
Fターム[4B065AA88]の内容
微生物、その培養処理 (127,014) | 微生物の種類 (32,496) | 動植物細胞又は組織 (14,879) | 植物細胞 (849)
Fターム[4B065AA88]に分類される特許
781 - 800 / 849
再構成ヒストンメチルトランスフェラーゼ複合体及びそのモジュレーターの識別方法
本発明は、EEDと、EZH2と、SUZ12とを包含する再構成複合体を提供し、ここで再構成複合体はヒストンH3のリジン27(H3−K27)に対してヒストンメチルトランスフェラーゼ(HMTase)活性を有する。再構成複合体は、RbAp48、AEBP2、あるいはそれらの両方をさらに含むことができる。再構成複合体の製造方法、再構成複合体のHTMase活性を阻害する化合物を識別する方法、及び、癌を治療するための候補化合物を識別する方法も開示する。再構成複合体を包含する試薬及びキットも更に提供される。  (もっと読む)
(もっと読む)
少なくとも一つのトレー状揺動プラットフォームを含むバイオリアクタアセンブリ
本発明は、2リットルから数千リットルまでの培養量の同一条件下で線形のスケールアップを可能にする、リアクタフレームを含むバイオリアクタアセンブリに関する。バイオリアクタは、少なくとも2リットルの全体積を受けるように適合された少なくとも一つの使い捨てバッグを保持することができる少なくとも一つの揺動プラットフォームを含む。前記少なくとも一つのプラットフォームは、バッグ中の流体を動かし、波状の動きを形成させるトレー状付形物を有する。前記少なくとも一つのプラットフォームは、リアクタフレームに取り付けられ、垂直軸に沿って互いの上に配置されている。特に、揺動中、揺動プラットフォームの重量分布が概ね平衡状態になる。本発明はさらに、バイオリアクタアセンブリを使用する方法に関する。 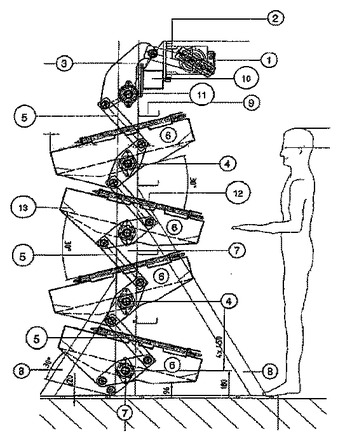 (もっと読む)
(もっと読む)
Nicotiana核酸分子およびその用途
本発明は、Nicotiana植物におけるNicotiana核酸配列、例えば構成的またはエチレンもしくは老化誘導性ポリペプチドをコードする配列、特にシトクロムp450酵素をコードする配列、ならびに例えば育種プロトコールを用いることにより、所望の形質を改変するための、これらの核酸配列および植物の使用方法に関する。 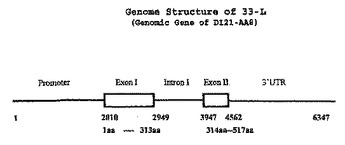 (もっと読む)
(もっと読む)
除草剤抵抗性遺伝子
本発明は、2,4-Dおよび他のフェノキシオーキシン除草剤に対してのみならず、アリールオキシフェノキシプロピオネート除草剤にもまた抵抗性である新規な植物を提供する。従来、これらの有利な特性の両方を有する植物が、単一の遺伝子の導入によって産生することができるという予測または示唆は存在しなかった。本発明はまた、より広くかつより強固な雑草の制御、処理の柔軟性の増加、および除草剤抵抗性管理の選択肢の改善を提供するために、本発明の1種または複数の酵素を、単独で、または別の除草剤抵抗性遺伝子、好ましくは、グリフォセート抵抗性遺伝子とともに「重ね合わせて」産生する植物を含む。より具体的には、本発明に従う使用のための好ましい酵素および遺伝子は、本明細書でAAD(アリールオキシシアルカノエート ジオキシゲナーゼ)遺伝子およびタンパク質と呼ばれる。α-ケトグルタル酸依存性ジオキシゲナーゼ酵素は、異なる化学クラスおよび作用の様式の除草剤を分解する能力を有することが以前には報告されていなかった。この非常に新規な発見は、顕著な除草剤耐性作物形質の機会ならびに選択マーカー技術の開発を基礎としている。本発明はまた、雑草を制御する関連方法も含む。本発明は、除草剤の新規な組み合わせが、新規な方法で使用されることを可能にする。さらに、本発明は、グリフォセートのような1種または複数の除草剤に対して抵抗性である(または天然により耐性である)雑草の形成を妨害し、かつその雑草を制御する新規な方法を提供する。 (もっと読む)
37℃での活性が増強されたI−DmoI誘導体及びその使用

37℃での開裂活性が増強されたI-DmoI誘導体、該変異形は、I-DmoIエンドヌクレアーゼの変異形又は第一I-DmoIドメインを少なくとも含むそのキメラ5誘導体の配列を含み、該配列は少なくとも:(i) 上記の第一I-DmoIドメインの位置4、20、49、52、92、94及び/又は95の残基の1つ、及び/又は(ii) I-DmoIのリンカー又は第二ドメインの始めの位置101、102及び/又は109の残基の1つ(存在する場合)の置換を含む。10上記の誘導体をコードするポリヌクレオチド、該ポリヌクレオチドを含む細胞、動物又は植物及び新規なDNA標的特異性を有するメガヌクレアーゼを単離するためのそれらの使用。 (もっと読む)
デンプンリン酸化酵素活性を有するタンパク質を同定するための方法
本発明は、デンプンのリン酸化に関わるタンパク質、およびそのようなタンパク質をコードする核酸を同定するための方法に関する。本発明はさらに、本発明による方法を用いて同定することができるタンパク質の変化した活性を示す植物細胞および植物に関する。この型の植物細胞および植物は、修飾されたデンプンを合成する。したがって、本発明はまた、本発明よる植物細胞および植物によって合成されるデンプン、ならびに、このデンプンの作製のための方法およびこの修飾デンプンのデンプン誘導体の作製に関する。 (もっと読む)
改善した生育特性を有する植物及びこれを作製する方法
本発明は、NAP1-様タンパク質をコードする核酸配列の発現をモジュレートすることによる、植物の生育特性を改善する方法に関する。本発明はまた、NAP1-様タンパク質をコードする核酸の発現がモジュレートされている、改善された生育特性を有するトランスジェニック植物に関する。  (もっと読む)
(もっと読む)
複数のデンプンリン酸化酵素の活性が増大した植物
本発明は、遺伝的に修飾された植物細胞および植物であって、遺伝的修飾が、遺伝的に修飾されていない対応する野生型植物細胞または野生型植物と比較して、デンプンリン酸化OK1タンパク質およびデンプンリン酸化R1タンパク質の活性の増大をもたらす、植物細胞および植物に関する。さらに、本発明はそのような植物細胞および植物の作製のための手段および方法に関する。この型の植物細胞および植物は、修飾デンプンを合成する。したがって、本発明はまた、本発明による植物細胞および植物によって合成されるデンプン、このデンプンの作製のための方法、およびこの修飾デンプンのデンプン誘導体の作製、ならびに本発明によるデンプンを含む穀粉に関する。さらに、本発明は、OK1タンパク質およびRIタンパク質をコードする配列を含む核酸分子およびベクター、ならびにこれらの核酸分子を含む宿主細胞に関する。 (もっと読む)
増大したデンプンリン酸化酵素活性を有する植物
本発明は、遺伝的に修飾された植物細胞および植物に関し、ここで遺伝的修飾は、遺伝的に修飾されていない対応する野生型植物細胞または野生型植物と比較して、OK1タンパク質のデンプンリン酸化活性の増大をもたらす。さらに本発明は、そのような植物細胞および植物を作製するための手段および方法に関する。これらの型の植物細胞および植物は、修飾デンプンを合成する。したがって、本発明はまた、本発明による植物細胞および植物から合成されたデンプン、これらのデンプンを作製する方法、およびこれらの修飾デンプンのデンプン誘導体の製造、ならびに本発明によるデンプンを含む穀粉に関する。さらに、本発明はまた、デンプンをリン酸化するOK1タンパク質をコードする核酸、そのような核酸分子を含むベクター、宿主細胞、植物細胞、および植物に関する。さらに本発明は、デンプンリン酸化活性を有するOK1タンパク質に関する。 (もっと読む)
遺伝子改変植物及びフィトレメディエーションにおけるそれらの使用
シュートに重金属を蓄積できる遺伝子改変植物、及び該遺伝子改変植物を用いて該重金属を除去し、できれば回収する方法。上記の遺伝子改変植物は、少なくともZn2+/Co2+/Cd2+/Pb2+サブクラスのP1B-タイプのATPアーゼをコードする配列の複数のコピーを含み、上記のP1B-タイプのATPアーゼを過剰発現する。 (もっと読む)
免疫抑制サイトカイン
EBI3-p35サイトカインが、サプレッサーT細胞に対する効果により、T細胞により媒介されるかまたは制御される免疫応答を阻害することができることが初めて示される。これは、EBI3-p35が関節炎、アテローム性動脈硬化症、移植片拒絶、および喘息などのアレルギーを含む様々な炎症状態および自己免疫状態において治療上有効であることを示唆している。 (もっと読む)
植物細胞における高効率ペプチド生産
本発明は、組換えペプチドの生産のための改善された方法を提供する。特に、本発明は、植物細胞懸濁培養においてインビボで集合され得るウイルスカプシド融合タンパク質の形態の組換えペプチドの生産のための改善された方法を提供する。本発明はまた、非感染性ウイルスカプシド融合ペプチド生産を可能にするプラスミド、配列及び植物細胞を含む。 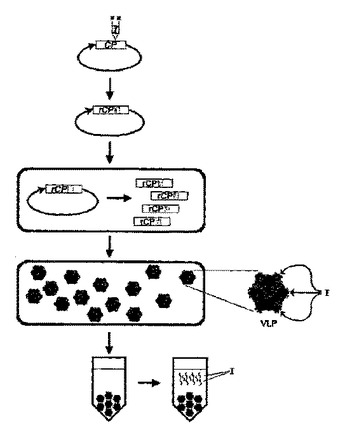 (もっと読む)
(もっと読む)
キチンオリゴ糖エリシター結合タンパク質
本発明者らは、APEA誘導体を用いたカラムの開発、非特異的吸着物質除去のためのプレカラム、効果的な溶出法を組み合わせることで収率良くエリシター結合タンパク質を単離精製した。これにより得られたN末端、及び内部鎖アミノ酸配列を利用して、イネcDNAライブラリーから本発明のタンパク質をコードするcDNAの単離に成功した。また、抗Con A-CEBiP抗体を精製し、エリシター応答性活性酸素生成に与える影響を調べたところ、該抗体で前処理することにより活性酸素生成は阻害され、本タンパク質がキチンオリゴ糖エリシター応答に関わる受容体タンパク質であることが示唆された。該エリシターは、イネにいもち抵抗性を誘導するので、本発明のタンパク質は、新規な病害防除技術の開発に応用できる。 (もっと読む)
生分解性ポリマー−ペプチド媒介トランスフェクションのための組成物および方法
ポリヌクレオチドと生分解性ポリアセタール-ペプチドとの組み合わせである複合体を開示する。開示する複合体は細胞トランスフェクションに有用である。 (もっと読む)
BacillusThuringiensisの新規な結晶ポリペプチド、ポリヌクレオチド、およびそれらの組成物
本発明は、Bacillus Cry2ポリペプチドに関連する殺虫性ポリペプチドを提供する。本発明のポリペプチドをコードする核酸もまた、提供される。昆虫による捕食に対する抵抗性を向上させる本発明のポリペプチドおよび核酸を使用するための方法が、包含される。本発明はまた、本発明の核酸および/またはポリペプチドを発現するトランスジェニック植物に関する。このトランスジェニック植物は、当該分野において公知である任意の方法で導入遺伝子を発現し得、この方法としては、構成的発現、発生的に制御された発現、組織特異的な発現などが挙げられるが、これらに限定されない。本発明のトランスジェニック植物から得られる種子もまた、本発明に包含される。  (もっと読む)
(もっと読む)
トリテルペン水酸化酵素
ソヤサポゲノールBは前駆体であるβ-アミリンの2段階の水酸化反応を経て生合成される。しかしながら、この反応に関与する水酸化酵素の遺伝子は明らかにされていなかった。そのため、水酸化酵素についての遺伝子工学的な利用が不可能であった。
発明者らは、ダイズ由来のシトクロームP450遺伝子CYP93E1に対応する配列がオレアナン型トリテルペンの24位を水酸化する酵素タンパクをコードしていることを明らかにするとともに、当該遺伝子を遺伝子工学的な手段を用いて利用する方法を提供する。
(もっと読む)
イチイ種(Taxussp.)における二次代謝を刺激するための、インダノイルアミドの使用
本発明は、インダノイルアミノ酸を含む栄養培地においてイチイ種の懸濁細胞を培養することによって、タキサンを生産するための方法を対象とする。インダノイルアミノ酸は、培養中の任意の時点に回分式で、または供給流中に添加してもよい。詳細には、合成化合物6−エチル−インダノイル−イソロイシン、6−ブロモインダノイルイソロイシン、および1−オキソ−インダン−カルボキシ−(L)−イソロイシン−メチルエステルアミド(1−OII)が、イチイ細胞培養物からのタキサン生産を増大させることが判明している。 (もっと読む)
ウキクサ中でのプラスミノ−ゲン及びミクロプラスミノ−ゲンの発現
本発明は、ウキクサ発現系中で組換えプラスミノゲン、ミクロプラスミノゲン、及びそれらの断片を生成するための方法及び組成物を提供する。ウキクサ発現系を使用して高レベルのプラスミノゲン及びミクロプラスミノゲンを生成することができることは、本発明の新規な発見である。ウキクサにおいて生成したプラスミノゲン及びミクロプラスミノゲンを活性化させて、プロテアーゼ活性を有するポリペプチドを生成することができる。したがって本発明は、プラスミノゲン、ミクロプラスミノゲン、及びその断片を発現させるための発現カセット、並びにプラスミノゲン、ミクロプラスミノゲン、及びその断片をコードするヌクレオチド配列を含む核酸を用いて形質転換したウキクサ、ウキクサ植物中でプラスミノゲン、ミクロプラスミノゲン、及びその断片を発現させるための方法を含み、これらのヌクレオチド配列を修飾してウキクサ中でのそれらの発現を増大させる。 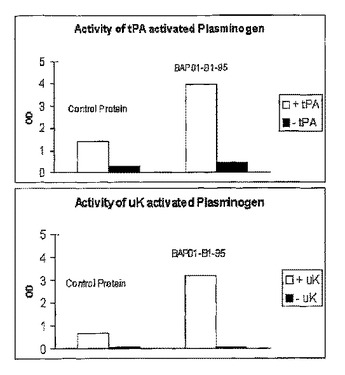 (もっと読む)
(もっと読む)
高リコペントマト品種及びその使用
本発明は、dark green(dg)遺伝子がホモ接合であり、現在市販されているトマト品種の果実の少なくとも2倍の平均リコペン含量を含む果実を産生する耐寒性トマト(Lycopersicon esculentum)品種に関するものであり、この品種は、商業規模での生育用に適合されており、その果実収穫物は、高い平均リコペン含量を維持している。本発明はさらに、生鮮果実及び加工果実の市場向けのトマトを作出するための、またリコペン及びそれを含む製品を製造するための前記品種の使用に関する。 (もっと読む)
受容体ポリペプチドを有する植物細胞
異種(例えばヒト)受容体ポリペプチドまたはその断片を含む新規なトランスジェニック植物細胞を開示する。これらの植物細胞を使用して、受容体ポリペプチドまたはその断片と相互作用することのできる分子(すなわちリガンド)を同定することが可能である。これらの植物細胞を使用して、トランスジェニック植物細胞に内因性のリガンド、または植物細胞に添加される外因性リガンドを同定することができる。このような受容体ポリペプチドリガンドは新規医薬の同定に使用される。 (もっと読む)
781 - 800 / 849
[ Back to top ]