによって本発明の化合物を投与するものを表す。このような薬学ビヒクルは、水および油のような液体であり、ピーナッツ油、大豆油、鉱油、ゴマ油など石油、動物、植物または合成起源のものであることができる。薬学ビヒクルは、塩水、ゼラチン、澱粉などであることができる。更に、補助剤、安定剤、増粘剤、滑沢剤および着色料を用いてもよい。食塩溶液、水性デキストロースおよびグリセロール溶液を、特に注射溶液の液体ビヒクルとして用いることもできる。適当な薬学ビヒクルとしては、澱粉、グルコース、ラクトース、スクロース、ゼラチン、ステアリン酸ナトリウム、モノステアリン酸グリセロール、塩化ナトリウム、乾燥スキムミルク、グリセロール、プロピレン、グリコール、水などの賦形剤も挙げられる。試験化合物の組成物は、所望ならば、少量の湿潤剤または乳化剤、またはpH緩衝剤を含むこともできる。本発明の組成物は、溶液、懸濁液、エマルション、錠剤、丸薬、ペレット、カプセル、液体を含むカプセル、散剤、徐放性処方物、座薬、エマルション、エアゾール、スプレー、懸濁液の形態、または使用に適当な任意の他の形態を採ることができる。前記組成物は、通常は経口投与または静脈内投与の目的でヒトに適合させた医薬組成物として常法に準じて処方される。活性化合物の量または治療に有効となる量は、標準的臨床手法によって決定することができる。更に、インビトロまたはインビボ(in vivo)アッセイを場合によって用いて、最適用量範囲を確認しやすくすることができる。用いられる正確な用量は投与経路および疾病の重篤度によって変化し、現場実務者の判断およびそれぞれの患者の環境によって決定されるべきである。しかしながら、経口、鼻内、皮内または静脈内投与は、通常は約0.01〜約75mg/kg体重/日、更に好ましくは約0.5〜5mg/kg体重/日である。
【0043】
第三の態様では、本発明は、内皮細胞、好ましくは血管または新生血管由来の内皮細胞の死を誘発することができる化合物を選択するためのキットであって、
ネトリン−1タンパク質またはネトリン−1受容体タンパク質と特異的に相互作用して結合対を形成することができるその断片、および
ネトリン−1受容体、好ましくはDCC、UNC5A、UNC5B、UNC5CおよびUNC5D、ネオゲニンおよびアデノシンA2bからなる群から選択され、更に好ましくは受容体UNC5Bを発現する内皮細胞
を含んでなる、キットに関する。
【0044】
好ましい態様では、前記キットは、内皮細胞HUVECまたはHUAEC細胞を含んでなる。
【0045】
本発明を上記態様に関して説明してきたが、上記説明および下記の実施例は本発明を例示するためのものであり、本発明の範囲を限定するためのものではないことを理解すべきである。本発明の範囲内における他の態様、利点および変更は、本発明が関係する当業者には明らかになるであろう。
【図面の簡単な説明】
【0046】
【図1A】図1A〜1H:ネトリン−1は、恐らくはUNC5Bによって引き起こされるアポトーシスを阻害することにより内皮細胞アポトーシスを防止する。 図1A:ヒト臍帯静脈/動脈内皮細胞(HUVEC/HUAEC)はUNC5AおよびUNC5B依存性受容体を発現するが、ネトリン−1を発現しない。定量RT−PCRは、実験手順に記載の通りに行った。
【図1B】図1B:ネトリン−1投与は、トリパンブルー排除によって測定される血清飢餓時に見られるHUVEC/HUAEC細胞死を阻害する。少なくとも100個の細胞/条件を計数した。相対指数は、平均値+/−s.e.m(n=3)として示す。*:p値<0.005;**:p値<0.0001(Student t検定)。
【図1C】図1C:ネトリン−1は、TUNEL染色によって測定したHUVECにおけるアポトーシスを防止する。TUNEL陽性細胞の定量は、平均値+/−s.e.mである(Student t検定)。
【図1D】図1D:ネトリン−1投与により、HUVECおよびHUAECにおけるカスパーゼ−3活性が減少する。カスパーゼ−3活性の相対指数は、平均値+/−s.e.m(n=3)として表される。開裂したカスパーゼ−3陽性細胞の免疫染色および相対指数(平均値+/−s.e.m)が示される(Student t検定)。
【図1E】図1E:ネトリン−1投与により、HUVECおよびHUAECにおけるカスパーゼ−3活性が減少する。カスパーゼ−3活性の相対指数は、平均値+/−s.e.m(n=3)として表される。開裂したカスパーゼ−3陽性細胞の免疫染色および相対指数(平均値+/−s.e.m)が示される(Student t検定)。
【図1F】図1F:siRNAによる、UNC5Aではなく、UNC5Bサイレンシングは、カスパーゼ−3活性の減少と関連している。値は、平均値およびs.e.m(n=3)である。総て、p<0.05(コントロール条件におけるレベルと比較したStudent t検定)。
【図1G】図1G:DAPキナーゼは、HUVECおよびHUAECにおけるUNC5Bによって引き起こされるアポトーシスを介在する。DAPキナーゼΔCaM(DAPキナーゼの構成性の活性突然変異体型)およびDAPキナーゼDD(DAPキナーゼのドミナントネガティブ型)の強制発現後の内皮細胞における細胞死(Toxilight)。値は平均値およびs.e.m.である。総て、p<0.001(コントロール条件におけるレベルと比較したStudent t検定)。
【図1H】図1H:ネトリン−1投与により、DAPキナーゼリン酸化が誘発される。抗ホスホ−DAPキナーゼを用いて、酵素活性のレベルを測定した。
【図2A】図2A〜2F:カスパーゼ阻害剤およびUNC5Bサイレンシングは、新生血管形成におけるネトリン−1効果に似ている。 図2A:ネトリン−1またはカスパーゼ阻害剤z−VAD−fmkおよびBAFは、エクスビボでのネズミ大動脈リングマトリゲルアッセイにおける微小血管形成を促進する。少なくとも8種類のマウスから切除された大動脈リングを、条件毎に示す。微小血管全長(ピクセル単位)の定量は、実験手順に記載の通りに行った。総てのp値を示す(χ2検定)。
【図2B】図2B:ネトリン−1またはカスパーゼ阻害剤z−VAD−fmkおよびBAFは、エクスビボでのネズミ大動脈リングマトリゲルアッセイにおける微小血管形成を促進する。少なくとも8種類のマウスから切除された大動脈リングを、条件毎に示す。微小血管全長(ピクセル単位)の定量は、実験手順に記載の通りに行った。総てのp値を示す(χ2検定)。
【図2C】図2C:ネトリン−1およびカスパーゼ阻害剤は、CAMアッセイにおける新生血管形成を誘発する。CAMモデルは、材料および方法に記載の通りに処理した8日齢のヒナドリ胚(n≧10)を用いて調製した。メチルセルロースディスクに輪郭を描く。デノボ(de novo)血管形成は、分岐点の数として表す(n≧13フィールド/条件)。平均値は、直線によって示す。*:p値<0.02;**:p値<0.0005(Mann−Whitney U検定)。
【図2D】図2D:ネトリン−1およびカスパーゼ阻害剤は、CAMアッセイにおける新生血管形成を誘発する。CAMモデルは、材料および方法に記載の通りに処理した8日齢のヒナドリ胚(n≧10)を用いて調製した。メチルセルロースディスクに輪郭を描く。デノボ血管形成は、分岐点の数として表す(n≧13フィールド/条件)。平均値は、直線によって示す。*:p値<0.02;**:p値<0.0005(Mann−Whitney U検定)。
【図2E】図2E:siRNAによるUNC5Bサイレンシングは、CAMアッセイにおける新生血管形成を増加する。実験手順に記載の通りに3回のsiRNA注射をE.5とE.9との間に行った。スクランブルsiRNAを、コントロールとして用いた。デノボ血管形成は、分岐点の数として表す(n>30異なるフィールド/条件)。平均値は、直線によって示す。P値=0.009(Mann−Whitney U検定)。
【図2F】図2F:siRNAによるUNC5Bサイレンシングは、CAMアッセイにおける新生血管形成を増加する。実験手順に記載の通りに3回のsiRNA注射をE.5とE.9との間に行った。スクランブルsiRNAを、コントロールとして用いた。デノボ血管形成は、分岐点の数として表す(n>30異なるフィールド/条件)。平均値は、直線によって示す。P値=0.009(Mann−Whitney U検定)。
【図3A】図3A〜3C:ノックダウンネトリン−1aゼブラフィッシュ胚の表現型は、アポトーシスの阻害によって救済される。 fli:egfpゼブラフィッシュ胚を、1〜4細胞期でコントロール(ctl)、スプライス(spl)または翻訳(tsl)遮断ネトリン1a、UNC5BおよびDAPキナーゼモルホリノと共に単独でまたは組み合わせて投与した。表現型を、24〜54hpfに分析した。主要血管構造の典型的なイメージを示す。左が先。カスパーゼ阻害剤BAFは、ネトリン−1a splモルホリノを投与したゼブラフィッシュ胚では傍脊索血管形成(星印)を回復し(図3A)、ネトリン−1a tslモルホリノを投与したゼブラフィッシュ胚ではISV形成を回復する(図3B)。同様の表現型の救済は、ゼブラフィッシュ胚におけるUNC5BまたはDAPキナーゼおよびネトリン1a tsl遮断モルホリノの同時投与によって観察された(図3B)。 図3A:定量を示す。p値を示す(χ2検定)。
【図3B】図3B:定量を示す。p値を示す(χ2検定)。
【図3C】図3C:BAF処理またはUNC5B遮断モルホリノ投与は、TUNEL染色(左パネル)またはカスパーゼ−3活性(右パネル)によって示されるようにゼブラフィッシュ胚におけるネトリン1a tsl遮断モルホリノ投与後に観察されるアポトーシスの増加を阻害する。カスパーゼ−3活性は、ネトリン−1a tslモルホリノのみを投与した胚に関する相対指数として表される。平均値およびs.e.m.は、3回の独立した実験から計算される。p値を示す(Student t検定)。
【図4A】図4A〜4G:ネトリン−1は、UNC5Bによって引き起こされる内皮細胞アポトーシスを防止する。 図4A〜4B:HUVECおよびHUAEC細胞におけるネトリン−1発現の評価。 図4A:特異的ネトリン−1抗体を用いる免疫蛍光を用いた。ネトリン−1標識は、MRC−5肺繊維芽細胞系とは反対に、HUVECおよびHUAEC細胞では検出されなかった。
【図4B】図4B:培地に分泌されたネトリン−1の定量は、Apotech Corporation製のネトリン−1(ヒト)ELISAキットを用いて行った。
【図4C】図4C:ネトリン−1処理により、HUVECおよびHUAECにおけるカスパーゼ−3活性は用量依存的に減少する。カスパーゼ−3活性の相対指数は、平均値+/−s.e.mと表される(n=3)。*:p<0.05;**p<0.0025(コントロール条件におけるレベルと比較したStudent t検定)。
【図4D】図4D:ネトリン−1処理により、50μMエトポシドで一晩処理したときに見られたHUVEC/HUAEC細胞死が阻害される。細胞死は、実験手順に記載のToxilightアッセイを用いて評価する。相対指数は、平均値およびs.e.mである(n=3)(p<0.05、Student t検定)。
【図4E】図4E:ラットUNC5H2の細胞外ドメインに対応する組換えタンパク質の添加により、ネトリン−1によって介在される細胞死阻害が防止される。結果は、それぞれの場合にコントロールに対する相対指数として表される(平均値+/−s.e.m,n=3、t検定)。
【図4F】図4F:スクランブルsiRNAでトランスフェクションしたコントロール細胞と比較したUNC5BsiRNAでトランスフェクションした細胞におけるUNC5B発現の定量RT−PCR分析。トランスフェクションの効率を示す、siRNAでトランスフェクションした細胞におけるGFP発現の典型的画像。
【図4G】図4G:スクランブルsiRNAでトランスフェクションしたコントロール細胞と比較したUNC5AsiRNAでトランスフェクションした細胞におけるUNC5A発現の定量RT−PCR分析。
【図5A】図5A〜5C:エクスビボセッティングにおけるUNC5Bの発現。 図5A:大動脈リングでのUNC5B発現の定量RT−PCR分析。
【図5B】図5B:CAMでの定量RT−PCRによるネトリン−1、UNC5H2(マウスUNC5B)およびDAPキナーゼ発現の分析。ネトリン−1とは反対に、UNC5H2およびその下流エフェクターが発現する。
【図5C】図5C:CAMでのUNC5B発現の定量RT−PCR分析、およびスクランブルsiRNAを用いてトランスフェクションしたコントロールと比較したCAMでのUNC5B発現に対する血液循環に投与したUNC5BsiRNAの効果。
【図6A】図6A〜6D:ゼブラフィッシュ胚におけるネトリン−1aおよびUNC5Bモルホリノの効果。 図6A:ネトリン−1bは、ゼブラフィッシュにおける発生上の血管形成に関与しているとは思われない。翻訳遮断ネトリン−1bモルホリノは、発生上の血管形成に対して重大な欠損を持っていない。欠損のない胚の割合および分析した胚の総数を示す。ネトリン−1オーソログ(otholog)遺伝子の系統発生的研究。100ブートストラップ複製を用いるJTTモデルでのタンパク質配列に関するML。ゼブラフィッシュにおけるネトリン−1オーソログであるネトリン−1aおよびネトリン−1bには、下線を付けている。
【図6B】図6B:ゼブラフィッシュ胚におけるネトリン−1aおよびUNC5B発現のRT−PCR分析。
【図6C】図6C:第三のネトリン−1a遮断モルホリノは、図3A〜3Cに記載のものと同様な血管の欠損を生じる。ISV欠損を有する胚の割合および分析した胚の総数を示す。
【図6D】図6D:ネトリン−1aサイレンシングは、全ゼブラフィッシュ胚のカスパーゼ−3活性を増加させる。カスパーゼ−3活性は、コントロールMOを投与した胚に関する相対指数として表される。平均値およびs.e.m.は、3回の独立した実験から計算される。
【図7】DCC−5Fbnデコイペプチドは、内皮細胞に対するネトリン生存効果を逆転させる。HUVECにおけるネトリン−1によるカスパーゼ−3活性化の減少は、DCCの第五の細胞外免疫グロブリンドメインに対応するデコイペプチドDCC−5Fbnによる細胞の同時処理によって減少する。カスパーゼ−3活性の相対的減少割合は、平均値+/−s.e.mとして表される(n=3)。Student t検定:p<0.015。
【0047】
材料および方法
A)内皮細胞培養およびトランスフェクション
内皮細胞培養およびトランスフェクション:
ヒト臍帯静脈および動脈細胞(Promocell,継代数<8)を、2%血清(キット、Promocell)を補足した内皮細胞成長培地(ECGM,Promocell)で80%コンフルエンスに達するまで培養した。HUVECを、AMAXA Nucleofectorシステムを用いて製造業者の指示に従ってトランスフェクションした。簡単に説明すれば、500,000個の細胞を100μlのHUVEC Nucleofector溶液に再懸濁し、1μgsiRNAまたは1.5μgプラスミドDNAおよび0.5μgGFPコードコントロールプラスミドを用いてトランスフェクションした。UNC5A、UNC5BおよびスクランブルsiRNAは、Sigma−Proligoによってデザインされた。突然変異体DAPキナーゼ(DAPキナーゼΔCamおよびDAPキナーゼDD)をコードするプラスミドは、以前に記載された(Llambi et al.,2005)。トランスフェクション後、細胞を1%成長培地で培養し、24時間後に回収した。
【0048】
B)細胞死アッセイ
細胞死アッセイは、上記に記載の通りに行った。簡単に説明すれば、内皮細胞を、10,000個/cm2を完全培地に16時間播種した。細胞死は、無血清培地(内皮細胞基礎培地、Promocell)での飢餓によりまたは50μMエトポシドに暴露することにより誘発した。ネトリン−1は、Axxora−Apotechから購入した。細胞死は、上記の通りにトリパンブルー排除アッセイによって評価した。DNA断片化を検出するため、カバーグラス上で培養した処理済み細胞を4%パラホルムアルデヒド(PFA)に20分間固定し、ターミナルデオキシヌクレオチジルトランスフェラーゼ介在dUTP−ビオチンニック末端標識(TUNEL)を、300U/mlTUNEL酵素(300U/ml)および6μMビオチニル化dUTP(Roche Diagnostics)で行った。カスパーゼ−3活性を、Clontech(米国)製ApoAlert CPP32キットを用いて、文献記載の抗活性カスパーゼ−3抗体(Cell signaling)を用いる固定細胞での免疫染色によって測定した(Llambi et al.,2005; Tauszig-Delamasure et al.,2007)。Toxilightアッセイは、製造業者の指示に従って行った(Lonza)。ネトリン−1滴定実験のため、ラット組換えUNC5H2−Fc(R&D)を1μg/mlで用いた。
【0049】
C)ウェスタンブロット分析および免疫蛍光
10,000個/cm2で播種し、無血清培地(内皮細胞基礎培地,Promocell)中でネトリン−1と共に24時間処理したまたは処理しなかった内皮細胞を回収して、プロテアーゼ阻害剤の存在下にて50mMHEPES pH7.6、125mMNaCl、5mMEDTAおよび0.1%NP−40緩衝液中で溶解した。タンパク質抽出物を、抗DAP−キナーゼおよび抗ホスホ−DAPキナーゼ抗体(Sigma)で免疫ブロットした。免疫蛍光分析のため、細胞をLabTekチャンバースライド上に播種し、4%PFAで15分間固定した。抗ネトリン−1ラットモノクローナル抗体(R&D)を1/150の希釈倍率で用い、特異的二次抗体(Alexa488 Donkey anti-Rat,Molecular Probes-Invitrogen)を用いて可視化した。
【0050】
D)定量RT−PCR
HUVEC/HUAEC、大動脈リング、CAMおよびゼブラフィッシュ胚におけるネトリン−1、依存性受容体、DCCおよびUNC5A−D、およびDAPキナーゼ発現を評価するため、総RNAをNucleospin RNAIIキット(Macherey-Nagel)を用いて抽出し、1μgをiScript cDNA Synthesisキット(BioRad)を用いて逆転写した。リアルタイム定量RT−PCRを、LightCyclerFastStartDNAMasterSYBERGreenIキット(Roche)を用いてLightCycler2.0装置(Roche)上で行った。総ての最適増幅のための反応条件並びにプライマー選択は、上記の通りに決定した。プライマーの配列は、必要であれば入手可能である。
【0051】
E)大動脈リングアッセイ
大動脈リングアッセイは、Nicosiaと共同研究者らによって記載されている方法によって行った(Nicosia and Ottinetti,1990)。大動脈は6〜8週齢のBalb/Cマウスから切除し、外膜周囲の脂肪および結合組織を除去し、ECGMで2回洗浄し、1〜1.5mmの長さのリングに切断した。成長因子減少マトリゲル(Growth factor reduced Matrigel; BD Biosciences)100μlを、48穴組織培養プレート(Costar)のそれぞれのウェルに添加し、37°Cで30分間固化させた。大動脈リングをマトリックス層に置き、Matrigel100μlに埋設し、完全成長培地(ECGM,Promocell)で24時間回復させた。次いで、それらをネトリン−1(150ng/ml)、z−VAD−fmk(Tebu;20nM)、BAF(Sigma;20nM)またはVEGF(Sigma;13ng/ml)で処理しまたは処理せずに、4日目に位相差顕微鏡で写真を撮った。微小血管生長の全長は、AxioVisionRelease4.6ソフトウェアを用いて計算した。
【0052】
F)CAMアッセイ
ヒナドリ漿尿膜(CAM)アッセイは、上記に記載の通りに行った。白色レグホンの卵を、加湿雰囲気下にて37°Cでインキュベーションした。72時間インキュベーションした後、3mlの卵白を除去し、四角い窓を殻に開けた。開口部は、8日目までセロテープで閉じた。次いで、ネトリン−1(150ng/ml)、z−VAD−fmk(Tebu;20nM)、BAF(Sigma;20nM)またはVEGF(Sigma)をPBSで希釈し、等容のオートクレーブ処理した1%メチルセルロース溶液と混合した。コントロールは、ビヒクルのみで調製した。それぞれの試料溶液(10μl)をパラフィルムに滴下して乾燥した。メチルセルロースディスクをパラフィルムから外して、8日齢ヒナドリ胚のCAM上に置いた。3日後、CAMを撮影して、血管形成応答をAxioVisionRelease4.6ソフトウェアを用いてメチルセルロースディスク下で二次異所性血管芽の数を測定することによって評価した。CAMにおけるUNC5Bサイレンシングは、E.5、E.7およびE.9でのCAM血液循環中でSigma−Proligoによってデザインされた特異的siRNAを投与することによって行った。分析は、E.11で行った。
【0053】
G)ゼブラフィッシュ胚のモルホリノノックダウン
トランスジェニックfli:egfpゼブラフィッシュをZebrafish International Resource Center(オレゴン大学)から入手し、28°Cで標準的飼育条件下に保持した(Lawson and Weinstein,2002)。胚をプロナーゼ酵素処理を用いてコリオン除去し、6ngのスプライス/翻訳遮断ネトリン−1、UNC5Bおよび予めデザインして特性決定したコントロールモルホリノを、単独でまたは組み合わせて1〜4細胞期の胚の高卵黄に投与した。第三のスプライス遮断モルホリノ特異ターゲッティングネトリン−1aをデザインし(GeneTools)、同じ用量で胚に投与した。最後に、DAPキナーゼ(GeneTools)の翻訳遮断モルホリノターゲッティングゼブラフィッシュオーソログ3ngを投与し、コントロールと比較してノックダウン胚におけるDAPキナーゼmRNA発現を35%減少させた。モルホリノの配列は、必要であれば入手可能である。胚を0.003%フェニルチオ尿素を補足したE3培地で培養し、色素形成を阻害した。ネトリン−1aサイレンシングによって誘発される表現型の救済を、モルホリノ投与の4時間後にE3培地に40nMBAFを加えることによって試験した。TUNEL染色のため、胚を4%PFA中で48hpfで4°Cにて一晩固定した。次いで、それらをPBS−Tween0.1%(PBST)で10分間洗浄し、連続的メタノール槽(25、50、75および100%)で累進的に脱水した。再水和(75、50および25%メタノール系)の後、胚をPBSTで洗浄し、PBS中で10μg/mlプロテイナーゼK(Roche)で5分間処理し、PBSTで再度洗浄し、PBS−BSA1%中で30分間インキュベーションした。TUNEL標識は、予め上記したとおりに行い、Cy3結合−ストレプトアビジン(1/400;Jackson Immunoresearch)を用いて可視化した。カスパーゼ−3活性アッセイは、24時間発生時に胚全体について上記の通りに行った。微速度撮影実験のため、ゼブラフィッシュ胚を0.016%トリカインを含む3%メチルセルロース溶液に埋設し、運動を抑制した(Isogai et al.,Development and Disease,2003)。透過ビデオ顕微鏡画像化を、Timelapse Axiovert100M(PLATIM,ENS Lyon)を用いて行った。
【0054】
DCC−5Fbn産生:DCC−5Fbn産生は、標準的手順を用いて行うことができる。簡単に説明すれば、BL21細胞をイミダゾールに応じてDCC−5Fbnを強制的に発現させ、BL21溶解物をFlag−アガロース(Sigma)を用いてアフィニティークロマトグラフィーにかけた。
【実施例】
【0055】
本発明者らは、本明細書において、これらの2つの明らかに対立する観察を、ネトリン−1の未結合UNC5B受容体によって誘発されるアポトーシスを遮断することにより、血管形成中に内皮細胞の生存を促進するネトリン−1の能力を説明することができるかどうかを検討した。実際に、ネトリン−1は、いわゆる依存性受容体DCCおよびUNC5A−Dについてのリガンドとして作用することによって上皮細胞死を予防し、それらのリガンドの存在下および非存在下のいずれにおいても活性な機能特性を共有しており、リガンド結合は正のシグナル伝達を誘発するが、依存性受容体は別のシグナル伝達カスケードを誘発し、それらの栄養リガンドから解放されたときに細胞死を生じる(Llambi et al.,2001; Mehlen et al.,1998; Tanikawa et al.,2003)(Bredesen et al.,2005; Mehlen and Bredesen,2004)。このスキームでは、UNC5Bの喪失、すなわちアポトーシス促進シグナル伝達の喪失は、実際に血管数の増加と関連している(Lu et al.,2004)。一方、ネトリン−1の喪失、すなわちアポトーシス促進シグナル伝達の増加は、血管消失と関係していると予想される(Wilson et al.,2006)。このような内皮細胞生存とアポトーシスとの間の動的バランスは、血管の完全性の保持を助長する可能性があり、これはまた、以前に報告されているように血管発生および病理学的血管形成の際の血管ホメオスタシスにも重要である(Alavi et al.,2003; Carmeliet,2005; Duval et al.,2003; Santoro et al.,2007; Winn et al.,2005)。
【0056】
例1:ネトリン−1はその未結合UNC5B受容体によって誘発される内皮細胞アポトーシスを遮断する
ネトリン−1がその未結合UNCB受容体によって誘発されるアポトーシスを遮断するかどうかを決定するため、どのネトリン−1依存性受容体、すなわちDCC、UNC5A、UNC5B、UNC5CおよびUNC5Dが、ヒト臍帯静脈/動脈内皮細胞(HUVEC/HUAEC)で発現するかを最初に決定した。定量RT−PCRによって示されているように、UNC5AおよびUNC5Bは、いずれもHUVECおよびHUAECで発現するが(図1A)、UNC5C、UNC5DおよびDCCを検出することはできなかった(データは示さず)。ネトリン−1は、これらの細胞では、RNA(図1A)またはタンパク質レベル(図4A〜4B)でも発現しなかった。次いで、ネトリン−1が内皮細胞の生存因子として働くかどうかを、HUVECおよびHUAECが血清飢餓下で経験する細胞死の自発的傾向を遮断するその能力を分析することによって検討した。ネトリンは、50ng/mlの用量が、生理学的かつ機能的であることが以前に報告されているので、総てのインビトロ実験でこの用量を用いた((Serafini et al.,1996; Serafini et al.,1994)および図4C)。図1Bに示されているように、ネトリン−1は、HUVECおよびHUAEC両方の細胞死を阻害した(p<0.005)。同様な結果は、エトポシドによって引き起こされる細胞死についても得られた(図4D)。ネトリン−1は、TUNEL陽性細胞数の減少(図1C)およびカスパーゼ−3活性の減少(図1D〜1E)によって示されるように、内皮細胞のアポトーシスを防止した(HUVECについては30%までおよびHUAECについては40%まで;総てp<0.005)。UNC5H2の細胞外ドメインに対応する組換えタンパク質の添加により、ネトリン−1によって介在される細胞死の阻害が防止され、生存効果はネトリン−1に特異的であるという結論を支持していた(図4E)。従って、ネトリン−1は、内皮細胞に対して生存因子として働く。
【0057】
UNC5Bおよび/またはUNC5Aが内皮細胞のアポトーシスを誘発するかどうかを決定するため、siRNA法を用いてそれらの発現を一過的にサイレンシングした(図1Fおよび図4F〜4G)。UNC5Bサイレンシングにより、カスパーゼ−3活性は減少したが、UNC5A siRNAはカスパーゼ−3活性に対して全く効果を示さなかった(図1F;総てp<0.05)。更に、UNC5Bサイレンシングと一緒のネトリン−1の添加は、追加的生存効果と関連しなかった(図1F)。従って、内皮細胞に対するネトリン−1生存効果は、UNC5B発現、および別の状況では未結合UNC5Bによって誘発されるアポトーシスの阻害からの同様な結果によって変化する。UNC5Bによって引き起こされるアポトーシスはDAPキナーゼによって介在されることが示されているので、最初にHUVECおよびHUAEC細胞で構成性の活性DAPキナーゼDAPKΔCaMを発現させた(Shohat et al.,2001)。図1Gに示されるように、DAPキナーゼの構成性の活性形態の発現により、細胞死が引き起こされた(総てp<0.0001)。次に、DAPキナーゼのドミナントネガティブ突然変異体DAPK DDを強制発現させた(Llambi et al.,2005)。ネトリン−1処理と同様に、DAPK DDは細胞死誘発を防止した。更に、UNC5Bアポトーシス促進シグナル伝達は、DAPキナーゼ自動リン酸化の阻害を介するDAPキナーゼの活性化によって介在されるので(Llambi et al.,2005)、次にDAPキナーゼリン酸化レベルを測定した。図1Hに示されるように、ネトリン−1処理により、恐らくはUNC5Bアポトーシス促進シグナル伝達カスケードの遮断によってHUVECにおけるDAPキナーゼリン酸化が増加した。UNC5B突然変異体胚における血管数の増加が見られたことと一緒に(Lu et al.,2004)、これらのインビトロデータは、ネトリン−1が血管形成の際にUNC5Bによって引き起こされるアポトーシスを遮断することができるという考察を支持している。
【0058】
例2:ネトリン−1の血管形成促進効果は、カスパーゼ阻害およびUNC5Bサイレンシングに似ている
次に、ネトリン−1の血管形成促進活性が、細胞生存を促進するその能力に関するという概念を、新血管形成のエクスビボモデルを用いて検討した。最初に、マウス大動脈リングアッセイを用いて、大動脈外植片におけるUNC5BのネズミオーソログであるUNC5H2の発現を評価した(図5A)。ネトリン−1は、外植片において発現されることは見られなかった(データは示さず)。VEGFで観察されたのと同様に、ネトリン−1処理では、コントロール条件と比較してマトリゲルに埋設した大動脈リングから毛細血管の出芽が増大し(図2A〜2B;p<0.001)、ネトリン−1の血管形成促進活性を支持していた。興味深いことには、カスパーゼ阻害薬z−VAD−fmkおよびBAFも新毛細血管形成を誘発し、ネトリン−1の効果に似ていた(図2A〜2B;p<0.002)。同じ結果は、ヒナドリ漿尿膜(CAM)アッセイで得られた(図2C〜2D、および発現分析についての図5B)。実際に、VEGF、ネトリン−1、z−VAD−fmkまたはBAFで浸漬したメチルセルロースディスク付近では、新血管形成の有意な増加が観察された(図2C〜2D;p<0.02)。従って、これらのエクスビボ生理学的モデルでは、アポトーシスの阻害はネトリン−1の効果に十分似ていると思われる。更に、CAMでのsiRNAによるUNC5Bサイレンシングにより新血管分枝が増加し(図5C)、ネトリン−1およびその受容体UNC5Bは、血管形成過程中に拮抗的に作用する(図2E〜2F;p=0.009)。
【0059】
例3:ゼブラフィッシュ発生中のネトリン−1aサイレンシングは、カスパーゼ阻害およびUNC5B/DAPキナーゼサイレンシングによって救済される血管欠損を生じる
ネトリン−1の血管形成促進および抗アポトーシス活性の間の原因と結果の関係をインビボで更に調査するため、抗アポトーシス処理がゼブラフィッシュ発生中にネトリン−1崩壊によって誘発される血管欠損を救済することができるかどうかを検討した。2種類のネトリン−1遺伝子オーソログであるネトリン−1aおよびネトリン−1bは、ゼブラフィッシュで特性決定されている(Lauderdale et al.,1997; Strahle et al.,1997)。ネトリン−1bサイレンシングは血管発生に対して何ら明らかな効果を持たないが(図6A)、EichmanのグループおよびLiのグループはそれぞれ異なるネトリン−1aモルホリノを用いて、ネトリン−1aのサイレンシングが血管発生欠損と関連していることを示した(Lu et al.,2004; Wilson et al.,2006)。スプライス遮断モルホリノ(spl−ネトリン−1a)を用いるネトリン−1aサイレンシングは、傍脊索血管(parachordal vessels:PAV)の形成を防止することが示されている(Wilson et al.,2006)。fli:egfpトランスジェニックゼブラフィッシュ、血管において特異的にGFPの発現をターゲッティングするトランスジェニックモデル(Lawson and Weinstein,2002)を用いて、受精から50〜54時間後(hpf)に、このようなspl−ネトリン−1aモルホリノを投与した胚の57.3%がPAVを欠いており(n=75)、節間血管(intersegmental vessels:ISV)および背側縦吻合血管(dorsal longitudinal anastomotic vessels:DLAV)の発生は正常である(図3A)ことを観察した。spl−ネトリン−1aを投与した胚を汎用カスパーゼ阻害剤BAFで処理すると、PAV形成は、未処理胚での42.7%と比較して処理済み胚の92.6%(n=54)で見られたので、この表現型を良好に逆転した(図3A;p<0.0001)。以前に報告されているように(Wilson et al.,2006)、翻訳遮断モルホリノ(tsl−ネトリン−1a、図6B)並びに第三のネトリン−1a遮断モルホリノ(図6C)を用いるネトリン−1aサイレンシングにより、ISVの組織崩壊、背側吻合血管(DLAV)の非存在(図3B)および発生上の欠損を生じた。ISV発生の微速度撮影分析により、ISVの非存在または未熟成長停止(premature growth arrest)を確かめた(例4参照)。同様の欠損は、アポトーシスタンパク質スルビビンの小阻害剤についてノックダウンしたゼブラフィッシュ胚に以前に報告されていた(Ma et al.,2007)。更に、カスパーゼ−3活性の増加が、ネトリン−1aノックダウン胚で観察された(図6D)。これらの胚のBAFによる処理は、コントロールでの23.0%と比較して処理済み胚の54.5%が少なくとも部分的ISVを示したので(図3B;p<0.001)、ISV形成の救済に十分であった。依存性受容体概念に従って、UNC5Bモルホリノの同時投与(図6B)は、胚の79.6%(n=54)においてtsl−ネトリン−1aモルホリノ投与によって誘発される欠損を修正することもできた(図3B;p<0.0001)。同様の救済は、tsl−ネトリン−1a遮断モルホリノとモルホリノターゲッティングDAPキナーゼの共投与によって得られた(図3B;p<0.0001)。
【0060】
BAF処理またはUNC5Bモルホリノ投与は、TUNEL免疫染色アッセイおよびカスパーゼ−3活性アッセイ(図3C)によって測定されるように、ネトリン−1aサイレンシング後の胚で観察される細胞死の増加を救済し、ネトリン−1aによって阻害された内皮細胞死は、血管発生中にUNC5Bによって特異的に介在されるという考えを更に支持する。
【0061】
例4:fli−EGFPゼブラフィッシュ胚における主要血管発生に対するネトリン−1aモルホリノ効果の微速度撮影分析
fli−EGFPトランスジェニックゼブラフィッシュにおける血管動力学の微速度撮影画像を得ることができる。従って、節間血管(ISV)の最初の出現は、3分間の時点間隔で収集した時点によって示すことができる。
【0062】
これらの微速度撮影画像(データは示さず)は、ISVおよび背側縦吻合血管(DLAV)形成は、通常はコントロールモルホリノを投与したfli−EGFPゼブラフィッシュ胚で起こることを示している。翻訳遮断ネトリン−1aモルホリノを投与したfli−EGFPゼブラフィッシュ胚では、ISVは未熟成長停止(premature growth arrest)を形成または提示することができず、DLAVは存在しない。
【0063】
本発明者らは、本明細書において、ネトリン−1が内皮細胞の生存を制御し、かつ、その未結合UNC5B受容体によって誘発されるアポトーシスを遮断することによって少なくとも部分的に血管形成を促進することを示した。この機能の調節緩和(deregulation)は、本発明者らおよび第三者が観察した血管形成の調節を説明することができる(Lu et al.,2004; Wilson et al.,2006)。このことは、発生および病理学的血管形成中の細胞死調節の重要性を記載している証拠の成長線と一致する(Birdsey et al.,2008; O'Connor et al.,2000; Santoro et al.,2007)。
【0064】
ネトリン−1依存性受容体は、別の状況では栄養リガンドの利用できないことのセッティングにおいて発生する上皮細胞のアポトーシスを誘発することによって腫瘍サプレッサーとして作用することが最近提案された(Grady,2007; Mehlen and Puisieux,2006)。従って、その依存性受容体のアポトーシス促進活性の喪失は、結直腸癌におけるDCCおよびUNC5C発現の頻繁な喪失によって例示されるように、腫瘍細胞についての選択的利点を表している(Bernet et al.,2007; Mazelin et al.,2004)。相補的に、ネトリン−1発現の利得は、腫瘍細胞についての同様な選択的利点を表しており、この現象は、転移性乳癌や肺癌などの幾つかの種類の侵攻性癌で実際に観察される(Delloye et al.,2008; Fitamant et al.,2008)。本明細書に示された結果を考慮して、本発明者らは、腫瘍上皮細胞における発現のこのオートクラインネトリン−1利得が2つの付加的効果を有することがあることを提案する。第一には、報告されているように、依存性受容体によって引き起こされる細胞死を阻害することによって上皮腫瘍細胞についての選択的利点を付与する(Fitamant et al.,2008)。第二には、それは、潜在的に血管保持および/または発生にも有利であり、従って癌の進行を促進する可能性がある。従って、ネトリン−1受容体との相互作用の滴定または阻害によるネトリン−1の機能の崩壊に基づく抗癌法は、腫瘍上皮細胞だけでなく腫瘍血管形成血管をも根絶するものであり、従って癌における決定的なターゲットとしてのネトリン−1の位置を更に強化する。
【0065】
例5:DCC−5Fbnデコイペプチドは、内皮細胞に対するネトリン−1生存効果を防止する
DCCの第五の免疫グロブリンドメインに対応するデコイペプチドDCC−5Fbnでの処理は、ネトリン−1受容体多量体化を阻害することによってネトリン−1生存効果を逆転することが以前に示された(Mille et al.,2009)。
【0066】
このようなペプチドが内皮細胞に対するネトリン−1効果を遮断できるかどうかを決定するため、内皮細胞をネトリン−1のみまたはネトリン−1およびDCC−5Fbnで処理したときのカスパーゼ−3活性の相対的減少を比較した。DCC−5Fbnを添加すると、カスパーゼ−3活性に対するネトリン−1効果は1/2に減少し、従って内皮細胞に対するその生存効果を強力に減少する(図7,p<0.015)。
【0067】
文献
Alavi et al.(2003). Science 301,94-96.
Bernet et al.(2007). Gastroenterology 133,1840-1848.
Birdsey et al.(2008). Blood 111,3498.
Bredesen et al.(2005). Cell Death Differ 12,1031-1043.
Carmeliet,P.(2005). Nature 438,932-936.
Cirulli et al.(2007). Nat Rev Mol Cell Biol 8,296-306.
Delloye et al.(2008). J Natl Cancer Ins in press.
Duval et al.(2003). Angiogenesis 6,171-183.
Fitamant et al.(2008). Proc Natl Acad Sci USA,105,4850-4855.
Grady,W.M.(2007). Gastroenterology 133,2045-2049.
Lauderdale et al.(1997). Mol Cell Neurosci 9,293-313.
Lawson et al.(2002). Dev Biol 248,307-318.
Liu et al.(2004). Curr Biol 14,897-905.
Llambi et al.(2001). Embo J 20,2715-2722.
Llambi et al.(2005). Embo J 24,1192-1201.
Lu et al.(2004). Nature 432,179-186.
Ma et al.(2007). BMC Dev Biol 7,50.
Mazelin et al.(2004). Nature 431,80-84.
Mehlen et al.(2004). Apoptosis 9,37-49.
Mehlen et al.(2006). Nat Rev Cancer 6,449-458.
Mehlen et al.(1998). Nature 395,801-804.
Mille et al.(2009). Cell Death Differ. 16(10):1344-51.
Nicosia et al.(1990). Lab Invest 63,115-122.
O'Connor et al.(2000). Am J Pathol 156,393-398.
Santoro et al.(2007). Nat Genet 39,1397-1402.
Serafini et al.(1996). Cell 87,1001-1014.
Serafini et al.(1994). Cell 78,409-424.
Shohat et al.(2001). J Biol Chem 276,47460-47467.
Strahle et al.(1997). Mech Dev 62,147-160.
Tanikawa et al.(2003). Nat Cell Biol 5,216-223.
Tauszig-Delamasure et al.(2007). Proc Natl Acad Sci USA,104,13361-13366.
Wilson et al.(2006). Science 313,640.
Winn et al.(2005). J Thromb Haemost 3,1815-1824.
【特許請求の範囲】
【請求項1】
内皮細胞死を誘発することができる化合物を選択する方法であって、
a)ネトリン−1、またはその断片、および内皮細胞を含む培地を有し、
b)前記培地を、試験する化合物と接触させ、
c)前記内皮細胞死を測定し、および
d)段階c)における測定が前記化合物の存在下にて内皮細胞死の有意な誘発を示しているときに、前記化合物を選択する
段階を含んでなる、方法。
【請求項2】
内皮細胞死を誘発することができる化合物を選択する方法であって、
a)少なくとも1種類のネトリン−1受容体を発現する内皮細胞を含む培地を有し、
b)前記培地を、試験する化合物と接触させ、および
c)前記の試験する化合物のこのネトリン−1受容体のアポトーシス促進活性を促進する能力を測定し、
d)段階c)における測定が、前記化合物が、このネトリン−1受容体の、好ましくは更にネトリン−1の存在下にてこのネトリン−1受容体の、アポトーシス促進活性を有意に促進することを示しているときに、前記化合物を内皮細胞死の誘導物質として選択する
段階を含んでなる、方法。
【請求項3】
段階c)の測定が、試験する化合物の細胞死関連タンパク質キナーゼ(DAPK)のキナーゼ活性の活性化を誘発する能力を測定することにあり、および
d)段階c)における測定が、前記化合物の存在下にて、好ましくは更にネトリン−1の存在下にて細胞死関連タンパク質キナーゼ(DAPK)のキナーゼ活性の有意な活性化を示しているときに、前記化合物を内皮細胞死の誘導物質として選択する、
請求項2に記載の内皮細胞死を誘発することができる化合物の選択方法。
【請求項4】
段階c)の測定が、試験する化合物のDAPKリン酸化を阻害する能力を測定することにあり、および
d)段階c)における測定が、前記化合物の存在下にて、好ましくは更にネトリン−1の存在下にてDAPKリン酸化の有意な阻害を示しているときに、前記化合物を内皮細胞死の誘導物質として選択する、
請求項2に記載の内皮細胞死を誘発することができる化合物の選択方法。
【請求項5】
内皮細胞死を誘発することができる化合物の選択方法であって、
a)少なくとも1種類のネトリン−1受容体を発現する内皮細胞を含む培地を有し、かつ前記ネトリン−1受容体細胞内ドメインがネトリン−1の存在下にて二量体化または多量体化することができ、
b)前記培地を、試験する化合物と接触させ、培地が更にネトリン−1、またはネトリン−1受容体の細胞外ドメインと相互作用することができるその断片を含み、
c)前記ネトリン−1受容体細胞内ドメインの二量体化または多量体化が前記の試験する化合物の存在下で阻害されるかどうかを決定し、および
d)段階c)における決定が、前記内皮細胞の前記ネトリン−1受容体の細胞内ドメインの二量体化または多量体化を有意に阻害することを示しているときに、前記化合物を内皮細胞死の誘導物質として選択する
段階を含んでなる、方法。
【請求項6】
血管または新生血管の死を誘発することができる化合物を選択するための、請求項1〜5のいずれか一項に記載の方法。
【請求項7】
段階a)において、前記内皮細胞が、DCC、UNC5A、UNC5B、UNC5CおよびUNC5E、ネオゲニンおよびアデノシンA2bの群から選択される少なくとも1種類のネトリン−1受容体、好ましくはUNC5Bを発現する内皮細胞である、請求項1〜6のいずれか一項に記載の方法。
【請求項8】
段階a)において、前記内皮細胞が、ネトリン−1受容体UNC5Bを発現する内皮細胞である、請求項7に記載の方法。
【請求項9】
段階a)において、前記内皮細胞が、ヒト臍帯静脈内皮細胞(HUVEC)およびヒト臍帯動脈内皮細胞(HUAEC)からなる群から選択される内皮細胞である、請求項1〜8のいずれか一項に記載の方法。
【請求項10】
ネトリン−1と前記ネトリン−1受容体との相互作用を特異的に阻害することができるネトリン−1受容体またはその断片の細胞外ドメインを含んでなる化合物、
ネトリン−1またはネトリン−1受容体に対して特異的な、特に前記ネトリン−1受容体の細胞外ドメインまたは前記ネトリン−1受容体の細胞外ドメインと相互作用することができるネトリン−1断片に対するモノクローナルまたはポリクローナル抗体、および
ネトリン−1タンパク質をコードする核酸に特異的なアンチセンスおよびRNAi(干渉RNA)オリゴヌクレオチドからなる群から選択されるネトリン−1発現を阻害することができる化合物
からなる群から選択される、内皮細胞死の誘導物質としての、化合物。
【請求項11】
血管または新生血管の死を誘発する目的で使用するための、請求項10に記載の化合物。
【請求項12】
腫瘍の血管または新生血管の死を誘発する目的で使用するための、請求項10または11に記載の化合物。
【請求項13】
前記腫瘍の腫瘍細胞がネトリン−1を発現または過剰発現する、請求項12に記載の化合物。
【請求項14】
内皮細胞、好ましくは血管または新生血管由来の内皮細胞の死を誘発することができる化合物を選択するためのキットであって、
ネトリン−1タンパク質またはネトリン−1受容体タンパク質と特異的に相互作用して結合対を形成することができるその断片、および
ネトリン−1受容体、好ましくはDCC、UNC5A、UNC5B、UNC5CおよびUNC5D、ネオゲニンおよびアデノシンA2bからなる群から選択され、更に好ましくはネトリン−1受容体UNC5Bを発現する内皮細胞
を含んでなる、キット。
【請求項15】
前記内皮細胞がHUVECおよびHUAEC細胞の群から選択される、請求項14に記載のキット。
【請求項1】
内皮細胞死を誘発することができる化合物を選択する方法であって、
a)ネトリン−1、またはその断片、および内皮細胞を含む培地を有し、
b)前記培地を、試験する化合物と接触させ、
c)前記内皮細胞死を測定し、および
d)段階c)における測定が前記化合物の存在下にて内皮細胞死の有意な誘発を示しているときに、前記化合物を選択する
段階を含んでなる、方法。
【請求項2】
内皮細胞死を誘発することができる化合物を選択する方法であって、
a)少なくとも1種類のネトリン−1受容体を発現する内皮細胞を含む培地を有し、
b)前記培地を、試験する化合物と接触させ、および
c)前記の試験する化合物のこのネトリン−1受容体のアポトーシス促進活性を促進する能力を測定し、
d)段階c)における測定が、前記化合物が、このネトリン−1受容体の、好ましくは更にネトリン−1の存在下にてこのネトリン−1受容体の、アポトーシス促進活性を有意に促進することを示しているときに、前記化合物を内皮細胞死の誘導物質として選択する
段階を含んでなる、方法。
【請求項3】
段階c)の測定が、試験する化合物の細胞死関連タンパク質キナーゼ(DAPK)のキナーゼ活性の活性化を誘発する能力を測定することにあり、および
d)段階c)における測定が、前記化合物の存在下にて、好ましくは更にネトリン−1の存在下にて細胞死関連タンパク質キナーゼ(DAPK)のキナーゼ活性の有意な活性化を示しているときに、前記化合物を内皮細胞死の誘導物質として選択する、
請求項2に記載の内皮細胞死を誘発することができる化合物の選択方法。
【請求項4】
段階c)の測定が、試験する化合物のDAPKリン酸化を阻害する能力を測定することにあり、および
d)段階c)における測定が、前記化合物の存在下にて、好ましくは更にネトリン−1の存在下にてDAPKリン酸化の有意な阻害を示しているときに、前記化合物を内皮細胞死の誘導物質として選択する、
請求項2に記載の内皮細胞死を誘発することができる化合物の選択方法。
【請求項5】
内皮細胞死を誘発することができる化合物の選択方法であって、
a)少なくとも1種類のネトリン−1受容体を発現する内皮細胞を含む培地を有し、かつ前記ネトリン−1受容体細胞内ドメインがネトリン−1の存在下にて二量体化または多量体化することができ、
b)前記培地を、試験する化合物と接触させ、培地が更にネトリン−1、またはネトリン−1受容体の細胞外ドメインと相互作用することができるその断片を含み、
c)前記ネトリン−1受容体細胞内ドメインの二量体化または多量体化が前記の試験する化合物の存在下で阻害されるかどうかを決定し、および
d)段階c)における決定が、前記内皮細胞の前記ネトリン−1受容体の細胞内ドメインの二量体化または多量体化を有意に阻害することを示しているときに、前記化合物を内皮細胞死の誘導物質として選択する
段階を含んでなる、方法。
【請求項6】
血管または新生血管の死を誘発することができる化合物を選択するための、請求項1〜5のいずれか一項に記載の方法。
【請求項7】
段階a)において、前記内皮細胞が、DCC、UNC5A、UNC5B、UNC5CおよびUNC5E、ネオゲニンおよびアデノシンA2bの群から選択される少なくとも1種類のネトリン−1受容体、好ましくはUNC5Bを発現する内皮細胞である、請求項1〜6のいずれか一項に記載の方法。
【請求項8】
段階a)において、前記内皮細胞が、ネトリン−1受容体UNC5Bを発現する内皮細胞である、請求項7に記載の方法。
【請求項9】
段階a)において、前記内皮細胞が、ヒト臍帯静脈内皮細胞(HUVEC)およびヒト臍帯動脈内皮細胞(HUAEC)からなる群から選択される内皮細胞である、請求項1〜8のいずれか一項に記載の方法。
【請求項10】
ネトリン−1と前記ネトリン−1受容体との相互作用を特異的に阻害することができるネトリン−1受容体またはその断片の細胞外ドメインを含んでなる化合物、
ネトリン−1またはネトリン−1受容体に対して特異的な、特に前記ネトリン−1受容体の細胞外ドメインまたは前記ネトリン−1受容体の細胞外ドメインと相互作用することができるネトリン−1断片に対するモノクローナルまたはポリクローナル抗体、および
ネトリン−1タンパク質をコードする核酸に特異的なアンチセンスおよびRNAi(干渉RNA)オリゴヌクレオチドからなる群から選択されるネトリン−1発現を阻害することができる化合物
からなる群から選択される、内皮細胞死の誘導物質としての、化合物。
【請求項11】
血管または新生血管の死を誘発する目的で使用するための、請求項10に記載の化合物。
【請求項12】
腫瘍の血管または新生血管の死を誘発する目的で使用するための、請求項10または11に記載の化合物。
【請求項13】
前記腫瘍の腫瘍細胞がネトリン−1を発現または過剰発現する、請求項12に記載の化合物。
【請求項14】
内皮細胞、好ましくは血管または新生血管由来の内皮細胞の死を誘発することができる化合物を選択するためのキットであって、
ネトリン−1タンパク質またはネトリン−1受容体タンパク質と特異的に相互作用して結合対を形成することができるその断片、および
ネトリン−1受容体、好ましくはDCC、UNC5A、UNC5B、UNC5CおよびUNC5D、ネオゲニンおよびアデノシンA2bからなる群から選択され、更に好ましくはネトリン−1受容体UNC5Bを発現する内皮細胞
を含んでなる、キット。
【請求項15】
前記内皮細胞がHUVECおよびHUAEC細胞の群から選択される、請求項14に記載のキット。
【図5A】



【図1A】

【図1B】

【図1C】

【図1D】

【図1E】

【図1F】

【図1G】

【図1H】

【図2A】

【図2B】

【図2C】

【図2D】
【図2E】

【図2F】

【図3A】

【図3B】

【図3C】

【図4A】

【図4B】

【図4C】

【図4D】

【図4E】

【図4F】

【図4G】

【図5B】

【図5C】

【図6A】

【図6B】

【図6C】

【図6D】

【図7】

【図1A】
【図1B】
【図1C】
【図1D】
【図1E】
【図1F】
【図1G】
【図1H】
【図2A】
【図2B】
【図2C】
【図2D】
【図2E】
【図2F】
【図3A】
【図3B】
【図3C】
【図4A】
【図4B】
【図4C】
【図4D】
【図4E】
【図4F】
【図4G】
【図5B】
【図5C】
【図6A】
【図6B】
【図6C】
【図6D】
【図7】
【公表番号】特表2012−514464(P2012−514464A)
【公表日】平成24年6月28日(2012.6.28)
【国際特許分類】
【出願番号】特願2011−544884(P2011−544884)
【出願日】平成22年1月11日(2010.1.11)
【国際出願番号】PCT/EP2010/050233
【国際公開番号】WO2010/079230
【国際公開日】平成22年7月15日(2010.7.15)
【公序良俗違反の表示】
(特許庁注:以下のものは登録商標)
1.セロテープ
【出願人】(594016872)サントル、ナショナール、ド、ラ、ルシェルシュ、シアンティフィク、(セーエヌエルエス) (83)
【出願人】(504217063)
【出願人】(510307222)
【氏名又は名称原語表記】ECOLE NORMALE SUPERIEURE DE LYON
【Fターム(参考)】
【公表日】平成24年6月28日(2012.6.28)
【国際特許分類】
【出願日】平成22年1月11日(2010.1.11)
【国際出願番号】PCT/EP2010/050233
【国際公開番号】WO2010/079230
【国際公開日】平成22年7月15日(2010.7.15)
【公序良俗違反の表示】
(特許庁注:以下のものは登録商標)
1.セロテープ
【出願人】(594016872)サントル、ナショナール、ド、ラ、ルシェルシュ、シアンティフィク、(セーエヌエルエス) (83)
【出願人】(504217063)
【出願人】(510307222)
【氏名又は名称原語表記】ECOLE NORMALE SUPERIEURE DE LYON
【Fターム(参考)】
[ Back to top ]