
ヒトおよび哺乳動物の幹細胞由来神経生存因子

【課題】ヒト、ラット、マウスの幹細胞由来神経生存因子ポリペプチド(SDNSF)を含有する中枢神経損傷治療剤、SDNSFを測定する方法、SDNSFを測定する試薬およびSDNSFを用いたスクリーニング方法を提供する。
【解決手段】実質的に純粋な形である配列番号4、8または12で示されるアミノ酸配列からなるポリペプチドの測定方法。実質的に純粋な形である配列番号4、8または12で示されるアミノ酸配列からなるポリペプチドに対するポリクローナルまたはモノクローナル抗体を用いることを特徴とする前記ポリペプチドの免疫化学的測定方法。実質的に純粋な形である配列番号4、8または12で示されるアミノ酸配列からなるポリペプチドを用いることを特徴とする該ポリペプチドに対するアンタゴニストまたはアゴニストとしての活性を有する化合物をスクリーニングする方法。
【解決手段】実質的に純粋な形である配列番号4、8または12で示されるアミノ酸配列からなるポリペプチドの測定方法。実質的に純粋な形である配列番号4、8または12で示されるアミノ酸配列からなるポリペプチドに対するポリクローナルまたはモノクローナル抗体を用いることを特徴とする前記ポリペプチドの免疫化学的測定方法。実質的に純粋な形である配列番号4、8または12で示されるアミノ酸配列からなるポリペプチドを用いることを特徴とする該ポリペプチドに対するアンタゴニストまたはアゴニストとしての活性を有する化合物をスクリーニングする方法。
【発明の詳細な説明】
【技術分野】
【0001】
本発明はヒト、ラット、マウスの幹細胞由来神経生存因子(Stem cell Derived-Neuron Survival Factor、以下、SDNSFと省略する。)に関する。
【0002】
さらに詳しく言えば、ヒト、ラット、マウスのSDNSF、それらの製造方法、SDNSFをコードするcDNA、そのcDNAからなるベクター、そのベクターで形質転換された宿主細胞、SDNSFの抗体、SDNSFまたは抗体を含有する薬学的組成物、SDNSFを測定する方法、SDNSFを測定する試薬およびSDNSFを用いたスクリーニング方法に関する。
【背景技術】
【0003】
成体脳では新しいニューロンは生まれないというのが長い間の定説であり、脳梗塞や脳出血、脊髄損傷などによる中枢神経の損傷、パーキンソン病や筋萎縮性側索硬化症(ALS)などの神経変性疾患などでは、神経細胞の細胞死により運動機能が失われ、いったん失われた機能の回復は難しいとされていた。しかし近年、ヒトやサルなどの高等哺乳動物の成体脳(海馬、大脳皮質連合野、側脳室)でもニューロンが新生することが見出され、これらの領域で新しく生まれるニューロンが神経幹細胞から発生することも示された。また、神経幹細胞が高齢者の脳内にも存在し、神経細胞に分化しうることも証明された。これらのことは、脳の再生治療が細胞を移植する細胞治療に限定されず、タンパク質医薬、薬物あるいは遺伝子治療などの方法により直接内在性の幹細胞を活性化する治療が可能であることを示唆している。
【0004】
従来、ある特定のポリペプチドまたはそれをコードするcDNAを得ようとする場合、組織や細胞培養液中に目的とする生物活性を確認し、次いでポリペプチドの単離精製を経て、遺伝子をクローニングするという方法、あるいはその生物活性を指標として遺伝子を発現クローニングする方法が一般的に用いられてきた。しかし、生体内生理活性ポリペプチドは、多様な生物活性を有している場合が多いので、あるひとつの活性を指標にして遺伝子をクローニングした結果、それが既知のポリペプチドと同一であることが後になって判明するという事例が増えている。また、微量しか産生されなかったり、特別な生理条件でのみ発現する因子も多く、そのことが単離、精製および生物活性の確認を困難なものとしている。
【発明の開示】
【0005】
本発明者らは、中枢神経の損傷または神経変性に係わる疾患の治療、診断、脳腫瘍の診断あるいは研究上有益な新規な因子(ポリペプチド)、特に分泌シグナルを有する分泌蛋白質および膜蛋白質に着目してそれを見出すべく、鋭意検討を行った。その結果、神経幹細胞に係わる新規なペプチド分子を単離し、上記のような再生治療に使用すること、および神経細胞の特異的マーカーとなることを見出した。
【0006】
本発明者らは、これまで造血系や免疫系で働く増殖分化因子の遺伝子のクローニングを研究してきた。そして、増殖分化因子(例えば、各種サイトカイン等)のような分泌蛋白質やそのレセプターのような膜蛋白質(以下、これらをまとめて分泌蛋白質等と呼ぶ。)の大部分がそのN末端にシグナルペプチドと呼ばれる配列を有していることに着目して、シグナルペプチドをコードする遺伝子を効率的かつ選択的にクローニングする方法を鋭意検討した。その結果、動物細胞を用いて、シグナルペプチドの有無を簡単に検索できる方法(シグナルシークエンストラップ(SST)法)を見出した(日本国特許第2,879,303号参照)。さらに同じ概念のもとに、酵母を用いてさらに大量かつ簡便にシグナルペプチドをコードする遺伝子を単離する方法(酵母SST法)も開発された(米国特許第5,536,637号)。
【0007】
本方法を用いて、成体ラット海馬由来神経幹細胞が産生している新規な分泌蛋白質、およびそれをコードするcDNAを同定することに成功し、その情報を基に全長cDNAを成体ラット海馬由来神経幹細胞より見出した。
【0008】
本ポリペプチドが、後に詳述するように、脳の一部のニューロン(海馬神経初代培養細胞および海馬由来幹細胞)に対し、生存支持活性を持つことを確認し、本発明を完成した。本ポリペプチドは、機能が特定された有用な因子である。
【0009】
本発明が提供するcDNA配列は、配列番号1または2で示されるラットSDNSFクローンとして同定され、成体ラット海馬由来神経幹細胞から作製したcDNAライブラリーより、酵母SST法を使用して得た情報をもとに単離された。配列番号1で示されるラットSDNSFクローンは分泌蛋白質(ここではラットSDNSF蛋白として表される。)をコードする完全なcDNA配列を含む全長鎖cDNAである。
【0010】
本発明が提供するcDNA配列は、配列番号5または6で示されるヒトSDNSFクローンとして同定され、成体ラット海馬由来神経幹細胞から作製したcDNAライブラリーより、酵母SST法を使用して得た情報をもとに単離された。配列番号5で示されるヒトSDNSFクローンは分泌蛋白質(ここではヒトSDNSF蛋白として表される。)をコードする完全なcDNA配列を含む全長鎖cDNAである。
【0011】
本発明が提供する配列番号9または10で示されるcDNA配列は、マウスSDNSFクローンとして同定され、成体ラット海馬由来神経幹細胞から作製したcDNAライブラリーより、酵母SST法を使用して得た情報をもとに単離された。配列番号9で示されるマウスSDNSFクローンは分泌蛋白質(ここではマウスSDNSF蛋白として表される。)をコードする完全なcDNA配列を含む全長鎖cDNAである。
【0012】
核酸配列データベースに登録されている既知の核酸配列に対してGenBankおよびNCBIを用いたBLASTN、FASTAおよびUNIGENEなどにより、またアミノ酸配列データベースに登録されている既知のポリペプチドのアミノ酸配列に対してBLASTP、Fly Database、SwissProtなどにより検索をした結果、本発明ポリペプチドであるラットSDNSFおよびそれらをコードする核酸配列と一致する配列はなかった。このことから、本発明のポリペプチドは、新規の分泌蛋白質であることが判明した。
【0013】
また、本発明者らは脳の一部のニューロンの生存支持活性を持つこと、分泌蛋白でありながらEFハンドモチーフを持ち、細胞外または分泌経路のオルガネラのカルシウムで調節を受けるサイトカインである可能性があること、および既知のいずれのニューロトロフィンとも相同性を有さないことから、既知のニューロトロフィン遺伝子欠損マウス解析からそれらへの依存性が知られている交感神経、知覚神経、脊髄運動神経核の神経、基底核コリン作動性神経以外のニューロンの形成や生存を支持するサイトカインである可能性と同時に、これまで原因不明であった神経変性疾患の病因解明および治療につながるものと考えられる。
【0014】
本発明は、
1.実質的に純粋な形である配列番号4、8または12で示されるアミノ酸配列からなるポリペプチド、そのホモローグ、そのフラグメントまたはそのフラグメントのホモローグ、
2.配列番号4、8または12で示されるアミノ酸配列からなる前記第1項記載のポリペプチド、
3.前記第1項または第2項記載のポリペプチドをコードするcDNA、
4.配列番号1、2、5、6、9または10で示される塩基配列を有する前記第3項記載のcDNA、またはその配列に選択的にハイブリダイズするフラグメント、
5.前記第3項または第4項記載のcDNAからなる複製または発現ベクター、
6.前記第5項記載の複製または発現ベクターで形質転換された宿主細胞、
7.前記第1項または第2項記載のポリペプチドを発現させるための条件下で前記第6項記載の宿主細胞を培養することを特徴とする該ポリペプチドの製造方法、
8.前記第1項または第2項記載のポリペプチドに対するポリクローナルまたはモノクローナル抗体、
9.前記第1項または第2項記載のポリペプチドまたは前記第8項記載の抗体および薬学的に許容される賦形剤および/または担体を含有することを特徴とする薬学的組成物、
10.前記第1項または第2項記載のポリペプチドまたは前記第8項記載の抗体および薬学的に許容される賦形剤および/または担体を含有することを特徴とする神経変性が係わる疾患の治療に有効な薬学的組成物、
11.神経変性が係わる疾患が、脳梗塞や脳出血、脊髄損傷などによる中枢神経の損傷である前記第10項記載の薬学的組成物、
12.前記第1項または第2項記載のポリペプチドの測定方法、
13.前記第8項記載の抗体を用いることを特徴とする前記第1項または第2項記載のポリペプチドの免疫化学的測定方法、
14.前記第12項または第13項記載の方法を用いることを特徴とする前記第1項または第2項記載のポリペプチドを検出する試薬、
15.前記第12項または第13項記載の方法を用いることを特徴とする腫瘍を検査する試薬、
16.腫瘍が脳腫瘍である前記第15項記載の試薬、
17.前記第1項または第2項記載のポリペプチドを用いることを特徴とする該ポリペプチドに対するアンタゴニストまたはアゴニストとしての活性を有する化合物をスクリーニングする方法、
18.前記第1項または第2項記載のポリペプチドを有効成分とする中枢神経損傷治療剤、
19.中枢神経損傷が脳梗塞による中枢神経損傷である前記第19項記載の試薬、
20.中枢神経損傷が脳出血による中枢神経損傷である前記第19項記載の試薬、および
21.中枢神経損傷が脊髄損傷による中枢神経損傷である前記第19項記載の試薬に関する。
【発明を実施するための最良の形態】
【0015】
実質的に純粋な形である配列番号4、8または12で示されるアミノ酸配列からなるポリペプチドとは、一般に、生産時のポリペプチドの90%以上、例えば、95、98または99%が配列番号4、8または12で示されるアミノ酸配列からなるポリペプチドであることを意味する。
【0016】
配列番号4、8または12で示されるアミノ酸配列からなるポリペプチドのホモローグとは、一般に少なくとも20個、好ましくは少なくとも30個、例えば40、60、80または100個の連続したアミノ酸領域で、少なくとも70%、好ましくは少なくとも80または90%、より好ましくは95%以上相同性であるものであり、そのようなホモローグは、以後本発明のポリペプチドとして記載される。
【0017】
さらに、配列番号4、8または12で示されるアミノ酸配列からなるポリペプチドのフラグメント、またはそれらのホモローグのフラグメントとは、少なくとも10アミノ酸、好ましくは少なくとも15アミノ酸、例えば20、25、30、40、50または60アミノ酸部分を意味する。
【0018】
配列番号1、2、5、6、9または10で示される塩基配列からなるcDNAに選択的にハイブリダイズするcDNAとは、一般に、少なくとも20個、好ましくは少なくとも30個、例えば40、60、80または100個の連続した塩基配列領域で、少なくとも70%、好ましくは少なくとも80または90%、より好ましくは95%以上相同性であるものであり、そのようなcDNAは、以後本発明のcDNAとして記載される。
【0019】
配列番号1、2、5、6、9または12で示される塩基配列からなるcDNAのフラグメントとは、少なくとも10塩基、好ましくは少なくとも15塩基、例えば20、25、30または40塩基部分を意味し、そのようなフラグメントも本発明のcDNAに含まれる。
【0020】
本発明のSDNSF蛋白は、脳腫瘍のうち未分化なニューロブラストーマとグリオブラストーマ細胞株から大量に分泌されるが、分化したグリオーマからは分泌されないことと、血中や髄液中に分泌されるニューロブラストーマとグリオブラストーマ脳腫瘍マーカーは無いので、SDNSF蛋白が、血中で検出可能な初の未分化脳腫瘍マーカーとすることができる。
【0021】
さらに、本発明には、本発明のcDNAからなる複製または発現ベクターが含まれる。ベクターとしては、例えば、ori領域と、必要により上記cDNAの発現のためのプロモーター、プロモーターの制御因子などからなるプラスミド、ウィルスまたはファージベクターが挙げられる。ベクターはひとつまたはそれ以上の選択的マーカー遺伝子、例えばアンピシリン耐性遺伝子を含んでいてもよい。ベクターは、イン・ビトロ(in vitro)において、例えばcDNAに対応するRNAの製造、宿主細胞の形質転換に用いることができる。
【0022】
さらに、本発明には、配列番号1、2、5、6、9また10で示される塩基配列、またはそれらのオープンリーディングフレームからなるcDNAを含む本発明のcDNAを複製または発現させるためのベクターで形質転換された宿主細胞も含まれる。細胞としては、例えば細菌、酵母、昆虫細胞または哺乳動物細胞が挙げられる。
【0023】
さらに、本発明には、本発明のポリペプチドを発現させるための条件下で、本発明の宿主細胞を培養することからなる本発明のポリペプチドの製造方法も含まれる。培養は、本発明のポリペプチドが発現し、宿主細胞より製造される条件下で行われることが好ましい。
【0024】
本発明のcDNAは、上記のようなベクターのアンチセンス領域に挿入することでアンチセンスRNAを製造することもできる。このようなアンチセンスRNAは、細胞中の本発明のポリペプチドのレベルを制御することに用いることもできる。
【0025】
本発明は、本発明におけるポリペプチドのモノクローナルまたはポリクローナル抗体をも含む。さらに本発明におけるポリペプチドのモノクローナルまたはポリクローナル抗体の製造方法をも含む。モノクローナル抗体は、本発明のペプチドまたは、その断片を抗原として用い、通常のハイブリドーマの技術により製造することができる。ポリクローナル抗体は、宿主動物(例えば、ラットやウサギ等)に本発明のポリペプチドを接種し、免疫血清を回収する、通常の方法により製造することができる。
【0026】
本発明には、本発明のポリペプチド、その抗体と薬学的に許容される賦形剤および/または担体を含有する薬学的組成物も含まれる。
【0027】
(A)前記1の本発明のポリペプチドとしては、配列番号4、8または12で示されたアミノ酸配列からなるもの以外に、その一部が欠損したもの(例えば、配列番号4中、生物活性の発現に必須な部分だけからなる成熟ポリペプチド等)、その一部が他のアミノ酸と置換したもの(例えば、物性の類似したアミノ酸に置換したもの)、および上記本発明のポリペプチドに他のアミノ酸が付加または挿入されたものも含まれる。
【0028】
よく知られているように、ひとつのアミノ酸をコードするコドンは1〜6種類(例えば、Metは1種類、Leuは6種類)知られている。従って、ポリペプチドのアミノ酸配列を変えることなくcDNAの塩基配列を変えることができる。
【0029】
(B)前記3で特定される本発明のcDNAには、(A)の配列番号4、8または12で示されるポリペプチドをコードするすべての塩基配列群が含まれる。塩基配列を変えることによって、ポリペプチドの生産性が向上することがある。
【0030】
(C)配列番号2、6または10で特定されるcDNAは、(B)で示されるcDNAの一態様であり、天然型配列を表わす。
【0031】
(D)配列番号1、5または9で示されるcDNAは、(C)で特定されるcDNAに天然の非翻訳部分を加えた配列を示す。
【0032】
配列番号1、5または9で示される塩基配列からなるcDNAの作製は、以下の方法に従って行われる。
【0033】
はじめに酵母SST法(米国特許第5,536,637号に記載)の概要について説明する。
【0034】
サッカロマイセス・セルビシエ(Saccharomyces cerevisiae)などの酵母がショ糖またはラフィノースをエネルギー源や炭素源として利用するためにはインベルターゼを培地中に分泌しなければならない(インベルターゼはラフィノースをショ糖とメリビオースに、ショ糖をフルクトースとグルコースに分解する酵素である。)。また数多くの既知の哺乳類のシグナルペプチドは酵母のインベルターゼを分泌させうることが知られている。
【0035】
これらの知見から、酵母のインベルターゼの分泌を可能にする新規のシグナルペプチドを哺乳類のcDNAライブラリーからラフィノース培地上での酵母の生育を指標にスクリーニングする方法として本方法は開発された。
【0036】
翻訳開始点ATGを削除した非分泌型のインベルターゼ遺伝子SUC2(GENBANK accession No.V01311)を酵母の発現ベクターに組み込んで酵母SST用ベクターpSUC2を作製した。発現ベクターには、AAH5プラスミド(Gammerer, Methods in Enzymol. 101,192-201,1983)由来の発現用プロモーター(ADHプロモーター)およびターミネーター(ADHターミネーター)が組み込まれ、酵母複製起点としては2μ ori、酵母選択マーカーにはTRP1、大腸菌複製起点としてはColE1 ori、大腸菌薬剤耐性マーカーにはアンピシリン耐性遺伝子がそれぞれ組み込まれている。そのSUC2遺伝子の上流に哺乳類のcDNAを組み込んで、酵母SST cDNAライブラリーを調製した。このライブラリーを分泌型インベルターゼを欠損している酵母に形質転換した。
【0037】
組み込まれた哺乳類cDNAがシグナルペプチドをコードしている場合、酵母で発現されたインベルターゼに対しても分泌作用をもつと考えられ、その結果ラフィノース培地上での生育が可能となる。そこで出現したコロニーから酵母を培養してプラスミドを調製し、インサートcDNAの塩基配列を決定することによって、新規シグナルペプチドの検索を迅速かつ容易にした。
【0038】
酵母SST cDNAライブラリーの作製は、
工程(1):対象となる細胞よりmRNAを単離し、特定の制限酵素(酵素I)サイトを連結したランダムプライマーを用いて二本鎖cDNAを合成し、
工程(2):酵素Iとは異なる特定の制限酵素(酵素II)サイトを含むアダプターを連結して、酵素Iで消化した後、適当なサイズで分画し、
工程(3):酵母発現ベクター内のシグナルペプチドを削除したインベルターゼ遺伝子の上流に得られたcDNA断片を連結し、形質転換する工程よりなる。
【0039】
各工程を詳しく説明すると、工程(1)では、対象となる哺乳類の臓器や細胞株などより、必要により適当な刺激剤で刺激した後、公知の方法(以下、公知の方法は特に記載がなければMolecular Cloning(Sambrook, J., Fritsch, E. F.およびManiatis, T. 著、Cold Spring Harbor Laboratory Pressより1989年に発刊)またはCurrent Protocol in Molecular Biology(F.M.Ausubelら編、John Wiley & Sons, Inc.より発刊)に記載の方法に従って行われる。)に従ってmRNAの単離が行われる。
【0040】
対象となる組織としては、マウス胎児心臓が挙げられる。ランダムプライマーを用いる二本鎖cDNAの合成は公知の方法により行われる。
【0041】
アダプターに連結される制限酵素(酵素I)サイトと次の工程(2)で用いられる制限酵素(酵素II)サイトは、互いに異なるものであれば何を用いてもよい。好ましくは、酵素IとしてXhoI、酵素IIとしてはEcoRIが用いられる。
【0042】
工程(2)ではT4DNAポリメラーゼで末端を平滑化し、酵素IIアダプターを連結した後、酵素Iで消化し、アガロース電気泳動(AGE)により300〜800bpのcDNAを分画する。酵素IIは、前記したように酵素Iと異なるものなら何でもよい。
【0043】
工程(3)は、酵母発現用プラスミドベクターに連結されたシグナルペプチドを削除したインベルターゼの遺伝子の上流に(2)で得られたcDNA断片を組み込んで大腸菌に形質転換する工程である。ここで酵母発現用プラスミドベクターとしては、種々のものが知られているが、例えば、大腸菌内でも機能するYEp24などが用いられるが、好適には前述したプラスミドpSUC2が用いられる。
【0044】
形質転換のための宿主大腸菌株はすでに多くのものが知られており、好ましくはDH10Bのコンピテントセルである。また形質転換方法は公知の方法のいずれを用いてもよいが、好ましくはエレクトロポレーション法により行われる。形質転換体は公知の方法により培養され、酵母SST用のcDNAライブラリーが得られる。
【0045】
このcDNAライブラリーでは、すべてのクローンcDNA断片が導入されているわけではないし、またすべてが未知の(新規の)シグナルペプチドをコードする遺伝子断片とは限らない。そこで、次に該ライブラリーから未知のシグナルペプチドをコードする遺伝子断片をスクリーニングする必要がある。
【0046】
遺伝子をもたない酵母Saccharomycs cerevisiae(例えばYT455株など)またはインベルターゼ遺伝子を人為的に欠損させた株(公知の方法に従い作製可能)に、該cDNAライブラリーを導入し、シグナルペプチドをコードする配列を有する断片のスクリーニングを行なう。酵母の形質転換は公知の方法、例えば酢酸リチウム法によって行われる。形質転換体を選択培地で生育後、ラフィノースを炭素源とする培地に移し、生育可能なコロニーを選択し、プラスミドを回収する。ラフィノースを炭素源として酵母が生育したということは、ライブラリー中に何らかの分泌蛋白質のシグナルペプチドが組み込まれていたことを示している。
【0047】
次に、単離した陽性クローンについて、塩基配列を決定し、未知の蛋白質をコードすることが明らかになったcDNAについては、それをプローブとして全長クローンを単離し、全長の塩基配列を決定することができる。これらの操作は、当業者にとってすべて公知の方法で行われる。
【0048】
配列番号1、5または9で示される塩基配列が、一部、好ましくは全てが確定されると哺乳類に存在する本発明の蛋白質をコードするcDNAもしくは本発明蛋白質のホモローグおよびサブセットをコードするcDNAを得ることができる。適当な塩基配列を有するオリゴヌクレオチドを合成し、それを用いて、哺乳類由来のcDNAライブラリーあるいはmRNAからPCR法により、あるいは適当な塩基配列の断片をプローブとしてハイブリダイズさせることにより、他の哺乳類cDNAライブラリーあるいは該ゲノムライブラリーから、他の哺乳類型の当該蛋白質をコードするcDNAを得ることができる。
【0049】
このようにして得られたcDNAが、SSTで得られたcDNA断片の塩基配列(またはその相同配列)を含んでいるならばシグナルペプチドをコードしていることになるので、該cDNAが全長、またはほぼ全長であることは明らかである。
(シグナルペプチドは例外なく蛋白質のN末端に存在することから、cDNAのオープンリーディングフレームの5’末端にコードされている。)
【0050】
さらに公知の方法に従い、該cDNAをプローブとしてノザン(Northern)解析によって全長の確認をしてもよい。ハイブリダイズしたバンドから得られるmRNAのサイズと該cDNAのサイズを比較し、ほぼ同じであれば該cDNAはほぼ全長であると考えられる。
【0051】
本発明は、開示されたタンパクの全長型並びに成熟型の両方を提供する。それらのタンパクの全長型は、配列番号4、8または12で示される塩基配列の翻訳されるアミノ酸配列で同定される。それらの成熟タンパクは、適当な哺乳類の細胞あるいはその他の宿主細胞で開示された配列番号1、5または9で示される全長DNAを適当な哺乳類の細胞あるいはその他の宿主細胞で発現させることによって得られる。成熟型のタンパクの配列は、全長型のアミノ酸配列より予測可能である。
【0052】
配列番号1、2、5、6、9または10で示される塩基配列が一旦確定されると、その後は、化学合成によって、あるいは該塩基配列の断片を化学合成し、これをプローブとしてハイブリダイズさせることにより、本発明のcDNAを得ることができる。さらに、本cDNAを含有するベクターcDNAを適当な宿主に導入し、これを増殖させることによって、目的とするcDNAを必要量得ることができる。
【0053】
本発明のポリペプチドを取得する方法としては、
(1)生体または培養細胞から精製単離する方法、
(2)ペプチド合成する方法、または
(3)遺伝子組み換え技術を用いて生産する方法、
などが挙げられるが、工業的には(3)に記載した方法が好ましい。
【0054】
遺伝子組み換え技術を用いてポリペプチドを生産するための発現系(宿主−ベクター系)としては、例えば、細菌、酵母、昆虫細胞および哺乳動物細胞の発現系が挙げられる。
【0055】
例えば、大腸菌で発現させる場合には、成熟蛋白部分をコードするcDNAの5’末端に開始コドン(ATG)を付加し、得られたcDNAを、適当なプロモーター(例えば、trpプロモーター、lacプロモーター、λPLプロモーター、T7プロモーター等)の下流に接続し、大腸菌内で機能するベクター(例えば、pBR322、pUC18、pUC19等)に挿入して発現ベクターを作製する。
【0056】
次に、この発現ベクターで形質転換した大腸菌(例えば、E. Coli DH1、E. Coli JM109、E. Coli HB101株等)を適当な培地で培養して、その菌体より目的とするポリペプチドを得ることができる。また、バクテリアのシグナルペプチド(例えば、pelBのシグナルペプチド)を利用すれば、ペリプラズム中に目的とするポリペプチドを分泌することもできる。さらに、他のポリペプチドとのフュージョン・プロテイン(fusion protein)を生産することもできる。
【0057】
また、哺乳動物細胞で発現させる場合には、例えば、配列番号1、2、5、6、9または10で示される塩基配列を適当なベクター(例えば、レトロウイルスベクター、パピローマウイルスベクター、ワクシニアウイルスベクター、SV40系ベクター等)中の適当なプロモーター(例えば、SV40プロモーター、LTRプロモーター、メタロチオネインプロモーター等)の下流に挿入して発現ベクターを作製する。次に、得られた発現ベクターで適当な哺乳動物細胞(例えば,サルCOS−1細胞、COS−7細胞、チャイニーズハムスターCHO細胞、マウスL細胞等)を形質転換し、形質転換体を適当な培地で培養することによって、分泌蛋白である本発明の蛋白は、その細胞上清中に目的とするポリペプチドとして発現される。さらに、その他のポリペプチド、例えば抗体の定常領域(Fc portion)をコードするcDNA断片と連結することによって、フュージョン・プロテイン(fusion protein)を生産することもできる。以上のようにして得られたポリペプチドは、一般的な生化学的方法によって単離精製することができる。
【産業上の利用可能性】
【0058】
本発明のポリペプチドおよびそれをコードするcDNAは、一つあるいはそれ以上の効果あるいは生物活性(以下に列挙するアッセイに関連するものを含む。)を示すことが考えられる。
【0059】
本発明の蛋白に関して記述される効果あるいは生物活性は、その蛋白の投与あるいは使用により、あるいは、その蛋白をコードするcDNAの投与あるいは使用(例えば、遺伝子療法(再生医療を含む。)やcDNA導入に適したベクター)により提供される。
【0060】
本発明のポリペプチドが、脳の一部のニューロン(海馬神経初代培養細胞および海馬由来幹細胞)の生存維持活性を有していることから、神経変性が関わる疾患(脳梗塞や脳出血、脊髄損傷などによる中枢神経の損傷)の治療に有用である。
【0061】
また、該ポリペプチドのポリクローナル抗体またはモノクローナル抗体を用いて、生体における該ポリペプチドの定量が行え、これによって該ポリペプチドと疾患との関係の研究あるいは疾患の診断等に利用することができる。ポリクローナル抗体およびモノクローナル抗体は該ポリペプチドあるいはその断片を抗原として用いて公知の方法により作製することができる。
【0062】
また、該ポリペプチドを用いることにより、例えば、アフィニティーカラムを作製して、本ポリペプチドと結合する既知または未知の蛋白(リガンド)の同定、精製あるいはその遺伝子クローニングを行うことができる。
【0063】
また、該ポリペプチドを用いて、例えば、ウエスト−ウエスタン法により、または該cDNA(好ましくは該ポリペプチドをコードするcDNA)を用いて、例えば、酵母Two-Hybrid法により該ポリペプチドと相互作用する分子の同定、遺伝子クローニングを行うこともできる。
【0064】
さらに、本ポリペプチドを用いることによって、本ポリペプチドレセプターアゴニスト、アンタゴニストおよび受容体−シグナル伝達分子間の阻害剤等のスクリーニングを行うこともできる。
【0065】
スクリーニングは、例えば、以下の方法により行なうことが出来る。すなわち、
(a)本発明のポリペプチド、スクリーニングすべき化合物、及び細胞を含む反応混合物を、細胞が該ペプチドにより正常に刺激される条件下に一緒にし(該反応混合物は細胞が増殖するに従い細胞中に導入される標識および該ペプチドの機能を効果的に観察させるための該ペプチド以外のペプチドを含む);ついで、
(b)細胞の増殖の程度を測定して、該化合物が有効なアンタゴニストまたはアゴニストであるかどうかを決定する。
【0066】
本発明のcDNAは、多大な有用性が期待される本発明のポリペプチドを生産する際の重要かつ必須の鋳型となるだけでなく、遺伝病の診断や治療(遺伝子欠損症の治療またはアンチセンスDNA(RNA)によって、ポリペプチドの発現を停止させることによる治療等)に利用できる。
また、本発明のcDNAをプローブとしてジェノミック(genomic)DNAを分離できる。
【0067】
前記の疾患に適応するために、本発明のポリペプチド、あるいは本発明のポリペプチドに対する抗体は通常、全身的または局所的に、一般的には経口または非経口の形で投与される。好ましくは、経口投与、静脈内投与および脳室内投与である。
【0068】
投与量は、年齢、体重、症状、治療効果、投与方法、処理時間等により異なるが、通常、成人一人あたり、一回につき、100μgから100mgの範囲で、一日一回から数回経口投与されるかまたは、成人一人あたり、一回につき、10μgから100mgの範囲で、一日一回から数回非経口投与される。
【0069】
もちろん前記したように、投与量は種々の条件により変動するので、上記投与量より少ない量で十分な場合もあるし、また範囲を越えて必要な場合もある。
【0070】
本発明化合物を投与する際には、経口投与のための固体組成物、液体組成物およびその他の組成物、非経口投与のための注射剤、外用剤、坐剤等として用いられる。
【0071】
経口投与のための固体組成物には、錠剤、丸剤、カプセル剤、散剤、顆粒剤等が含まれる。カプセルには、ソフトカプセルおよびハードカプセルが含まれる。
【0072】
このような固体組成物においては、一つまたはそれ以上の活性物質が、少なくとも一つの不活性な希釈剤(例えば、ラクトース、マンニトール、グルコース、ヒドロキシプロピルセルロース、微結晶セルロース、デンプン、ポリビニルピロリドン、メタケイ酸アルミン酸マグネシウム等)と混合される。組成物は、常法に従って、不活性な希釈剤以外の添加物、例えば、潤滑剤(ステアリン酸マグネシウム等)、崩壊剤(繊維素グリコール酸カルシウム等)、安定化剤(ヒト血清アルブミン、ラクトース等)、溶解補助剤(アルギニン、アスパラギン酸等)を含有していてもよい。
【0073】
錠剤または丸剤は、必要により白糖、ゼラチン、ヒドロキシプロピルセルロース、ヒドロキシプロピルメチルセルロースフタレート等の胃溶性あるいは腸溶性のフィルムで被膜してもよいし、また2以上の層で被膜してもよい。さらにゼラチンのような吸収されうる物質のカプセルも包含される。
【0074】
経口投与のための液体組成物は、薬学的に許容される乳濁剤、溶液剤、懸濁剤、シロップ剤、エリキシル剤等を含み、一般に用いられる不活性な希釈剤(例えば、精製水、エタノール等)を含んでいてもよい。この様な組成物は、不活性な希釈剤以外に湿潤剤、懸濁剤のような補助剤、甘味剤、風味剤、芳香剤、防腐剤を含有していてもよい。
【0075】
経口投与のためのその他の組成物としては、ひとつまたはそれ以上の活性物質を含み、それ自体公知の方法により処方されるスプレー剤が含まれる。この組成物は不活性な希釈剤以外に亜硫酸水素ナトリウムのような安定剤と等張性を与えるような安定化剤、塩化ナトリウム、クエン酸ナトリウムあるいはクエン酸のような等張剤を含有していてもよい。スプレー剤の製造方法は、例えば米国特許第2,868,691号および同第3,095,355号明細書に詳しく記載されている。
【0076】
本発明による非経口投与のための注射剤としては、無菌の水性または非水性の溶液剤、懸濁剤、乳濁剤を包含する。水性または非水性の溶液剤、懸濁剤としては、一つまたはそれ以上の活性物質が、少なくとも一つの不活性な希釈剤と混合される。水性の希釈剤としては、例えば注射用蒸留水および生理食塩水が挙げられる。非水性の希釈剤としては、例えばプロピレングリコール、ポリエチレングリコール、オリーブ油のような植物油、エタノールのようなアルコール類、ポリソルベート80(登録商標)等が挙げられる。
【0077】
このような組成物は、さらに防腐剤、湿潤剤、乳化剤、分散剤、安定化剤(例えば、ヒト血清アルブミン、ラクトース等)、溶解補助剤(例えば、アルギニン、アスパラギン酸等)のような補助剤を含んでいてもよい。
【実施例】
【0078】
以下に本発明のSDNSFに関する実施例を挙げて本発明をより具体的に説明するが、これらは本発明の範囲を制限するものではない。
【0079】
実施例1:poly(A)+RNAの調製
成体ラット海馬由来神経幹細胞をクローン化したPZ5細胞よりTRIzol reagent(商品名、Life Technologies,Incより購入)を用いて全RNAを抽出し、OligotexdT30<Super>(商品名、Rocheより購入)を用いてpoly(A)+RNAを精製した。
【0080】
実施例2:酵母SST cDNAライブラリーの作製
上記poly(A)+RNAを鋳型にXhoI部位を連結した9mer
5'-TCC CGA TTG AAT TCT AGA CCT GCC TCG AGN NNN NNN NN-3' (配列番号13)
をプライマーとして、Super Script Choice System(商品名、Life Technologies,Incより購入)を用いて2本鎖cDNAの合成を行った。EcoRIアダプター(GIBCOERLより購入)をDNA ligation kit Ver.2(商品名、宝酒造(株)より購入。以後、cDNAの連結はすべて本キットを使用した。)を用いて連結した後、XhoIで消化し、アガロース電気泳動で400〜800bpのcDNAを切り出して分画し、pSuc2t7lori(米国特許5,536,637号参照)のEcoRI/XhoI部位に連結し、大腸菌DH10株にエレクトロポレーション法で形質転換して酵母SST用のcDNAライブラリーを得た。
【0081】
実施例3:SSTによるスクリーニングおよびSST陽性クローンの塩基配列の決定
このcDNAライブラリーのプラスミドを調製し、酢酸リチウム法(Current Protocols In Molecular Biology 13.7.1を参照)により酵母YTK12株を形質転換し、トリプトファン(Trp)不含の酵母形質転換体の選択培地(CMO-Trp培地)のプレート上にまいた。30℃で48時間インキュベートした後、Accutran Replica Plater(商品名、Schleicher & Schuellより購入)を用いて得られたコロニー(形質転換体)のレプリカをラフィノースを炭素源とするYRPプレートにとり、30℃で14日間インキュベートした。
3日目以降、出現してきた各々のコロニーを一つずつ再度YPRプレートにストリークして、30℃で48時間インキュベーション後、シングルコロニーをYPD培地に植菌し、さらに、30℃で48時間インキュベーションした後、プラスミドを調製した。続いて、pSUC2のクローニングサイトの両端の配列の2種類のプライマー(センス鎖はビオチン化プライマー)を用いて公知の方法に従ってPCRを行い、インサートcDNAを増幅した後、Dynabeads(商品名、DYNALより購入)を用いてビオチン化1本鎖cDNAを精製し、塩基配列の決定を行った。塩基配列の決定はDNA Sequencing kit(Dye Terminator Cycle Sequencing Ready Reaction)(商品名、Applied Biosystems Inc.より購入)を用いた蛍光ダイターミネーターサイクルシークエンス法で反応を行い、自動DNAシークエンサー373(Applied Biosystems Inc.)で読み取りを行った(以下の塩基配列決定もすべて本方法で行った)。
得られた塩基配列および推定されるアミノ酸配列についてデータベースとの相同性検索を行い、SDNSFと名付けられたクローンがデータベースに登録されていない新規のcDNAであることが明らかとなった。そこで、このSDNSFクローンの断片cDNA(以後、SDNSF SST断片cDNAと呼ぶ)について全長cDNAのクローニングを試みた。また、推定されるアミノ酸配列を既知のシグナルペプチドと比較することによりSDNSF SST断片cDNAが機能的かつ構造的にもシグナルペプチドを有することを確認した。
【0082】
実施例4:全長cDNAのクローニングおよび塩基配列の決定
PZ5cDNAライブラリーより得られた100万プラークをナイロンメンブレンにトランスファーした。32P標識したラットSDNSF SST断片cDNAをプローブとして、公知の方法に従いハイブリダイゼーションを行い、多数の陽性クローンを得た。その中の1クローンを単離して、大腸菌DH5αに形質転換してプラスミドを調製した。初めに5’側の塩基配列を決定してラットSDNSF SST断片cDNAの塩基配列が存在することを確認した後、全塩基配列を決定し、配列番号1に示す配列を得た。さらにオープンリーディングフレームを決定し、配列番号2に示すアミノ酸翻訳領域および配列番号4に示す推定アミノ酸配列を得た。
NCBIデータベース検索の結果、本発明のポリペプチド(ラットSDNSFポリペプチドと呼ぶ)およびコードする核酸配列と一致する配列はなかった。さらに、ラットSDNSFポリペプチドは膜貫通領域を持たないことも明らかとなり、本発明のラットSDNSFポリペプチドは新規の分泌蛋白質であることが判明した。
モチーフ検索の結果から、SDNSFはシグナルペプチドと下流に2つのEFハンドモチーフ(カルシウム結合モチーフ)を有することが判明した(第1図)。EFハンドモチーフはカルシウム分泌蛋白であるcalmodulinのEFハンドモチーフと共通の配列を有している(第2図)。EFハンドモチーフは分泌蛋白ではあまりみられない構造ではあるが、同様の構造を有する蛋白としてはBM-40とその関連蛋白が報告されている。BM-40においてはEFハンドモチーフが分泌小胞内のカルシウム濃度に応じて蛋白質高次構造の変化に寄与し、分泌効率に関与しているのではないかとされている(文献1:Busch E et.al., Calcium affinity cooperativity and domain interaction of extracellular EF-hands present in BM-40., J. Biol. Chem., 275(33), 25508-15(2000))。
【0083】
実施例5:ヒトおよびマウスSDNSF遺伝子の塩基配列の決定
他の哺乳類のESTsおよびUNIGENE DNAデータベースの相同性検索の結果、本発明者らはラットSDNSFと相同するヒトおよびマウスEST配列を見い出した。
そこで、本発明者らは、その配列情報を用いて、常法によりヒトおよびマウスSDNSFの全長遺伝子を単離し、全塩基配列を決定した。その結果、配列番号5および9に示す塩基配列を得た。さらにオープンリーディングフレームを決定し、配列番号8および12に示す推定アミノ酸配列を得た。以上のことから、該ヒトクローンおよびマウスクローンは全長であること、およびラットSDNSFに対してアミノ酸レベルで、それぞれ87%、90%一致していることが判明した。
このヒトSDNSFおよびマウスSDNSFについても核酸配列データベースおよびアミノ酸配列データベースを検索したが、ラットSDNSFと同様に一致する配列はなかった。このことから、本発明のポリペプチドも、新規の分泌蛋白質であることが判明した。
【0084】
実施例6:哺乳類以外の動物との相同性検索
線虫、およびショウジョウバエの遺伝子データベース検索の結果から線虫の仮想的な蛋白質として報告されているF55A11.1あるいはショウジョウバエの遺伝子産物として報告されているCG12817と20−30%のアミノ酸が一致する相同性を有することが確認された。このことから、種間の保存が保たれていると考えられた。
【0085】
実施例7:抗SDNSF蛋白ポリクローナル抗体の作製
固相法により合成し、キーホール リムペット ヘモシアニン(Keyhole limpet hemocyanin;KLH)コンジュゲートした3種類のラットSDNSFポリペプチドの部分ペプチド
Asp Lys Ser Thr Val His Asp Gln Glu His Ile Met Glu His Leu Glu Cys-KLH
(配列番号4の15〜30)
His Lys Glu Glu Gly Ser Glu Gln Val Pro Pro Met Ser Glu Asp Glu Cys-KLH
(配列番号4の74〜89)
KLH-Cys Asp Gly Tyr Ile Asp Tyr Ala Glu Phe Ala Lys Ser Leu Gln
(配列番号4の106〜119)
を免疫原としてウサギに免疫して、抗体価の測定後血清を採取した。得られた血清を各々免疫原としたペプチド断片を結合させたアフィニティーカラムにより抗SDNSF蛋白ポリクローナル抗体を精製した。
【0086】
実施例8:SDNSFの分泌経路の検討
ラットSDNSFのアミノ基末端にFLAG構造を付加したSDNSF合成蛋白(FLAG-SDNSF)およびカルボキシル基末端にFLAG-6His構造を付加したSDNSF合成蛋白(SDNSF-C'FLAG-6His)を293T細胞に強制発現させた。それぞれの培養上清中にSDNSF合成蛋白が分泌されていることを抗SDNSF抗体および抗FLAG抗体を用いたウェスタンブロット(Western Blot)法により確認した(第3図)。
【0087】
実施例9:ラットSDNSFの発現部位の同定
成体ラット脳、心臓、肺、肝臓、脾臓、腎臓、精巣、骨格筋、胸腺の組織よりTRIzol reagent(商品名、Life Technologies,Incより購入)を用いて全RNAを抽出し、OligotexdT30<Super>(商品名、Rocheより購入)を用いてpoly(A)+RNAを精製した。
それぞれのpoly(A)+RNAをホルムアルデヒドゲル電気泳動し、(Sambrook, et.al., 1989[Molecular Cloning])の方法に従い、ブロッティングを行った。ゲル精製を行った、32PラベルcDNAフラグメントをプローブとして、検出を行ったところ、第4図に示すように全ての臓器で発現が認められた。
ラット胎児の全身および脳、出生後7日までの脳の組織および神経系の初代培養細胞および細胞株よりTRIzol reagent(商品名、Life Technologies,Incより購入)を用いて全RNAを抽出し、RT-PCR法によりmRNAの発現量を調べた。第5図に示すように、培養幹細胞、神経初代培養細胞、神経幹細胞にSDNSFの発現が認められた。また、ヒト神経膠芽種細胞UG251およびマウス神経芽細胞N18ではSDNSFの発現を認めたが、グリア初代培養細胞およびラットグリオーマ細胞C6では発現を認めなかった。また、ウェスタンブロット(Western Blot)法によって、ヒト神経膠芽種細胞UG251およびマウス神経芽細胞N18でSDNSFタンパク質の発現が確認された(第6図)。
【0088】
実施例10:SDNSFの神経細胞および幹細胞生存支持活性の測定
SDNSFの発現部位より、神経細胞および幹細胞に対する機能を調べた。
His構造を利用しNi-NTA法により精製した合成SDNSF蛋白(SDNSF-FLAG-6His)をラット海馬神経初代培養細胞およびラット海馬由来幹細胞の培養上清に加えて、それぞれの培養条件下における培養4日目の生存細胞数をWST reduction法を用いて検討したところ、第7図に示すように神経初代細胞においてはSDNSF100ng/mlの濃度において無添加群と比較して細胞生存に有効であった。また、神経幹細胞においてもSDNSF添加群において無添加群と比較して生存細胞数が多い傾向が認められた。また、FGF−2(線維芽細胞増殖因子2)を除いた培地でのラット海馬由来幹細胞の5日目での生存活性は、SDNSF添加濃度に依存して増強した(第8図および第9図)。
【0089】
実施例11:SDNSFの神経幹細胞の自己再生および分化に対する効果の測定
ラット海馬由来幹細胞をSDNSF存在/FGF−2欠如培地で5日間培養後、さらに、非コート培養ディッシュ中にてFGF−2(20ng/ml)添加培地で6日間培養した後の神経肝細胞のニューロスフェア(神経細胞塊)を計数した。また、神経細胞の分化マーカーであるTuj-1陽性細胞を計測した。第10図および第11図に示すように、SDNSF添加神経幹細胞は、無添加群(Ctrl)と比較して有意なニューロスフェアの増加を呈した。さらに、そのうちの分化した神経細胞の割合は、無添加群(Ctrl)と比較して有意な増加を示した。これらの結果から、SDNSFは、FGF−2と同様に神経幹細胞の自己再生能を維持し、神経細胞やグリア細胞への分化を促進する活性を有すると考えられる。
【図面の簡単な説明】
【0090】
【図1】(A)はヒト、マウスおよびラットSDNSFのアミノ酸配列の相同性アライメントを示し、(B)はシグナルペプチドの下流に2つのEFハンドモチーフ(カルシウム結合モチーフ)を有することを示す。
【図2】SDNSFのEFハンドモチーフがカルシウム分泌蛋白であるカルモデュリン(calmodulin)のEFハンドモチーフと共通の配列を有していることを示す。
【図3】培養上清中にSDNSF合成蛋白が分泌されていることを抗SDNSF抗体および抗FLAG抗体を用いたウェスタンブロット(Western Blot)法により確認した様子を示す。
【図4】ゲル精製を行った、32PラベルcDNAフラグメントをプローブとして、検出を行った様子を示す。
【図5】神経初代培養細胞(Primary neuron)、神経膠星状細胞(astrocyte)、神経幹細胞(胎児神経幹細胞(embryo NSC)、未分化成人神経幹細胞(adult NSC undiff)、分化成人神経幹細胞(adult NSC diff))、ラットグリオーマ細胞C6、マウス神経芽細胞N18、ヒト神経膠芽種細胞UG251でのSDNSFmRNAの発現を示す。
【図6】初代神経膠星状細胞(primary astrocyte)、神経初代培養細胞(Primary neuron)、成人神経幹細胞(adult NSC)、マウス神経芽細胞N18、ヒト神経膠芽種細胞UG251でのSDNSFタンパク質の発現を示す。
【図7】(A)(B)は、神経初代細胞(hippocampal neuron)および神経幹細胞(neuron stem cells)におけるSDNSF添加による細胞生存活性に対する効果を示す。
【図8】SDNSFの添加が、濃度依存的に神経初代細胞の生存活性に有効であることを示す(*は、SDNSF非添加群に対する統計的有意差(P<0.01)を示す。)。
【図9】FGF−2非存在下で培養した神経幹細胞の生存活性に対するSDNSFの効果を示す(*は、無添加群(Ctrl)に対する統計的有意差(P<0.01)を示す。)。
【図10】FGF−2非存在下で培養した神経幹細胞のニューロスフェア(神経細胞塊)を計数することによる神経幹細胞の自己再生能に対するSDNSFの効果を示す(*は、無添加群(Ctrl)に対する統計的有意差(P<0.01)を示す。)。
【図11】FGF−2非存在下で培養した神経幹細胞のニューロスフェア(神経細胞塊)の神経細胞への分化能に対するSDNSFの効果を示す(*は、SDNSF添加群、FGF−2添加群および無添加群(Ctrl)間それぞれの統計的有意差(P<0.01)を示す。)。
【技術分野】
【0001】
本発明はヒト、ラット、マウスの幹細胞由来神経生存因子(Stem cell Derived-Neuron Survival Factor、以下、SDNSFと省略する。)に関する。
【0002】
さらに詳しく言えば、ヒト、ラット、マウスのSDNSF、それらの製造方法、SDNSFをコードするcDNA、そのcDNAからなるベクター、そのベクターで形質転換された宿主細胞、SDNSFの抗体、SDNSFまたは抗体を含有する薬学的組成物、SDNSFを測定する方法、SDNSFを測定する試薬およびSDNSFを用いたスクリーニング方法に関する。
【背景技術】
【0003】
成体脳では新しいニューロンは生まれないというのが長い間の定説であり、脳梗塞や脳出血、脊髄損傷などによる中枢神経の損傷、パーキンソン病や筋萎縮性側索硬化症(ALS)などの神経変性疾患などでは、神経細胞の細胞死により運動機能が失われ、いったん失われた機能の回復は難しいとされていた。しかし近年、ヒトやサルなどの高等哺乳動物の成体脳(海馬、大脳皮質連合野、側脳室)でもニューロンが新生することが見出され、これらの領域で新しく生まれるニューロンが神経幹細胞から発生することも示された。また、神経幹細胞が高齢者の脳内にも存在し、神経細胞に分化しうることも証明された。これらのことは、脳の再生治療が細胞を移植する細胞治療に限定されず、タンパク質医薬、薬物あるいは遺伝子治療などの方法により直接内在性の幹細胞を活性化する治療が可能であることを示唆している。
【0004】
従来、ある特定のポリペプチドまたはそれをコードするcDNAを得ようとする場合、組織や細胞培養液中に目的とする生物活性を確認し、次いでポリペプチドの単離精製を経て、遺伝子をクローニングするという方法、あるいはその生物活性を指標として遺伝子を発現クローニングする方法が一般的に用いられてきた。しかし、生体内生理活性ポリペプチドは、多様な生物活性を有している場合が多いので、あるひとつの活性を指標にして遺伝子をクローニングした結果、それが既知のポリペプチドと同一であることが後になって判明するという事例が増えている。また、微量しか産生されなかったり、特別な生理条件でのみ発現する因子も多く、そのことが単離、精製および生物活性の確認を困難なものとしている。
【発明の開示】
【0005】
本発明者らは、中枢神経の損傷または神経変性に係わる疾患の治療、診断、脳腫瘍の診断あるいは研究上有益な新規な因子(ポリペプチド)、特に分泌シグナルを有する分泌蛋白質および膜蛋白質に着目してそれを見出すべく、鋭意検討を行った。その結果、神経幹細胞に係わる新規なペプチド分子を単離し、上記のような再生治療に使用すること、および神経細胞の特異的マーカーとなることを見出した。
【0006】
本発明者らは、これまで造血系や免疫系で働く増殖分化因子の遺伝子のクローニングを研究してきた。そして、増殖分化因子(例えば、各種サイトカイン等)のような分泌蛋白質やそのレセプターのような膜蛋白質(以下、これらをまとめて分泌蛋白質等と呼ぶ。)の大部分がそのN末端にシグナルペプチドと呼ばれる配列を有していることに着目して、シグナルペプチドをコードする遺伝子を効率的かつ選択的にクローニングする方法を鋭意検討した。その結果、動物細胞を用いて、シグナルペプチドの有無を簡単に検索できる方法(シグナルシークエンストラップ(SST)法)を見出した(日本国特許第2,879,303号参照)。さらに同じ概念のもとに、酵母を用いてさらに大量かつ簡便にシグナルペプチドをコードする遺伝子を単離する方法(酵母SST法)も開発された(米国特許第5,536,637号)。
【0007】
本方法を用いて、成体ラット海馬由来神経幹細胞が産生している新規な分泌蛋白質、およびそれをコードするcDNAを同定することに成功し、その情報を基に全長cDNAを成体ラット海馬由来神経幹細胞より見出した。
【0008】
本ポリペプチドが、後に詳述するように、脳の一部のニューロン(海馬神経初代培養細胞および海馬由来幹細胞)に対し、生存支持活性を持つことを確認し、本発明を完成した。本ポリペプチドは、機能が特定された有用な因子である。
【0009】
本発明が提供するcDNA配列は、配列番号1または2で示されるラットSDNSFクローンとして同定され、成体ラット海馬由来神経幹細胞から作製したcDNAライブラリーより、酵母SST法を使用して得た情報をもとに単離された。配列番号1で示されるラットSDNSFクローンは分泌蛋白質(ここではラットSDNSF蛋白として表される。)をコードする完全なcDNA配列を含む全長鎖cDNAである。
【0010】
本発明が提供するcDNA配列は、配列番号5または6で示されるヒトSDNSFクローンとして同定され、成体ラット海馬由来神経幹細胞から作製したcDNAライブラリーより、酵母SST法を使用して得た情報をもとに単離された。配列番号5で示されるヒトSDNSFクローンは分泌蛋白質(ここではヒトSDNSF蛋白として表される。)をコードする完全なcDNA配列を含む全長鎖cDNAである。
【0011】
本発明が提供する配列番号9または10で示されるcDNA配列は、マウスSDNSFクローンとして同定され、成体ラット海馬由来神経幹細胞から作製したcDNAライブラリーより、酵母SST法を使用して得た情報をもとに単離された。配列番号9で示されるマウスSDNSFクローンは分泌蛋白質(ここではマウスSDNSF蛋白として表される。)をコードする完全なcDNA配列を含む全長鎖cDNAである。
【0012】
核酸配列データベースに登録されている既知の核酸配列に対してGenBankおよびNCBIを用いたBLASTN、FASTAおよびUNIGENEなどにより、またアミノ酸配列データベースに登録されている既知のポリペプチドのアミノ酸配列に対してBLASTP、Fly Database、SwissProtなどにより検索をした結果、本発明ポリペプチドであるラットSDNSFおよびそれらをコードする核酸配列と一致する配列はなかった。このことから、本発明のポリペプチドは、新規の分泌蛋白質であることが判明した。
【0013】
また、本発明者らは脳の一部のニューロンの生存支持活性を持つこと、分泌蛋白でありながらEFハンドモチーフを持ち、細胞外または分泌経路のオルガネラのカルシウムで調節を受けるサイトカインである可能性があること、および既知のいずれのニューロトロフィンとも相同性を有さないことから、既知のニューロトロフィン遺伝子欠損マウス解析からそれらへの依存性が知られている交感神経、知覚神経、脊髄運動神経核の神経、基底核コリン作動性神経以外のニューロンの形成や生存を支持するサイトカインである可能性と同時に、これまで原因不明であった神経変性疾患の病因解明および治療につながるものと考えられる。
【0014】
本発明は、
1.実質的に純粋な形である配列番号4、8または12で示されるアミノ酸配列からなるポリペプチド、そのホモローグ、そのフラグメントまたはそのフラグメントのホモローグ、
2.配列番号4、8または12で示されるアミノ酸配列からなる前記第1項記載のポリペプチド、
3.前記第1項または第2項記載のポリペプチドをコードするcDNA、
4.配列番号1、2、5、6、9または10で示される塩基配列を有する前記第3項記載のcDNA、またはその配列に選択的にハイブリダイズするフラグメント、
5.前記第3項または第4項記載のcDNAからなる複製または発現ベクター、
6.前記第5項記載の複製または発現ベクターで形質転換された宿主細胞、
7.前記第1項または第2項記載のポリペプチドを発現させるための条件下で前記第6項記載の宿主細胞を培養することを特徴とする該ポリペプチドの製造方法、
8.前記第1項または第2項記載のポリペプチドに対するポリクローナルまたはモノクローナル抗体、
9.前記第1項または第2項記載のポリペプチドまたは前記第8項記載の抗体および薬学的に許容される賦形剤および/または担体を含有することを特徴とする薬学的組成物、
10.前記第1項または第2項記載のポリペプチドまたは前記第8項記載の抗体および薬学的に許容される賦形剤および/または担体を含有することを特徴とする神経変性が係わる疾患の治療に有効な薬学的組成物、
11.神経変性が係わる疾患が、脳梗塞や脳出血、脊髄損傷などによる中枢神経の損傷である前記第10項記載の薬学的組成物、
12.前記第1項または第2項記載のポリペプチドの測定方法、
13.前記第8項記載の抗体を用いることを特徴とする前記第1項または第2項記載のポリペプチドの免疫化学的測定方法、
14.前記第12項または第13項記載の方法を用いることを特徴とする前記第1項または第2項記載のポリペプチドを検出する試薬、
15.前記第12項または第13項記載の方法を用いることを特徴とする腫瘍を検査する試薬、
16.腫瘍が脳腫瘍である前記第15項記載の試薬、
17.前記第1項または第2項記載のポリペプチドを用いることを特徴とする該ポリペプチドに対するアンタゴニストまたはアゴニストとしての活性を有する化合物をスクリーニングする方法、
18.前記第1項または第2項記載のポリペプチドを有効成分とする中枢神経損傷治療剤、
19.中枢神経損傷が脳梗塞による中枢神経損傷である前記第19項記載の試薬、
20.中枢神経損傷が脳出血による中枢神経損傷である前記第19項記載の試薬、および
21.中枢神経損傷が脊髄損傷による中枢神経損傷である前記第19項記載の試薬に関する。
【発明を実施するための最良の形態】
【0015】
実質的に純粋な形である配列番号4、8または12で示されるアミノ酸配列からなるポリペプチドとは、一般に、生産時のポリペプチドの90%以上、例えば、95、98または99%が配列番号4、8または12で示されるアミノ酸配列からなるポリペプチドであることを意味する。
【0016】
配列番号4、8または12で示されるアミノ酸配列からなるポリペプチドのホモローグとは、一般に少なくとも20個、好ましくは少なくとも30個、例えば40、60、80または100個の連続したアミノ酸領域で、少なくとも70%、好ましくは少なくとも80または90%、より好ましくは95%以上相同性であるものであり、そのようなホモローグは、以後本発明のポリペプチドとして記載される。
【0017】
さらに、配列番号4、8または12で示されるアミノ酸配列からなるポリペプチドのフラグメント、またはそれらのホモローグのフラグメントとは、少なくとも10アミノ酸、好ましくは少なくとも15アミノ酸、例えば20、25、30、40、50または60アミノ酸部分を意味する。
【0018】
配列番号1、2、5、6、9または10で示される塩基配列からなるcDNAに選択的にハイブリダイズするcDNAとは、一般に、少なくとも20個、好ましくは少なくとも30個、例えば40、60、80または100個の連続した塩基配列領域で、少なくとも70%、好ましくは少なくとも80または90%、より好ましくは95%以上相同性であるものであり、そのようなcDNAは、以後本発明のcDNAとして記載される。
【0019】
配列番号1、2、5、6、9または12で示される塩基配列からなるcDNAのフラグメントとは、少なくとも10塩基、好ましくは少なくとも15塩基、例えば20、25、30または40塩基部分を意味し、そのようなフラグメントも本発明のcDNAに含まれる。
【0020】
本発明のSDNSF蛋白は、脳腫瘍のうち未分化なニューロブラストーマとグリオブラストーマ細胞株から大量に分泌されるが、分化したグリオーマからは分泌されないことと、血中や髄液中に分泌されるニューロブラストーマとグリオブラストーマ脳腫瘍マーカーは無いので、SDNSF蛋白が、血中で検出可能な初の未分化脳腫瘍マーカーとすることができる。
【0021】
さらに、本発明には、本発明のcDNAからなる複製または発現ベクターが含まれる。ベクターとしては、例えば、ori領域と、必要により上記cDNAの発現のためのプロモーター、プロモーターの制御因子などからなるプラスミド、ウィルスまたはファージベクターが挙げられる。ベクターはひとつまたはそれ以上の選択的マーカー遺伝子、例えばアンピシリン耐性遺伝子を含んでいてもよい。ベクターは、イン・ビトロ(in vitro)において、例えばcDNAに対応するRNAの製造、宿主細胞の形質転換に用いることができる。
【0022】
さらに、本発明には、配列番号1、2、5、6、9また10で示される塩基配列、またはそれらのオープンリーディングフレームからなるcDNAを含む本発明のcDNAを複製または発現させるためのベクターで形質転換された宿主細胞も含まれる。細胞としては、例えば細菌、酵母、昆虫細胞または哺乳動物細胞が挙げられる。
【0023】
さらに、本発明には、本発明のポリペプチドを発現させるための条件下で、本発明の宿主細胞を培養することからなる本発明のポリペプチドの製造方法も含まれる。培養は、本発明のポリペプチドが発現し、宿主細胞より製造される条件下で行われることが好ましい。
【0024】
本発明のcDNAは、上記のようなベクターのアンチセンス領域に挿入することでアンチセンスRNAを製造することもできる。このようなアンチセンスRNAは、細胞中の本発明のポリペプチドのレベルを制御することに用いることもできる。
【0025】
本発明は、本発明におけるポリペプチドのモノクローナルまたはポリクローナル抗体をも含む。さらに本発明におけるポリペプチドのモノクローナルまたはポリクローナル抗体の製造方法をも含む。モノクローナル抗体は、本発明のペプチドまたは、その断片を抗原として用い、通常のハイブリドーマの技術により製造することができる。ポリクローナル抗体は、宿主動物(例えば、ラットやウサギ等)に本発明のポリペプチドを接種し、免疫血清を回収する、通常の方法により製造することができる。
【0026】
本発明には、本発明のポリペプチド、その抗体と薬学的に許容される賦形剤および/または担体を含有する薬学的組成物も含まれる。
【0027】
(A)前記1の本発明のポリペプチドとしては、配列番号4、8または12で示されたアミノ酸配列からなるもの以外に、その一部が欠損したもの(例えば、配列番号4中、生物活性の発現に必須な部分だけからなる成熟ポリペプチド等)、その一部が他のアミノ酸と置換したもの(例えば、物性の類似したアミノ酸に置換したもの)、および上記本発明のポリペプチドに他のアミノ酸が付加または挿入されたものも含まれる。
【0028】
よく知られているように、ひとつのアミノ酸をコードするコドンは1〜6種類(例えば、Metは1種類、Leuは6種類)知られている。従って、ポリペプチドのアミノ酸配列を変えることなくcDNAの塩基配列を変えることができる。
【0029】
(B)前記3で特定される本発明のcDNAには、(A)の配列番号4、8または12で示されるポリペプチドをコードするすべての塩基配列群が含まれる。塩基配列を変えることによって、ポリペプチドの生産性が向上することがある。
【0030】
(C)配列番号2、6または10で特定されるcDNAは、(B)で示されるcDNAの一態様であり、天然型配列を表わす。
【0031】
(D)配列番号1、5または9で示されるcDNAは、(C)で特定されるcDNAに天然の非翻訳部分を加えた配列を示す。
【0032】
配列番号1、5または9で示される塩基配列からなるcDNAの作製は、以下の方法に従って行われる。
【0033】
はじめに酵母SST法(米国特許第5,536,637号に記載)の概要について説明する。
【0034】
サッカロマイセス・セルビシエ(Saccharomyces cerevisiae)などの酵母がショ糖またはラフィノースをエネルギー源や炭素源として利用するためにはインベルターゼを培地中に分泌しなければならない(インベルターゼはラフィノースをショ糖とメリビオースに、ショ糖をフルクトースとグルコースに分解する酵素である。)。また数多くの既知の哺乳類のシグナルペプチドは酵母のインベルターゼを分泌させうることが知られている。
【0035】
これらの知見から、酵母のインベルターゼの分泌を可能にする新規のシグナルペプチドを哺乳類のcDNAライブラリーからラフィノース培地上での酵母の生育を指標にスクリーニングする方法として本方法は開発された。
【0036】
翻訳開始点ATGを削除した非分泌型のインベルターゼ遺伝子SUC2(GENBANK accession No.V01311)を酵母の発現ベクターに組み込んで酵母SST用ベクターpSUC2を作製した。発現ベクターには、AAH5プラスミド(Gammerer, Methods in Enzymol. 101,192-201,1983)由来の発現用プロモーター(ADHプロモーター)およびターミネーター(ADHターミネーター)が組み込まれ、酵母複製起点としては2μ ori、酵母選択マーカーにはTRP1、大腸菌複製起点としてはColE1 ori、大腸菌薬剤耐性マーカーにはアンピシリン耐性遺伝子がそれぞれ組み込まれている。そのSUC2遺伝子の上流に哺乳類のcDNAを組み込んで、酵母SST cDNAライブラリーを調製した。このライブラリーを分泌型インベルターゼを欠損している酵母に形質転換した。
【0037】
組み込まれた哺乳類cDNAがシグナルペプチドをコードしている場合、酵母で発現されたインベルターゼに対しても分泌作用をもつと考えられ、その結果ラフィノース培地上での生育が可能となる。そこで出現したコロニーから酵母を培養してプラスミドを調製し、インサートcDNAの塩基配列を決定することによって、新規シグナルペプチドの検索を迅速かつ容易にした。
【0038】
酵母SST cDNAライブラリーの作製は、
工程(1):対象となる細胞よりmRNAを単離し、特定の制限酵素(酵素I)サイトを連結したランダムプライマーを用いて二本鎖cDNAを合成し、
工程(2):酵素Iとは異なる特定の制限酵素(酵素II)サイトを含むアダプターを連結して、酵素Iで消化した後、適当なサイズで分画し、
工程(3):酵母発現ベクター内のシグナルペプチドを削除したインベルターゼ遺伝子の上流に得られたcDNA断片を連結し、形質転換する工程よりなる。
【0039】
各工程を詳しく説明すると、工程(1)では、対象となる哺乳類の臓器や細胞株などより、必要により適当な刺激剤で刺激した後、公知の方法(以下、公知の方法は特に記載がなければMolecular Cloning(Sambrook, J., Fritsch, E. F.およびManiatis, T. 著、Cold Spring Harbor Laboratory Pressより1989年に発刊)またはCurrent Protocol in Molecular Biology(F.M.Ausubelら編、John Wiley & Sons, Inc.より発刊)に記載の方法に従って行われる。)に従ってmRNAの単離が行われる。
【0040】
対象となる組織としては、マウス胎児心臓が挙げられる。ランダムプライマーを用いる二本鎖cDNAの合成は公知の方法により行われる。
【0041】
アダプターに連結される制限酵素(酵素I)サイトと次の工程(2)で用いられる制限酵素(酵素II)サイトは、互いに異なるものであれば何を用いてもよい。好ましくは、酵素IとしてXhoI、酵素IIとしてはEcoRIが用いられる。
【0042】
工程(2)ではT4DNAポリメラーゼで末端を平滑化し、酵素IIアダプターを連結した後、酵素Iで消化し、アガロース電気泳動(AGE)により300〜800bpのcDNAを分画する。酵素IIは、前記したように酵素Iと異なるものなら何でもよい。
【0043】
工程(3)は、酵母発現用プラスミドベクターに連結されたシグナルペプチドを削除したインベルターゼの遺伝子の上流に(2)で得られたcDNA断片を組み込んで大腸菌に形質転換する工程である。ここで酵母発現用プラスミドベクターとしては、種々のものが知られているが、例えば、大腸菌内でも機能するYEp24などが用いられるが、好適には前述したプラスミドpSUC2が用いられる。
【0044】
形質転換のための宿主大腸菌株はすでに多くのものが知られており、好ましくはDH10Bのコンピテントセルである。また形質転換方法は公知の方法のいずれを用いてもよいが、好ましくはエレクトロポレーション法により行われる。形質転換体は公知の方法により培養され、酵母SST用のcDNAライブラリーが得られる。
【0045】
このcDNAライブラリーでは、すべてのクローンcDNA断片が導入されているわけではないし、またすべてが未知の(新規の)シグナルペプチドをコードする遺伝子断片とは限らない。そこで、次に該ライブラリーから未知のシグナルペプチドをコードする遺伝子断片をスクリーニングする必要がある。
【0046】
遺伝子をもたない酵母Saccharomycs cerevisiae(例えばYT455株など)またはインベルターゼ遺伝子を人為的に欠損させた株(公知の方法に従い作製可能)に、該cDNAライブラリーを導入し、シグナルペプチドをコードする配列を有する断片のスクリーニングを行なう。酵母の形質転換は公知の方法、例えば酢酸リチウム法によって行われる。形質転換体を選択培地で生育後、ラフィノースを炭素源とする培地に移し、生育可能なコロニーを選択し、プラスミドを回収する。ラフィノースを炭素源として酵母が生育したということは、ライブラリー中に何らかの分泌蛋白質のシグナルペプチドが組み込まれていたことを示している。
【0047】
次に、単離した陽性クローンについて、塩基配列を決定し、未知の蛋白質をコードすることが明らかになったcDNAについては、それをプローブとして全長クローンを単離し、全長の塩基配列を決定することができる。これらの操作は、当業者にとってすべて公知の方法で行われる。
【0048】
配列番号1、5または9で示される塩基配列が、一部、好ましくは全てが確定されると哺乳類に存在する本発明の蛋白質をコードするcDNAもしくは本発明蛋白質のホモローグおよびサブセットをコードするcDNAを得ることができる。適当な塩基配列を有するオリゴヌクレオチドを合成し、それを用いて、哺乳類由来のcDNAライブラリーあるいはmRNAからPCR法により、あるいは適当な塩基配列の断片をプローブとしてハイブリダイズさせることにより、他の哺乳類cDNAライブラリーあるいは該ゲノムライブラリーから、他の哺乳類型の当該蛋白質をコードするcDNAを得ることができる。
【0049】
このようにして得られたcDNAが、SSTで得られたcDNA断片の塩基配列(またはその相同配列)を含んでいるならばシグナルペプチドをコードしていることになるので、該cDNAが全長、またはほぼ全長であることは明らかである。
(シグナルペプチドは例外なく蛋白質のN末端に存在することから、cDNAのオープンリーディングフレームの5’末端にコードされている。)
【0050】
さらに公知の方法に従い、該cDNAをプローブとしてノザン(Northern)解析によって全長の確認をしてもよい。ハイブリダイズしたバンドから得られるmRNAのサイズと該cDNAのサイズを比較し、ほぼ同じであれば該cDNAはほぼ全長であると考えられる。
【0051】
本発明は、開示されたタンパクの全長型並びに成熟型の両方を提供する。それらのタンパクの全長型は、配列番号4、8または12で示される塩基配列の翻訳されるアミノ酸配列で同定される。それらの成熟タンパクは、適当な哺乳類の細胞あるいはその他の宿主細胞で開示された配列番号1、5または9で示される全長DNAを適当な哺乳類の細胞あるいはその他の宿主細胞で発現させることによって得られる。成熟型のタンパクの配列は、全長型のアミノ酸配列より予測可能である。
【0052】
配列番号1、2、5、6、9または10で示される塩基配列が一旦確定されると、その後は、化学合成によって、あるいは該塩基配列の断片を化学合成し、これをプローブとしてハイブリダイズさせることにより、本発明のcDNAを得ることができる。さらに、本cDNAを含有するベクターcDNAを適当な宿主に導入し、これを増殖させることによって、目的とするcDNAを必要量得ることができる。
【0053】
本発明のポリペプチドを取得する方法としては、
(1)生体または培養細胞から精製単離する方法、
(2)ペプチド合成する方法、または
(3)遺伝子組み換え技術を用いて生産する方法、
などが挙げられるが、工業的には(3)に記載した方法が好ましい。
【0054】
遺伝子組み換え技術を用いてポリペプチドを生産するための発現系(宿主−ベクター系)としては、例えば、細菌、酵母、昆虫細胞および哺乳動物細胞の発現系が挙げられる。
【0055】
例えば、大腸菌で発現させる場合には、成熟蛋白部分をコードするcDNAの5’末端に開始コドン(ATG)を付加し、得られたcDNAを、適当なプロモーター(例えば、trpプロモーター、lacプロモーター、λPLプロモーター、T7プロモーター等)の下流に接続し、大腸菌内で機能するベクター(例えば、pBR322、pUC18、pUC19等)に挿入して発現ベクターを作製する。
【0056】
次に、この発現ベクターで形質転換した大腸菌(例えば、E. Coli DH1、E. Coli JM109、E. Coli HB101株等)を適当な培地で培養して、その菌体より目的とするポリペプチドを得ることができる。また、バクテリアのシグナルペプチド(例えば、pelBのシグナルペプチド)を利用すれば、ペリプラズム中に目的とするポリペプチドを分泌することもできる。さらに、他のポリペプチドとのフュージョン・プロテイン(fusion protein)を生産することもできる。
【0057】
また、哺乳動物細胞で発現させる場合には、例えば、配列番号1、2、5、6、9または10で示される塩基配列を適当なベクター(例えば、レトロウイルスベクター、パピローマウイルスベクター、ワクシニアウイルスベクター、SV40系ベクター等)中の適当なプロモーター(例えば、SV40プロモーター、LTRプロモーター、メタロチオネインプロモーター等)の下流に挿入して発現ベクターを作製する。次に、得られた発現ベクターで適当な哺乳動物細胞(例えば,サルCOS−1細胞、COS−7細胞、チャイニーズハムスターCHO細胞、マウスL細胞等)を形質転換し、形質転換体を適当な培地で培養することによって、分泌蛋白である本発明の蛋白は、その細胞上清中に目的とするポリペプチドとして発現される。さらに、その他のポリペプチド、例えば抗体の定常領域(Fc portion)をコードするcDNA断片と連結することによって、フュージョン・プロテイン(fusion protein)を生産することもできる。以上のようにして得られたポリペプチドは、一般的な生化学的方法によって単離精製することができる。
【産業上の利用可能性】
【0058】
本発明のポリペプチドおよびそれをコードするcDNAは、一つあるいはそれ以上の効果あるいは生物活性(以下に列挙するアッセイに関連するものを含む。)を示すことが考えられる。
【0059】
本発明の蛋白に関して記述される効果あるいは生物活性は、その蛋白の投与あるいは使用により、あるいは、その蛋白をコードするcDNAの投与あるいは使用(例えば、遺伝子療法(再生医療を含む。)やcDNA導入に適したベクター)により提供される。
【0060】
本発明のポリペプチドが、脳の一部のニューロン(海馬神経初代培養細胞および海馬由来幹細胞)の生存維持活性を有していることから、神経変性が関わる疾患(脳梗塞や脳出血、脊髄損傷などによる中枢神経の損傷)の治療に有用である。
【0061】
また、該ポリペプチドのポリクローナル抗体またはモノクローナル抗体を用いて、生体における該ポリペプチドの定量が行え、これによって該ポリペプチドと疾患との関係の研究あるいは疾患の診断等に利用することができる。ポリクローナル抗体およびモノクローナル抗体は該ポリペプチドあるいはその断片を抗原として用いて公知の方法により作製することができる。
【0062】
また、該ポリペプチドを用いることにより、例えば、アフィニティーカラムを作製して、本ポリペプチドと結合する既知または未知の蛋白(リガンド)の同定、精製あるいはその遺伝子クローニングを行うことができる。
【0063】
また、該ポリペプチドを用いて、例えば、ウエスト−ウエスタン法により、または該cDNA(好ましくは該ポリペプチドをコードするcDNA)を用いて、例えば、酵母Two-Hybrid法により該ポリペプチドと相互作用する分子の同定、遺伝子クローニングを行うこともできる。
【0064】
さらに、本ポリペプチドを用いることによって、本ポリペプチドレセプターアゴニスト、アンタゴニストおよび受容体−シグナル伝達分子間の阻害剤等のスクリーニングを行うこともできる。
【0065】
スクリーニングは、例えば、以下の方法により行なうことが出来る。すなわち、
(a)本発明のポリペプチド、スクリーニングすべき化合物、及び細胞を含む反応混合物を、細胞が該ペプチドにより正常に刺激される条件下に一緒にし(該反応混合物は細胞が増殖するに従い細胞中に導入される標識および該ペプチドの機能を効果的に観察させるための該ペプチド以外のペプチドを含む);ついで、
(b)細胞の増殖の程度を測定して、該化合物が有効なアンタゴニストまたはアゴニストであるかどうかを決定する。
【0066】
本発明のcDNAは、多大な有用性が期待される本発明のポリペプチドを生産する際の重要かつ必須の鋳型となるだけでなく、遺伝病の診断や治療(遺伝子欠損症の治療またはアンチセンスDNA(RNA)によって、ポリペプチドの発現を停止させることによる治療等)に利用できる。
また、本発明のcDNAをプローブとしてジェノミック(genomic)DNAを分離できる。
【0067】
前記の疾患に適応するために、本発明のポリペプチド、あるいは本発明のポリペプチドに対する抗体は通常、全身的または局所的に、一般的には経口または非経口の形で投与される。好ましくは、経口投与、静脈内投与および脳室内投与である。
【0068】
投与量は、年齢、体重、症状、治療効果、投与方法、処理時間等により異なるが、通常、成人一人あたり、一回につき、100μgから100mgの範囲で、一日一回から数回経口投与されるかまたは、成人一人あたり、一回につき、10μgから100mgの範囲で、一日一回から数回非経口投与される。
【0069】
もちろん前記したように、投与量は種々の条件により変動するので、上記投与量より少ない量で十分な場合もあるし、また範囲を越えて必要な場合もある。
【0070】
本発明化合物を投与する際には、経口投与のための固体組成物、液体組成物およびその他の組成物、非経口投与のための注射剤、外用剤、坐剤等として用いられる。
【0071】
経口投与のための固体組成物には、錠剤、丸剤、カプセル剤、散剤、顆粒剤等が含まれる。カプセルには、ソフトカプセルおよびハードカプセルが含まれる。
【0072】
このような固体組成物においては、一つまたはそれ以上の活性物質が、少なくとも一つの不活性な希釈剤(例えば、ラクトース、マンニトール、グルコース、ヒドロキシプロピルセルロース、微結晶セルロース、デンプン、ポリビニルピロリドン、メタケイ酸アルミン酸マグネシウム等)と混合される。組成物は、常法に従って、不活性な希釈剤以外の添加物、例えば、潤滑剤(ステアリン酸マグネシウム等)、崩壊剤(繊維素グリコール酸カルシウム等)、安定化剤(ヒト血清アルブミン、ラクトース等)、溶解補助剤(アルギニン、アスパラギン酸等)を含有していてもよい。
【0073】
錠剤または丸剤は、必要により白糖、ゼラチン、ヒドロキシプロピルセルロース、ヒドロキシプロピルメチルセルロースフタレート等の胃溶性あるいは腸溶性のフィルムで被膜してもよいし、また2以上の層で被膜してもよい。さらにゼラチンのような吸収されうる物質のカプセルも包含される。
【0074】
経口投与のための液体組成物は、薬学的に許容される乳濁剤、溶液剤、懸濁剤、シロップ剤、エリキシル剤等を含み、一般に用いられる不活性な希釈剤(例えば、精製水、エタノール等)を含んでいてもよい。この様な組成物は、不活性な希釈剤以外に湿潤剤、懸濁剤のような補助剤、甘味剤、風味剤、芳香剤、防腐剤を含有していてもよい。
【0075】
経口投与のためのその他の組成物としては、ひとつまたはそれ以上の活性物質を含み、それ自体公知の方法により処方されるスプレー剤が含まれる。この組成物は不活性な希釈剤以外に亜硫酸水素ナトリウムのような安定剤と等張性を与えるような安定化剤、塩化ナトリウム、クエン酸ナトリウムあるいはクエン酸のような等張剤を含有していてもよい。スプレー剤の製造方法は、例えば米国特許第2,868,691号および同第3,095,355号明細書に詳しく記載されている。
【0076】
本発明による非経口投与のための注射剤としては、無菌の水性または非水性の溶液剤、懸濁剤、乳濁剤を包含する。水性または非水性の溶液剤、懸濁剤としては、一つまたはそれ以上の活性物質が、少なくとも一つの不活性な希釈剤と混合される。水性の希釈剤としては、例えば注射用蒸留水および生理食塩水が挙げられる。非水性の希釈剤としては、例えばプロピレングリコール、ポリエチレングリコール、オリーブ油のような植物油、エタノールのようなアルコール類、ポリソルベート80(登録商標)等が挙げられる。
【0077】
このような組成物は、さらに防腐剤、湿潤剤、乳化剤、分散剤、安定化剤(例えば、ヒト血清アルブミン、ラクトース等)、溶解補助剤(例えば、アルギニン、アスパラギン酸等)のような補助剤を含んでいてもよい。
【実施例】
【0078】
以下に本発明のSDNSFに関する実施例を挙げて本発明をより具体的に説明するが、これらは本発明の範囲を制限するものではない。
【0079】
実施例1:poly(A)+RNAの調製
成体ラット海馬由来神経幹細胞をクローン化したPZ5細胞よりTRIzol reagent(商品名、Life Technologies,Incより購入)を用いて全RNAを抽出し、OligotexdT30<Super>(商品名、Rocheより購入)を用いてpoly(A)+RNAを精製した。
【0080】
実施例2:酵母SST cDNAライブラリーの作製
上記poly(A)+RNAを鋳型にXhoI部位を連結した9mer
5'-TCC CGA TTG AAT TCT AGA CCT GCC TCG AGN NNN NNN NN-3' (配列番号13)
をプライマーとして、Super Script Choice System(商品名、Life Technologies,Incより購入)を用いて2本鎖cDNAの合成を行った。EcoRIアダプター(GIBCOERLより購入)をDNA ligation kit Ver.2(商品名、宝酒造(株)より購入。以後、cDNAの連結はすべて本キットを使用した。)を用いて連結した後、XhoIで消化し、アガロース電気泳動で400〜800bpのcDNAを切り出して分画し、pSuc2t7lori(米国特許5,536,637号参照)のEcoRI/XhoI部位に連結し、大腸菌DH10株にエレクトロポレーション法で形質転換して酵母SST用のcDNAライブラリーを得た。
【0081】
実施例3:SSTによるスクリーニングおよびSST陽性クローンの塩基配列の決定
このcDNAライブラリーのプラスミドを調製し、酢酸リチウム法(Current Protocols In Molecular Biology 13.7.1を参照)により酵母YTK12株を形質転換し、トリプトファン(Trp)不含の酵母形質転換体の選択培地(CMO-Trp培地)のプレート上にまいた。30℃で48時間インキュベートした後、Accutran Replica Plater(商品名、Schleicher & Schuellより購入)を用いて得られたコロニー(形質転換体)のレプリカをラフィノースを炭素源とするYRPプレートにとり、30℃で14日間インキュベートした。
3日目以降、出現してきた各々のコロニーを一つずつ再度YPRプレートにストリークして、30℃で48時間インキュベーション後、シングルコロニーをYPD培地に植菌し、さらに、30℃で48時間インキュベーションした後、プラスミドを調製した。続いて、pSUC2のクローニングサイトの両端の配列の2種類のプライマー(センス鎖はビオチン化プライマー)を用いて公知の方法に従ってPCRを行い、インサートcDNAを増幅した後、Dynabeads(商品名、DYNALより購入)を用いてビオチン化1本鎖cDNAを精製し、塩基配列の決定を行った。塩基配列の決定はDNA Sequencing kit(Dye Terminator Cycle Sequencing Ready Reaction)(商品名、Applied Biosystems Inc.より購入)を用いた蛍光ダイターミネーターサイクルシークエンス法で反応を行い、自動DNAシークエンサー373(Applied Biosystems Inc.)で読み取りを行った(以下の塩基配列決定もすべて本方法で行った)。
得られた塩基配列および推定されるアミノ酸配列についてデータベースとの相同性検索を行い、SDNSFと名付けられたクローンがデータベースに登録されていない新規のcDNAであることが明らかとなった。そこで、このSDNSFクローンの断片cDNA(以後、SDNSF SST断片cDNAと呼ぶ)について全長cDNAのクローニングを試みた。また、推定されるアミノ酸配列を既知のシグナルペプチドと比較することによりSDNSF SST断片cDNAが機能的かつ構造的にもシグナルペプチドを有することを確認した。
【0082】
実施例4:全長cDNAのクローニングおよび塩基配列の決定
PZ5cDNAライブラリーより得られた100万プラークをナイロンメンブレンにトランスファーした。32P標識したラットSDNSF SST断片cDNAをプローブとして、公知の方法に従いハイブリダイゼーションを行い、多数の陽性クローンを得た。その中の1クローンを単離して、大腸菌DH5αに形質転換してプラスミドを調製した。初めに5’側の塩基配列を決定してラットSDNSF SST断片cDNAの塩基配列が存在することを確認した後、全塩基配列を決定し、配列番号1に示す配列を得た。さらにオープンリーディングフレームを決定し、配列番号2に示すアミノ酸翻訳領域および配列番号4に示す推定アミノ酸配列を得た。
NCBIデータベース検索の結果、本発明のポリペプチド(ラットSDNSFポリペプチドと呼ぶ)およびコードする核酸配列と一致する配列はなかった。さらに、ラットSDNSFポリペプチドは膜貫通領域を持たないことも明らかとなり、本発明のラットSDNSFポリペプチドは新規の分泌蛋白質であることが判明した。
モチーフ検索の結果から、SDNSFはシグナルペプチドと下流に2つのEFハンドモチーフ(カルシウム結合モチーフ)を有することが判明した(第1図)。EFハンドモチーフはカルシウム分泌蛋白であるcalmodulinのEFハンドモチーフと共通の配列を有している(第2図)。EFハンドモチーフは分泌蛋白ではあまりみられない構造ではあるが、同様の構造を有する蛋白としてはBM-40とその関連蛋白が報告されている。BM-40においてはEFハンドモチーフが分泌小胞内のカルシウム濃度に応じて蛋白質高次構造の変化に寄与し、分泌効率に関与しているのではないかとされている(文献1:Busch E et.al., Calcium affinity cooperativity and domain interaction of extracellular EF-hands present in BM-40., J. Biol. Chem., 275(33), 25508-15(2000))。
【0083】
実施例5:ヒトおよびマウスSDNSF遺伝子の塩基配列の決定
他の哺乳類のESTsおよびUNIGENE DNAデータベースの相同性検索の結果、本発明者らはラットSDNSFと相同するヒトおよびマウスEST配列を見い出した。
そこで、本発明者らは、その配列情報を用いて、常法によりヒトおよびマウスSDNSFの全長遺伝子を単離し、全塩基配列を決定した。その結果、配列番号5および9に示す塩基配列を得た。さらにオープンリーディングフレームを決定し、配列番号8および12に示す推定アミノ酸配列を得た。以上のことから、該ヒトクローンおよびマウスクローンは全長であること、およびラットSDNSFに対してアミノ酸レベルで、それぞれ87%、90%一致していることが判明した。
このヒトSDNSFおよびマウスSDNSFについても核酸配列データベースおよびアミノ酸配列データベースを検索したが、ラットSDNSFと同様に一致する配列はなかった。このことから、本発明のポリペプチドも、新規の分泌蛋白質であることが判明した。
【0084】
実施例6:哺乳類以外の動物との相同性検索
線虫、およびショウジョウバエの遺伝子データベース検索の結果から線虫の仮想的な蛋白質として報告されているF55A11.1あるいはショウジョウバエの遺伝子産物として報告されているCG12817と20−30%のアミノ酸が一致する相同性を有することが確認された。このことから、種間の保存が保たれていると考えられた。
【0085】
実施例7:抗SDNSF蛋白ポリクローナル抗体の作製
固相法により合成し、キーホール リムペット ヘモシアニン(Keyhole limpet hemocyanin;KLH)コンジュゲートした3種類のラットSDNSFポリペプチドの部分ペプチド
Asp Lys Ser Thr Val His Asp Gln Glu His Ile Met Glu His Leu Glu Cys-KLH
(配列番号4の15〜30)
His Lys Glu Glu Gly Ser Glu Gln Val Pro Pro Met Ser Glu Asp Glu Cys-KLH
(配列番号4の74〜89)
KLH-Cys Asp Gly Tyr Ile Asp Tyr Ala Glu Phe Ala Lys Ser Leu Gln
(配列番号4の106〜119)
を免疫原としてウサギに免疫して、抗体価の測定後血清を採取した。得られた血清を各々免疫原としたペプチド断片を結合させたアフィニティーカラムにより抗SDNSF蛋白ポリクローナル抗体を精製した。
【0086】
実施例8:SDNSFの分泌経路の検討
ラットSDNSFのアミノ基末端にFLAG構造を付加したSDNSF合成蛋白(FLAG-SDNSF)およびカルボキシル基末端にFLAG-6His構造を付加したSDNSF合成蛋白(SDNSF-C'FLAG-6His)を293T細胞に強制発現させた。それぞれの培養上清中にSDNSF合成蛋白が分泌されていることを抗SDNSF抗体および抗FLAG抗体を用いたウェスタンブロット(Western Blot)法により確認した(第3図)。
【0087】
実施例9:ラットSDNSFの発現部位の同定
成体ラット脳、心臓、肺、肝臓、脾臓、腎臓、精巣、骨格筋、胸腺の組織よりTRIzol reagent(商品名、Life Technologies,Incより購入)を用いて全RNAを抽出し、OligotexdT30<Super>(商品名、Rocheより購入)を用いてpoly(A)+RNAを精製した。
それぞれのpoly(A)+RNAをホルムアルデヒドゲル電気泳動し、(Sambrook, et.al., 1989[Molecular Cloning])の方法に従い、ブロッティングを行った。ゲル精製を行った、32PラベルcDNAフラグメントをプローブとして、検出を行ったところ、第4図に示すように全ての臓器で発現が認められた。
ラット胎児の全身および脳、出生後7日までの脳の組織および神経系の初代培養細胞および細胞株よりTRIzol reagent(商品名、Life Technologies,Incより購入)を用いて全RNAを抽出し、RT-PCR法によりmRNAの発現量を調べた。第5図に示すように、培養幹細胞、神経初代培養細胞、神経幹細胞にSDNSFの発現が認められた。また、ヒト神経膠芽種細胞UG251およびマウス神経芽細胞N18ではSDNSFの発現を認めたが、グリア初代培養細胞およびラットグリオーマ細胞C6では発現を認めなかった。また、ウェスタンブロット(Western Blot)法によって、ヒト神経膠芽種細胞UG251およびマウス神経芽細胞N18でSDNSFタンパク質の発現が確認された(第6図)。
【0088】
実施例10:SDNSFの神経細胞および幹細胞生存支持活性の測定
SDNSFの発現部位より、神経細胞および幹細胞に対する機能を調べた。
His構造を利用しNi-NTA法により精製した合成SDNSF蛋白(SDNSF-FLAG-6His)をラット海馬神経初代培養細胞およびラット海馬由来幹細胞の培養上清に加えて、それぞれの培養条件下における培養4日目の生存細胞数をWST reduction法を用いて検討したところ、第7図に示すように神経初代細胞においてはSDNSF100ng/mlの濃度において無添加群と比較して細胞生存に有効であった。また、神経幹細胞においてもSDNSF添加群において無添加群と比較して生存細胞数が多い傾向が認められた。また、FGF−2(線維芽細胞増殖因子2)を除いた培地でのラット海馬由来幹細胞の5日目での生存活性は、SDNSF添加濃度に依存して増強した(第8図および第9図)。
【0089】
実施例11:SDNSFの神経幹細胞の自己再生および分化に対する効果の測定
ラット海馬由来幹細胞をSDNSF存在/FGF−2欠如培地で5日間培養後、さらに、非コート培養ディッシュ中にてFGF−2(20ng/ml)添加培地で6日間培養した後の神経肝細胞のニューロスフェア(神経細胞塊)を計数した。また、神経細胞の分化マーカーであるTuj-1陽性細胞を計測した。第10図および第11図に示すように、SDNSF添加神経幹細胞は、無添加群(Ctrl)と比較して有意なニューロスフェアの増加を呈した。さらに、そのうちの分化した神経細胞の割合は、無添加群(Ctrl)と比較して有意な増加を示した。これらの結果から、SDNSFは、FGF−2と同様に神経幹細胞の自己再生能を維持し、神経細胞やグリア細胞への分化を促進する活性を有すると考えられる。
【図面の簡単な説明】
【0090】
【図1】(A)はヒト、マウスおよびラットSDNSFのアミノ酸配列の相同性アライメントを示し、(B)はシグナルペプチドの下流に2つのEFハンドモチーフ(カルシウム結合モチーフ)を有することを示す。
【図2】SDNSFのEFハンドモチーフがカルシウム分泌蛋白であるカルモデュリン(calmodulin)のEFハンドモチーフと共通の配列を有していることを示す。
【図3】培養上清中にSDNSF合成蛋白が分泌されていることを抗SDNSF抗体および抗FLAG抗体を用いたウェスタンブロット(Western Blot)法により確認した様子を示す。
【図4】ゲル精製を行った、32PラベルcDNAフラグメントをプローブとして、検出を行った様子を示す。
【図5】神経初代培養細胞(Primary neuron)、神経膠星状細胞(astrocyte)、神経幹細胞(胎児神経幹細胞(embryo NSC)、未分化成人神経幹細胞(adult NSC undiff)、分化成人神経幹細胞(adult NSC diff))、ラットグリオーマ細胞C6、マウス神経芽細胞N18、ヒト神経膠芽種細胞UG251でのSDNSFmRNAの発現を示す。
【図6】初代神経膠星状細胞(primary astrocyte)、神経初代培養細胞(Primary neuron)、成人神経幹細胞(adult NSC)、マウス神経芽細胞N18、ヒト神経膠芽種細胞UG251でのSDNSFタンパク質の発現を示す。
【図7】(A)(B)は、神経初代細胞(hippocampal neuron)および神経幹細胞(neuron stem cells)におけるSDNSF添加による細胞生存活性に対する効果を示す。
【図8】SDNSFの添加が、濃度依存的に神経初代細胞の生存活性に有効であることを示す(*は、SDNSF非添加群に対する統計的有意差(P<0.01)を示す。)。
【図9】FGF−2非存在下で培養した神経幹細胞の生存活性に対するSDNSFの効果を示す(*は、無添加群(Ctrl)に対する統計的有意差(P<0.01)を示す。)。
【図10】FGF−2非存在下で培養した神経幹細胞のニューロスフェア(神経細胞塊)を計数することによる神経幹細胞の自己再生能に対するSDNSFの効果を示す(*は、無添加群(Ctrl)に対する統計的有意差(P<0.01)を示す。)。
【図11】FGF−2非存在下で培養した神経幹細胞のニューロスフェア(神経細胞塊)の神経細胞への分化能に対するSDNSFの効果を示す(*は、SDNSF添加群、FGF−2添加群および無添加群(Ctrl)間それぞれの統計的有意差(P<0.01)を示す。)。
【特許請求の範囲】
【請求項1】
実質的に純粋な形である配列番号4、8または12で示されるアミノ酸配列からなるポリペプチドの測定方法。
【請求項2】
実質的に純粋な形である配列番号4、8または12で示されるアミノ酸配列からなるポリペプチドに対するポリクローナルまたはモノクローナル抗体を用いることを特徴とする前記ポリペプチドの免疫化学的測定方法。
【請求項3】
請求項1または2に記載の方法を用いることを特徴とする実質的に純粋な形である配列番号4、8または12で示されるアミノ酸配列からなるポリペプチドを検出する試薬。
【請求項4】
請求項1または2に記載の方法を用いることを特徴とする腫瘍を検査する試薬。
【請求項5】
腫瘍が脳腫瘍である請求項4記載の試薬。
【請求項6】
実質的に純粋な形である配列番号4、8または12で示されるアミノ酸配列からなるポリペプチドを用いることを特徴とする該ポリペプチドに対するアンタゴニストまたはアゴニストとしての活性を有する化合物をスクリーニングする方法。
【請求項7】
実質的に純粋な形である配列番号4、8または12で示されるアミノ酸配列からなるポリペプチドを有効成分とする中枢神経損傷治療剤。
【請求項8】
中枢神経損傷が脳梗塞による中枢神経損傷である請求項7記載の治療剤。
【請求項9】
中枢神経損傷が脳出血による中枢神経損傷である請求項7記載の治療剤。
【請求項10】
中枢神経損傷が脊髄損傷による中枢神経損傷である請求項7記載の治療剤。
【請求項1】
実質的に純粋な形である配列番号4、8または12で示されるアミノ酸配列からなるポリペプチドの測定方法。
【請求項2】
実質的に純粋な形である配列番号4、8または12で示されるアミノ酸配列からなるポリペプチドに対するポリクローナルまたはモノクローナル抗体を用いることを特徴とする前記ポリペプチドの免疫化学的測定方法。
【請求項3】
請求項1または2に記載の方法を用いることを特徴とする実質的に純粋な形である配列番号4、8または12で示されるアミノ酸配列からなるポリペプチドを検出する試薬。
【請求項4】
請求項1または2に記載の方法を用いることを特徴とする腫瘍を検査する試薬。
【請求項5】
腫瘍が脳腫瘍である請求項4記載の試薬。
【請求項6】
実質的に純粋な形である配列番号4、8または12で示されるアミノ酸配列からなるポリペプチドを用いることを特徴とする該ポリペプチドに対するアンタゴニストまたはアゴニストとしての活性を有する化合物をスクリーニングする方法。
【請求項7】
実質的に純粋な形である配列番号4、8または12で示されるアミノ酸配列からなるポリペプチドを有効成分とする中枢神経損傷治療剤。
【請求項8】
中枢神経損傷が脳梗塞による中枢神経損傷である請求項7記載の治療剤。
【請求項9】
中枢神経損傷が脳出血による中枢神経損傷である請求項7記載の治療剤。
【請求項10】
中枢神経損傷が脊髄損傷による中枢神経損傷である請求項7記載の治療剤。
【図1】



【図2】

【図3】
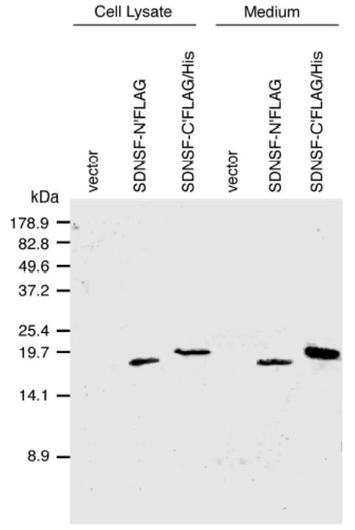
【図4】

【図5】
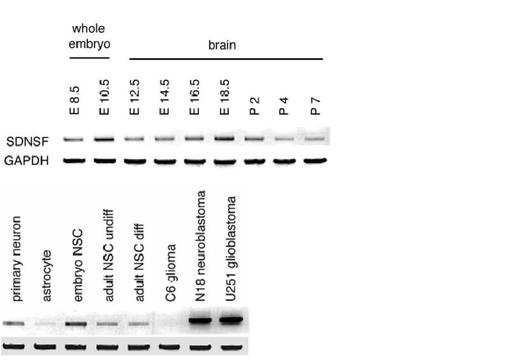
【図6】
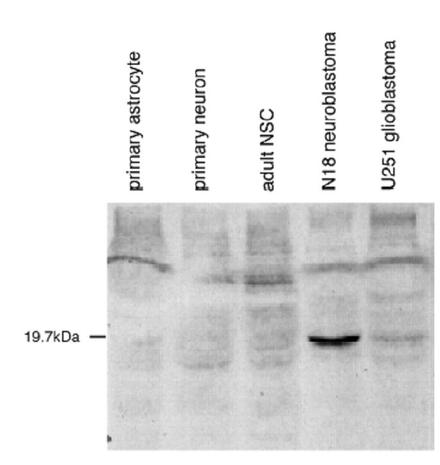
【図7】
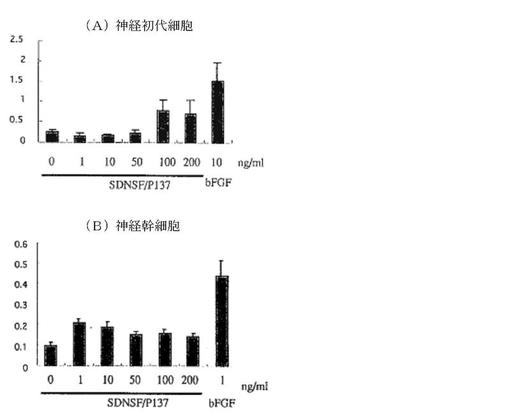
【図8】

【図9】

【図10】
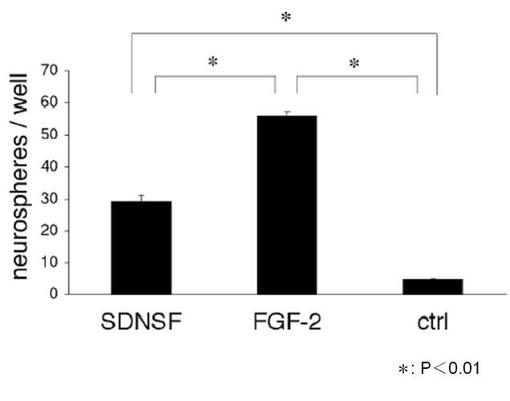
【図11】
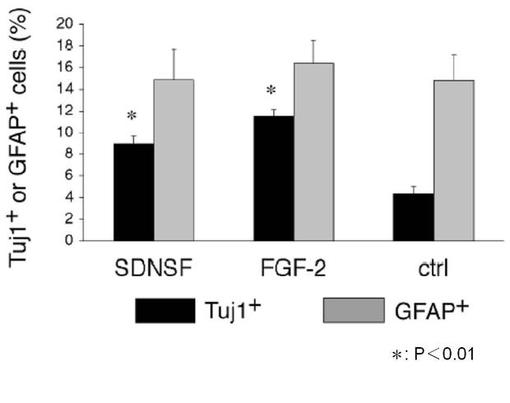
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【公開番号】特開2008−191146(P2008−191146A)
【公開日】平成20年8月21日(2008.8.21)
【国際特許分類】
【出願番号】特願2008−13506(P2008−13506)
【出願日】平成20年1月24日(2008.1.24)
【分割の表示】特願2003−538372(P2003−538372)の分割
【原出願日】平成14年10月22日(2002.10.22)
【出願人】(396023812)
【出願人】(000185983)小野薬品工業株式会社 (180)
【Fターム(参考)】
【公開日】平成20年8月21日(2008.8.21)
【国際特許分類】
【出願日】平成20年1月24日(2008.1.24)
【分割の表示】特願2003−538372(P2003−538372)の分割
【原出願日】平成14年10月22日(2002.10.22)
【出願人】(396023812)
【出願人】(000185983)小野薬品工業株式会社 (180)
【Fターム(参考)】
[ Back to top ]