
Fターム[4B024FA10]の内容
突然変異又は遺伝子工学 (218,933) | ベクターの改良、機能 (7,615) | マーカー (1,459)
Fターム[4B024FA10]に分類される特許
1,281 - 1,300 / 1,459
活性シナプスの可視化のための破傷風毒素融合タンパク質
活性シナプスを可視化する方法であって、前記方法が、(a)活性シナプスを形成する細胞を、少なくとも破傷風毒素のC断片およびリポータータンパク質を含むバイオマーカーに暴露するステップと、(b)当該バイオマーカーを可視化するステップとを含み、ここで細胞の樹状突起棘へのバイオマーカーの蓄積により活性シナプスの可視化が可能になる方法を提供する。また、シナプス活性をモジュレーションすることが可能な分子をスクリーニングする方法も提供する。神経変性疾患の早期診断に有用なキットは、少なくとも破傷風毒素のC断片およびリポータータンパク質を含むバイオマーカーを含む。 (もっと読む)
CD23に結合するペプチドおよびペプチド模倣体
本発明は、CD23に結合可能な新規のそして有用なペプチドおよびペプチド模倣体を含む化合物を記載する。これらは、自己免疫疾患、慢性炎症性疾患、アレルギー、および哺乳動物免疫系に仲介されるものなどの他の炎症状態に関連する炎症反応を減少させることが可能である。本発明の化合物は、CD23結合性ペプチドであって、前記ペプチドが、X1−X2−X3−X4−X5−X6−X7−X8のアミノ酸配列(式中、X1は、Pheであるか、または存在せず;X2は、HisまたはAlaであり;X3は、Glu、Ser、Ala、Asn、Lys、またはCysであり;X4は、Asn、Phe、Gln、Pro、Ser、またはAlaであり;X5は、Trpであり;X6は、Pro、Arg、Glu、Gly、Cys、またはLysであり;X7は、Ser、Pro、Leu、Thr、Ala、Gly、Asnであるか、または存在せず;そしてX8は、Phe、Glyであるか、または存在しない)を含む、前記ペプチドに関する。 (もっと読む)
HAUSP−Mdm2相互作用及びその使用
本発明は、新形成の診断、新形成治療の有効性の評価、新形成罹患対象者の予後評価のための方法を提供する。本発明はまた、新形成の検出で使用されるキットを提供する。本発明はさらに、細胞内でMdm2を脱ユビキチン化及び/又は安定化させる方法、及び細胞内でp53の脱ユビキチン化及び/又は安定性を調節する方法を提供する。さらにまた、本発明は、Mdm2-HAUSP相互作用の調節因子を同定する方法を提供する。さらにまた本方法で同定された調節因子、前記調節因子を含む医薬組成物、および新形成を治療する方法での前記調節因子の使用が提供される。本発明はさらに、Mdm2と反応する作用因子及びHAUSPと反応する作用因子を同定する方法を提供する。さらにまた、これらの方法によって同定された作用因子が提供される。最後に、本発明はMdm2及びHAUSPを含む複合体を提供する。 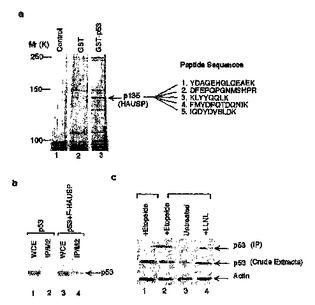 (もっと読む)
(もっと読む)
改善された相同的組換え効率を有する糸状菌変異体
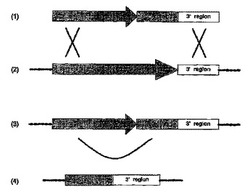
本発明は、NHR選好を有する糸状菌細胞のゲノムへのポリヌクレオチドの所定の部位に対する標的取込みの効率を増大させるための方法に関し、前記ポリヌクレオチドは、前記所定の部位との相同の領域を有し、取込み経路をHRへ導くステップを含んで成る。本発明は、親細胞に由来する変異体糸状菌にも関し、前記変異体は高い効率のHR経路および/または低い効率のNHR経路および/または同じ条件下の前記親細胞の前記HRおよび/またはNHR効率および/またはNHR/HR比と比べ減少した効率のNHR/HR比を有する。 (もっと読む)
デンプンリン酸化酵素活性を有するタンパク質を同定するための方法
本発明は、デンプンのリン酸化に関わるタンパク質、およびそのようなタンパク質をコードする核酸を同定するための方法に関する。本発明はさらに、本発明による方法を用いて同定することができるタンパク質の変化した活性を示す植物細胞および植物に関する。この型の植物細胞および植物は、修飾されたデンプンを合成する。したがって、本発明はまた、本発明よる植物細胞および植物によって合成されるデンプン、ならびに、このデンプンの作製のための方法およびこの修飾デンプンのデンプン誘導体の作製に関する。 (もっと読む)
生物工学的産生のために使用される、バシラス・リケニホルミス由来のRecA因子およびRecA不活性化安全な系統
本発明は、Bacillus licheniformis DSM 13 由来のRecA因子(配列番号:2)、と共にその関連遺伝子recA(配列番号:1)にも関し、関連タンパク質およびそれらの遺伝子、例えばそれらの中で、特に配列番号31および32として示した変異体を包含する。本発明に従って、遺伝子recAは、生物学的産生のために、それらの中でグラム陽性細菌の安全な系統を構築するために、その機能を不活性化することによって使用される。特定の実施態様において、該系統は、IV期の胞子形成遺伝子、好ましくは遺伝子spoIV(Bacillus licheniformis)、遺伝子yqfD(Bacillus subtilis)、または該系統が天然に胞子を形成できる場合はそれらに対して相同である各遺伝子のさらなる機能削除により提供される。さらに、新規RecAは、分子生物学的アッセイまたは細胞の分子生物学的活性について、特にDNA重合化または組み換え過程との関連において使用され得るタンパク質を示す。  (もっと読む)
(もっと読む)
成長因子、NsG33の治療上の使用
本発明は、蛋白質、遺伝子および細胞の治療上の使用に関する。より具体的には、本発明は、分泌型治療用蛋白質、NsG33の生物学的機能に基づく治療、特に、神経系の障害の処置についての治療に関する。NsG33は、ニューロン潜在能を有する細胞系のおける抗アポトーシス効果、および神経前駆細胞系および初代線条体培養物における神経保護および/または神経発生効果を有する、神経の生存および成長因子である。本発明はまた、新規対生物活性NsG33ポリペプチド断片およびその対応物をコードするDNA配列にも関する。 (もっと読む)
細胞内パラメータを改変することができる分子をスクリーニングするための発光タンパク質プローブを用いて、前記パラメータを検出するための方法
本発明は、標的細胞内のパラメータを改変することができる分子をスクリーニングするための発光組換えタンパク質プローブによって、前記細胞内パラメータを検出するための方法に関する。 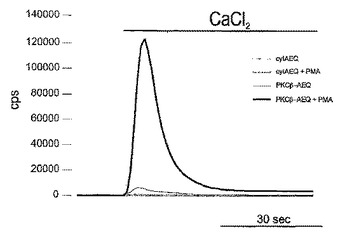 (もっと読む)
(もっと読む)
エキソ−エンドセルラーゼ融合構築体

本発明は異種エキソ−エンドセルラーゼ融合構築体に関する。該構築体は、糸状菌エキソセロビオハイドロラーゼ由来の触媒ドメイン及びエンドグルカナーゼ由来の触媒ドメインを含むセルロース分解活性を有する融合タンパク質をコードする。本発明は前記異種エキソ−エンドセルラーゼ融合タンパク質及びセルラーゼ酵素組成物を生産する方法だけでなく、異種エキソ−エンドセルラーゼ融合構築体を含むベクター及び宿主細胞にも関する。 (もっと読む)
視神経症に対する感受性を診断または予測するための方法
緑内障およびレーベル病を含む視神経症に連関する遺伝子多型のセットを開示する。これらの多型は視神経症に対する感受性を診断し予測するのに有用である。 (もっと読む)
選択的スプライシングを用いて真核細胞においてポリペプチドマルチマーを発現するための方法および構築物
本発明は単一の発現ベクターを用いて真核細胞において抗体または抗体フラグメントのような複数のポリペプチドを生成する方法を提供する。この発現ベクターは、単一のプロモーターの制御下に2つまたはそれより多い発現カセットを含むように操作され、この発現カセットは、その選択的スプライシングおよび望ましい比率での2つまたはそれより多い別個の遺伝子産物としての発現を可能にするスプライシング部位を有する。真核宿主細胞における組換え抗体の効率的な発現のためのベクターの使用ならびに診断適用および治療適用におけるかかる抗体の使用を開示する。 (もっと読む)
生物活性DNA結合部位及び関連する方法
本発明は、細胞中の目的のタンパク質が結合する生物活性DNA結合部位の同定に関する。本発明はまた、タンパク質が結合する生物活性DNA結合部位を変化させる薬剤及び条件の同定に関する。本発明の一態様はまた、転写調節因子によって調節される経路の同定方法及び経路の活性の調整方法を提供する。 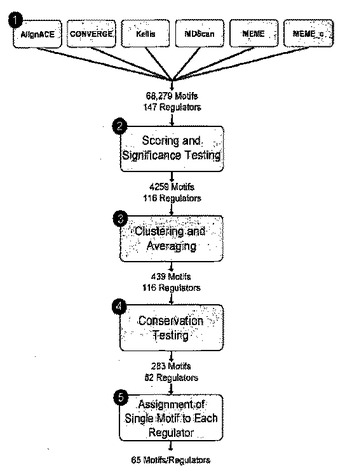 (もっと読む)
(もっと読む)
ポリペプチドの精製
大腸菌に対して異種性のポリペプチドを産生する方法であって、ポリペプチドをコードする核酸を含む大腸菌細胞が、核酸が発現するように、輸送可能な有機リン酸塩を培養液に与えながら培養液中で培養される方法が記載されている。次いで細胞からポリペプチドが回収される。 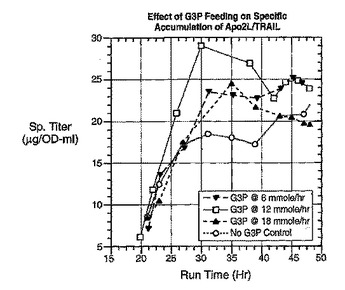 (もっと読む)
(もっと読む)
CD38スプライスバリアント及びその使用
本発明は、本明細書でCD38JLと称される、実質的に精製されたポリペプチドを提供する。CD38JLは、配列番号1のポリペプチド又はそのフラグメントを含むCD38のスプライスバリアントである。本発明はまた、CD38JLの発現に付随する疾患を治療、予防及び診断する方法を提供する。 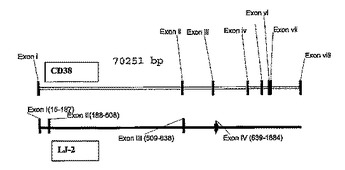 (もっと読む)
(もっと読む)
ステロイドホルモン受容体を調節するリガンドを産生またはスクリーニングする機能的方法
この出願は、核ホルモンリガンド活性化受容体スーパーファミリーのメンバーであるビタミンD3受容体(VDR)と、足場タンパク質(scaffolding protein)であるMNARとの相互作用の発見を示すものである。この相互作用の結果、VDR、MNARおよびSrcまたはチロシンキナーゼのファミリーであるPI3 キナーゼ間の三元複合体が形成され、細胞、特に骨芽細胞おけるシグナル伝達を媒介する。 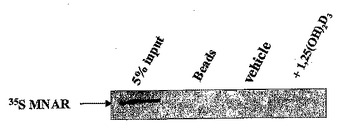 (もっと読む)
(もっと読む)
トランスジェニック植物における多価不飽和脂肪酸の製造方法
【課題】 多価不飽和脂肪酸を用いて食物および/または飼料の栄養価を高めるためには、植物系、特にトランスジェニック植物の種子において多価不飽和脂肪酸を簡便で安価に製造する方法が強く切望されている。
【解決手段】 本発明は、トランスジェニック植物の種子中に多価不飽和脂肪酸を産生させる方法に関する。この方法において、ω-3-デサチュラーゼ、Δ-12-デサチュラーゼ、Δ-6-デサチュラーゼ、Δ-6-エロンガーゼ、Δ-5-デサチュラーゼ、Δ-5-エロンガーゼおよび/またはΔ-4-デサチュラーゼ活性を有するポリペプチド、好ましくはΔ-6-デサチュラーゼ、Δ-6-エロンガーゼおよびΔ-5-デサチュラーゼ活性を有するポリペプチドをコードする核酸を生物に導入する。核酸配列は、配列番号11、配列番号27、配列番号193、配列番号197、配列番号199および配列番号201に示されている。有利に、これらの核酸配列は、場合により脂肪酸の生合成または脂質代謝のポリペプチドをコードする他の核酸配列と共に、生物中で発現させることができる。Δ-6-デサチュラーゼ、Δ-5-デサチュラーゼ、Δ-4-デサチュラーゼ、Δ-12-デサチュラーゼおよび/またはΔ-6-エロンガーゼ活性をコードする核酸配列は特に都合がよい。有利に、これらのデサチュラーゼおよびエロンガーゼは、珪藻(Thalassiosira)、ミドリムシ(Euglena)またはオストレオコッカス(Ostreococcus)に由来する。本発明はまた、増加した長鎖多価不飽和脂肪酸含量を有する油および/またはトリアシルグリセリドの製造方法に関する。好ましい実施形態において、本発明はまたアラキドン酸、エイコサペンタエン酸、またはドコサヘキサエン酸の製造方法に関し、増加した不飽和脂肪酸(特にアラキドン酸、エイコサペンタエン酸、および/またはドコサヘキサエン酸)含量を有するトリグリセリドを、トランスジェニック植物中、好ましくはトランスジェニック植物の種子中で産生させる方法に関する。本発明は更に、本発明の方法に用いるエロンガーゼおよびデサチュラーゼの発現に基づいて多価不飽和脂肪酸(特にアラキドン酸、エイコサペンタエン酸、および/またはドコサヘキサエン酸)含量の増加したトランスジェニック植物の製造に関する。本発明はまた、Δ-6-デサチュラーゼ、Δ-6-エロンガーゼ、Δ-5-デサチュラーゼおよびΔ-5-エロンガーゼ活性を有するポリペプチドをコードする核酸配列を一緒にまたは別々に含有する組換え核酸分子、ならびに、これらの組換え核酸分子を含有するトランスジェニック植物に関する。本発明のさらなる態様は、本発明の方法によって産生される油、脂質、および/または脂肪酸、ならびにこれらの使用に関する。さらに本発明は不飽和脂肪酸および不飽和脂肪酸含量の増大したトリグリセリド、ならびにこれらの使用に関する。
(もっと読む)
2種の染色体外エレメントを使用することによる原核細胞中での組換え遺伝子の生成
本発明は全体として、2種の異なる染色体外エレメントを用いることにより、原核細胞中で、具体的には細菌中で、組換えDNA配列を生成し検出する方法、および本発明の方法の実施に使用することができる染色体外エレメント、具体的にはプラスミドに関する。この方法が関係するDNA配列は、タンパク質コード配列および非コード配列を含む。  (もっと読む)
(もっと読む)
菌類における菌糸の育成
本発明は所望の菌糸成長に関係するタンパク質をコードしている核酸を修飾することを含む、タンパク質及び及び化学物質を菌類の宿主細胞の中で生産する、方法を提供する。hbrA及びhbrBのアミノ酸及び核酸配列も提供する。 (もっと読む)
L−アミノ酸の生産方法
腸内細菌科に属するL−アミノ酸生産細菌であって、pgl遺伝子(ybhE ORF)によりコードされる6−ホスホグルコノラクトナーゼの活性を増強することによりL−アミノ酸生産能を増大させた細菌を使用して、L−トリプトファン、L−フェニルアラニン及びL−チロシンなどのL−アミノ酸を生産する方法を開示する。 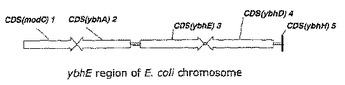 (もっと読む)
(もっと読む)
体細胞核初期化物質のスクリーニング方法
(a) ECAT遺伝子の発現調節領域により発現調節を受ける位置にマーカー遺伝子を存在させた遺伝子を含有する体細胞と被験物質とを接触させる工程、および(b) 前記(a)の工程の後、マーカー遺伝子発現細胞の出現の有無を調べ、該細胞を出現させた被験物質を体細胞の核初期化候補物質として選択する工程、を含む、体細胞の核初期化物質のスクリーニング方法等を提供する。 (もっと読む)
1,281 - 1,300 / 1,459
[ Back to top ]