
Fターム[4B065AA88]の内容
微生物、その培養処理 (127,014) | 微生物の種類 (32,496) | 動植物細胞又は組織 (14,879) | 植物細胞 (849)
Fターム[4B065AA88]に分類される特許
761 - 780 / 849
葉緑素を酵素的に脱色するための組成物及び方法
本発明は、葉緑素を含む組成物、例えば藻類調製物、葉緑素含有若しくは葉緑素夾雑飼料、食品又は油、例えば植物油(油種子(例えばキャノーラ(ナタネ)油又はダイズ油)又は油果実(例えばヤシ油)から加工された油を含む)の酵素的処理(“漂白”又は“脱色”)のための組成物及び方法を提供する。ある特徴では、本発明は、藻類、動物(例えば魚)若しくは植物調製物、食品、又は油中の葉緑素の酵素的加水分解のためにクロロフィラーゼ酵素を用いる方法を提供する。ある特徴では、前記クロロフィラーゼはシリカ上に固定される。本発明はまた工業的製造のための組成物及び洗剤の組成物を提供する。 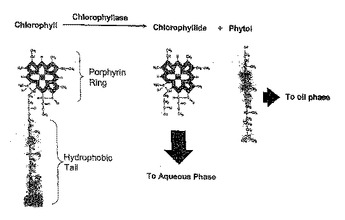 (もっと読む)
(もっと読む)
低下したアレルギー誘発性および維持されたT細胞反応性を有するイネ科からのI型のアレルゲンの変異体
本発明は、既知の野生型アレルゲンと比較して低下したIgE反応性および同時に十分に維持されたTリンパ球との反応性を特徴とする、イネ科(ドジョウツナギ)のI型のアレルゲンの変異体の調製および使用に関する。これらの低アレルギー誘発性のアレルゲンの変異体は、草花粉症を有する患者の特異的な免疫療法(減感作療法)に、または草花粉症の予防的免疫療法に用いることができる。 (もっと読む)
デンプン合成に関わる酵素をコードするDNA分子、ならびに該DNA分子を含むベクター、細菌、トランスジェニック植物細胞およびトランスジェニック植物体
【課題】可溶性デンプン合成酵素のDNA、およびトランスジェニック植物細部並びに植物体の提供。
【解決手段】植物のデンプン合成に関わる酵素は、可溶性デンプン合成酵素およびデンプン粒結合性デンプン合成酵素の二つ異なるアイソタイプよりなる。デンプン合成に関わる酵素をコードするDNA分子、ならびに該DNA分子を含むベクター、細菌、トランスジェニック植物細胞およびトランスジェニック植物を提供する。さらに、該タンパク質の増加または減少した活性を有する植物体から単離できるデンプンを提供する。
(もっと読む)
植物α−ファルネセン合成酵素およびそれをコードするポリヌクレオチド
本発明は、単離α−ファルネセン合成酵素およびその酵素をコードするポリヌクレオチド配列を提供する。本発明はまた、そのポリヌクレオチド配列を組み込んでいる核酸構築物、ベクターおよび宿主細胞をも提供する。これはさらに、この酵素を用いたα−ファルネセン産生、植物内でのα−ファルネセン合成の調節、およびα−ファルネセン合成酵素活性が変化した植物の選択にも関する。 (もっと読む)
I−SceI酵素をコ−ドするヌクレオチド配列、及びその使用

【課題】染色体DNAに部位特異的に、二本鎖の切れ目を少なくとも一つ誘導する方法により得られる組換え細胞の提供。
【解決手段】I-SceI酵素をコ−ドした、単離DNAを提供する。該DNA配列は、クロ−ニングベクタ及び発現ベクタ、形質転換細胞及びトランスジェニック動物へ導入することが可能である。該ベクタは遺伝子マッピングと、遺伝子の部位特異的導入に有益である。また、染色体DNA中に少なくとも一つの、部位特異的な切れ目を誘導し、ポリペプチドをコードするDNAを挿入する方法により毛質転換する組換え細胞の提供は(a)I-SceI制限部位を具備した二本鎖DNAを有する細胞を用意する;(b)I-SceIをコ−ドするDNAを具備した、少なくとも一つのプラスミドで前記細胞を形質転換する;および、(c)少なくとも一つの、二本鎖の切れ目が誘導された細胞を選択する。
(もっと読む)
クロロフィル分解に関与する遺伝子NYC1とその利用
【課題】 植物体のクロロフィル分解(黄化)を抑制し、緑色を維持する能力を高めることを課題とする。
【解決手段】 老化時にも葉が黄化しない突然変異体nyc1-1から、ポジショナルクローニングにより、その原因遺伝子の単離を試みた結果、黄化関連遺伝子NYC1がP0452F10.5遺伝子とP0443E07.24遺伝子との間に存在することが明らかとなった。突然変異体nyc1-1についてこれら2つの遺伝子間の領域の塩基配列を解析したところ、両遺伝子間に存在するP0452F10.4遺伝子の1塩基置換の存在が確認され、突然変異体nyc1-1ではこの遺伝子のスプライシングの異常が起きていることが推定された。さらに、緑色維持の表現型を示す培養変異体nyc1-2について上記領域を解析したところ、P0452F10.4遺伝子がトランスポゾンによって破壊されていることが判明した。野生型のNYC1を含むゲノム断片を緑色維持変異体nyc1-2に導入したところ、野生型のように黄化を示すことが確認された。黄化関連遺伝子NYC1の機能を抑制することにより、緑色を維持する能力を高めることが可能である。
(もっと読む)
組換え微生物によるフラボノイドの生成
微生物宿主におけるフラボノイドの生成のための方法および組成物が提供される。組成物は、フェニルプロパノイドの種々のフラボノイドへの変換のための生合成経路中の1以上のステップに関与する酵素をコードする遺伝子のセットを含む。方法は、遺伝子のセットを異種宿主細胞に導入して、前記遺伝子の発現が酵素の生成を生じるよう、適当な培地中で細胞を増殖させるステップを含む。特的の基質(複数も可)を形質転換細胞に供給すると、酵素が基質に作用して所望のフラボノイドを生成する。 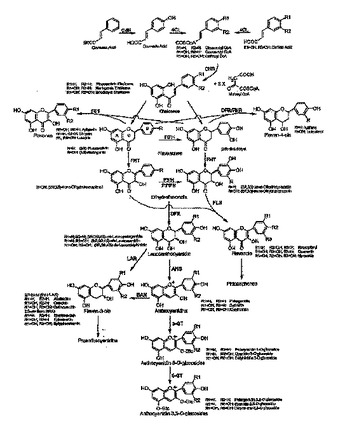 (もっと読む)
(もっと読む)
ファゴピリトールシンターゼ遺伝子およびその使用法
本発明はファゴピリトールシンターゼをコードする単離されたDNA分子に関する。ファゴピリトール、インスリンメディエーター、インスリンメディエーターアナログ、インスリンメディエーターホモログ、またはインスリンメディエーターインヒビターを製造するための方法もまた、記載される。本方法は、ファゴピリトールシンターゼを提供する工程、ガラクトシルドナーおよびガラクトシルアクセプターを含む基質を提供する工程、ならびに、ファゴピリトール、インスリンメディエーター、インスリンメディエーターアナログ、インスリンメディエーターホモログ、またはインスリンメディエーターインヒビターを有効に製造するする条件下で、ファゴピリトールシンターゼと基質とを組み合わせる工程、を含む。 (もっと読む)
急速に成熟する蛍光タンパク質およびその使用法
急速に成熟する蛍光タンパク質およびその非凝集変種(およびその変異体)をコードする核酸組成、ならびにおなじものをコードするタンパク質を提供する。関心対象のタンパク質は蛍光性で、この特徴はそのタンパク質の2残基以上の相互作用から生じる。本発明のタンパク質は、ある態様において、花虫類(Anthozoan)のような非生物発光刺胞動物(Cnidarian)、もしくは花虫類非ウミエラ(Pennatulacean)(シーペン(sea pen))種のいずれかから得られた野生型タンパク質の変異体であるという特徴をさらに有する。ある態様について、本発明のタンパク質は野生型イソギンチャクモドキ(Discosoma)種「赤色」蛍光タンパク質の変異体である。実質的に上記特定のタンパク質と同類のタンパク質、またはそれらの変異体もまた関心対象である。また、核酸の断片およびそれをコードするペプチド、ならびに本発明のタンパク質、形質転換細胞、および形質転換組織に対する抗体も提供される。本発明のタンパク質組成および核酸組成は様々な異った適用で使用される。最後に、このような適用に用いるキット(たとえば本発明の核酸組成を含むようなキット)が提供される。 (もっと読む)
イミダゾリノン耐性植物作出のための成熟型AHASLタンパク質をコードするポリヌクレオチド

成熟型野性型イミダゾリノン耐性アセトヒドロキシ酸合成酵素の大サブユニット(AHASL)ポリペプチドをコードする、単離されたポリヌクレオチド分子と、これらのポリペプチドをコードするアミノ酸配列を開示する。本発明のポリヌクレオチド分子を含む発現カセットと形質転換ベクター並びにポリヌクレオチド分子、発現カセット及び形質転換ベクターにより形質転換される植物と宿主細胞を開示する。植物の除草剤に対する抵抗性を亢進するようにポリヌクレオチド分子を用いる方法と、このような植物の近傍に存在する雑草を抑制する方法も開示する。 (もっと読む)
細菌ロドバクタースフェロイデスから得られるチロシンアンモニアリアーゼ酵素のDNAおよびアミノ酸配列
細菌ロドバクタースフェロイデス(Rhodobacter sphaeroides)において、新規のチロシンアンモニアリアーゼ酵素が同定された。この酵素は、フェニルアラニンに対してよりもチロシンに対してより高い活性を有し、チロシンから直接パラ−ヒドロキシケイ皮酸を生成するために有用である。この酵素をコードする遺伝子が、ゲノムDNAを用いる直接的増幅によってクローン化され、大腸菌中で発現された。 (もっと読む)
植物における生物学的に安全な一過性のタンパク質発現
植物又は植物の葉において目的配列から目的タンパク質を発現させることによって目的タンパク質を産生する方法であって:a)レプリコンをコードする配列部分を有する異種DNA配列をT−DNA中に含有するアグロバクテリウム株を補完因子の存在下で植物又は植物の葉に浸潤させることによって植物又は植物の葉にトランスフェクションし、レプリコンをコードする配列は、植物ウイルスに由来する、レプリコンのレプリコン機能に必要な配列、及びレプリコンから発現されるべき目的配列を含有し、b)場合により、工程(a)で浸潤させた植物又は植物の葉から目的タンパク質を単離する、ことを含み、アグロバクテリウム株は、補完因子の非存在下では生物へのT−DNAのトランスフェクションを不良にする第1の遺伝子改変が提供されている、前記方法。 (もっと読む)
篩管局在性グルタチオンSトランスフェラーゼ
篩管局在性グルタチオンSトランスフェラーゼを提供する。イネの篩管液に存在するタンパク質を分析し、植物のグルタチオンSトランスフェラーゼと高い相同性を有するタンパク質を同定する。 (もっと読む)
植物に於ける組織因子の産生
本発明は、植物に於いて作動可能な第一のプロモーター、核酸配列が前記プロモーターに作動可能に会合する哺乳動物の組織因子タンパク質又はその機能断片をコードする発現可能な核酸配列、及び前記核酸配列に作動可能に会合する終止配列を含んで成る植物を発現するベクターを提供するものである。本発明はさらに、請求の範囲に記載のベクター及び植物を用いて前記組織因子タンパク質又はその機能的な断片を作製する方法、及び対象に組織因子タンパク質を投与することにより、対象に於いて失血を治療又は予防する、創傷治癒を促進させる、血管形成又は血管の再構築を促進させる方法を提供するものである。 (もっと読む)
セスキテルペン合成及び使用方法
本発明は、セスキテルペン合成酵素及びその製造方法及びその使用に関する。一実施態様では、本発明は、少なくとも1種のセスキテルペン合成酵素をコードする前記のヌクレオチド配列を有する核酸を提供する。更なる実施態様では、また本発明はセスキテルペン合成酵素及びこれらの酵素の製造方法及び使用方法を提供する。例えば本発明のセスキテルペン合成酵素を用いて、ファルネシルピロリン酸を種々に酸素化された及び脂肪族のセスキテルペン、例えばバレンセン、ビシクロゲルマクレン、クベボール及びδ−カジネンに変換することができる。 (もっと読む)
被子植物遺伝子導入植物を作製するためのベクター
任意の被子植物細胞の色素体ゲノム中の異種遺伝子を安定的に挿入し発現するDNAベクターを提供する。本ベクターでは、異なる分類の植物(双子葉植物及び単子葉植物)に属するatpBオペロンとrbcLオペロンの組み合わせから得られた人工遺伝子間領域に位置する多重クローニング部位に外来遺伝子を挿入することができる。ベクターのatpB及びrbcLボーダー配列と対応する色素体ゲノムの相同領域との間の相同組換えによって、色素体ゲノムにこの発現カセットを挿入する。かかる方法により、目的の2種以上の遺伝子が、atpB遺伝子をコードするボーダー領域に隣接するrbcLプロモーターの転写調節下で発現可能である。 (もっと読む)
アルデヒドデヒドロゲナーゼ遺伝子
本発明は、アルデヒドデヒドロゲナーゼ(SNDH)をコードするDNA、前記DNAを含む発現ベクター、および前記DNAを含む組換え微生物に関する。さらに、本発明は、組換えアルデヒドデヒドロゲナーゼタンパク質を産生するプロセス、および、組換えアルデヒドデヒドロゲナーゼタンパク質または発現ベクターを含む組換え微生物を使用することにより、L−ソルボソンからL−アスコルビン酸(ビタミンC)および/または2−ケト−L−グロン酸(2−KGA)を産生するプロセスにも関する。また、前記アルデヒドデヒドロゲナーゼをコードする遺伝子が破壊されている微生物を用いて、2−KGAを産生するプロセスも提供する。 (もっと読む)
トランスジェニック植物の病原体抵抗性をペルオキシダーゼ発現により増大させる方法
本発明は、ペルオキシダーゼ活性を有するタンパク質をコードするDNA配列を植物に挿入し、その中で発現させることを特徴とする、増大した病原体抵抗性を有するトランスジェニック植物及び/又は植物細胞の作製方法に関する。本発明はまた、増大した病原体抵抗性を有するトランスジェニック植物又は植物細胞を作製するための、ペルオキシダーゼをコードする核酸の使用に関する。さらに、本発明はオオムギ由来のペルオキシダーゼをコードする核酸配列に関する。 (もっと読む)
腫瘍特異的抗体
本発明は、腫瘍特異的抗体の重鎖および軽鎖相補性決定領域のアミノ酸および核酸配列を提供する。さらに、本発明は、腫瘍特異的抗体および毒素または標識に結合した腫瘍特異的抗体を含む免疫複合体、ならびにそれらの方法および使用を提供する。本発明はまた、本発明の腫瘍特異的抗体を使用する診断方法およびキットに関する。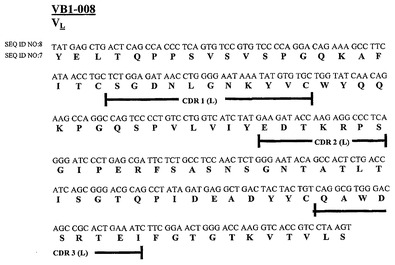
 (もっと読む)
(もっと読む)
4−アミノ−4−デオキシコリスミ酸(ADC)および[3R,4R]−4−アミノ−3−ヒドロキシシクロヘキサ−1,5−ジエン−1−カルボン酸(3,4−CHA)の生合成生産
本発明は、高められたレベルの活性で、4−アミノ−4−デオキシコリスミ酸シンターゼ、好ましくはPabAB二部分タンパク質(それは融合タンパク質であってもよい)を使ってインビボで発酵により行われ、それによって、回収されるADCおよび4−アミノ−4−デオキシプレフェネート(ADP)を含む培養液を得る、4−アミノ−4−デオキシコリスミ酸(ADC)の生合成生産方法に関する。本発明はまた、p−アミノフェニルアラニンへのADPのさらなる転化方法にも関する。本発明はさらに、3,4−CHAの回収を含む、かかる4−アミノ−4−デオキシコリスミ酸シンターゼのおよびイソコリメートを[5S,6S]−5,6−ジヒドロキシシクロヘキサ−1,3−ジエン−1カルボン酸(2,3−CHD)へ転化することができる酵素、好ましくはフェナジン生合成タンパク質PhzDの共同作用による[3R,4R]−4−アミノ−3−ヒドロキシシクロヘキサ−1,5−ジエン−1−カルボン酸(3,4−CHA)の生合成生産に関する。本発明はまた、かかる方法の任意のものでの使用のための発現ベクターおよびホスト細胞にも関する。本発明はさらに、触媒活性生成物としての、特にキラル触媒としての3,4−CHAの使用に関する。そして本発明は最後に3,4−CHAからのリン酸オセルタミビルの合成に関する。 (もっと読む)
761 - 780 / 849
[ Back to top ]