
Fターム[4B065CA13]の内容
微生物、その培養処理 (127,014) | 微生物の生産物質;用途 (34,848) | 有機化合物 (9,808) | 酸素原子(O)を含むもの (1,098) | 油脂、高級脂肪酸(C7以上) (179)
Fターム[4B065CA13]に分類される特許
161 - 179 / 179
脂質含量の高い真核微生物及び該微生物を用いた脂質の製造方法
【課題】トリアシルグリセロールなどの脂質を高含量で含む真核微生物変異株を取得するとともに、これを用いて脂質を製造する。
【課題解決手段】
トランスポゾンを挿入させた変異ライブラリーを用いて、真核微生物を変異させ、脂質の蓄積含量の高い変異株をスクリーニングし、その変異株のトランスポゾン挿入位置の同定により変異遺伝子を決定し、それらの遺伝子の破壊株を作成し、これを用いて脂質を製造する。
(もっと読む)
不飽和脂肪酸の製造方法
本発明は、鱗翅目昆虫からのデサチュラーゼの組換え発現によって、不飽和脂肪酸、好ましくは共役リノール酸(CLA)などの共役多価不飽和脂肪酸を製造する方法に関するものである。発現は好ましくは、植物、酵母、真菌およびソウ類の群から選択される生物で行う。さらに本発明によれば、鱗翅目昆虫からのデサチュラーゼの組換え発現のための組換え発現カセット、ならびにそれによって形質転換されたトランスジェニック生物がある。 (もっと読む)
オレフィン性もしくはアセチレン性基質を酵母によって結合させることによるポリ不飽和脂肪酸の製造方法
本発明は、ポリ不飽和脂肪酸の製造方法であって、炭素原子数が13〜18の鎖長を有するオレフィン系もしくはアセチレン系脂肪酸(基質)を、該基質の9位において配置Zを有する新結合を形成させるビール酵母菌イーストW303aまたはΔElo1のハプロイドもしくはディプロイド野生菌株に取込ませることによる該製造方法に関する。新たな不飽和化は、被処理物中の元の不飽和結合が配置Eを有しているか、またはアセチレン結合であるときにのみおこなわれる。さらに、元の結合が配置Zを有するときには、基質は無変化で回収される。 (もっと読む)
ホスホリパーゼ、それらをコードする核酸並びにそれらの製造方法及び使用方法
本発明は、ホスホリパーゼ活性(例えばホスホリパーゼA、B、C及びD活性、パタチン活性、ホスファチジン酸ホスファターゼ(PAP)及び/又は脂質アシルヒドロラーゼ(LAH)活性を含む)を有する新規なポリペプチド、前記ポリペプチドをコードする核酸、前記ポリペプチドと結合する抗体を提供する。これらホスホリパーゼの使用を含む工業的方法(例えば油の脱ガム)及び製品もまた提供される。  (もっと読む)
(もっと読む)
生物学的細胞における多価不飽和脂肪酸の含有量を変更するための組成物および方法
本発明は、組成物(例えば、恒常的に活性な、または組織に特異的なプロモーターもしくは他の制御配列に任意選択で操作可能に結合している、fat−1をコードする核酸、およびその核酸または生物学的に活性なその変種を含む薬学的に許容できる製剤)、ならびに動物細胞(すなわち、シー・エレガンス(C.elegans)の細胞以外の細胞、例えば、筋肉細胞、ニューロン(末梢神経系でも中枢神経系でも)、脂肪細胞、内皮細胞、および癌細胞など、鳥類または魚類の細胞)におけるPUFAの含有量を効果的に変更するために用いることができる方法を特徴とする。組成物および方法は、少なくとも1つの最適化されたコドンを含むように変更されたfat−1遺伝子を含む。改変された細胞は、インビボでもイクスビボ(例えば、組織培養物中で)でも、それらを含むトランスジェニック動物(特に、魚類および鳥類)、ならびにそれらの動物から得られた食品(例えば、肉、または動物の他の可食部(例えば、肝臓、腎臓、またはスイートブレッド))も、本発明の範囲内である。 (もっと読む)
ゼニゴケ由来の不飽和脂肪酸合成系酵素遺伝子及びその利用
同一種のゼニゴケから、Δ5脂肪酸不飽和化酵素遺伝子、Δ6脂肪酸不飽和化酵素遺伝子及びΔ6脂肪酸鎖長延長酵素遺伝子を単離する。これらの遺伝子を高等植物に導入することにより、アラキドン酸やエイコサペンタエン酸(EPA)を生産し得る形質転換植物体を取得する。 (もっと読む)
アラキドン酸を含有する植物体およびその利用
アラキドン酸を含有するアラキドン酸植物体やダイズおよびその利用法を提供する。アラキドン酸生合成に関与する脂肪酸合成酵素遺伝子を植物体に導入し、アラキドン酸を生産させるアラキドン酸生産工程を含む植物体の生産方法を用いて得られる油脂原料植物体によれば、簡便にアラキドン酸を含有する植物体やダイズを取得することができる。それゆえ、アラキドン酸を大量かつ安価に取得することができる。 (もっと読む)
飽和脂肪酸のレベルが低減されたトランスジェニック植物およびその製造方法
【課題】 低減されたレベルの飽和脂肪酸を含有する種油を提供することのできる植物を提供する。
【解決手段】 本発明は、種油中の飽和脂肪酸のレベルが低減されたトランスジェニック植物およびその製造方法を提供する。この方法により発生したトランスジェニック植物は種油中に低減されたレベルの飽和脂肪酸を含有するが、この低減されたレベルは小胞体保留および回収シグナル配列に機能的に連結された原核生物のデルタ-9不飽和化酵素(すなわち飽和脂肪酸のデルタ-9位にcis二重結合を導入する酵素)の発現に起因するものである。本発明の一つの例はシアノバクテリアであるAnacystis nidulans由来の異種デルタ-9不飽和化酵素を発現する植物であるが、この酵素はKKSS(配列番号5)小胞体保留および回収シグナル配列と機能的に連結されており、脂質結合型16:0および18:0脂肪酸をそれぞれ16:1および18:1へと変換するものである。
(もっと読む)
油性植物および酵母中の多価不飽和脂肪酸レベルを変更するのに適したΔ15デサチュラーゼ
本発明は、リノール酸(18:2、LA)からα−リノレン酸(18:3、ALA)への転換を触媒できる真菌Δ15脂肪酸デサチュラーゼに関する。デサチュラーゼをコードする核酸配列、それにハイブリダイズする核酸配列、デサチュラーゼ遺伝子を含んでなるDNAコンストラクト、および増大したレベルのデサチュラーゼを発現する組換え宿主植物および微生物について記載されている。15脂肪酸デサチュラーゼの過剰発現によって特定のω−3およびω−6脂肪酸の生成の増大方法についてもまた本明細書に記載されている。 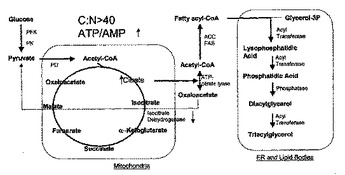 (もっと読む)
(もっと読む)
NADPH消費生合成経路のために最適化された微生物菌株
【課題】本発明は、NADPH消費生合成経路を有する、分子のバイオトランスフォーメーションによる生成のために最適化された微生物菌株に関する。
【解決手段】本発明の菌株はNADPHを消費するバイオトランスフォーメーションプロセスで利用できる。それらの菌株は、NADPHを酸化する1つまたは複数の活性が制限されていることを特徴とする。
(もっと読む)
ヤブレツボカビ属の微生物の培養方法
本発明は、ヤブレツボカビ(Thraustochytriales)目の微生物を炭酸カルシウムを用いてpH安定化されており、3〜15g/LのCaCO3を含む発酵培地において培養し、その後PUFAを微生物及び/または培地から単離することによるPUFAを産生するための最適方法に関する。本発明は特に、異なるCaCO3含量を有する新規な最適化培地に関する。適当量のCaCO3を使用することにより、この方法はpHをコントロールすることなくヤブレツボカビ微生物を発酵させることができるように発酵中かなり簡素化され得、大量のDHAをバイオマス中の高い油含量で得ることができ、これによりPUFA産生が実質的に改良され、かなり簡素化される。 (もっと読む)
最適化低塩培地を用いるヤブレツボカビ属の微生物の培養方法
本発明は、ヤブレツボカビ目の微生物をナトリウム塩及び塩化物を添加せずに、総塩含量が3.5g/L未満(海水塩含量の10%未満に相当)である低塩培地において培養し、PUFAを微生物及び/または培地から単離する前記微生物を培養するための最適化方法に関する。本発明は特に、総塩含量を実質的に低減させた、特にNaCl含量を著しく低減させた新規な最適化培地に関する。PUFAの産生は、Na+イオンとCl−イオンの全重量比が1.75g/Lを越えない培地組成として各種塩の新規組合せを用いることにより実質的に改善され、十分に簡素化され得る。更に、培地は好ましくは添加したナトリウム塩及び塩化物を全く含まず、塩を含有する廃水により生ずる環境ダメージを防止するのを助ける。 (もっと読む)
海洋微生物のバイオマス及び/又はそのバイオマスの成分の収量を高める方法
本発明は、Y g/Lの細胞乾燥物質(CDM)(ここで、Yは、100〜300g/Lの範囲で存在する)で、好気性条件下で発酵槽において栄養要求性海洋微生物を連続して培養するための最適化された方法を提供し、ここで培養液1L当たり(Y×h)g(hは、1.1〜3.0の範囲である)の量で徐々に添加され、そして20〜100時間の滞留時間をもって、炭素源を含んで成る培養培地において、前記栄養要求性海洋微生物を培養することを含んで成る。本発明は、細胞脂質、特にポリエン酸の高い生産性を維持する。 (もっと読む)
脂質生産菌の育種方法
抗生物質によらず、モルティエレラ属に属する脂質生産菌の育種を有効かつ効率的に行うことができる育種方法を提供する。 モルティエレラ(Mortierella)属に属する脂質生産菌の栄養要求性株、例えばウラシル要求性株を宿主とし、形質転換工程を行う。具体的には、マーカー遺伝子としてこの栄養要求性を相補する遺伝子を用いて、当該マーカー遺伝子を宿主に導入する(遺伝子導入段階)。その後、上記宿主の栄養要求性の回復をマーカーとして形質転換株を選抜する(選抜段階)。上記脂質生産菌としては、例えばモルティエレラ アルピナ(M.alpina)を挙げることができる。 (もっと読む)
植物中の油レベルの上昇
本発明は、HOI001 GBSS対立遺伝子の発現によって、トウモロコシの穀穀粒組織中の油レベルを増加させるための方法を提供する。また、本発明は、HOI001 GBSSポリペプチドをコードする単離された核酸分子を提供する。 (もっと読む)
フィードバック耐性メバロン酸キナーゼ
本発明は、フィードバック阻害に対する感受性の低い修飾メバロン酸キナーゼおよびそれをコードするポリヌクレオチドに関する。本発明はさらにこれらポリヌクレオチドを含むベクターおよびそのようなベクターを含む宿主細胞に関係する。本発明は、修飾酵素の製法およびイソプレノイド化合物の製法を提供する。 (もっと読む)
油性酵母菌における多不飽和脂肪酸生産のためのコドン最適化遺伝子
本発明は、リノール酸(LA)からγ−リノレン酸(GLA)、α−リノール酸(ALA)からステアリドン酸(STA)、GLAからジホモ−γ−リノール酸(DGLA)、STAからエイコサテトラエン酸(ETA)、DGLAからETA、エイコサペンタエン酸(EPA)からドコサペンタエン酸(DPA)、およびアラキドン酸(ARA)からEPAへの転換を触媒できる脂肪酸デサチュラーゼおよび鎖長延長酵素に関する。コドン最適化デサチュラーゼおよび鎖長延長酵素をコードする核酸配列、それにハイブリダイズする核酸配列、コドン最適化デサチュラーゼまたは鎖長延長酵素を含んでなるDNA構築物、および増大するレベルのデサチュラーゼまたは鎖長延長酵素を発現する組換え宿主微生物について記載される。 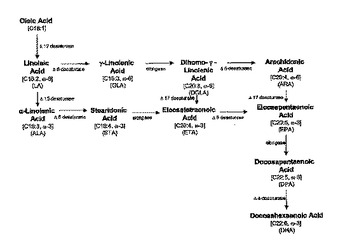 (もっと読む)
(もっと読む)
油性酵母菌における多不飽和脂肪酸の生成
本発明は、油性酵母菌におけるω−3および/またはω−6脂肪酸の生成方法に関する。したがってARAおよびEPAの合成のために、リノール酸(LA)からγ−リノレン酸(GLA)、α−リノール酸(ALA)からステアリドン酸(STA)、GLAからジホモ−γ−リノール酸(DGLA)、STAからエイコサテトラエン酸(ETA)、DGLAからアラキドン酸(ARA)、ETAからエイコサペンタエン酸(EPA)、DGLAからETA、EPAからドコサペンタエン酸(DPA)、およびARAからEPAへの転換を触媒できるデサチュラーゼおよびエロンガーゼが、ヤロウイア(Yarrowia)のゲノムに導入された。
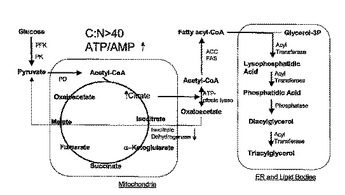 (もっと読む)
(もっと読む)
PUFAポリケチドシンターゼ系およびこれらの使用
本発明は概して、高度不飽和脂肪酸(PUFA)ポリケチドシンターゼ(PKS)系に関し、これらの相同体に関し、このようなPUFA PKS系の生物学的に活性なドメインをコードする単離された核酸分子および組換え核酸分子に関し、PUFA PKS系を含む遺伝子改変された生物に関し、関心対象の生理活性分子の産生のためにこのような系を作製および使用する方法に関し、かつこのようなPUFA PKS系を有する新たな非細菌性および細菌性の微生物を同定するための新規の方法に関する。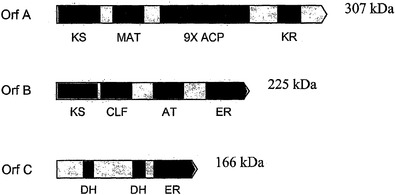
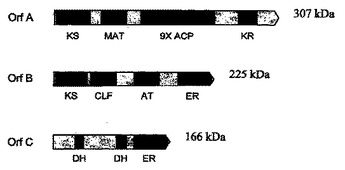 (もっと読む)
(もっと読む)
161 - 179 / 179
[ Back to top ]