
プラスミドの維持
本発明は、プラスミドの安定した維持のための系、この系において用いるための宿主細胞、ならびに医療への適用において有用なプラスミドを得るためにこの系を用いる方法に関する。特に、本発明は、インビボでの宿主細胞におけるプラスミドの安定した維持のための方法を提供する。例えば、本発明の宿主細胞は、形質転換された宿主細胞であって、i)細胞増殖を阻害する染色体遺伝子;およびii)アンチセンス配列をコードするプラスミド、を含み、該プラスミドによってコードされるアンチセンス配列は、該染色体遺伝子の作用を阻害し、それによって細胞増殖を許容する、形質転換された宿主細胞である。

【発明の詳細な説明】
【技術分野】
【0001】
(発明の分野)
本発明は、プラスミドの安定した維持のための系、この系においての使用のための宿主細胞、ならびに医療への適用において有用なプラスミドを得るためにこの系を用いる方法に関する。
【0002】
本明細書において引用される全ての文献は、参考として援用される。
【背景技術】
【0003】
(発明の背景)
プラスミドは、組換えタンパク質の調製においておよび遺伝子治療目的のためのDNAの調製において、慣用的に用いられる。宿主細胞内のプラスミドの安定した維持は、これらの産物の効率的な調製のために重要である。しかし、宿主細胞において運ばれる染色体外DNAは、プラスミドを含む細胞での、プラスミドが無い細胞に比べて増大した代謝的負荷に起因して、もともと不安定である。プラスミドの安定性を維持し代謝的負荷を軽減するため、プラスミドは、優性な選択マーカーを含むように操作されている。
【0004】
培養中の細胞内にプラスミドを維持する従来の方法は、抗生物質耐性遺伝子をそのプラスミドに含め、そして適切な抗生物質の存在中でその細胞を培養することである。治療用途が意図される細胞もしくはプラスミドに関して、このことは、抗生物質耐性遺伝子を含むプラスミドの使用が抗生物質耐性の拡がりの一因になり得るという不利益を有する。
【0005】
いくつかのプラスミド維持法は、プラスミドによって運ばれる遺伝子(plasmid−borne gene)によって制御される天然に存在する分離後殺傷(post segregational killing)機構を利用することを試みてきた。例えば、hok/sok系、srnB系、およびpnd系は、キラー(killer)タンパク質を含み、そのキラータンパク質は、安定なmRNAによってコードされ、キラーRNAに結合してキラーRNAを不活性化する小さい不安定なアンチセンスRNAによって調節される。キラーRNAは、アンチセンスRNAが分解された後、プラスミドが無い分離体(segregant)において保持され、致死タンパク質へと翻訳される。hok/sok系を用いるプラスミド維持は、弱毒化生ベクターワクチン株Salmonella typhi CDV908 htr−A(非特許文献1)において研究された。しかし、そのような分離後殺傷機構は、形質転換後のプラスミド選択を可能とせず、ゆえに依然としてプラスミドにおける抗生物質耐性遺伝子の存在に依存している。
【0006】
抗生物質選択を用いずにプラスミドを維持するためおよびプラスミドを選択するためのいくつかの代替的な方法が開発されてきており、それらの方法においては、プラスミドが宿主細胞の栄養要求性を補完する遺伝子をコードする。例えば、宿主細胞は、必須アミノ酸代謝産物を合成することが出来ず、このアミノ酸の合成のための失われたエレメントをコードしている遺伝子を含むプラスミドが存在する場合においてのみ、そのアミノ酸を欠乏している培地中で生存し得る、変異細胞であり得る(非特許文献2)。しかし、このアプローチは、そのアミノ酸が省かれなければならないために、増殖培地の組成を制限する。複雑な培地において用いられ得る代替的な方法は、温度感受性のtRNAシンテターゼ遺伝子を有する変異宿主細胞を用いる。この変異宿主細胞は、野生型tRNAシンテターゼ遺伝子を含むプラスミドが存在する場合のみ、非許容(non−permissive)温度で生存し得る(非特許文献3)。別の選択方法は、変異宿主細胞における必須な染色体遺伝子中のナンセンス変異を補完するプラスミドによって運ばれるtRNA遺伝子を用いる(非特許文献4)。あるいは、pilオペロンのような、細胞に対する代謝的負荷を増大する遺伝子が、対応するリプレッサータンパク質をコードするプラスミドの存在する場合においてのみ宿主細胞が生存するように、宿主の染色体に配置され得る(非特許文献5)。
【0007】
特許文献1は、インビトロ培養中の宿主細胞内のプラスミドをオペレーターリプレッサータイトレーション(operator repressor titration)によって維持する方法を説明している。その方法は、その宿主細胞が、リプレッサーをコードする第一の染色体遺伝子、および、その調節領域においてリプレッサーについてのオペレーター配列を有する細胞増殖のために必須である第二の染色体遺伝子を含むようにする、宿主細胞の操作を伴う。プラスミドが存在しない場合、第二の染色体遺伝子の発現は、リプレッサーのオペレーターへの結合によって阻害され、そしてその細胞は死ぬ。この宿主細胞における維持のためのプラスミドは、オペレーター配列を含むように操作されており、その結果、プラスミドが存在する場合、リプレッサーは細胞増殖のために必須である遺伝子のオペレーターから結合除去(titrate away)され、その遺伝子が発現されその細胞が生存する。この機構はまた、Williamsら(非特許文献6)に、およびCranenburghら(非特許文献7)に、記載される。
【特許文献1】欧州特許第0851932号明細書
【非特許文献1】Galenら、Infect.Immunol.,1999年、第67巻、p.6424−6433
【非特許文献2】Wang M−Dら、J.Bacteriol.,1987年、第169巻、p.5610−5614
【非特許文献3】Skogmanら、Gene,1984年、第31巻、p.117−122
【非特許文献4】Zengelら、J.Bacteriol.,1981年、第145巻、p.459−465
【非特許文献5】Ogdenら、Biotech.Bioeng.,1992年、第40巻、p.1027−1038
【非特許文献6】Williamsら、Nucleic Acids Research,1999年、第26巻、第9号、p.2120−2124
【非特許文献7】Cranenburghら、Nucleic Acids Research,2001年、第29巻、第5号、p.e26−e27
【発明の開示】
【発明が解決しようとする課題】
【0008】
抗生物質選択に依存しないプラスミドの維持および選択のいくつかの機構は公知であるが、治療への適用のためのDNAおよび組換えタンパク質の産生におけるプラスミドの重要性が増大していることを考慮して、さらなる方法の開発についての必要性は存続する。加えて、今までに開発された、プラスミドの維持および選択の系は、これらの系における使用のために特別に改変されているプラスミドの使用を必要とする。抗生物質耐性を伴わず、当該分野において一般的であり特別な改変を必要としないプラスミドを使用する、プラスミドの維持および選択の系についての必要性が存続する。
【課題を解決するための手段】
【0009】
(発明の要旨)
本発明の第一の局面に従って、形質転換された宿主細胞が提供される。その宿主細胞は、
i)細胞増殖を阻害する染色体遺伝子;および
ii)アンチセンス配列をコードするプラスミド
を含み、上記プラスミドによってコードされるアンチセンス配列は、上記染色体遺伝子の作用を阻害し、それによって細胞増殖を許容する。
【0010】
上記プラスミドによってコードされるアンチセンス配列は、このプラスミドの他の領域によってコードされてもよいが、好ましくはこのプラスミドの複製起点によってコードされる。
【0011】
本明細書において用いられる場合、用語「アンチセンス配列」とは、核酸配列であって、その核酸配列の標的配列に対して実質的に相補的であり、その標的配列に特異的にハイブリダイズする能力を有する核酸配列を、指す。好ましくは、上記のプラスミドによってコードされるアンチセンス配列は、ストリンジェントな条件下でそのアンチセンス配列の標的配列にハイブリダイズする。高ストリンジェンシーハイブリダイゼーション条件は、以下のように定義される:50%のホルムアミド、5×SSC(150mMのNaCl、15mMのクエン酸三ナトリウム)、50mMのリン酸ナトリウム(pH7.6)、5×デンハルト液、10%のデキストラン硫酸、および20マイクログラム/mlの変性剪断されたサケ精子DNAを含む溶液中での、42℃での一晩のインキュベーションと、その後の0.1×SSC中でのおよそ65℃でのフィルターの洗浄。
【0012】
本明細書において用いられる場合、「細胞増殖」とは、培養培地中の細胞数の経時的な増加、および、生細胞の数が経時的に減少しない場合は、細胞の生存率を、指す。「細胞増殖を阻害する」によって、染色体遺伝子が細胞にとって致死性であり、その結果、培養培地中の生細胞の数が経時的に減少すること、または、染色体遺伝子が細胞増殖を妨げ、その結果、培養培地中の細胞の数が時間にわたって増加しないことが、意味される。
【0013】
本発明のこの第一の局面に従う形質転換された宿主細胞の増殖は、プラスミドの存在に依存しており、このプラスミドを維持する細胞の選択をもたらす。宿主細胞からこのプラスミドが失われた場合、この染色体遺伝子の作用はもはやプラスミドによってコードされるアンチセンス配列によって妨げられず、細胞増殖が阻害される。いくつかの実施形態において、この染色体遺伝子の発現が阻害され、それによって細胞増殖を許容するためには、ほんの1コピー(copy)のこのプラスミドが必要であり得る。他の実施形態において、染色体遺伝子の発現が阻害されるために、プラスミドが複数コピーで存在することが必要であり得る。
【0014】
この染色体遺伝子は、細胞増殖を直接的に阻害し得る。例えば、この染色体遺伝子の転写および翻訳は、トキシンのような、細胞にとって致死性のタンパク質を産生し得る。あるいは、この染色体遺伝子は、細胞増殖を間接的に阻害し得る。例えば、この染色体遺伝子の転写および翻訳は、第二の染色体遺伝子によってコードされる細胞増殖のために必須であるタンパク質の転写および翻訳を阻害するリプレッサータンパク質を、産生し得る。この染色体遺伝子の転写はまた、細胞増殖のために必須な第二の染色体遺伝子の転写および/もしくは翻訳を、第二の染色体遺伝子に結合することまたは第二の染色体遺伝子から転写されるmRNAに結合することのいずれかによって阻害するアンチセンス配列を、産生し得る。
【0015】
いくつかの実施形態において、この染色体遺伝子は、より複雑な反応カスケードによって細胞増殖を阻害し得る。例えば、この染色体遺伝子は、第二の染色体遺伝子の発現を活性化するアクチベータータンパク質をコードし得、この第二の染色体遺伝子は、次に、細胞増殖のために必須なタンパク質をコードする第三の染色体遺伝子の発現を阻害するリプレッサータンパク質をコードする。あるいは、第一の染色体遺伝子は、第二の染色体遺伝子のアンチセンスインヒビターをコードし得、この第二の染色体遺伝子のアンチセンスインヒビターは、次に、細胞増殖を阻害するタンパク質をコードする第三の染色体遺伝子の発現を抑制するリプレッサータンパク質をコードする。そのようなカスケードは三つより多くの染色体遺伝子を含み得る。例えば、そのようなカスケードは、4、5、6、7もしくはそれより多くの染色体遺伝子を含み得る。そのようなカスケードの全てにおいて、第一の染色体遺伝子を阻害するプラスミドが存在しないことにより、第一の染色体遺伝子にカスケード反応を開始させ、このことから最終的に細胞増殖の阻害がもたらされる。細胞増殖を阻害する適切な染色体遺伝子の例は、本明細書において提供される。
【0016】
本発明の形質転換された宿主細胞は、好ましくは、治療用途が意図されるプラスミドを維持するために用いられる。従って、このプラスミドが目的の遺伝子の挿入のためのクローニング部位を含むことが、好ましい。好ましくは、このプラスミドはさらに、目的の遺伝子を含む。この目的の遺伝子は哺乳類細胞において、好ましくはヒト細胞において、発現し得ることが好ましい。この目的の遺伝子は、治療用途のために、目的のRNAもしくは目的のタンパク質を発現し得る。プラスミドに含まれ得るそのような目的の遺伝子の例は、当該分野において公知であり、本明細書において提供される。
【0017】
上記プラスミドによってコードされるアンチセンス配列は、上記染色体遺伝子自体に結合することによってこの染色体遺伝子の作用を阻害し得、それによってこの遺伝子の転写を阻害し得る。あるいは、上記プラスミドによってコードされるアンチセンス配列は、上記染色体遺伝子から転写されるmRNAに結合することによってこの染色体遺伝子の作用を阻害し得る。この染色体遺伝子の性質に依存して、この染色体遺伝子から転写されるmRNAへの結合は、この染色体遺伝子の翻訳を阻害し得るか、もしくはこの染色体遺伝子にコードされるアンチセンス配列がこのアンチセンス配列の標的へ結合するのを妨ぎ得る。
【0018】
本発明の第一の局面の好ましい実施形態において、調節配列は、上記染色体遺伝子と作動的(operatively)に連結される。「作動的に連結される」によって、この調節配列が、読み枠をあわせて(in−frame)この染色体遺伝子と連結され、その結果、この調節配列がこの染色体遺伝子と同時に転写されることが、意味される。好ましくは、この調節配列は、この染色体遺伝子の上流に作動的に連結される。しかし、あるいはこの調節配列は、この染色体遺伝子の下流に作動的に連結されてもよい。この調節配列がこの染色体遺伝子の上流に作動的に連結された場合、この調節配列は、好ましくはリボソーム結合部位内に挿入される。あるいは、この調節配列は、リボソーム結合部位の上流に挿入されてもよい。この染色体遺伝子もしくは調節遺伝子に作動的に連結されたこの染色体遺伝子は、構成的プロモーターもしくは誘導性プロモーターの制御下にあってもよい。好ましくは、この染色体遺伝子もしくは調節遺伝子に作動的に連結されたこの染色体遺伝子は、構成的プロモーターの制御下にある。
【0019】
上記プラスミドによってコードされるアンチセンス配列は、この調節配列に結合することによってもしくはこの調節配列から転写されたmRNAに結合することによって、上記染色体遺伝子の作用を阻害し得る。本発明のこの実施形態において、上記プラスミドによってコードされるアンチセンス配列は、好ましくはこのプラスミドの複製起点によってコードされる。
【0020】
全てのプラスミドの複製起点は、転写されたRNAを産生する。いくつかの場合において、プラスミド(例えば、プラスミドR1、プラスミドRK6、プラスミドpT181、プラスミドpMV158、およびプラスミドpIP501)の複製起点から転写されたRNAは、複製の調節に直接的に用いられる。他の場合において、転写されたRNAは、翻訳されてプラスミド(例えば、pSC101、pPS10、p15A、F、R100、R453、P1、RK2、RA1、RSF10110、pColIV−K30、ColE2、COlE3、Rts1、pLS20、およびpUB110)の複製のために必要とされるタンパク質を提供する(Solarら、1998,Microbiol and Molec.Biol.Rev.62:434−363を参照)。
【0021】
本発明は、上記プラスミドによってコードされるアンチセンス配列がこれらのプラスミドのうちのいずれか一つの複製起点から転写されるRNAである、形質転換された宿主細胞を含む。この実施形態によると、上記染色体遺伝子に作動的に連結された調節配列、もしくはこの調節配列から転写されるmRNAは、このプラスミドの複製起点から転写されるRNAに対してアンチセンスである。このプラスミドが存在しない場合において、この染色体遺伝子に作動的に連結された調節配列は、転写され、そして/もしくは翻訳され、細胞増殖の阻害をもたらす。プラスミドが存在する場合において、複製起点から転写されるRNAは、この染色体遺伝子の作用を阻害し、それによって細胞増殖を可能にする。
【0022】
本発明のこの局面の好ましい実施形態によれば、上記プラスミドによってコードされるアンチセンス配列は、RNAIであり、上記染色体遺伝子の上流にある調節配列は、RNAIIもしくはその一部をコードする。あるいは、このプラスミドによってコードされるアンチセンス配列は、RNAIIであり、この染色体遺伝子の上流にある調節配列は、RNAIもしくはその一部をコードする。
【0023】
RNAIおよびRNAIIは、研究用途および商業的用途の事実上全てのプラスミドの複製起点によってコードされる、二つの重なるRNA転写産物である。RNAIIは、プラスミドDNA合成の開始を始め、E.coliにおいて自律的プラスミド複製がおこることを可能にする。RNAIは、RNAIIのアンチセンスインヒビターであり、過剰のプラスミド複製を妨げ、プラスミドのコピー数が、宿主細胞によって支持され得る数を上回らないことを保証する。現在、このアンチセンス機構が、形質転換された宿主細胞におけるプラスミド維持を促進するために利用され得ることが、見出されている。
【0024】
この実施形態によれば、本発明は、形質転換された宿主細胞を提供する。この宿主細胞は、
i)染色体遺伝子に作動的に連結された、RNAIIコード領域もしくはRNAIIコード領域の一部であって、その染色体遺伝子は細胞増殖を阻害する、RNAIIコード領域もしくはRNAIIコード領域の一部:および
ii)RNAIコード領域もしくはRNAIコード領域の一部を含む、プラスミド、
を含み、上記RNAIコード領域もしくは上記RNAIコード領域の一部から転写されるRNAIは、上記RNAIIコード領域もしくは上記RNAIIコード領域の一部から転写されるRNAIIに結合し、上記染色体遺伝子の作用を阻害し、それによって細胞増殖を許容する。
【0025】
上記染色体遺伝子に作動的に連結されたRNAIIコード領域もしくはRNAIIコード領域の一部は、転写されて、RNAII−染色体遺伝子mRNAが生成される。RNAIをコードするプラスミドが存在しない場合において、細胞増殖は阻害される。細胞増殖は、このRNAII−染色体遺伝子mRNAが、細胞増殖のために必須の第二の染色体遺伝子もしくはそのような第二の染色体遺伝子から転写されるmRNAに結合することによって阻害され得る。あるいは、このRNAII−染色体遺伝子は翻訳され得、そしてこの染色体遺伝子によってコードされる毒性タンパク質により、もしくは、細胞増殖のために必須の第二の染色体遺伝子を阻害する、この染色体遺伝子によってコードされるリプレッサータンパク質により、細胞増殖が阻害され得る。このプラスミドが存在する場合において、このプラスミドから転写されたRNAIは、RNAII−染色体遺伝子mRNA中のRNAIIに結合し、染色体遺伝子が細胞増殖を阻害することを防ぐ。
【0026】
本発明はさらに、形質転換された宿主細胞を提供する。この宿主細胞は、
i)染色体遺伝子に作動的に連結されたRNAIコード領域もしくはRNAIコード領域の一部であって、この染色体遺伝子は細胞増殖を阻害する、RNAIコード領域もしくはRNAIコード領域の一部:および
ii)RNAIIコード領域もしくはRNAIIコード領域の一部を含む、プラスミド、
を含み、上記RNAIIコード領域もしくはRNAIIコード領域の一部から転写されるRNAIIは、上記RNAIコード領域もしくはRNAIコード領域の一部から転写されるRNAIに結合し、上記染色体遺伝子の作用を阻害し、それによって細胞増殖を許容する。
【0027】
上記の染色体遺伝子に作動的に連結されたRNAIコード領域もしくはRNAIコード領域の一部は、転写されて、RNAI−染色体遺伝子mRNAを生成する。RNAIIをコードするプラスミドが存在しない場合において、細胞増殖は阻害される。細胞増殖は、このRNAI−染色体遺伝子mRNAが、細胞増殖に必須な第二の染色体遺伝子もしくはそのような第二の染色体遺伝子から転写されるmRNAに結合することによって、阻害され得る。あるいは、このRNAI−染色体遺伝子は翻訳され得、この染色体遺伝子によってコードされる毒性タンパク質によって、もしくは、細胞増殖に必須な第二の染色体遺伝子を阻害する、この染色体遺伝子によってコードされるリプレッサータンパク質によって、細胞増殖が阻害され得る。このプラスミドが存在する場合、このプラスミドから転写されるRNAIIは、RNAI−染色体遺伝子mRNA中のRNAIに結合し、この染色体遺伝子が細胞増殖を阻害することを妨げる。
【0028】
全てのプラスミドは、複製起点から転写されるRNAを産生し、ほとんどの商業的に入手し得るプラスミドは、RNAIコード領域およびRNAIIコード領域を含む複製起点を含む。用いられるプラスミド内へのlacオペレーター系のクローニングにプラスミドの維持を依存している、EP0851932において開示される系とは異なり、本発明は、形質転換される宿主細胞における、任意の複製起点を含む任意の商業的に入手し得るプラスミドの維持を可能にし、これらのプラスミド内に任意のさらなる配列をクローニングすることを必要とせずに、これらのプラスミドを維持する宿主細胞の選択を可能にする。このことは、プラスミドの大きさが縮小されて、DNAワクチンおよび遺伝子治療への適用におけるプラスミドの有効用量を増加するという利点を有する。不必要な細菌DNAの除去もまた、CpGジヌクレオチドに対する、免疫応答の危険性を低下させる。
【0029】
加えて、プラスミドの維持は、抗生物質選択を用いずに達成され得る。このプラスミドは、それゆえに、抗生物質耐性遺伝子の存在を必要としない。このプラスミドに抗生物質耐性遺伝子が存在しないことは、特にこのプラスミドもしくはこのプラスミドによってコードされる産物が治療に用いられることが意図される状況において、多くの利点を有している。第一に、このプラスミドに抗生物質耐性遺伝子が存在しないことは、抗生物質耐性遺伝子が潜在的な病原体を含めた環境内生物体に移動する危険性を取り除く。このプラスミドに抗生物質耐性遺伝子が存在しないことは、また、細胞から単離されたプラスミドがプラスミド選択のために用いられた残留抗生物質によって汚染されることを防止し、従って、残留抗生物質の存在についてプラスミドを試験する必要性、および抗生物質耐性遺伝子が患者の細胞において有害な効果をもたらす危険性を、取り除く。選択が抗生物質耐性に依存しないという事実は、培養中の抗生物質分解に起因するプラスミド選択圧の欠如の問題を回避する。
【0030】
本発明において使用するためのプラスミドとしては、複製起点から転写されるRNAが複製の調節に直接的に用いられるプラスミド(例えば、プラスミドR1、RK6、pT181、pMV158、およびpIP501)、および、複製起点から転写されるRNAが翻訳されてプラスミドの複製のために必要とされるタンパク質を提供するプラスミド(例えば、pSC101、pPS10、p15A、F、R100、R453、P1、RK2、RA1、RSF10110、pColIV−K30、ColE2、ColE3、Rts1、pLS20、およびpUB110)が挙げられる。好ましくは、本発明において使用するためのプラスミドは、RNAIコード領域およびRNAIIコード領域を含む複製起点を、含む。RNAIコード領域およびRNAIIコード領域を含む複製起点を含むプラスミドとしては、ColE1oriおよびpMB1oriのような、ColE1型oriを含むプラスミドが挙げられる。
【0031】
細胞増殖を阻害する染色体遺伝子に作動的に連結された、RNAIIコード領域、RNAIコード領域、もしくはそれらの一部は、構成的プロモーターもしくは誘導性プロモーターの制御下にあってもよい。好ましくは、細胞増殖を阻害する染色体遺伝子に作動的に連結された、RNAIIコード領域、RNAIコード領域、もしくはそれらの一部は、構成的プロモーターの制御下にある。適切な構成的プロモーターは、当該分野において公知であり、適切な構成的プロモーターとしては、例えば、trcプロモーターのようなE.coliプロモーターからの−35配列および−10配列を含むプロモーターが挙げられる。
【0032】
本発明の第二の局面によれば、細胞増殖を阻害する染色体遺伝子を含む宿主細胞が提供され、この染色体遺伝子は、その上流に位置する調節配列に作動的に連結されており、この調節配列は、プラスミドの複製起点から転写されるRNAの配列に対してアンチセンスである配列を含むか、もしくは、この調節配列は、プラスミド複製起点から転写されるRNAの配列に対してアンチセンスであるRNA配列をコードする。この調節配列の性質は、この宿主細胞が形質転換されることが意図されるプラスミドの特性に依存する。適切なプラスミド複製起点の例は、上記に提供される。これらの複製起点から転写されるRNAに対してアンチセンスである調節配列を設計する方法は、当業者にとって明らかである。
【0033】
好ましくは、この調節配列は、RNAIコード領域もしくはRNAIコード領域の一部、またはRNAIIコード領域もしくはRNAIIコード領域の一部を含む。細胞増殖を阻害する染色体遺伝子に作動的に連結された、これらのRNAIIコード領域、RNAIコード領域、もしくはそれらの一部は、構成的プロモーターもしくは誘導性プロモーターの制御下にあってもよい。好ましくは、細胞増殖を阻害する染色体遺伝子に作動的に連結された、これらのRNAIIコード領域、RNAIコード領域、もしくはそれらの一部は、構成的プロモーターの制御下にある。適切な構成的プロモーターは、当該分野において公知であり、適切な構成的プロモーターとしては、例えば、trcプロモーターのような、E.coliプロモーターからの−35配列および−10配列を含むプロモーターが挙げられる。
【0034】
本発明のこの第二の局面による宿主細胞は、本発明の第一の局面による形質転換された宿主細胞を産生するために、RNAIコード領域およびRNAIIコード領域を含むプラスミドを用いて形質転換されてもよい。
【0035】
本発明のこの宿主細胞および形質転換された宿主細胞は、インビトロにおいて培養中であってもよい。適切な宿主細胞の例は、本明細書中に提供される。
【0036】
本発明の第三の局面によれば、インビトロで宿主細胞内にプラスミドを維持する方法が提供され、この方法は、上記において記載されるように形質転換された宿主細胞を、この細胞が増殖することを許容するのに十分な条件下で培養する工程を含む。この方法は、抗生物質耐性が存在しない場合において、プラスミドの維持および選択が行われることを可能にし、このことは、この方法を治療への適用のためのプラスミドDNAもしくは組換えタンパク質の産生のために特に適切にする。本発明の第四の局面によれば、プラスミドDNAを産生する方法が提供され、この方法は、本発明の第三の局面の方法をおこなう工程およびプラスミドDNAを単離する工程を包含する。本発明の第五の局面によれば、組換えタンパク質を産生する方法が提供され、この方法は、本発明の第三の局面の方法に従う目的のタンパク質をコードするプラスミドを含む形質転換された宿主細胞を培養する工程、およびそのタンパク質をその細胞から単離する工程、を包含する。
【0037】
本発明の形質転換された宿主細胞および宿主細胞は、それら自体が治療への適用を有し得る。本発明の第六の局面によれば、治療における使用のための、本発明の第一の局面による形質転換された宿主細胞が提供される。
【0038】
本発明の第七の局面によれば、薬学的組成物が提供され、この薬学的組成物は、本発明の第一の局面による形質転換された宿主細胞と、それに加えて、薬学的に受容可能な賦形剤、希釈剤もしくは緩衝剤(例えば、当業者にとって周知である薬学的に受容可能な賦形剤、希釈剤もしくは緩衝剤)を含む。この局面の一つの実施形態によれば、この薬学的組成物は、ワクチン組成物である。このワクチン組成物は、賦形剤、希釈剤もしくは緩衝剤に加えて、一つ以上のアジュバントを含んでもよい。
【0039】
本発明の第八の局面によれば、薬の製造において、本発明の第一の局面による形質転換された宿主細胞の使用が、提供される。好ましくは、この形質転換された宿主細胞は、遺伝子送達系として用いられてもよい。例えば、この形質転換された宿主細胞は、遺伝子治療目的でプラスミドにある目的の遺伝子を送達するために用いられてもよい。この形質転換された細胞はまた、プラスミドにある目的の(その転写産物が治療的効果を有する)遺伝子(例えば、アンチセンスオリゴヌクレオチド)を送達するために、もしくは治療的効果を有するタンパク質をコードする目的の遺伝子を送達するために、用いられてもよい。
【0040】
あるいは、この形質転換された宿主細胞は、免疫原を送達するために用いられてもよい。この実施形態において、プラスミドにある目的の遺伝子は、免疫応答を惹起する一つ以上の抗原をコードしてもよい。当業者は、免疫応答を惹起する抗原をコードしていて本発明の形質転換された細胞内のプラスミドに含まれ得る、目的の遺伝子が分かる。
【0041】
本発明の第九の局面によれば、処置の方法が提供され、この処置の方法は、上記に記載された形質転換された宿主細胞、もしくはそれらを含む組成物を患者に投与する工程を、包含する。特に、目的の遺伝子を患者に送達する方法が提供され、この方法は、上記に記載される形質転換された宿主細胞もしくはそれらを含む組成物を患者に投与する工程を、包含する。上記に記載されるように、形質転換された宿主細胞内のプラスミドにある目的の遺伝子は、それ自体が遺伝子治療目的のために有用であってもよく、または、この目的の遺伝子の転写産物もしくは転写および翻訳の産物が、治療的特性を有してもよい。
【0042】
本発明の第十の局面によれば、病原体によって引き起こされる疾患に対して患者を免疫化する方法が提供され、この方法は、上記に記載される形質転換された宿主細胞もしくはそれらを含む組成物を患者に投与する工程を、包含する。この細胞内のプラスミドにおいて見出される目的の遺伝子は、この病原体に対する免疫応答を誘導する抗原をコードする。細胞の性質および目的の遺伝子の発現を制御するプロモーターの性質によって、この抗原はこの細胞が患者に投与される前にこの細胞において発現されてもよく、もしくは、投与後に患者において発現されるこの抗原と共に、このプラスミドを含む細胞が患者に投与されてもよい。好ましくは、上記で記載したように、この細胞は細菌細胞であり、この目的の遺伝子は、細菌のプロモーターもしくは真核細胞のプロモーターのいずれかに機能的に連結される。
【0043】
本発明の第十一の局面によれば、インビボの宿主細胞内にプラスミドを維持する方法が提供され、この方法は、本発明の第一の局面による形質転換された宿主細胞をレシピエント生物体に供給する工程を包含する。好ましくは、このレシピエント生物体は哺乳類、好ましくはヒトである。本発明のこの局面によれば、上記染色体遺伝子は、好ましくは、インビボで細胞増殖を阻害し、その結果、このプラスミドを維持する宿主細胞のみが、レシピエント生物体においてインビボで生存する。例えば、この染色体遺伝子は、この細胞の完全性にとって必須であるタンパク質をコードする第二の染色体遺伝子についてのリプレッサータンパク質をコードしてもよい。
【0044】
本発明における使用のために適切なプラスミドに運ばれる目的の遺伝子、宿主細胞、および細胞増殖を阻害する染色体遺伝子の例、ならびに本発明の形質転換された宿主細胞の医療への適用についての情報は、以下に提供される。
【発明を実施するための最良の形態】
【0045】
(染色体遺伝子)
本発明の宿主細胞および形質転換された宿主細胞内に含まれる染色体遺伝子は、増殖を阻害する任意の遺伝子であり得る。
【0046】
(増殖を直接的に阻害する染色体遺伝子)
細胞増殖を直接的に阻害する好ましい染色体遺伝子としては、トキシンをコードする遺伝子が挙げられる。本発明において用いられ得る、トキシンをコードする遺伝子の例としては、B.subtilisからのsacB遺伝子、E.coliのFプラスミド内のccdBによってコードされるCcdB、プラスミドRK2からのparD遺伝子によってコードされるKidタンパク質、制限エンドヌクレアーゼ遺伝子および抗生物質遺伝子が挙げられる。
【0047】
(増殖のために必須の遺伝子を阻害することによって増殖を間接的に阻害する染色体遺伝子)
増殖を間接的に阻害する好ましい染色体遺伝子としては、細胞増殖のために必須の第二の染色体遺伝子を阻害するタンパク質リプレッサーをコードする染色体遺伝子、および、細胞増殖のために必須の第二の染色体遺伝子を阻害するアンチセンス配列をコードする染色体遺伝子が挙げられる。本発明の一つの実施形態において、この第二の染色体遺伝子は、細胞増殖のために条件的に必須である。
【0048】
この染色体遺伝子がタンパク質リプレッサーをコードする場合、第二の染色体遺伝子は、好ましくは、そのリプレッサーによって従来的に阻害されるオペレーターおよびプロモーターに作動的に連結される。
【0049】
例えば、この染色体遺伝子がタンパク質リプレッサーLacIをコードするならば、細胞増殖のために必須の第二の染色体遺伝子は、好ましくはlacオペレーターおよびlacプロモーターに作動的に連結される。E.coliのlacリプレッサーは、「The Lactose Operon」(J.Beckwith、Escherichia coli and Salmonella typhimurium、J.L.Ingrahamら編、1987年、Amer.Soc.Micro.、1444〜1452頁)およびDicksonら(1975年、Science 187:27−35)において記載される。lacオペロンは以下のように調節される。非誘導条件下(例えば、ブドウ糖での増殖)では、LacIはlacオペロンのオペレーターに結合し、β−ガラクトシダーゼ(LacZ)、ラクトースパーミアーゼ(LacY)、およびトランスアセチラーゼ(LacA)の転写を妨げる。誘導性条件下(例えば、ラクトースでの増殖、もしくは代謝できないアナログであるIPTGの添加)では、リプレッサーはもはやオペレーターに結合せず、転写が起こる。オペロンの発現は、β−ガラクトシダーゼについてのアッセイによって容易に検出される。
【0050】
本発明に従う有用な他のリプレッサー系としては、真核細胞における遺伝子活性の調節において用いるためのtetリプレッサー系(Gossenら(1994)Current Opinions in Biotechnology,5,516−520)が挙げられる。tetリプレッサー系は、酵母、タマホコリカビ(dictiostelium)、植物細胞およびタバコ植物において用いられている。本発明に従うさらなる有用なリプレッサー系は、ArgRNVリプレッサー系(Burkeら(1994)Mol.Microbiol.13,609−618)である。ArgRリプレッサーは、正常にはアルギニンが存在する場合においてのみそのオペレーターに結合する。しかし、変異ArgRNVリプレッサーは、アルギニンが存在しない場合においてそのオペレーターに結合し、アルギニンが存在する場合においても結合したままであり、トランスドミナント(transdominant)効果を有する。二つの対称なArgボックスを有する理想化されたArgR結合部位(オペレーター)は、目的のプラスミド内へ操作され得、ArgR結合部位によって発現が制御される必須遺伝子からの、ArgRNVの結合除去を可能とする。
【0051】
本発明に従えば、E.coliのtrpリプレッサーもまた有用である(「The tryptophan Operon」、Yanof skyおよびCrawford、Escherichia coli and Salmonella typhimurium、J.L.Ingrahamら編、1987年、Amer.Soc.Micro.、1453〜1472頁を参照)。trpリプレッサーは、約50コピー/細胞で存在し、発酵培地中にリプレッサー結合の誘導因子としてトリプトファンの存在を必要とする。
【0052】
本発明に従えば、E.coliのgalRリプレッサーは有用である(「The Galactose Operon」、S.Adhya、Escherichia coli and Salmonella typhimurium、J.L.Ingrahamら編、1987年、Amer.Soc.Micro.、1503〜1512頁を参照)。
【0053】
本発明に従えば、E.coliのaraCリプレッサーもまた有用である(「The L−Arabinose Operon」、R.Schlief、Escherichia coli and Salmonella typhimurium、J.L.Ingrahamら編、1987年、Amer.Soc.Micro.、1473〜1481頁;Dunnら、1984年、Proc.Nat.Aca.Sci.81:5017−5020を参照)。araCリプレッサーは、アラビノースの存在する場合において結合親和性が増大する。
【0054】
最後に、本発明に従えば、Aリプレッサーは有用である(Introduction to Lambda Phages、Current Protocols in Molecular Biology、Ausubelら編、1994年、第3節、ユニット1.9;Hochschildら、1986年、Cell 47(5):807−816)。
【0055】
細胞増殖のために必須の第二の染色体遺伝子は、細胞の代謝産物の生合成に関係する産物をコードする遺伝子、その遺伝子の産物が炭素代謝に関与する遺伝子、または、高分子の生合成もしくは調節をコードする遺伝子(例えば、DNAおよび/もしくはRNAの合成機能および複製機能のために必須の遺伝子)であってもよい。
【0056】
細胞の構成要素の供給、特に細胞壁の前駆体の供給と関与する酵素をコードする特定の遺伝子は、細胞増殖のために必須であり、本発明に従って、有用である。例えば、細菌の細胞壁は、メソジアミノピメリン酸(DAP)を含み、この構成要素を合成することが出来ないことは、溶菌をもたらす。DAP生合成経路内の遺伝子、すなわち、dapA遺伝子、dapB遺伝子、dapC遺伝子、dapD遺伝子、およびdapE遺伝子は、それゆえ、第一の染色体遺伝子によって阻害される、細胞増殖のために必須の第二の染色体遺伝子として用いられ得る。dapAおよびdapBは、クローニングされて塩基配列が決定されており、dapBはひとつのシストロンとして利用可能である(Richaudら、J.Bacteriol.166:297−300,1986年;Bouvierら、J.Biol.Chem.259:14829−14834,1984年)。
【0057】
他の細胞壁構成要素の生合成(例えば、D−アラニン生合成)に関与する遺伝子もまた本発明に従って有用である(Walsh、1989年、J.Biol.Chem.264(5):2393−2396)。
【0058】
脂肪酸生合成に関与する遺伝子もまた、本発明に従って有用である。一つの例は、fabAであり、fabAは、3−ヒドロキシデカノイル−ACPデヒドラーゼをコードしており、不飽和脂肪酸生合成と飽和脂肪酸生合成とが分岐する点での、伸長する脂肪酸鎖への二重結合の導入の原因である(Cronan Jr.J.E.およびRock.C.O.(1987)Biosynthesis of membrane lipids、「Escherichia coli and Salmonella typhimurium:cellular and molecular biology」。Neidhardt.F.C.編、American Society of Microbiology,Washington D.C.)。この酵素は比較的高い濃度で存在し、fabA変異体は、不飽和脂肪酸が補われないと溶菌する。
【0059】
あるいは、細胞増殖に必須の第二の染色体遺伝子は、炭素源の利用と関与する遺伝子であってもよい。具体的には、この第二の染色体遺伝子は、ラクトースオペロンの遺伝子を含んでもよく、その結果、ラクトースが唯一の炭素源である条件において、プラスミドを維持しておりlacオペロンの遺伝子が発現される細胞のみが、生存し得る。他の改変は、当業者にとって明らかである。
【0060】
グルタミンシンテターゼは、NSOミエローマ細胞株(Bebbingtonら、(1992)Bio/Technology 10,169−175)のような真核細胞にとっての必須遺伝子であって、好ましくは、宿主細胞が真核細胞である場合に第二の染色体遺伝子として用いられる。
【0061】
この第二の染色体遺伝子はまた、宿主細胞のDNAおよび/またはRNAの合成タンパク質または複製タンパク質をコードする必須遺伝子であってもよい。E.coliおよびSalmonellaのような細菌におけるこれらの必須機能に関するそのような遺伝子の例は、McMackenら(Escherichia coli and Salmonella typhimurium,Cellular and Molecular Biology、Neidhardtら編、Amer.Soc.Micro,,Wash.D.C.,1987年、564〜612頁)において提供され、以下の遺伝子を含むが、以下の遺伝子に限定されない:dnaA、dnaB、dnaC、ssb、dnaG、polC(dnaE)、dnaQ(mutD)、dnaN、dnaZX、gyrA、gyrB、polA、lig、dnaT、rpoA、rpoB、rpoC、およびrpoD。
【0062】
好ましくないが、この第二の染色体遺伝子は、抗生物質耐性遺伝子であってもよく、その結果、プラスミドを用いるその株の形質転換が、その抗生物質の存在する場合における増殖を可能にする。
【0063】
医療への適用に使用するための形質転換された宿主細胞に対して、投与経路に依存して、局所環境内の多くの要因が選択圧をかけ得る。そのような圧力に抵抗する能力が同定される遺伝子に連鎖しているならば、この遺伝子は、選択マーカーとして用いられ得る。例えば、細胞、特に経口で投与されることになる細菌細胞にとって、胃の内容物の低いpHへの抵抗性は、重要な選択的利点を構成する。このことと関連することが示される遺伝子のうちには、Streptococcus mutansのuvrA遺伝子(Hannaら、2001年、J.Bacteriol 183:5964)、E.coli.のgadA遺伝子、gadB遺伝子、gadC遺伝子(Castanie−CornetおよびFoster、2001年 Microbiology 147:709)、Salmonella typhimuriumのrpoS遺伝子、fur遺伝子、atp遺伝子、およびatbR遺伝子(Baikら、1996年 Microbiology 142:3195)、Helicobacter pyloriのlepA遺伝子、Frasel遺伝子、czcA遺伝子、uvrA遺伝子、atpF’遺伝子、およびアルド−ケトレダクターゼ遺伝子(Bijlsmaら、2000年、J Infect Dis 182:1566)、ならびに、Listeria monocytogenesのF0F1ATPアーゼをコードする遺伝子(Cotterら、2000年、Int J Food Microbial 60:137)がある。
【0064】
細胞内での生存を与える遺伝子(例えば、Listeria monocytogenesのSvpA(Borezeeら、2001年、Microbiology 147:2913)もしくは、リソソームのような特定の細胞内局在に対する抵抗力もまた、適切な宿主細胞において有用であり得る。
【0065】
細胞増殖を間接的に阻害する好ましい遺伝子としては、細胞増殖のために必須な第二の染色体遺伝子に結合することによって、または細胞増殖のために必須な第二の染色体遺伝子から転写されるmRNAに結合することによって、この第二の染色体遺伝子を阻害するアンチセンス配列をコードする、染色体遺伝子も挙げられる。
【0066】
アンチセンス配列とは、このアンチセンス配列の標的配列に実質的に相補的である配列であって、その標的配列に特異的にハイブリダイゼーションする能力を有する。このアンチセンス配列は、細胞増殖のために必須な第二の染色体遺伝子に直接的に結合するか、またはその第二の染色体遺伝子から転写されるmRNAに結合することのいずれかによって作用し得るので、この標的配列は、この第二の染色体のコード鎖もしくは非コード鎖のいずれかの配列、またはそれらの配列の一部であってもよい。標的配列に特異的にハイブリダイゼーションする能力を有するアンチセンス配列を調製する方法は、当業者によって公知である(例えば、Cohen,J.S.,Trends in Pharm.Sci.,10,435(1989)、Okano,J.Neurochem.56,560(1991);O’Connor,J.Neurochem 56,560(1991);Leeら、Nucleic Acids Res 6,3073(1979);Cooneyら、Science 241,456(1988);Dervanら、Science 251,1360(1991)を参照)。
【0067】
好ましくは、このアンチセンス配列は、ストリンジェントな条件の下で、その標的配列にハイブリダイゼーションする。高ストリンジェンシーハイブリダイゼーション条件は、以下のように定義される:50%のホルムアミド、5×SSC(150mM NaCl、15mMのクエン酸三ナトリウム)、50mMのリン酸ナトリウム(pH7.6)、5×デンハルト液、10%のデキストラン硫酸、および20マイクログラム/mlの変性剪断サケ精子DNAを含む溶液中での、42℃での一晩のインキュベーションと、その後の0.1×SSC中でのおよそ65℃でのフィルターの洗浄。
【0068】
上記に示されるように、第一の染色体遺伝子は、細胞増殖のために必須な遺伝子の阻害もしくは細胞増殖を阻害する遺伝子の活性化を最終的にもたらす、多くの遺伝子を含む反応のカスケードを開始することによって、細胞増殖を阻害し得る。例えば、この第一の染色体遺伝子は、第二の染色体遺伝子を阻害するリプレッサータンパク質をコードし得、この第二の遺伝子は、細胞増殖のために必須な第三の染色体遺伝子を他の状態では活性化するアクチベーターをコードする。あるいは、この第一の染色体遺伝子は、第二の染色体遺伝子を阻害するリプレッサータンパク質をコードし得、この第二の染色体遺伝子は、細胞増殖を阻害する第三の染色体遺伝子を他の状態では阻害するアンチセンス配列をコードする。これらが本発明の必要条件を満たす遺伝子の多くのカスケードの例に過ぎないことは、当業者にとって明らかである。
【0069】
(改変された宿主細胞の産生)
上記の細胞増殖を阻害する染色体遺伝子は、宿主細胞のゲノム内に天然に存在する遺伝子であってもよく、あるいは外来遺伝子であってもよい。
【0070】
プラスミドによってコードされるアンチセンス配列が、上記の染色体遺伝子に作動的に連結される調節配列に結合することによってこの染色体遺伝子を阻害する場合、この染色体遺伝子は、外因性の調節配列に作動的に連結されている、天然に存在する遺伝子であってもよい。例えば、天然に存在するトキシン遺伝子の発現を通常では制御する配列が除去されてもよく、このトキシン遺伝子が代わりに、構成的プロモーターの制御下で、外因性のRNAIコード配列に作動的に結合されてもよい。同様に、天然に存在するリプレッサータンパク質の発現を通常では制御する配列が除去されてもよく、このリプレッサータンパク質遺伝子が代わりに、構成的プロモーターもしくは誘導性プロモーターの制御下で、外因性のRNAIIコード配列に作動的に連結されてもよい。従って、本明細書中の実施例2において、天然に存在するリプレッサータンパク質であるlacIは、RNAIIコード領域に作動的に結合される。
【0071】
あるいは、この調節配列がRNAIコード配列もしくはRNAIIコード配列であって、この染色体遺伝子が細胞増殖のために必要である第二の染色体遺伝子を阻害するアンチセンス配列をコードする場合のように、この調節配列およびこの染色体遺伝子の両方が外因性であってもよい。
【0072】
細胞増殖のために必須な第二の染色体遺伝子が存在するならば、この第二の染色体遺伝子もまた、天然に存在する遺伝子または外来遺伝子のいずれであってもよい。
【0073】
細胞増殖を阻害する染色体遺伝子は、特に宿主細胞が真核細胞の宿主細胞である場合において、この細胞のゲノム内に複数コピーで存在し得る。
【0074】
本発明において用いるために適切な宿主細胞を産生するために、細胞を操作する方法は、当業者にとって明らかである。
【0075】
(宿主細胞)
本発明は、動物細胞(例えば、哺乳類細胞および昆虫細胞)、植物細胞、真菌(例えば、酵母)、細菌、および古細菌を含む、全ての細胞型に対して適用され得る。
【0076】
宿主細胞が細菌細胞である場合、この宿主細胞は、グラム陰性細菌細胞であってもよく、グラム陽性細菌細胞であってもよい。この細胞がグラム陰性細菌細胞である場合、この細胞は、好ましくはE.coli細胞もしくはSalmonella細胞である。この細胞がグラム陽性細菌細胞である場合、この細胞は、好ましくはBacillus、Streptomyces、Lactobacillus、もしくはLactococcus細胞である。
【0077】
特に有用であるのは、レシピエント生物体における治療用途が自由な、受容可能な程度にまで副作用が無いように弱毒化されている、弱毒化された宿主細胞である。この細胞の特性に依存して、この細胞は、獣医学的用途もしくはヒトの治療用途のために適切であり得る。
【0078】
入手し得る弱毒化された細菌の中では、以下のものがある:
SL3261株(aroA変異体、Titballら、1997年、Infection and Immunity 65:1926、Titballら、1995年、Infection and Immunity 63:563)、VNP20009株
(purlおよびmsbBの変異体、Tosoら、2002年、J.Clin Oncol 20:142)を含む、Salmonella typhimurium;
CVD 908−htrA株、χ4073株、χ4632株、Ty800株、CVD 909株、およびCVD 915株を含む、Salmonella typhi(Galenら、1999年、Infect.Immun.67:6424−6433;Moronaら、1991年、Gene 107:139−144;Tacketら、1997年、Infect.Immun.65:3381−3385;Garmoryら、2002年、FEMS Microbiology Reviews 26:339−353);
9R株を含む、Salmonella gallinarum(鳥類のワクチン接種のため、Feberweeら、2001年、Avian Dis 45:1024);
aroA変異体、cnfl変異体、およびOMP変異体を含む、Escherichia coli(Rippere−Lampeら、2001年、Infect Immunol 69:3954);
S.flexneri 2a guaBA株を含む、Shigella flexneri(Altboumら、2001年、Infect Immunol 69:3150);
Peru2株を含む、Vibrio cholerae(Ryanら、2000年、Infect.Immun.68:221−226);
PrfA変異体およびSvpA変異体を含む、Listeria monocytogenes(Sheehanら、1996年、Mol Microbiol 20:785;Borezeeら、2001年、Microbiology 747:2913);
RB51株を含む、Brucella abortus(Vemulapalliら、2000年、Infect Immunol 68:3290);
Calmette−Guerin(BCG)株を含む、Mycobacterium bovis;および
Mycobacterium tuberculosis。
【0079】
(本発明に従って有用である、プラスミドによって運ばれる遺伝子)
本発明の形質転換された宿主細胞内のプラスミドで運ばれる目的の遺伝子は、任意の遺伝子であり得る。この遺伝子は、レシピエント生物体に送達された場合に治療効果を有する遺伝子であり得る。例えば、遺伝子治療目的のためにレシピエント生物体にこの遺伝子を送達することが所望され得る。
【0080】
あるいは、遺伝子の転写産物もしくは転写および翻訳の産物が治療効果を有するので、この遺伝子をレシピエント生物体に送達することが所望され得る。例えば、目的の遺伝子の転写産物は、アンチセンスオリゴヌクレオチドであってもよく、あるいは、この目的の遺伝子は、治療効果を有する抗原をコードしてもよい。特に、目的の遺伝子は、レシピエント生物体において免疫応答を惹起する一つ以上の抗原をコードしてもよい。レシピエント生物体において免疫応答を誘導するためにプラスミドに含まれ得る目的の遺伝子の例は、cafオペロンのcafI遺伝子、caf1A遺伝子、caf1M遺伝子がある。
【0081】
目的の遺伝子が転写されるもしくは転写されて翻訳されることが望ましい場合、この目的の遺伝子は、好ましくはプロモーターと機能的に結合する。好ましくは、このプロモーターは真核生物のプロモーターもしくは細菌のプロモーターである。目的の遺伝子と機能的に結合するプロモーターの選択、および宿主細胞の選択は、宿主細胞において、レシピエント生物体への投与に先立って、もしくはレシピエント生物体への投与の後でのみ、目的の遺伝子が転写されるか、または転写されて翻訳されるかを決定する。
【0082】
いくつかの場合において、目的の遺伝子を既に発現している宿主細胞に送達することが、所望され得る。この場合において、この目的の遺伝子と機能的に結合しているプロモーターは、その結果として、このプロモーターがこの宿主細胞内で作動し、この宿主細胞内におけるこの目的の遺伝子の転写および翻訳を促進するように、選択される。例えば、この宿主細胞が細菌細胞である場合、この状況においてこの目的の遺伝子と機能的に結合しているプロモーターは、細菌のプロモーターであってもよい。
【0083】
他の場合において、この目的の遺伝子を含むプラスミドを含む宿主細胞をレシピエント生物体に送達するが、この目的の遺伝子の発現をこの細胞のレシピエント生物体への送達後にまで遅延させることが、所望され得る。例えば、この宿主細胞が細菌細胞である場合、この状況においてこの目的遺伝子と機能的に結合しているプロモーターは、この宿主細胞においてこの目的の遺伝子の転写および翻訳を促進しないが、レシピエント生物体への投与の後にこの目的の遺伝子の転写および翻訳を促進する、真核生物のプロモーターであってもよい。
【0084】
(本発明に従って有用であるワクチン接種の方法)
本発明によって形質転換された宿主細胞を用いるワクチン接種は、多くの投与経路のいずれによってであってもよいことが、予想される。Mycobacteriumについては、例えば、有用な経路は、皮内注射もしくは皮下注射によってである。Salmonella、Shigella、もしくはE.coliのような腸内細菌については、経口投与(特に、耐酸性形質転換体の経口投与)または直腸投与が好ましい。いくつかの状況においては、粘膜投与、静脈内投与、もしくは腹腔内投与が、好適であり得る。
【0085】
(他のインビボでの適用のための投与)
非ワクチン接種適用のための多くの投与方法が用いられ得ることが、予想される。
【0086】
ワクチン接種のために用いられる経腸投与経路および非経口投与経路に加えて、非ワクチン適用は、他の手順を必要とし得る。他の手順の中では、本発明による細胞が中に含まれる移植可能な浸透性の容器の使用がある。
【0087】
本発明は、ここで、一例としてプラスミドの複製起点を選択マーカーとして使用するプラスミド維持系を参照して、さらに詳細に記載される。実施例において記載される系に対して変更がなされ得ることが、理解される。
【実施例】
【0088】
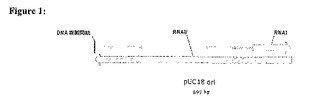
本発明は、複製起点(pMB1 oriもしくはColEI ori)のみを選択マーカーとして用いてプラスミドの選択およびプラスミドの維持を可能とし、本明細書中でoriSELECTと言及される。図1において示すように、これらのColEI適合性oriは、二つの重なるRNA転写産物をコードする:RNAIIは、プラスミドDNA合成の開始を始め、RNAIは、RNAIIのアンチセンスインヒビターである(Scottら、1984,Microbiol.Rev.48:1−23;Chanら、1985,J.Biol.Chem.260:8925−8935)。研究用途もしくは商業的用途の事実上全てのプラスミドは、E.coli内での自律複製をこれらの複製起点に依存している。
【0089】
本発明は、あるオペロンにおいて阻害的遺伝子エレメントに連結されたRNAIIコード領域を含むようにゲノムが改変されているE.coliの形質転換株の構築を、含み得る。プラスミド産生性RNAIのRNAII−mRNA融合体へ結合は、その阻害遺伝子mRNAの転写を妨害し、それによって細胞の生存および増殖を可能にする。このプラスミド維持系の3つの提案される機構は、以下に記載される。
【0090】
(実施例1:リプレッサーoriSELECT)
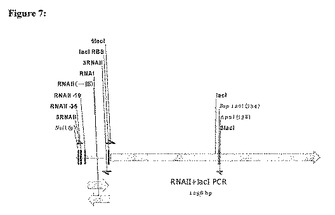
図2において示すように、リプレッサーoriSELECTにおける使用のための宿主細胞のゲノムは、リプレッサー遺伝子シストロンに連結されたRNAIIコード領域を含み、RNAIIコード領域およびリプレッサー遺伝子シストロンの両方は、構成的RNAIIプロモーターの制御下にある。リプレッサーシストロンに連結されたRNAIIコード領域はまた、図8において示されるような、E.coliのTrcプロモーターにおいて見出される−10配列および−35配列ならびにスペーシングに基づくプロモーターのような、代替的な構成的プロモーターの制御下にあり得る。宿主細胞のゲノムはまた、誘導性プロモーター(inducible promoter)の制御下での細胞増殖のために必須な遺伝子を含む(図8においては示さない)。
【0091】
プラスミドが無い場合において、リプレッサータンパク質が発現し、そして必須遺伝子(essential gene)の発現を制御するオペレーター/プロモーターに結合することによって必須遺伝子を抑制し、細胞増殖を阻害する。プラスミド産生性RNAIの、RNAII−リプレッサーmRNA融合体内のRNAII転写産物への結合は、リプレッサーの翻訳を妨げ、必須遺伝子の発現を可能にし、それによって細胞増殖を可能にする。
【0092】
リプレッサーoriSELECTにおける使用のためにこの宿主細胞を構築するために、dapD(Richaudら、1984,J.Biol.Chem.259:14824−14828)もしくはfabA(Cronanら、1987,Biosynthesis of membrane lipids、「Escherichia coli and Salmonella typhimurium:cellular and molecular biology」,Neidhardt,F.C.編、American Society of Microbiology,Washington D.C.)のような必須遺伝子を、細菌株の染色体から欠失させ、誘導性プロモーター(例えば、Plac)の制御下に置き、そしてその染色体内へ挿入し戻す。その誘導性プロモーターに結合している、リプレッサーをコードしている遺伝子(この場合においてはlacI)を、染色体から欠失させる。このリプレッサー遺伝子を、両方の転写を駆動する構成的RNAIIプロモーター(もしくは任意の他の構成的プロモーター、例えばE.coliのtrcプロモーターにおいて見出される−10配列および−35配列ならびにスペーシングに基づくプロモーター)と共に、RNAIIの一部をコードしている領域の下流のオペロン内に置く。RNAIIコード領域の3’末端を、リプレッサーのリボソーム結合部位を含むように改変し、このためRNAIIコード領域の3’末端は重なり、RNAIによって妨げられる。いくつかの場合においては、RNAIIコード領域の3’末端とlacのリボソーム結合部位との間の距離を増大させることによって、LacIリプレッサータンパク質の発現を増加させ得る。この構築物を、染色体の中へ挿入する。
【0093】
その細胞はそれからリプレッサーを構成的に産生する。リプレッサータンパク質は、必須遺伝子を制御している染色体オペレーターに結合し、それによって細胞増殖を妨害する。その細胞をColEI適合性oriを含むプラスミドを用いて形質転換するとき、RNAIは、RNAIIに結合し、そして下流のリプレッサー遺伝子の翻訳を妨げ、細胞増殖を可能にする。
【0094】
染色体の必須遺伝子がその必須遺伝子自体の結合したリプレッサーを有するのであれば、このリプレッサー(マイナスそのリプレッサーのプロモーター)をRNAIIの下流のオペロン内へ操作してもよく、そしてこのオペロンを野生型リプレッサーと置き換えさせるために細菌の染色体内へ挿入してもよい。
【0095】
上記の必須遺伝子は、条件的に必須な外来遺伝子(例えば、抗生物質耐性遺伝子)であり得る。
【0096】
必須な外来遺伝子が誘導性であり、かつその遺伝子自体の結合した外来リプレッサーを有するのであれば、このリプレッサー遺伝子をRNAIIの下流のオペロン内へ操作してもよく、そしてこのオペロンを細菌の染色体内へ挿入してもよい。
【0097】
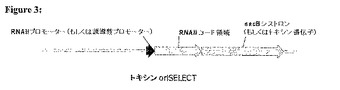
(実施例2:トキシン遺伝子oriSELECT)
図3において示すように、トキシン遺伝子oriSELECTにおける使用のための宿主細胞のゲノムは、RNAIIの下流に配置されたBacillus subtilisからのレバンスクラーゼ(Linkら、1997,J.Bacteriol.179:6228−6237)をコードするsacB遺伝子を含む。ショ糖が存在する場合において、sacBは毒性の産物を合成し、RNAIを発現するプラスミドを導入しなければ、細胞は死ぬ。誘導性プロモーターによって調節される別の毒性遺伝子を、sacBの代わりにRNAIIコード領域へ融合してもよい。
【0098】
構築物を、RNAIIコード領域およびsacBをコードする下流の遺伝子からなるオペロンを用いて操作する。RNAIIコード領域の3’末端を、sacBのリボソーム結合部位を含むように改変する。構成的RNAIIプロモーターは、遺伝子発現を駆動する。構成的RNAIIプロモーターを細菌の染色体内へ挿入する場合、その細胞は、ショ糖がない場合においても増殖し得るが、ショ糖を加えると、トキシンが産生されそして細胞は死ぬ。その細胞を、ColEI適合性oriを含むプラスミドを用いて形質転換する場合、RNAIがRNAII−トキシン遺伝子mRNAに結合し、そして翻訳を妨げ、ショ糖が存在する場合においても細胞増殖を可能にする。
【0099】
温度感受性プロモーターもしくは化学的に誘導されるプロモーターを、RNAII−トキシン遺伝子融合体の発現を駆動するために用いてもよい。許容温度において、もしくは誘導物質が存在する場合においては、RNAIを発現するプラスミドを導入しなければ、細胞は死ぬ。
【0100】
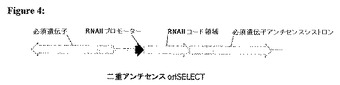
(実施例3:二重アンチセンスoriSELECT)
図4において示すように、二重アンチセンスoriSELECTにおいて、宿主細胞のゲノムは、細胞増殖のために必須である遺伝子、および細胞増殖のために必須である遺伝子に対するアンチセンスである遺伝子に連結されたRNAIIコード領域を、含む。
【0101】
必須遺伝子(例えば、dapDもしくはfabA)のアンチセンスバージョンを、RNAIIコード領域の下流のオペロン内に、必須遺伝子のアンチセンスバージョンおよびRNAIIコード領域の両方の転写を駆動する構成的RNAIIプロモーターと共に置く。RNAIコード領域の3’末端を、必須遺伝子アンチセンス転写産物の一部を含むように、それによってRNAIコード領域の3’末端がRNAIIと部分的に重なるように改変してもよい。この構築物を、細菌の染色体内へ挿入する。
【0102】
その細胞はそれからRNAIIおよびアンチセンスRNAを構成的に産生する。アンチセンスRNAは、必須遺伝子mRNAに結合し、それによって必須遺伝子の翻訳および細胞増殖を妨害する。
【0103】
その細胞をColEI適合性oriを含むプラスミドを用いて形質転換する場合、RNAIはRNAII−アンチセンスRNAに結合し、RNAII−アンチセンスRNAが必須遺伝子mRNAに結合することを妨害し、細胞増殖を可能にする。
【0104】
(実施例4:リプレッサーoriSELECT株の製作)
ORT株DH1lackan(Williansら、1998)、DH1lacdapDおよびDH1lacP2dapD(Cranenburghら、2001,Nucleic Acid Res.29:e26)を、RNAII−lacIオペロンの染色体のlacIリプレッサー遺伝子への組込みによって(そしてそれによってRNAII−lacIオペロンで染色体lacIリプレッサー遺伝子を置き換えることによって)oriSELECT株に転換してもよい。pKO3組込み系(Linkら、1997,J.Bacteriol.179:6228−6237)に基づいて構築された単一プラスミドは、以下のように、全てのORT株において、野生型lacIをRNAII−lacIで置換するために用いられる。
【0105】
1.lacIを取り囲む染色体lacオペロンの一部を、PCRによって増幅し、クローニングする;
2.lacIリプレッサー遺伝子の上流に挿入されたRNAIIコード領域からなるオペロンを操作し、天然のプロモーターと置き換える。lacIのリボソーム結合部位は、mRNA上でアンチセンスRNAIIの結合によって妨げられるように、配置されるべきである。
【0106】
3.RNAII−lacIオペロンの遺伝子座を、組込みプラスミドpKO3内へクローニングする。
【0107】
4.RNAII−lacIオペロンの遺伝子座を、DH1lackan、DH1lacdapD、およびDH1lacP2dapDの野生型lacオペロンの遺伝子座内へ組込んで、oriSELECT株を産生する。
【0108】
5.これらの株におけるプラスミドの選択およびプラスミドの維持を、ColE1/pMB1 ori配列を保有しかつlacO配列を保有しない(ORT選択を避けるため)プラスミドを用いて試験する。
【0109】
このoriSELECT株の構築のために必要であるクローニングの工程を、より詳細に以下に記載する。
【0110】
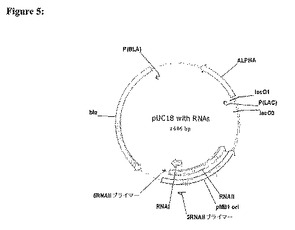
(RNAII−lacI融合体を産生するスプライシングPCR)
1.図5において示すように、pUC18から、RNAIIの5’末端の一部を、以下のプライマー(産物=176bp)を用いるPCRによって増幅する。
【0111】
5RNAII:GAATGCATCAAAGGATCTTCTTGAGA(26nt)
3RNAII:ACATTCACCACCGTATTTGGTATCTGCGCTCTGCTGAAGCCA(42nt)。
【0112】
2.図6に示すように、DH1 gDNAから、lacIの5’末端の一部を、以下のプライマーを用いるPCRによって増幅する(産物=597bp)。
【0113】
5lacI:GATACCAAATACGGTGGTGAATGTGAAACCAGTAACGTTATA(42nt)
3lacI:ACAGAACTTAATGGGCCCGCTAACA(25nt)。
【0114】
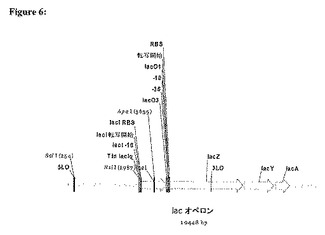
3.図7において示すように、5RNAIIプライマーおよび3lacIプライマーを用いて、一回のPCRにおいてこれら二つのPCR産物を組み合わせ、増幅して、749bpのRNAII−lacI遺伝子融合体PCR産物(以下の5RNAIIプライマーと3lacIプライマーとの間の領域)を生成する。
【0115】
4.スプライシングされたPCR産物を、NsiIおよびApaIを用いて切断して、lacIプロモーターおよび5’末端の位置においてラクトースオペロン内へクローニングするための断片を生成する。
【0116】
(ラクトースオペロンの一部のクローニング、およびRNAII−lacIの挿入)
1.DH1 gDNAからのlacオペロンを、以下のプライマーを用いて増幅する。
【0117】
【化1】
プライマー5LOは、天然のSalI部位(下線)を含むが、3LOは、1ヌクレオチド変化(太字)を有するSalI部位を導入する。総PCR産物の大きさは、5803bpである。
【0118】
2.上記のPCR産物をSalIで切断し、そしてSalIを用いて切断されたpUC18内へクローニングする。
【0119】
3.このプラスミドをNsiIおよびApaIで切断し、そして工程4による断片にクローニングし、プロモーターおよびlacIの5’末端をRNAII−lacI遺伝子融合体で置換する。
【0120】
4.挿入部分をSalIを用いて切断し、そしてSalIを用いて切断されたpKO3recA内へライゲーションする。
【0121】
5.oriSELECT株を産生するために、DH1lacdapD、DH1lacP2dapD、およびDH1lackanの染色体内へ組込む。
【0122】
6.pMB1複製起点を有するプラスミドを用いて、プラスミドの選択およびプラスミドの維持を試験する。
【0123】
RNAII構成的プロモーターをRNAII−遺伝子融合体発現を駆動するために用いる場合、必要であれば、任意の他の構成的プロモーターを置換してもよい。そのRNAIIプロモーターをこれらの実施例において既に存在するものとして用いる。代替的な好ましい構成的プロモーターは、至適なE.coliのtrcプロモーターにおいて見出される−10配列および−35配列ならびにスペーシングに基づくプロモーターである。このE.coliプロモーターは、RNAIIプロモーターより強く、それゆえに、より大量のLacIリプレッサータンパク質を産生し、それによってプラスミドが存在しない場合において増殖が妨げられることを保証することが望まれる場合に用いてもよい。LacIリプレッサータンパク質の発現はまた、RNAIIコード領域の3’末端とlaciIのリボソーム結合部位との間の距離を長くすることによって増大されてもよい。
【0124】
また、遺伝子カセットを挿入する代替的な方法を、上記に記載したpKO3法の代わりに用いてもよいことが理解される。例えば、ラムダRed組換え系を用いて、直鎖状PCR産物としてこのカセットを組込んでもよい(Murphy,1998,J.Bacteriol.,180:2063−2071)。
【0125】
これらの実施例は、アンチセンスインヒビターとして機能するRNAIを用いるRNAII−遺伝子融合体発現カセットに基づく。このことは、なぜなら、プラスミドを含む細胞においては、RNAIはRNAIIを五倍の過剰量で上回って存在するからであり(Liangら、1999,J.Mol.Biol.292:19−37)、ゆえに、モル過剰のRNAIが利用可能であるかのように、RNAIがより効果的なインヒビターとして働き得るからである。しかし、RNAIIよりもRNAIを用いて遺伝子融合物を構築することが有利である状況はあり得る。この場合において、アンチセンスインヒビターとして働くのは、プラスミドからのRNAII転写産物である。
【0126】
ColEI/pMB1 oriからのRomタンパク質を発現する遺伝子の染色体組込みは、この染色体組み込みがRNAIのRNAIIとの結合親和性を上昇させるので、oriSELECT株において必要であり得る(Chanら、1984,J.Biol.Chem.260:8925−8935)。
【図面の簡単な説明】
【0127】
【図1】pUC18oriの図であり、RNAI転写産物およびRNAII転写産物の位置を示す。RNAIIは、プラスミドのDNA合成の開始を始め、RNAIは、RNAIIのアンチセンスインヒビターである。
【図2】リプレッサーoriSELECTにおいて用いるための宿主細胞のゲノムの例。宿主細胞のゲノムは、リプレッサー遺伝子のシストロンに連結されたRNAIIコード領域を含み、これらの両方が構成的RNAIIプロモーターの制御下にある。このゲノムはさらに、誘導性プロモーターの制御下にある必須遺伝子を含む。プラスミドが存在しない場合において、リプレッサータンパク質が発現され、必須遺伝子を抑制し、細胞増殖を阻害する。プラスミドによって生成されるRNAIの、RNAII−リプレッサーmRNA融合体のRNAII転写産物への結合は、リプレッサータンパク質の翻訳を遮断し、必須遺伝子の発現および細胞増殖を可能にする。
【図3】トキシン遺伝子oriSELECTにおいて用いるための宿主細胞のゲノムの例。このゲノムは、Bacillus subtilis由来のレバンスクラーゼをコードし(Linkら、1997年、J.Bacteriol.179:6228−6237)RNAIIの下流に配置されたsacB遺伝子を含む。ショ糖が存在する場合において、sacBは毒性の産物を合成し、トキシン遺伝子のmRNAの翻訳を遮断するRNAIを発現するプラスミドが存在しない限り、ショ糖が存在する場合においてその細胞は殺される。
【図4】アンチセンスoriSELECTにおいて用いるための宿主細胞のゲノムの例。この宿主細胞のゲノムは、細胞増殖のために必須である遺伝子、および、細胞増殖のために必須な遺伝子に対するアンチセンスである遺伝子に連結されたRNAIIコード領域を含む。プラスミドが存在しない場合において、このアンチセンスRNAは必須遺伝子のmRNAに結合し、それによってこの必須遺伝子の翻訳および細胞増殖を妨げる。この細胞がRNAIを与えるプラスミドを用いて形質転換される場合、RNAIはRNAII−アンチセンスRNAに結合し、RNAII−アンチセンスRNAが必須遺伝子のmRNAに結合することを妨げ、細胞増殖を可能にする。
【図5】PCRによるpUC18からのRNAIIの5’末端の増幅。RNAI転写産物およびRNAII転写産物の位置は、リプレッサーoriSELECTにおいて用いるための宿主細胞のゲノムへの挿入のための、リプレッサーlacIをコードする遺伝子の上流へのRNAIIの挿入に先立って、RANIIの5’末端を増幅するために用いられるプライマーの位置であるとして、示される。
【図6】PCRを用いる、DH1 ORT株のゲノムDNA由来のlacIの5’末端の増幅。この増幅において用いられるプライマー5lacIおよび3lacIの位置が示される。
【図7】図5中のプライマーを用いて産生されるRNAII増幅産物と、図6中のプライマーを用いて産生されるlacI増幅物とを組み合わせることによって形成される産物。この組合せ産物の増幅において用いられるプライマーの位置が示される。
【図8】リプレッサーoriSELECTにおいて用いるための宿主細胞のゲノムのさらなる例。この宿主細胞のゲノムは、リプレッサー遺伝子のシストロン(lacI)に連結されたRNAIIコード領域を含み、これらの両方が、E.coliのtrcプロモーターにおいて見出される−10配列および−35配列ならびにスペーシングに基づく構成的プロモーターの制御下にある。このゲノムはさらに、誘導性プロモーターの制御下にある必須遺伝子を含む(示さず)。プラスミドが存在しない場合において、このリプレッサータンパク質が発現され、必須遺伝子を抑制し、細胞増殖を阻害する。プラスミドによって生成されるRNAIの、RNAII−リプレッサーmRNA融合体のRNAII転写産物への結合は、リプレッサータンパク質の翻訳を遮断し、必須遺伝子の発現および細胞増殖を可能にする。
【技術分野】
【0001】
(発明の分野)
本発明は、プラスミドの安定した維持のための系、この系においての使用のための宿主細胞、ならびに医療への適用において有用なプラスミドを得るためにこの系を用いる方法に関する。
【0002】
本明細書において引用される全ての文献は、参考として援用される。
【背景技術】
【0003】
(発明の背景)
プラスミドは、組換えタンパク質の調製においておよび遺伝子治療目的のためのDNAの調製において、慣用的に用いられる。宿主細胞内のプラスミドの安定した維持は、これらの産物の効率的な調製のために重要である。しかし、宿主細胞において運ばれる染色体外DNAは、プラスミドを含む細胞での、プラスミドが無い細胞に比べて増大した代謝的負荷に起因して、もともと不安定である。プラスミドの安定性を維持し代謝的負荷を軽減するため、プラスミドは、優性な選択マーカーを含むように操作されている。
【0004】
培養中の細胞内にプラスミドを維持する従来の方法は、抗生物質耐性遺伝子をそのプラスミドに含め、そして適切な抗生物質の存在中でその細胞を培養することである。治療用途が意図される細胞もしくはプラスミドに関して、このことは、抗生物質耐性遺伝子を含むプラスミドの使用が抗生物質耐性の拡がりの一因になり得るという不利益を有する。
【0005】
いくつかのプラスミド維持法は、プラスミドによって運ばれる遺伝子(plasmid−borne gene)によって制御される天然に存在する分離後殺傷(post segregational killing)機構を利用することを試みてきた。例えば、hok/sok系、srnB系、およびpnd系は、キラー(killer)タンパク質を含み、そのキラータンパク質は、安定なmRNAによってコードされ、キラーRNAに結合してキラーRNAを不活性化する小さい不安定なアンチセンスRNAによって調節される。キラーRNAは、アンチセンスRNAが分解された後、プラスミドが無い分離体(segregant)において保持され、致死タンパク質へと翻訳される。hok/sok系を用いるプラスミド維持は、弱毒化生ベクターワクチン株Salmonella typhi CDV908 htr−A(非特許文献1)において研究された。しかし、そのような分離後殺傷機構は、形質転換後のプラスミド選択を可能とせず、ゆえに依然としてプラスミドにおける抗生物質耐性遺伝子の存在に依存している。
【0006】
抗生物質選択を用いずにプラスミドを維持するためおよびプラスミドを選択するためのいくつかの代替的な方法が開発されてきており、それらの方法においては、プラスミドが宿主細胞の栄養要求性を補完する遺伝子をコードする。例えば、宿主細胞は、必須アミノ酸代謝産物を合成することが出来ず、このアミノ酸の合成のための失われたエレメントをコードしている遺伝子を含むプラスミドが存在する場合においてのみ、そのアミノ酸を欠乏している培地中で生存し得る、変異細胞であり得る(非特許文献2)。しかし、このアプローチは、そのアミノ酸が省かれなければならないために、増殖培地の組成を制限する。複雑な培地において用いられ得る代替的な方法は、温度感受性のtRNAシンテターゼ遺伝子を有する変異宿主細胞を用いる。この変異宿主細胞は、野生型tRNAシンテターゼ遺伝子を含むプラスミドが存在する場合のみ、非許容(non−permissive)温度で生存し得る(非特許文献3)。別の選択方法は、変異宿主細胞における必須な染色体遺伝子中のナンセンス変異を補完するプラスミドによって運ばれるtRNA遺伝子を用いる(非特許文献4)。あるいは、pilオペロンのような、細胞に対する代謝的負荷を増大する遺伝子が、対応するリプレッサータンパク質をコードするプラスミドの存在する場合においてのみ宿主細胞が生存するように、宿主の染色体に配置され得る(非特許文献5)。
【0007】
特許文献1は、インビトロ培養中の宿主細胞内のプラスミドをオペレーターリプレッサータイトレーション(operator repressor titration)によって維持する方法を説明している。その方法は、その宿主細胞が、リプレッサーをコードする第一の染色体遺伝子、および、その調節領域においてリプレッサーについてのオペレーター配列を有する細胞増殖のために必須である第二の染色体遺伝子を含むようにする、宿主細胞の操作を伴う。プラスミドが存在しない場合、第二の染色体遺伝子の発現は、リプレッサーのオペレーターへの結合によって阻害され、そしてその細胞は死ぬ。この宿主細胞における維持のためのプラスミドは、オペレーター配列を含むように操作されており、その結果、プラスミドが存在する場合、リプレッサーは細胞増殖のために必須である遺伝子のオペレーターから結合除去(titrate away)され、その遺伝子が発現されその細胞が生存する。この機構はまた、Williamsら(非特許文献6)に、およびCranenburghら(非特許文献7)に、記載される。
【特許文献1】欧州特許第0851932号明細書
【非特許文献1】Galenら、Infect.Immunol.,1999年、第67巻、p.6424−6433
【非特許文献2】Wang M−Dら、J.Bacteriol.,1987年、第169巻、p.5610−5614
【非特許文献3】Skogmanら、Gene,1984年、第31巻、p.117−122
【非特許文献4】Zengelら、J.Bacteriol.,1981年、第145巻、p.459−465
【非特許文献5】Ogdenら、Biotech.Bioeng.,1992年、第40巻、p.1027−1038
【非特許文献6】Williamsら、Nucleic Acids Research,1999年、第26巻、第9号、p.2120−2124
【非特許文献7】Cranenburghら、Nucleic Acids Research,2001年、第29巻、第5号、p.e26−e27
【発明の開示】
【発明が解決しようとする課題】
【0008】
抗生物質選択に依存しないプラスミドの維持および選択のいくつかの機構は公知であるが、治療への適用のためのDNAおよび組換えタンパク質の産生におけるプラスミドの重要性が増大していることを考慮して、さらなる方法の開発についての必要性は存続する。加えて、今までに開発された、プラスミドの維持および選択の系は、これらの系における使用のために特別に改変されているプラスミドの使用を必要とする。抗生物質耐性を伴わず、当該分野において一般的であり特別な改変を必要としないプラスミドを使用する、プラスミドの維持および選択の系についての必要性が存続する。
【課題を解決するための手段】
【0009】
(発明の要旨)
本発明の第一の局面に従って、形質転換された宿主細胞が提供される。その宿主細胞は、
i)細胞増殖を阻害する染色体遺伝子;および
ii)アンチセンス配列をコードするプラスミド
を含み、上記プラスミドによってコードされるアンチセンス配列は、上記染色体遺伝子の作用を阻害し、それによって細胞増殖を許容する。
【0010】
上記プラスミドによってコードされるアンチセンス配列は、このプラスミドの他の領域によってコードされてもよいが、好ましくはこのプラスミドの複製起点によってコードされる。
【0011】
本明細書において用いられる場合、用語「アンチセンス配列」とは、核酸配列であって、その核酸配列の標的配列に対して実質的に相補的であり、その標的配列に特異的にハイブリダイズする能力を有する核酸配列を、指す。好ましくは、上記のプラスミドによってコードされるアンチセンス配列は、ストリンジェントな条件下でそのアンチセンス配列の標的配列にハイブリダイズする。高ストリンジェンシーハイブリダイゼーション条件は、以下のように定義される:50%のホルムアミド、5×SSC(150mMのNaCl、15mMのクエン酸三ナトリウム)、50mMのリン酸ナトリウム(pH7.6)、5×デンハルト液、10%のデキストラン硫酸、および20マイクログラム/mlの変性剪断されたサケ精子DNAを含む溶液中での、42℃での一晩のインキュベーションと、その後の0.1×SSC中でのおよそ65℃でのフィルターの洗浄。
【0012】
本明細書において用いられる場合、「細胞増殖」とは、培養培地中の細胞数の経時的な増加、および、生細胞の数が経時的に減少しない場合は、細胞の生存率を、指す。「細胞増殖を阻害する」によって、染色体遺伝子が細胞にとって致死性であり、その結果、培養培地中の生細胞の数が経時的に減少すること、または、染色体遺伝子が細胞増殖を妨げ、その結果、培養培地中の細胞の数が時間にわたって増加しないことが、意味される。
【0013】
本発明のこの第一の局面に従う形質転換された宿主細胞の増殖は、プラスミドの存在に依存しており、このプラスミドを維持する細胞の選択をもたらす。宿主細胞からこのプラスミドが失われた場合、この染色体遺伝子の作用はもはやプラスミドによってコードされるアンチセンス配列によって妨げられず、細胞増殖が阻害される。いくつかの実施形態において、この染色体遺伝子の発現が阻害され、それによって細胞増殖を許容するためには、ほんの1コピー(copy)のこのプラスミドが必要であり得る。他の実施形態において、染色体遺伝子の発現が阻害されるために、プラスミドが複数コピーで存在することが必要であり得る。
【0014】
この染色体遺伝子は、細胞増殖を直接的に阻害し得る。例えば、この染色体遺伝子の転写および翻訳は、トキシンのような、細胞にとって致死性のタンパク質を産生し得る。あるいは、この染色体遺伝子は、細胞増殖を間接的に阻害し得る。例えば、この染色体遺伝子の転写および翻訳は、第二の染色体遺伝子によってコードされる細胞増殖のために必須であるタンパク質の転写および翻訳を阻害するリプレッサータンパク質を、産生し得る。この染色体遺伝子の転写はまた、細胞増殖のために必須な第二の染色体遺伝子の転写および/もしくは翻訳を、第二の染色体遺伝子に結合することまたは第二の染色体遺伝子から転写されるmRNAに結合することのいずれかによって阻害するアンチセンス配列を、産生し得る。
【0015】
いくつかの実施形態において、この染色体遺伝子は、より複雑な反応カスケードによって細胞増殖を阻害し得る。例えば、この染色体遺伝子は、第二の染色体遺伝子の発現を活性化するアクチベータータンパク質をコードし得、この第二の染色体遺伝子は、次に、細胞増殖のために必須なタンパク質をコードする第三の染色体遺伝子の発現を阻害するリプレッサータンパク質をコードする。あるいは、第一の染色体遺伝子は、第二の染色体遺伝子のアンチセンスインヒビターをコードし得、この第二の染色体遺伝子のアンチセンスインヒビターは、次に、細胞増殖を阻害するタンパク質をコードする第三の染色体遺伝子の発現を抑制するリプレッサータンパク質をコードする。そのようなカスケードは三つより多くの染色体遺伝子を含み得る。例えば、そのようなカスケードは、4、5、6、7もしくはそれより多くの染色体遺伝子を含み得る。そのようなカスケードの全てにおいて、第一の染色体遺伝子を阻害するプラスミドが存在しないことにより、第一の染色体遺伝子にカスケード反応を開始させ、このことから最終的に細胞増殖の阻害がもたらされる。細胞増殖を阻害する適切な染色体遺伝子の例は、本明細書において提供される。
【0016】
本発明の形質転換された宿主細胞は、好ましくは、治療用途が意図されるプラスミドを維持するために用いられる。従って、このプラスミドが目的の遺伝子の挿入のためのクローニング部位を含むことが、好ましい。好ましくは、このプラスミドはさらに、目的の遺伝子を含む。この目的の遺伝子は哺乳類細胞において、好ましくはヒト細胞において、発現し得ることが好ましい。この目的の遺伝子は、治療用途のために、目的のRNAもしくは目的のタンパク質を発現し得る。プラスミドに含まれ得るそのような目的の遺伝子の例は、当該分野において公知であり、本明細書において提供される。
【0017】
上記プラスミドによってコードされるアンチセンス配列は、上記染色体遺伝子自体に結合することによってこの染色体遺伝子の作用を阻害し得、それによってこの遺伝子の転写を阻害し得る。あるいは、上記プラスミドによってコードされるアンチセンス配列は、上記染色体遺伝子から転写されるmRNAに結合することによってこの染色体遺伝子の作用を阻害し得る。この染色体遺伝子の性質に依存して、この染色体遺伝子から転写されるmRNAへの結合は、この染色体遺伝子の翻訳を阻害し得るか、もしくはこの染色体遺伝子にコードされるアンチセンス配列がこのアンチセンス配列の標的へ結合するのを妨ぎ得る。
【0018】
本発明の第一の局面の好ましい実施形態において、調節配列は、上記染色体遺伝子と作動的(operatively)に連結される。「作動的に連結される」によって、この調節配列が、読み枠をあわせて(in−frame)この染色体遺伝子と連結され、その結果、この調節配列がこの染色体遺伝子と同時に転写されることが、意味される。好ましくは、この調節配列は、この染色体遺伝子の上流に作動的に連結される。しかし、あるいはこの調節配列は、この染色体遺伝子の下流に作動的に連結されてもよい。この調節配列がこの染色体遺伝子の上流に作動的に連結された場合、この調節配列は、好ましくはリボソーム結合部位内に挿入される。あるいは、この調節配列は、リボソーム結合部位の上流に挿入されてもよい。この染色体遺伝子もしくは調節遺伝子に作動的に連結されたこの染色体遺伝子は、構成的プロモーターもしくは誘導性プロモーターの制御下にあってもよい。好ましくは、この染色体遺伝子もしくは調節遺伝子に作動的に連結されたこの染色体遺伝子は、構成的プロモーターの制御下にある。
【0019】
上記プラスミドによってコードされるアンチセンス配列は、この調節配列に結合することによってもしくはこの調節配列から転写されたmRNAに結合することによって、上記染色体遺伝子の作用を阻害し得る。本発明のこの実施形態において、上記プラスミドによってコードされるアンチセンス配列は、好ましくはこのプラスミドの複製起点によってコードされる。
【0020】
全てのプラスミドの複製起点は、転写されたRNAを産生する。いくつかの場合において、プラスミド(例えば、プラスミドR1、プラスミドRK6、プラスミドpT181、プラスミドpMV158、およびプラスミドpIP501)の複製起点から転写されたRNAは、複製の調節に直接的に用いられる。他の場合において、転写されたRNAは、翻訳されてプラスミド(例えば、pSC101、pPS10、p15A、F、R100、R453、P1、RK2、RA1、RSF10110、pColIV−K30、ColE2、COlE3、Rts1、pLS20、およびpUB110)の複製のために必要とされるタンパク質を提供する(Solarら、1998,Microbiol and Molec.Biol.Rev.62:434−363を参照)。
【0021】
本発明は、上記プラスミドによってコードされるアンチセンス配列がこれらのプラスミドのうちのいずれか一つの複製起点から転写されるRNAである、形質転換された宿主細胞を含む。この実施形態によると、上記染色体遺伝子に作動的に連結された調節配列、もしくはこの調節配列から転写されるmRNAは、このプラスミドの複製起点から転写されるRNAに対してアンチセンスである。このプラスミドが存在しない場合において、この染色体遺伝子に作動的に連結された調節配列は、転写され、そして/もしくは翻訳され、細胞増殖の阻害をもたらす。プラスミドが存在する場合において、複製起点から転写されるRNAは、この染色体遺伝子の作用を阻害し、それによって細胞増殖を可能にする。
【0022】
本発明のこの局面の好ましい実施形態によれば、上記プラスミドによってコードされるアンチセンス配列は、RNAIであり、上記染色体遺伝子の上流にある調節配列は、RNAIIもしくはその一部をコードする。あるいは、このプラスミドによってコードされるアンチセンス配列は、RNAIIであり、この染色体遺伝子の上流にある調節配列は、RNAIもしくはその一部をコードする。
【0023】
RNAIおよびRNAIIは、研究用途および商業的用途の事実上全てのプラスミドの複製起点によってコードされる、二つの重なるRNA転写産物である。RNAIIは、プラスミドDNA合成の開始を始め、E.coliにおいて自律的プラスミド複製がおこることを可能にする。RNAIは、RNAIIのアンチセンスインヒビターであり、過剰のプラスミド複製を妨げ、プラスミドのコピー数が、宿主細胞によって支持され得る数を上回らないことを保証する。現在、このアンチセンス機構が、形質転換された宿主細胞におけるプラスミド維持を促進するために利用され得ることが、見出されている。
【0024】
この実施形態によれば、本発明は、形質転換された宿主細胞を提供する。この宿主細胞は、
i)染色体遺伝子に作動的に連結された、RNAIIコード領域もしくはRNAIIコード領域の一部であって、その染色体遺伝子は細胞増殖を阻害する、RNAIIコード領域もしくはRNAIIコード領域の一部:および
ii)RNAIコード領域もしくはRNAIコード領域の一部を含む、プラスミド、
を含み、上記RNAIコード領域もしくは上記RNAIコード領域の一部から転写されるRNAIは、上記RNAIIコード領域もしくは上記RNAIIコード領域の一部から転写されるRNAIIに結合し、上記染色体遺伝子の作用を阻害し、それによって細胞増殖を許容する。
【0025】
上記染色体遺伝子に作動的に連結されたRNAIIコード領域もしくはRNAIIコード領域の一部は、転写されて、RNAII−染色体遺伝子mRNAが生成される。RNAIをコードするプラスミドが存在しない場合において、細胞増殖は阻害される。細胞増殖は、このRNAII−染色体遺伝子mRNAが、細胞増殖のために必須の第二の染色体遺伝子もしくはそのような第二の染色体遺伝子から転写されるmRNAに結合することによって阻害され得る。あるいは、このRNAII−染色体遺伝子は翻訳され得、そしてこの染色体遺伝子によってコードされる毒性タンパク質により、もしくは、細胞増殖のために必須の第二の染色体遺伝子を阻害する、この染色体遺伝子によってコードされるリプレッサータンパク質により、細胞増殖が阻害され得る。このプラスミドが存在する場合において、このプラスミドから転写されたRNAIは、RNAII−染色体遺伝子mRNA中のRNAIIに結合し、染色体遺伝子が細胞増殖を阻害することを防ぐ。
【0026】
本発明はさらに、形質転換された宿主細胞を提供する。この宿主細胞は、
i)染色体遺伝子に作動的に連結されたRNAIコード領域もしくはRNAIコード領域の一部であって、この染色体遺伝子は細胞増殖を阻害する、RNAIコード領域もしくはRNAIコード領域の一部:および
ii)RNAIIコード領域もしくはRNAIIコード領域の一部を含む、プラスミド、
を含み、上記RNAIIコード領域もしくはRNAIIコード領域の一部から転写されるRNAIIは、上記RNAIコード領域もしくはRNAIコード領域の一部から転写されるRNAIに結合し、上記染色体遺伝子の作用を阻害し、それによって細胞増殖を許容する。
【0027】
上記の染色体遺伝子に作動的に連結されたRNAIコード領域もしくはRNAIコード領域の一部は、転写されて、RNAI−染色体遺伝子mRNAを生成する。RNAIIをコードするプラスミドが存在しない場合において、細胞増殖は阻害される。細胞増殖は、このRNAI−染色体遺伝子mRNAが、細胞増殖に必須な第二の染色体遺伝子もしくはそのような第二の染色体遺伝子から転写されるmRNAに結合することによって、阻害され得る。あるいは、このRNAI−染色体遺伝子は翻訳され得、この染色体遺伝子によってコードされる毒性タンパク質によって、もしくは、細胞増殖に必須な第二の染色体遺伝子を阻害する、この染色体遺伝子によってコードされるリプレッサータンパク質によって、細胞増殖が阻害され得る。このプラスミドが存在する場合、このプラスミドから転写されるRNAIIは、RNAI−染色体遺伝子mRNA中のRNAIに結合し、この染色体遺伝子が細胞増殖を阻害することを妨げる。
【0028】
全てのプラスミドは、複製起点から転写されるRNAを産生し、ほとんどの商業的に入手し得るプラスミドは、RNAIコード領域およびRNAIIコード領域を含む複製起点を含む。用いられるプラスミド内へのlacオペレーター系のクローニングにプラスミドの維持を依存している、EP0851932において開示される系とは異なり、本発明は、形質転換される宿主細胞における、任意の複製起点を含む任意の商業的に入手し得るプラスミドの維持を可能にし、これらのプラスミド内に任意のさらなる配列をクローニングすることを必要とせずに、これらのプラスミドを維持する宿主細胞の選択を可能にする。このことは、プラスミドの大きさが縮小されて、DNAワクチンおよび遺伝子治療への適用におけるプラスミドの有効用量を増加するという利点を有する。不必要な細菌DNAの除去もまた、CpGジヌクレオチドに対する、免疫応答の危険性を低下させる。
【0029】
加えて、プラスミドの維持は、抗生物質選択を用いずに達成され得る。このプラスミドは、それゆえに、抗生物質耐性遺伝子の存在を必要としない。このプラスミドに抗生物質耐性遺伝子が存在しないことは、特にこのプラスミドもしくはこのプラスミドによってコードされる産物が治療に用いられることが意図される状況において、多くの利点を有している。第一に、このプラスミドに抗生物質耐性遺伝子が存在しないことは、抗生物質耐性遺伝子が潜在的な病原体を含めた環境内生物体に移動する危険性を取り除く。このプラスミドに抗生物質耐性遺伝子が存在しないことは、また、細胞から単離されたプラスミドがプラスミド選択のために用いられた残留抗生物質によって汚染されることを防止し、従って、残留抗生物質の存在についてプラスミドを試験する必要性、および抗生物質耐性遺伝子が患者の細胞において有害な効果をもたらす危険性を、取り除く。選択が抗生物質耐性に依存しないという事実は、培養中の抗生物質分解に起因するプラスミド選択圧の欠如の問題を回避する。
【0030】
本発明において使用するためのプラスミドとしては、複製起点から転写されるRNAが複製の調節に直接的に用いられるプラスミド(例えば、プラスミドR1、RK6、pT181、pMV158、およびpIP501)、および、複製起点から転写されるRNAが翻訳されてプラスミドの複製のために必要とされるタンパク質を提供するプラスミド(例えば、pSC101、pPS10、p15A、F、R100、R453、P1、RK2、RA1、RSF10110、pColIV−K30、ColE2、ColE3、Rts1、pLS20、およびpUB110)が挙げられる。好ましくは、本発明において使用するためのプラスミドは、RNAIコード領域およびRNAIIコード領域を含む複製起点を、含む。RNAIコード領域およびRNAIIコード領域を含む複製起点を含むプラスミドとしては、ColE1oriおよびpMB1oriのような、ColE1型oriを含むプラスミドが挙げられる。
【0031】
細胞増殖を阻害する染色体遺伝子に作動的に連結された、RNAIIコード領域、RNAIコード領域、もしくはそれらの一部は、構成的プロモーターもしくは誘導性プロモーターの制御下にあってもよい。好ましくは、細胞増殖を阻害する染色体遺伝子に作動的に連結された、RNAIIコード領域、RNAIコード領域、もしくはそれらの一部は、構成的プロモーターの制御下にある。適切な構成的プロモーターは、当該分野において公知であり、適切な構成的プロモーターとしては、例えば、trcプロモーターのようなE.coliプロモーターからの−35配列および−10配列を含むプロモーターが挙げられる。
【0032】
本発明の第二の局面によれば、細胞増殖を阻害する染色体遺伝子を含む宿主細胞が提供され、この染色体遺伝子は、その上流に位置する調節配列に作動的に連結されており、この調節配列は、プラスミドの複製起点から転写されるRNAの配列に対してアンチセンスである配列を含むか、もしくは、この調節配列は、プラスミド複製起点から転写されるRNAの配列に対してアンチセンスであるRNA配列をコードする。この調節配列の性質は、この宿主細胞が形質転換されることが意図されるプラスミドの特性に依存する。適切なプラスミド複製起点の例は、上記に提供される。これらの複製起点から転写されるRNAに対してアンチセンスである調節配列を設計する方法は、当業者にとって明らかである。
【0033】
好ましくは、この調節配列は、RNAIコード領域もしくはRNAIコード領域の一部、またはRNAIIコード領域もしくはRNAIIコード領域の一部を含む。細胞増殖を阻害する染色体遺伝子に作動的に連結された、これらのRNAIIコード領域、RNAIコード領域、もしくはそれらの一部は、構成的プロモーターもしくは誘導性プロモーターの制御下にあってもよい。好ましくは、細胞増殖を阻害する染色体遺伝子に作動的に連結された、これらのRNAIIコード領域、RNAIコード領域、もしくはそれらの一部は、構成的プロモーターの制御下にある。適切な構成的プロモーターは、当該分野において公知であり、適切な構成的プロモーターとしては、例えば、trcプロモーターのような、E.coliプロモーターからの−35配列および−10配列を含むプロモーターが挙げられる。
【0034】
本発明のこの第二の局面による宿主細胞は、本発明の第一の局面による形質転換された宿主細胞を産生するために、RNAIコード領域およびRNAIIコード領域を含むプラスミドを用いて形質転換されてもよい。
【0035】
本発明のこの宿主細胞および形質転換された宿主細胞は、インビトロにおいて培養中であってもよい。適切な宿主細胞の例は、本明細書中に提供される。
【0036】
本発明の第三の局面によれば、インビトロで宿主細胞内にプラスミドを維持する方法が提供され、この方法は、上記において記載されるように形質転換された宿主細胞を、この細胞が増殖することを許容するのに十分な条件下で培養する工程を含む。この方法は、抗生物質耐性が存在しない場合において、プラスミドの維持および選択が行われることを可能にし、このことは、この方法を治療への適用のためのプラスミドDNAもしくは組換えタンパク質の産生のために特に適切にする。本発明の第四の局面によれば、プラスミドDNAを産生する方法が提供され、この方法は、本発明の第三の局面の方法をおこなう工程およびプラスミドDNAを単離する工程を包含する。本発明の第五の局面によれば、組換えタンパク質を産生する方法が提供され、この方法は、本発明の第三の局面の方法に従う目的のタンパク質をコードするプラスミドを含む形質転換された宿主細胞を培養する工程、およびそのタンパク質をその細胞から単離する工程、を包含する。
【0037】
本発明の形質転換された宿主細胞および宿主細胞は、それら自体が治療への適用を有し得る。本発明の第六の局面によれば、治療における使用のための、本発明の第一の局面による形質転換された宿主細胞が提供される。
【0038】
本発明の第七の局面によれば、薬学的組成物が提供され、この薬学的組成物は、本発明の第一の局面による形質転換された宿主細胞と、それに加えて、薬学的に受容可能な賦形剤、希釈剤もしくは緩衝剤(例えば、当業者にとって周知である薬学的に受容可能な賦形剤、希釈剤もしくは緩衝剤)を含む。この局面の一つの実施形態によれば、この薬学的組成物は、ワクチン組成物である。このワクチン組成物は、賦形剤、希釈剤もしくは緩衝剤に加えて、一つ以上のアジュバントを含んでもよい。
【0039】
本発明の第八の局面によれば、薬の製造において、本発明の第一の局面による形質転換された宿主細胞の使用が、提供される。好ましくは、この形質転換された宿主細胞は、遺伝子送達系として用いられてもよい。例えば、この形質転換された宿主細胞は、遺伝子治療目的でプラスミドにある目的の遺伝子を送達するために用いられてもよい。この形質転換された細胞はまた、プラスミドにある目的の(その転写産物が治療的効果を有する)遺伝子(例えば、アンチセンスオリゴヌクレオチド)を送達するために、もしくは治療的効果を有するタンパク質をコードする目的の遺伝子を送達するために、用いられてもよい。
【0040】
あるいは、この形質転換された宿主細胞は、免疫原を送達するために用いられてもよい。この実施形態において、プラスミドにある目的の遺伝子は、免疫応答を惹起する一つ以上の抗原をコードしてもよい。当業者は、免疫応答を惹起する抗原をコードしていて本発明の形質転換された細胞内のプラスミドに含まれ得る、目的の遺伝子が分かる。
【0041】
本発明の第九の局面によれば、処置の方法が提供され、この処置の方法は、上記に記載された形質転換された宿主細胞、もしくはそれらを含む組成物を患者に投与する工程を、包含する。特に、目的の遺伝子を患者に送達する方法が提供され、この方法は、上記に記載される形質転換された宿主細胞もしくはそれらを含む組成物を患者に投与する工程を、包含する。上記に記載されるように、形質転換された宿主細胞内のプラスミドにある目的の遺伝子は、それ自体が遺伝子治療目的のために有用であってもよく、または、この目的の遺伝子の転写産物もしくは転写および翻訳の産物が、治療的特性を有してもよい。
【0042】
本発明の第十の局面によれば、病原体によって引き起こされる疾患に対して患者を免疫化する方法が提供され、この方法は、上記に記載される形質転換された宿主細胞もしくはそれらを含む組成物を患者に投与する工程を、包含する。この細胞内のプラスミドにおいて見出される目的の遺伝子は、この病原体に対する免疫応答を誘導する抗原をコードする。細胞の性質および目的の遺伝子の発現を制御するプロモーターの性質によって、この抗原はこの細胞が患者に投与される前にこの細胞において発現されてもよく、もしくは、投与後に患者において発現されるこの抗原と共に、このプラスミドを含む細胞が患者に投与されてもよい。好ましくは、上記で記載したように、この細胞は細菌細胞であり、この目的の遺伝子は、細菌のプロモーターもしくは真核細胞のプロモーターのいずれかに機能的に連結される。
【0043】
本発明の第十一の局面によれば、インビボの宿主細胞内にプラスミドを維持する方法が提供され、この方法は、本発明の第一の局面による形質転換された宿主細胞をレシピエント生物体に供給する工程を包含する。好ましくは、このレシピエント生物体は哺乳類、好ましくはヒトである。本発明のこの局面によれば、上記染色体遺伝子は、好ましくは、インビボで細胞増殖を阻害し、その結果、このプラスミドを維持する宿主細胞のみが、レシピエント生物体においてインビボで生存する。例えば、この染色体遺伝子は、この細胞の完全性にとって必須であるタンパク質をコードする第二の染色体遺伝子についてのリプレッサータンパク質をコードしてもよい。
【0044】
本発明における使用のために適切なプラスミドに運ばれる目的の遺伝子、宿主細胞、および細胞増殖を阻害する染色体遺伝子の例、ならびに本発明の形質転換された宿主細胞の医療への適用についての情報は、以下に提供される。
【発明を実施するための最良の形態】
【0045】
(染色体遺伝子)
本発明の宿主細胞および形質転換された宿主細胞内に含まれる染色体遺伝子は、増殖を阻害する任意の遺伝子であり得る。
【0046】
(増殖を直接的に阻害する染色体遺伝子)
細胞増殖を直接的に阻害する好ましい染色体遺伝子としては、トキシンをコードする遺伝子が挙げられる。本発明において用いられ得る、トキシンをコードする遺伝子の例としては、B.subtilisからのsacB遺伝子、E.coliのFプラスミド内のccdBによってコードされるCcdB、プラスミドRK2からのparD遺伝子によってコードされるKidタンパク質、制限エンドヌクレアーゼ遺伝子および抗生物質遺伝子が挙げられる。
【0047】
(増殖のために必須の遺伝子を阻害することによって増殖を間接的に阻害する染色体遺伝子)
増殖を間接的に阻害する好ましい染色体遺伝子としては、細胞増殖のために必須の第二の染色体遺伝子を阻害するタンパク質リプレッサーをコードする染色体遺伝子、および、細胞増殖のために必須の第二の染色体遺伝子を阻害するアンチセンス配列をコードする染色体遺伝子が挙げられる。本発明の一つの実施形態において、この第二の染色体遺伝子は、細胞増殖のために条件的に必須である。
【0048】
この染色体遺伝子がタンパク質リプレッサーをコードする場合、第二の染色体遺伝子は、好ましくは、そのリプレッサーによって従来的に阻害されるオペレーターおよびプロモーターに作動的に連結される。
【0049】
例えば、この染色体遺伝子がタンパク質リプレッサーLacIをコードするならば、細胞増殖のために必須の第二の染色体遺伝子は、好ましくはlacオペレーターおよびlacプロモーターに作動的に連結される。E.coliのlacリプレッサーは、「The Lactose Operon」(J.Beckwith、Escherichia coli and Salmonella typhimurium、J.L.Ingrahamら編、1987年、Amer.Soc.Micro.、1444〜1452頁)およびDicksonら(1975年、Science 187:27−35)において記載される。lacオペロンは以下のように調節される。非誘導条件下(例えば、ブドウ糖での増殖)では、LacIはlacオペロンのオペレーターに結合し、β−ガラクトシダーゼ(LacZ)、ラクトースパーミアーゼ(LacY)、およびトランスアセチラーゼ(LacA)の転写を妨げる。誘導性条件下(例えば、ラクトースでの増殖、もしくは代謝できないアナログであるIPTGの添加)では、リプレッサーはもはやオペレーターに結合せず、転写が起こる。オペロンの発現は、β−ガラクトシダーゼについてのアッセイによって容易に検出される。
【0050】
本発明に従う有用な他のリプレッサー系としては、真核細胞における遺伝子活性の調節において用いるためのtetリプレッサー系(Gossenら(1994)Current Opinions in Biotechnology,5,516−520)が挙げられる。tetリプレッサー系は、酵母、タマホコリカビ(dictiostelium)、植物細胞およびタバコ植物において用いられている。本発明に従うさらなる有用なリプレッサー系は、ArgRNVリプレッサー系(Burkeら(1994)Mol.Microbiol.13,609−618)である。ArgRリプレッサーは、正常にはアルギニンが存在する場合においてのみそのオペレーターに結合する。しかし、変異ArgRNVリプレッサーは、アルギニンが存在しない場合においてそのオペレーターに結合し、アルギニンが存在する場合においても結合したままであり、トランスドミナント(transdominant)効果を有する。二つの対称なArgボックスを有する理想化されたArgR結合部位(オペレーター)は、目的のプラスミド内へ操作され得、ArgR結合部位によって発現が制御される必須遺伝子からの、ArgRNVの結合除去を可能とする。
【0051】
本発明に従えば、E.coliのtrpリプレッサーもまた有用である(「The tryptophan Operon」、Yanof skyおよびCrawford、Escherichia coli and Salmonella typhimurium、J.L.Ingrahamら編、1987年、Amer.Soc.Micro.、1453〜1472頁を参照)。trpリプレッサーは、約50コピー/細胞で存在し、発酵培地中にリプレッサー結合の誘導因子としてトリプトファンの存在を必要とする。
【0052】
本発明に従えば、E.coliのgalRリプレッサーは有用である(「The Galactose Operon」、S.Adhya、Escherichia coli and Salmonella typhimurium、J.L.Ingrahamら編、1987年、Amer.Soc.Micro.、1503〜1512頁を参照)。
【0053】
本発明に従えば、E.coliのaraCリプレッサーもまた有用である(「The L−Arabinose Operon」、R.Schlief、Escherichia coli and Salmonella typhimurium、J.L.Ingrahamら編、1987年、Amer.Soc.Micro.、1473〜1481頁;Dunnら、1984年、Proc.Nat.Aca.Sci.81:5017−5020を参照)。araCリプレッサーは、アラビノースの存在する場合において結合親和性が増大する。
【0054】
最後に、本発明に従えば、Aリプレッサーは有用である(Introduction to Lambda Phages、Current Protocols in Molecular Biology、Ausubelら編、1994年、第3節、ユニット1.9;Hochschildら、1986年、Cell 47(5):807−816)。
【0055】
細胞増殖のために必須の第二の染色体遺伝子は、細胞の代謝産物の生合成に関係する産物をコードする遺伝子、その遺伝子の産物が炭素代謝に関与する遺伝子、または、高分子の生合成もしくは調節をコードする遺伝子(例えば、DNAおよび/もしくはRNAの合成機能および複製機能のために必須の遺伝子)であってもよい。
【0056】
細胞の構成要素の供給、特に細胞壁の前駆体の供給と関与する酵素をコードする特定の遺伝子は、細胞増殖のために必須であり、本発明に従って、有用である。例えば、細菌の細胞壁は、メソジアミノピメリン酸(DAP)を含み、この構成要素を合成することが出来ないことは、溶菌をもたらす。DAP生合成経路内の遺伝子、すなわち、dapA遺伝子、dapB遺伝子、dapC遺伝子、dapD遺伝子、およびdapE遺伝子は、それゆえ、第一の染色体遺伝子によって阻害される、細胞増殖のために必須の第二の染色体遺伝子として用いられ得る。dapAおよびdapBは、クローニングされて塩基配列が決定されており、dapBはひとつのシストロンとして利用可能である(Richaudら、J.Bacteriol.166:297−300,1986年;Bouvierら、J.Biol.Chem.259:14829−14834,1984年)。
【0057】
他の細胞壁構成要素の生合成(例えば、D−アラニン生合成)に関与する遺伝子もまた本発明に従って有用である(Walsh、1989年、J.Biol.Chem.264(5):2393−2396)。
【0058】
脂肪酸生合成に関与する遺伝子もまた、本発明に従って有用である。一つの例は、fabAであり、fabAは、3−ヒドロキシデカノイル−ACPデヒドラーゼをコードしており、不飽和脂肪酸生合成と飽和脂肪酸生合成とが分岐する点での、伸長する脂肪酸鎖への二重結合の導入の原因である(Cronan Jr.J.E.およびRock.C.O.(1987)Biosynthesis of membrane lipids、「Escherichia coli and Salmonella typhimurium:cellular and molecular biology」。Neidhardt.F.C.編、American Society of Microbiology,Washington D.C.)。この酵素は比較的高い濃度で存在し、fabA変異体は、不飽和脂肪酸が補われないと溶菌する。
【0059】
あるいは、細胞増殖に必須の第二の染色体遺伝子は、炭素源の利用と関与する遺伝子であってもよい。具体的には、この第二の染色体遺伝子は、ラクトースオペロンの遺伝子を含んでもよく、その結果、ラクトースが唯一の炭素源である条件において、プラスミドを維持しておりlacオペロンの遺伝子が発現される細胞のみが、生存し得る。他の改変は、当業者にとって明らかである。
【0060】
グルタミンシンテターゼは、NSOミエローマ細胞株(Bebbingtonら、(1992)Bio/Technology 10,169−175)のような真核細胞にとっての必須遺伝子であって、好ましくは、宿主細胞が真核細胞である場合に第二の染色体遺伝子として用いられる。
【0061】
この第二の染色体遺伝子はまた、宿主細胞のDNAおよび/またはRNAの合成タンパク質または複製タンパク質をコードする必須遺伝子であってもよい。E.coliおよびSalmonellaのような細菌におけるこれらの必須機能に関するそのような遺伝子の例は、McMackenら(Escherichia coli and Salmonella typhimurium,Cellular and Molecular Biology、Neidhardtら編、Amer.Soc.Micro,,Wash.D.C.,1987年、564〜612頁)において提供され、以下の遺伝子を含むが、以下の遺伝子に限定されない:dnaA、dnaB、dnaC、ssb、dnaG、polC(dnaE)、dnaQ(mutD)、dnaN、dnaZX、gyrA、gyrB、polA、lig、dnaT、rpoA、rpoB、rpoC、およびrpoD。
【0062】
好ましくないが、この第二の染色体遺伝子は、抗生物質耐性遺伝子であってもよく、その結果、プラスミドを用いるその株の形質転換が、その抗生物質の存在する場合における増殖を可能にする。
【0063】
医療への適用に使用するための形質転換された宿主細胞に対して、投与経路に依存して、局所環境内の多くの要因が選択圧をかけ得る。そのような圧力に抵抗する能力が同定される遺伝子に連鎖しているならば、この遺伝子は、選択マーカーとして用いられ得る。例えば、細胞、特に経口で投与されることになる細菌細胞にとって、胃の内容物の低いpHへの抵抗性は、重要な選択的利点を構成する。このことと関連することが示される遺伝子のうちには、Streptococcus mutansのuvrA遺伝子(Hannaら、2001年、J.Bacteriol 183:5964)、E.coli.のgadA遺伝子、gadB遺伝子、gadC遺伝子(Castanie−CornetおよびFoster、2001年 Microbiology 147:709)、Salmonella typhimuriumのrpoS遺伝子、fur遺伝子、atp遺伝子、およびatbR遺伝子(Baikら、1996年 Microbiology 142:3195)、Helicobacter pyloriのlepA遺伝子、Frasel遺伝子、czcA遺伝子、uvrA遺伝子、atpF’遺伝子、およびアルド−ケトレダクターゼ遺伝子(Bijlsmaら、2000年、J Infect Dis 182:1566)、ならびに、Listeria monocytogenesのF0F1ATPアーゼをコードする遺伝子(Cotterら、2000年、Int J Food Microbial 60:137)がある。
【0064】
細胞内での生存を与える遺伝子(例えば、Listeria monocytogenesのSvpA(Borezeeら、2001年、Microbiology 147:2913)もしくは、リソソームのような特定の細胞内局在に対する抵抗力もまた、適切な宿主細胞において有用であり得る。
【0065】
細胞増殖を間接的に阻害する好ましい遺伝子としては、細胞増殖のために必須な第二の染色体遺伝子に結合することによって、または細胞増殖のために必須な第二の染色体遺伝子から転写されるmRNAに結合することによって、この第二の染色体遺伝子を阻害するアンチセンス配列をコードする、染色体遺伝子も挙げられる。
【0066】
アンチセンス配列とは、このアンチセンス配列の標的配列に実質的に相補的である配列であって、その標的配列に特異的にハイブリダイゼーションする能力を有する。このアンチセンス配列は、細胞増殖のために必須な第二の染色体遺伝子に直接的に結合するか、またはその第二の染色体遺伝子から転写されるmRNAに結合することのいずれかによって作用し得るので、この標的配列は、この第二の染色体のコード鎖もしくは非コード鎖のいずれかの配列、またはそれらの配列の一部であってもよい。標的配列に特異的にハイブリダイゼーションする能力を有するアンチセンス配列を調製する方法は、当業者によって公知である(例えば、Cohen,J.S.,Trends in Pharm.Sci.,10,435(1989)、Okano,J.Neurochem.56,560(1991);O’Connor,J.Neurochem 56,560(1991);Leeら、Nucleic Acids Res 6,3073(1979);Cooneyら、Science 241,456(1988);Dervanら、Science 251,1360(1991)を参照)。
【0067】
好ましくは、このアンチセンス配列は、ストリンジェントな条件の下で、その標的配列にハイブリダイゼーションする。高ストリンジェンシーハイブリダイゼーション条件は、以下のように定義される:50%のホルムアミド、5×SSC(150mM NaCl、15mMのクエン酸三ナトリウム)、50mMのリン酸ナトリウム(pH7.6)、5×デンハルト液、10%のデキストラン硫酸、および20マイクログラム/mlの変性剪断サケ精子DNAを含む溶液中での、42℃での一晩のインキュベーションと、その後の0.1×SSC中でのおよそ65℃でのフィルターの洗浄。
【0068】
上記に示されるように、第一の染色体遺伝子は、細胞増殖のために必須な遺伝子の阻害もしくは細胞増殖を阻害する遺伝子の活性化を最終的にもたらす、多くの遺伝子を含む反応のカスケードを開始することによって、細胞増殖を阻害し得る。例えば、この第一の染色体遺伝子は、第二の染色体遺伝子を阻害するリプレッサータンパク質をコードし得、この第二の遺伝子は、細胞増殖のために必須な第三の染色体遺伝子を他の状態では活性化するアクチベーターをコードする。あるいは、この第一の染色体遺伝子は、第二の染色体遺伝子を阻害するリプレッサータンパク質をコードし得、この第二の染色体遺伝子は、細胞増殖を阻害する第三の染色体遺伝子を他の状態では阻害するアンチセンス配列をコードする。これらが本発明の必要条件を満たす遺伝子の多くのカスケードの例に過ぎないことは、当業者にとって明らかである。
【0069】
(改変された宿主細胞の産生)
上記の細胞増殖を阻害する染色体遺伝子は、宿主細胞のゲノム内に天然に存在する遺伝子であってもよく、あるいは外来遺伝子であってもよい。
【0070】
プラスミドによってコードされるアンチセンス配列が、上記の染色体遺伝子に作動的に連結される調節配列に結合することによってこの染色体遺伝子を阻害する場合、この染色体遺伝子は、外因性の調節配列に作動的に連結されている、天然に存在する遺伝子であってもよい。例えば、天然に存在するトキシン遺伝子の発現を通常では制御する配列が除去されてもよく、このトキシン遺伝子が代わりに、構成的プロモーターの制御下で、外因性のRNAIコード配列に作動的に結合されてもよい。同様に、天然に存在するリプレッサータンパク質の発現を通常では制御する配列が除去されてもよく、このリプレッサータンパク質遺伝子が代わりに、構成的プロモーターもしくは誘導性プロモーターの制御下で、外因性のRNAIIコード配列に作動的に連結されてもよい。従って、本明細書中の実施例2において、天然に存在するリプレッサータンパク質であるlacIは、RNAIIコード領域に作動的に結合される。
【0071】
あるいは、この調節配列がRNAIコード配列もしくはRNAIIコード配列であって、この染色体遺伝子が細胞増殖のために必要である第二の染色体遺伝子を阻害するアンチセンス配列をコードする場合のように、この調節配列およびこの染色体遺伝子の両方が外因性であってもよい。
【0072】
細胞増殖のために必須な第二の染色体遺伝子が存在するならば、この第二の染色体遺伝子もまた、天然に存在する遺伝子または外来遺伝子のいずれであってもよい。
【0073】
細胞増殖を阻害する染色体遺伝子は、特に宿主細胞が真核細胞の宿主細胞である場合において、この細胞のゲノム内に複数コピーで存在し得る。
【0074】
本発明において用いるために適切な宿主細胞を産生するために、細胞を操作する方法は、当業者にとって明らかである。
【0075】
(宿主細胞)
本発明は、動物細胞(例えば、哺乳類細胞および昆虫細胞)、植物細胞、真菌(例えば、酵母)、細菌、および古細菌を含む、全ての細胞型に対して適用され得る。
【0076】
宿主細胞が細菌細胞である場合、この宿主細胞は、グラム陰性細菌細胞であってもよく、グラム陽性細菌細胞であってもよい。この細胞がグラム陰性細菌細胞である場合、この細胞は、好ましくはE.coli細胞もしくはSalmonella細胞である。この細胞がグラム陽性細菌細胞である場合、この細胞は、好ましくはBacillus、Streptomyces、Lactobacillus、もしくはLactococcus細胞である。
【0077】
特に有用であるのは、レシピエント生物体における治療用途が自由な、受容可能な程度にまで副作用が無いように弱毒化されている、弱毒化された宿主細胞である。この細胞の特性に依存して、この細胞は、獣医学的用途もしくはヒトの治療用途のために適切であり得る。
【0078】
入手し得る弱毒化された細菌の中では、以下のものがある:
SL3261株(aroA変異体、Titballら、1997年、Infection and Immunity 65:1926、Titballら、1995年、Infection and Immunity 63:563)、VNP20009株
(purlおよびmsbBの変異体、Tosoら、2002年、J.Clin Oncol 20:142)を含む、Salmonella typhimurium;
CVD 908−htrA株、χ4073株、χ4632株、Ty800株、CVD 909株、およびCVD 915株を含む、Salmonella typhi(Galenら、1999年、Infect.Immun.67:6424−6433;Moronaら、1991年、Gene 107:139−144;Tacketら、1997年、Infect.Immun.65:3381−3385;Garmoryら、2002年、FEMS Microbiology Reviews 26:339−353);
9R株を含む、Salmonella gallinarum(鳥類のワクチン接種のため、Feberweeら、2001年、Avian Dis 45:1024);
aroA変異体、cnfl変異体、およびOMP変異体を含む、Escherichia coli(Rippere−Lampeら、2001年、Infect Immunol 69:3954);
S.flexneri 2a guaBA株を含む、Shigella flexneri(Altboumら、2001年、Infect Immunol 69:3150);
Peru2株を含む、Vibrio cholerae(Ryanら、2000年、Infect.Immun.68:221−226);
PrfA変異体およびSvpA変異体を含む、Listeria monocytogenes(Sheehanら、1996年、Mol Microbiol 20:785;Borezeeら、2001年、Microbiology 747:2913);
RB51株を含む、Brucella abortus(Vemulapalliら、2000年、Infect Immunol 68:3290);
Calmette−Guerin(BCG)株を含む、Mycobacterium bovis;および
Mycobacterium tuberculosis。
【0079】
(本発明に従って有用である、プラスミドによって運ばれる遺伝子)
本発明の形質転換された宿主細胞内のプラスミドで運ばれる目的の遺伝子は、任意の遺伝子であり得る。この遺伝子は、レシピエント生物体に送達された場合に治療効果を有する遺伝子であり得る。例えば、遺伝子治療目的のためにレシピエント生物体にこの遺伝子を送達することが所望され得る。
【0080】
あるいは、遺伝子の転写産物もしくは転写および翻訳の産物が治療効果を有するので、この遺伝子をレシピエント生物体に送達することが所望され得る。例えば、目的の遺伝子の転写産物は、アンチセンスオリゴヌクレオチドであってもよく、あるいは、この目的の遺伝子は、治療効果を有する抗原をコードしてもよい。特に、目的の遺伝子は、レシピエント生物体において免疫応答を惹起する一つ以上の抗原をコードしてもよい。レシピエント生物体において免疫応答を誘導するためにプラスミドに含まれ得る目的の遺伝子の例は、cafオペロンのcafI遺伝子、caf1A遺伝子、caf1M遺伝子がある。
【0081】
目的の遺伝子が転写されるもしくは転写されて翻訳されることが望ましい場合、この目的の遺伝子は、好ましくはプロモーターと機能的に結合する。好ましくは、このプロモーターは真核生物のプロモーターもしくは細菌のプロモーターである。目的の遺伝子と機能的に結合するプロモーターの選択、および宿主細胞の選択は、宿主細胞において、レシピエント生物体への投与に先立って、もしくはレシピエント生物体への投与の後でのみ、目的の遺伝子が転写されるか、または転写されて翻訳されるかを決定する。
【0082】
いくつかの場合において、目的の遺伝子を既に発現している宿主細胞に送達することが、所望され得る。この場合において、この目的の遺伝子と機能的に結合しているプロモーターは、その結果として、このプロモーターがこの宿主細胞内で作動し、この宿主細胞内におけるこの目的の遺伝子の転写および翻訳を促進するように、選択される。例えば、この宿主細胞が細菌細胞である場合、この状況においてこの目的の遺伝子と機能的に結合しているプロモーターは、細菌のプロモーターであってもよい。
【0083】
他の場合において、この目的の遺伝子を含むプラスミドを含む宿主細胞をレシピエント生物体に送達するが、この目的の遺伝子の発現をこの細胞のレシピエント生物体への送達後にまで遅延させることが、所望され得る。例えば、この宿主細胞が細菌細胞である場合、この状況においてこの目的遺伝子と機能的に結合しているプロモーターは、この宿主細胞においてこの目的の遺伝子の転写および翻訳を促進しないが、レシピエント生物体への投与の後にこの目的の遺伝子の転写および翻訳を促進する、真核生物のプロモーターであってもよい。
【0084】
(本発明に従って有用であるワクチン接種の方法)
本発明によって形質転換された宿主細胞を用いるワクチン接種は、多くの投与経路のいずれによってであってもよいことが、予想される。Mycobacteriumについては、例えば、有用な経路は、皮内注射もしくは皮下注射によってである。Salmonella、Shigella、もしくはE.coliのような腸内細菌については、経口投与(特に、耐酸性形質転換体の経口投与)または直腸投与が好ましい。いくつかの状況においては、粘膜投与、静脈内投与、もしくは腹腔内投与が、好適であり得る。
【0085】
(他のインビボでの適用のための投与)
非ワクチン接種適用のための多くの投与方法が用いられ得ることが、予想される。
【0086】
ワクチン接種のために用いられる経腸投与経路および非経口投与経路に加えて、非ワクチン適用は、他の手順を必要とし得る。他の手順の中では、本発明による細胞が中に含まれる移植可能な浸透性の容器の使用がある。
【0087】
本発明は、ここで、一例としてプラスミドの複製起点を選択マーカーとして使用するプラスミド維持系を参照して、さらに詳細に記載される。実施例において記載される系に対して変更がなされ得ることが、理解される。
【実施例】
【0088】
本発明は、複製起点(pMB1 oriもしくはColEI ori)のみを選択マーカーとして用いてプラスミドの選択およびプラスミドの維持を可能とし、本明細書中でoriSELECTと言及される。図1において示すように、これらのColEI適合性oriは、二つの重なるRNA転写産物をコードする:RNAIIは、プラスミドDNA合成の開始を始め、RNAIは、RNAIIのアンチセンスインヒビターである(Scottら、1984,Microbiol.Rev.48:1−23;Chanら、1985,J.Biol.Chem.260:8925−8935)。研究用途もしくは商業的用途の事実上全てのプラスミドは、E.coli内での自律複製をこれらの複製起点に依存している。
【0089】
本発明は、あるオペロンにおいて阻害的遺伝子エレメントに連結されたRNAIIコード領域を含むようにゲノムが改変されているE.coliの形質転換株の構築を、含み得る。プラスミド産生性RNAIのRNAII−mRNA融合体へ結合は、その阻害遺伝子mRNAの転写を妨害し、それによって細胞の生存および増殖を可能にする。このプラスミド維持系の3つの提案される機構は、以下に記載される。
【0090】
(実施例1:リプレッサーoriSELECT)
図2において示すように、リプレッサーoriSELECTにおける使用のための宿主細胞のゲノムは、リプレッサー遺伝子シストロンに連結されたRNAIIコード領域を含み、RNAIIコード領域およびリプレッサー遺伝子シストロンの両方は、構成的RNAIIプロモーターの制御下にある。リプレッサーシストロンに連結されたRNAIIコード領域はまた、図8において示されるような、E.coliのTrcプロモーターにおいて見出される−10配列および−35配列ならびにスペーシングに基づくプロモーターのような、代替的な構成的プロモーターの制御下にあり得る。宿主細胞のゲノムはまた、誘導性プロモーター(inducible promoter)の制御下での細胞増殖のために必須な遺伝子を含む(図8においては示さない)。
【0091】
プラスミドが無い場合において、リプレッサータンパク質が発現し、そして必須遺伝子(essential gene)の発現を制御するオペレーター/プロモーターに結合することによって必須遺伝子を抑制し、細胞増殖を阻害する。プラスミド産生性RNAIの、RNAII−リプレッサーmRNA融合体内のRNAII転写産物への結合は、リプレッサーの翻訳を妨げ、必須遺伝子の発現を可能にし、それによって細胞増殖を可能にする。
【0092】
リプレッサーoriSELECTにおける使用のためにこの宿主細胞を構築するために、dapD(Richaudら、1984,J.Biol.Chem.259:14824−14828)もしくはfabA(Cronanら、1987,Biosynthesis of membrane lipids、「Escherichia coli and Salmonella typhimurium:cellular and molecular biology」,Neidhardt,F.C.編、American Society of Microbiology,Washington D.C.)のような必須遺伝子を、細菌株の染色体から欠失させ、誘導性プロモーター(例えば、Plac)の制御下に置き、そしてその染色体内へ挿入し戻す。その誘導性プロモーターに結合している、リプレッサーをコードしている遺伝子(この場合においてはlacI)を、染色体から欠失させる。このリプレッサー遺伝子を、両方の転写を駆動する構成的RNAIIプロモーター(もしくは任意の他の構成的プロモーター、例えばE.coliのtrcプロモーターにおいて見出される−10配列および−35配列ならびにスペーシングに基づくプロモーター)と共に、RNAIIの一部をコードしている領域の下流のオペロン内に置く。RNAIIコード領域の3’末端を、リプレッサーのリボソーム結合部位を含むように改変し、このためRNAIIコード領域の3’末端は重なり、RNAIによって妨げられる。いくつかの場合においては、RNAIIコード領域の3’末端とlacのリボソーム結合部位との間の距離を増大させることによって、LacIリプレッサータンパク質の発現を増加させ得る。この構築物を、染色体の中へ挿入する。
【0093】
その細胞はそれからリプレッサーを構成的に産生する。リプレッサータンパク質は、必須遺伝子を制御している染色体オペレーターに結合し、それによって細胞増殖を妨害する。その細胞をColEI適合性oriを含むプラスミドを用いて形質転換するとき、RNAIは、RNAIIに結合し、そして下流のリプレッサー遺伝子の翻訳を妨げ、細胞増殖を可能にする。
【0094】
染色体の必須遺伝子がその必須遺伝子自体の結合したリプレッサーを有するのであれば、このリプレッサー(マイナスそのリプレッサーのプロモーター)をRNAIIの下流のオペロン内へ操作してもよく、そしてこのオペロンを野生型リプレッサーと置き換えさせるために細菌の染色体内へ挿入してもよい。
【0095】
上記の必須遺伝子は、条件的に必須な外来遺伝子(例えば、抗生物質耐性遺伝子)であり得る。
【0096】
必須な外来遺伝子が誘導性であり、かつその遺伝子自体の結合した外来リプレッサーを有するのであれば、このリプレッサー遺伝子をRNAIIの下流のオペロン内へ操作してもよく、そしてこのオペロンを細菌の染色体内へ挿入してもよい。
【0097】
(実施例2:トキシン遺伝子oriSELECT)
図3において示すように、トキシン遺伝子oriSELECTにおける使用のための宿主細胞のゲノムは、RNAIIの下流に配置されたBacillus subtilisからのレバンスクラーゼ(Linkら、1997,J.Bacteriol.179:6228−6237)をコードするsacB遺伝子を含む。ショ糖が存在する場合において、sacBは毒性の産物を合成し、RNAIを発現するプラスミドを導入しなければ、細胞は死ぬ。誘導性プロモーターによって調節される別の毒性遺伝子を、sacBの代わりにRNAIIコード領域へ融合してもよい。
【0098】
構築物を、RNAIIコード領域およびsacBをコードする下流の遺伝子からなるオペロンを用いて操作する。RNAIIコード領域の3’末端を、sacBのリボソーム結合部位を含むように改変する。構成的RNAIIプロモーターは、遺伝子発現を駆動する。構成的RNAIIプロモーターを細菌の染色体内へ挿入する場合、その細胞は、ショ糖がない場合においても増殖し得るが、ショ糖を加えると、トキシンが産生されそして細胞は死ぬ。その細胞を、ColEI適合性oriを含むプラスミドを用いて形質転換する場合、RNAIがRNAII−トキシン遺伝子mRNAに結合し、そして翻訳を妨げ、ショ糖が存在する場合においても細胞増殖を可能にする。
【0099】
温度感受性プロモーターもしくは化学的に誘導されるプロモーターを、RNAII−トキシン遺伝子融合体の発現を駆動するために用いてもよい。許容温度において、もしくは誘導物質が存在する場合においては、RNAIを発現するプラスミドを導入しなければ、細胞は死ぬ。
【0100】
(実施例3:二重アンチセンスoriSELECT)
図4において示すように、二重アンチセンスoriSELECTにおいて、宿主細胞のゲノムは、細胞増殖のために必須である遺伝子、および細胞増殖のために必須である遺伝子に対するアンチセンスである遺伝子に連結されたRNAIIコード領域を、含む。
【0101】
必須遺伝子(例えば、dapDもしくはfabA)のアンチセンスバージョンを、RNAIIコード領域の下流のオペロン内に、必須遺伝子のアンチセンスバージョンおよびRNAIIコード領域の両方の転写を駆動する構成的RNAIIプロモーターと共に置く。RNAIコード領域の3’末端を、必須遺伝子アンチセンス転写産物の一部を含むように、それによってRNAIコード領域の3’末端がRNAIIと部分的に重なるように改変してもよい。この構築物を、細菌の染色体内へ挿入する。
【0102】
その細胞はそれからRNAIIおよびアンチセンスRNAを構成的に産生する。アンチセンスRNAは、必須遺伝子mRNAに結合し、それによって必須遺伝子の翻訳および細胞増殖を妨害する。
【0103】
その細胞をColEI適合性oriを含むプラスミドを用いて形質転換する場合、RNAIはRNAII−アンチセンスRNAに結合し、RNAII−アンチセンスRNAが必須遺伝子mRNAに結合することを妨害し、細胞増殖を可能にする。
【0104】
(実施例4:リプレッサーoriSELECT株の製作)
ORT株DH1lackan(Williansら、1998)、DH1lacdapDおよびDH1lacP2dapD(Cranenburghら、2001,Nucleic Acid Res.29:e26)を、RNAII−lacIオペロンの染色体のlacIリプレッサー遺伝子への組込みによって(そしてそれによってRNAII−lacIオペロンで染色体lacIリプレッサー遺伝子を置き換えることによって)oriSELECT株に転換してもよい。pKO3組込み系(Linkら、1997,J.Bacteriol.179:6228−6237)に基づいて構築された単一プラスミドは、以下のように、全てのORT株において、野生型lacIをRNAII−lacIで置換するために用いられる。
【0105】
1.lacIを取り囲む染色体lacオペロンの一部を、PCRによって増幅し、クローニングする;
2.lacIリプレッサー遺伝子の上流に挿入されたRNAIIコード領域からなるオペロンを操作し、天然のプロモーターと置き換える。lacIのリボソーム結合部位は、mRNA上でアンチセンスRNAIIの結合によって妨げられるように、配置されるべきである。
【0106】
3.RNAII−lacIオペロンの遺伝子座を、組込みプラスミドpKO3内へクローニングする。
【0107】
4.RNAII−lacIオペロンの遺伝子座を、DH1lackan、DH1lacdapD、およびDH1lacP2dapDの野生型lacオペロンの遺伝子座内へ組込んで、oriSELECT株を産生する。
【0108】
5.これらの株におけるプラスミドの選択およびプラスミドの維持を、ColE1/pMB1 ori配列を保有しかつlacO配列を保有しない(ORT選択を避けるため)プラスミドを用いて試験する。
【0109】
このoriSELECT株の構築のために必要であるクローニングの工程を、より詳細に以下に記載する。
【0110】
(RNAII−lacI融合体を産生するスプライシングPCR)
1.図5において示すように、pUC18から、RNAIIの5’末端の一部を、以下のプライマー(産物=176bp)を用いるPCRによって増幅する。
【0111】
5RNAII:GAATGCATCAAAGGATCTTCTTGAGA(26nt)
3RNAII:ACATTCACCACCGTATTTGGTATCTGCGCTCTGCTGAAGCCA(42nt)。
【0112】
2.図6に示すように、DH1 gDNAから、lacIの5’末端の一部を、以下のプライマーを用いるPCRによって増幅する(産物=597bp)。
【0113】
5lacI:GATACCAAATACGGTGGTGAATGTGAAACCAGTAACGTTATA(42nt)
3lacI:ACAGAACTTAATGGGCCCGCTAACA(25nt)。
【0114】
3.図7において示すように、5RNAIIプライマーおよび3lacIプライマーを用いて、一回のPCRにおいてこれら二つのPCR産物を組み合わせ、増幅して、749bpのRNAII−lacI遺伝子融合体PCR産物(以下の5RNAIIプライマーと3lacIプライマーとの間の領域)を生成する。
【0115】
4.スプライシングされたPCR産物を、NsiIおよびApaIを用いて切断して、lacIプロモーターおよび5’末端の位置においてラクトースオペロン内へクローニングするための断片を生成する。
【0116】
(ラクトースオペロンの一部のクローニング、およびRNAII−lacIの挿入)
1.DH1 gDNAからのlacオペロンを、以下のプライマーを用いて増幅する。
【0117】
【化1】
プライマー5LOは、天然のSalI部位(下線)を含むが、3LOは、1ヌクレオチド変化(太字)を有するSalI部位を導入する。総PCR産物の大きさは、5803bpである。
【0118】
2.上記のPCR産物をSalIで切断し、そしてSalIを用いて切断されたpUC18内へクローニングする。
【0119】
3.このプラスミドをNsiIおよびApaIで切断し、そして工程4による断片にクローニングし、プロモーターおよびlacIの5’末端をRNAII−lacI遺伝子融合体で置換する。
【0120】
4.挿入部分をSalIを用いて切断し、そしてSalIを用いて切断されたpKO3recA内へライゲーションする。
【0121】
5.oriSELECT株を産生するために、DH1lacdapD、DH1lacP2dapD、およびDH1lackanの染色体内へ組込む。
【0122】
6.pMB1複製起点を有するプラスミドを用いて、プラスミドの選択およびプラスミドの維持を試験する。
【0123】
RNAII構成的プロモーターをRNAII−遺伝子融合体発現を駆動するために用いる場合、必要であれば、任意の他の構成的プロモーターを置換してもよい。そのRNAIIプロモーターをこれらの実施例において既に存在するものとして用いる。代替的な好ましい構成的プロモーターは、至適なE.coliのtrcプロモーターにおいて見出される−10配列および−35配列ならびにスペーシングに基づくプロモーターである。このE.coliプロモーターは、RNAIIプロモーターより強く、それゆえに、より大量のLacIリプレッサータンパク質を産生し、それによってプラスミドが存在しない場合において増殖が妨げられることを保証することが望まれる場合に用いてもよい。LacIリプレッサータンパク質の発現はまた、RNAIIコード領域の3’末端とlaciIのリボソーム結合部位との間の距離を長くすることによって増大されてもよい。
【0124】
また、遺伝子カセットを挿入する代替的な方法を、上記に記載したpKO3法の代わりに用いてもよいことが理解される。例えば、ラムダRed組換え系を用いて、直鎖状PCR産物としてこのカセットを組込んでもよい(Murphy,1998,J.Bacteriol.,180:2063−2071)。
【0125】
これらの実施例は、アンチセンスインヒビターとして機能するRNAIを用いるRNAII−遺伝子融合体発現カセットに基づく。このことは、なぜなら、プラスミドを含む細胞においては、RNAIはRNAIIを五倍の過剰量で上回って存在するからであり(Liangら、1999,J.Mol.Biol.292:19−37)、ゆえに、モル過剰のRNAIが利用可能であるかのように、RNAIがより効果的なインヒビターとして働き得るからである。しかし、RNAIIよりもRNAIを用いて遺伝子融合物を構築することが有利である状況はあり得る。この場合において、アンチセンスインヒビターとして働くのは、プラスミドからのRNAII転写産物である。
【0126】
ColEI/pMB1 oriからのRomタンパク質を発現する遺伝子の染色体組込みは、この染色体組み込みがRNAIのRNAIIとの結合親和性を上昇させるので、oriSELECT株において必要であり得る(Chanら、1984,J.Biol.Chem.260:8925−8935)。
【図面の簡単な説明】
【0127】
【図1】pUC18oriの図であり、RNAI転写産物およびRNAII転写産物の位置を示す。RNAIIは、プラスミドのDNA合成の開始を始め、RNAIは、RNAIIのアンチセンスインヒビターである。
【図2】リプレッサーoriSELECTにおいて用いるための宿主細胞のゲノムの例。宿主細胞のゲノムは、リプレッサー遺伝子のシストロンに連結されたRNAIIコード領域を含み、これらの両方が構成的RNAIIプロモーターの制御下にある。このゲノムはさらに、誘導性プロモーターの制御下にある必須遺伝子を含む。プラスミドが存在しない場合において、リプレッサータンパク質が発現され、必須遺伝子を抑制し、細胞増殖を阻害する。プラスミドによって生成されるRNAIの、RNAII−リプレッサーmRNA融合体のRNAII転写産物への結合は、リプレッサータンパク質の翻訳を遮断し、必須遺伝子の発現および細胞増殖を可能にする。
【図3】トキシン遺伝子oriSELECTにおいて用いるための宿主細胞のゲノムの例。このゲノムは、Bacillus subtilis由来のレバンスクラーゼをコードし(Linkら、1997年、J.Bacteriol.179:6228−6237)RNAIIの下流に配置されたsacB遺伝子を含む。ショ糖が存在する場合において、sacBは毒性の産物を合成し、トキシン遺伝子のmRNAの翻訳を遮断するRNAIを発現するプラスミドが存在しない限り、ショ糖が存在する場合においてその細胞は殺される。
【図4】アンチセンスoriSELECTにおいて用いるための宿主細胞のゲノムの例。この宿主細胞のゲノムは、細胞増殖のために必須である遺伝子、および、細胞増殖のために必須な遺伝子に対するアンチセンスである遺伝子に連結されたRNAIIコード領域を含む。プラスミドが存在しない場合において、このアンチセンスRNAは必須遺伝子のmRNAに結合し、それによってこの必須遺伝子の翻訳および細胞増殖を妨げる。この細胞がRNAIを与えるプラスミドを用いて形質転換される場合、RNAIはRNAII−アンチセンスRNAに結合し、RNAII−アンチセンスRNAが必須遺伝子のmRNAに結合することを妨げ、細胞増殖を可能にする。
【図5】PCRによるpUC18からのRNAIIの5’末端の増幅。RNAI転写産物およびRNAII転写産物の位置は、リプレッサーoriSELECTにおいて用いるための宿主細胞のゲノムへの挿入のための、リプレッサーlacIをコードする遺伝子の上流へのRNAIIの挿入に先立って、RANIIの5’末端を増幅するために用いられるプライマーの位置であるとして、示される。
【図6】PCRを用いる、DH1 ORT株のゲノムDNA由来のlacIの5’末端の増幅。この増幅において用いられるプライマー5lacIおよび3lacIの位置が示される。
【図7】図5中のプライマーを用いて産生されるRNAII増幅産物と、図6中のプライマーを用いて産生されるlacI増幅物とを組み合わせることによって形成される産物。この組合せ産物の増幅において用いられるプライマーの位置が示される。
【図8】リプレッサーoriSELECTにおいて用いるための宿主細胞のゲノムのさらなる例。この宿主細胞のゲノムは、リプレッサー遺伝子のシストロン(lacI)に連結されたRNAIIコード領域を含み、これらの両方が、E.coliのtrcプロモーターにおいて見出される−10配列および−35配列ならびにスペーシングに基づく構成的プロモーターの制御下にある。このゲノムはさらに、誘導性プロモーターの制御下にある必須遺伝子を含む(示さず)。プラスミドが存在しない場合において、このリプレッサータンパク質が発現され、必須遺伝子を抑制し、細胞増殖を阻害する。プラスミドによって生成されるRNAIの、RNAII−リプレッサーmRNA融合体のRNAII転写産物への結合は、リプレッサータンパク質の翻訳を遮断し、必須遺伝子の発現および細胞増殖を可能にする。
【特許請求の範囲】
【請求項1】
形質転換された宿主細胞であって、
i)細胞増殖を阻害する染色体遺伝子;および
ii)アンチセンス配列をコードするプラスミド、
を含み、該プラスミドによってコードされるアンチセンス配列は、該染色体遺伝子の作用を阻害し、それによって細胞増殖を許容する、形質転換された宿主細胞。
【請求項2】
前記プラスミドは目的の遺伝子の挿入のためのクローニング部位を含む、請求項1に記載の形質転換された宿主細胞。
【請求項3】
前記プラスミドはさらに目的の遺伝子を含む、請求項1もしくは請求項2に記載の形質転換された宿主細胞。
【請求項4】
前記プラスミドによってコードされるアンチセンス配列は、前記染色体遺伝子に結合することによって該染色体遺伝子の作用を阻害する、請求項1〜3のいずれか1項に記載の形質転換された宿主細胞。
【請求項5】
前記プラスミドによってコードされるアンチセンス配列は、前記染色体遺伝子から転写されるmRNAに結合することによって該染色体遺伝子の作用を阻害する、請求項1〜3のいずれか1項に記載の形質転換された宿主細胞。
【請求項6】
調節配列が前記染色体遺伝子に作動的に連結される、請求項1〜5のいずれか1項に記載の形質転換された宿主細胞。
【請求項7】
前記プラスミドによってコードされるアンチセンス配列は、前記調節配列に結合することによって前記染色体遺伝子の作用を阻害する、請求項6に記載の形質転換された宿主細胞。
【請求項8】
前記プラスミドによってコードされるアンチセンス配列は、前記調節配列から転写されるmRNAに結合することによって前記染色体遺伝子の作用を阻害する、請求項6に記載の形質転換された宿主細胞。
【請求項9】
前記プラスミドによってコードされるアンチセンス配列は、該プラスミドの複製起点によってコードされる、請求項8に記載の形質転換された宿主細胞。
【請求項10】
前記プラスミドによってコードされるアンチセンス配列は、RNAIもしくはRNAIの一部であり、前記染色体遺伝子に作動的に連結される調節配列は、RNAIIもしくはRNAIIの一部をコードする、請求項9に記載の形質転換された宿主細胞。
【請求項11】
前記プラスミドによってコードされるアンチセンス配列は、RNAIIもしくはRNAIIの一部であり、前記染色体遺伝子に作動的に連結される調節配列は、RNAIもしくはRNAIの一部である、請求項10に記載の形質転換された宿主細胞。
【請求項12】
細胞増殖を阻害する染色体遺伝子を含む宿主細胞であって、該染色体遺伝子は、該染色体遺伝子の上流に位置する調節配列に作動的に連結されており、該調節配列は、RNAI遺伝子もしくはRNAI遺伝子の一部、またはRNAII遺伝子もしくはRNAII遺伝子の一部をコードする、宿主細胞。
【請求項13】
前記細胞はインビトロで培養中である、請求項1〜11のいずれか1項に記載の形質転換された宿主細胞、もしくは請求項12に記載の宿主細胞。
【請求項14】
原核細胞である、請求項1〜11のいずれか1項に記載の形質転換された宿主細胞、もしくは請求項12および請求項13に記載の宿主細胞。
【請求項15】
細菌細胞である、請求項14に記載の形質転換された宿主細胞もしくは宿主細胞。
【請求項16】
前記細胞はグラム陰性細菌細胞である、請求項15に記載の形質転換された宿主細胞もしくは宿主細胞。
【請求項17】
前記細胞はE.coli細胞もしくはSalmonella細胞である、請求項16に記載の形質転換された宿主細胞もしくは宿主細胞。
【請求項18】
前記細胞はグラム陽性細菌細胞である、請求項15に記載の形質転換された宿主細胞もしくは宿主細胞。
【請求項19】
前記細胞はBacillus細胞である、請求項18に記載の形質転換された宿主細胞もしくは宿主細胞。
【請求項20】
弱毒化された細胞である、請求項15〜19のいずれか1項に記載の形質転換された宿主細胞もしくは宿主細胞。
【請求項21】
前記細胞は真核細胞である、請求項1〜13のいずれか1項に記載の形質転換された宿主細胞もしくは宿主細胞。
【請求項22】
前記細胞は酵母のような真菌である、請求項21に記載の形質転換された宿主細胞。
【請求項23】
前記細胞は植物細胞である、請求項21に記載の形質転換された宿主細胞。
【請求項24】
前記細胞は、哺乳類細胞もしくは昆虫細胞のような動物細胞である、請求項21に記載の形質転換された宿主細胞。
【請求項25】
前記染色体遺伝子はトキシン遺伝子である、請求項1〜24のいずれか1項に記載の形質転換された宿主細胞もしくは宿主細胞。
【請求項26】
前記トキシン遺伝子はsacBである、請求項25に記載の形質転換された宿主細胞もしくは宿主細胞。
【請求項27】
前記染色体遺伝子は、細胞増殖のために必須な第二の染色体遺伝子の発現を阻害するリプレッサータンパク質をコードする、請求項1〜24のいずれか1項に記載の形質転換された宿主細胞もしくは宿主細胞。
【請求項28】
前記第二の染色体遺伝子は、細胞増殖のために条件的に必須である、請求項27に記載の形質転換された宿主細胞もしくは宿主細胞。
【請求項29】
前記染色体遺伝子はリプレッサーlacIをコードし、前記第二の染色体遺伝子はlacIのオペレーターおよびプロモーターに作動的に連結される、請求項27もしくは請求項28に記載の形質転換された宿主細胞もしくは宿主細胞。
【請求項30】
前記染色体遺伝子はdapDもしくはfabAである、請求項27もしくは請求項28に記載の形質転換された宿主細胞もしくは宿主細胞。
【請求項31】
前記染色体遺伝子は、細胞増殖のために必須な第二の染色体遺伝子の発現を阻害するアンチセンス配列をコードする、請求項1〜24のいずれか1項に記載の形質転換された宿主細胞もしくは宿主細胞。
【請求項32】
前記染色体遺伝子によってコードされるアンチセンス配列は、前記第二の染色体遺伝子に結合することによって該第二の染色体遺伝子の発現を阻害する、請求項31に記載の形質転換された宿主細胞もしくは宿主細胞。
【請求項33】
前記染色体遺伝子によってコードされるアンチセンス配列は、前記第二の染色体遺伝子から転写されるmRNAに結合することによって該第二の染色体遺伝子の発現を阻害する、請求項31に記載の形質転換された宿主細胞もしくは宿主細胞。
【請求項34】
前記第二の染色体遺伝子は、細胞増殖のために条件的に必須である、請求項31〜33のいずれか1項に記載の形質転換された宿主細胞もしくは宿主細胞。
【請求項35】
前記第二の染色体遺伝子は、dapDもしくはfabAである、請求項31〜33のいずれか1項に記載の形質転換された宿主細胞もしくは宿主細胞。
【請求項36】
前記染色体遺伝子もしくは前記調節配列−染色体遺伝子融合体は、構成的プロモーターの制御下にある、請求項1〜35のいずれか1項に記載の形質転換された宿主細胞もしくは宿主細胞。
【請求項37】
前記染色体遺伝子、もしくは前記調節配列−染色体遺伝子融合体は、誘導性プロモーターの制御下にある、請求項1〜35のいずれか1項に記載の形質転換された宿主細胞もしくは宿主細胞。
【請求項38】
インビトロで宿主細胞内にプラスミドを維持する方法であって、該方法は、請求項1〜11のいずれか1項もしくは請求項13〜37のいずれか1項に記載の形質転換された宿主細胞を、該細胞が増殖することを許容するために十分な条件下で培養する工程を包含する、方法。
【請求項39】
プラスミドDNAを産生する方法であって、該方法は、形質転換された宿主細胞を請求項38に記載の方法に従って培養する工程、および該プラスミドDNAを単離する工程を包含する、方法。
【請求項40】
組換えタンパク質を産生する方法であって、該方法は、目的のタンパク質をコードするプラスミドを含む形質転換された宿主細胞を請求項38に記載の方法に従って培養する工程、および該タンパク質を該細胞から単離する工程を包含する、方法。
【請求項41】
薬学的組成物であって、該薬学的組成物は、請求項1〜37のいずれか1項に記載の形質転換された宿主細胞もしくは宿主細胞を、薬学的に受容可能な賦形剤、希釈剤、もしくは緩衝剤と共に含む、薬学的組成物。
【請求項42】
治療用途のための、請求項1〜37のいずれか1項に記載の形質転換された宿主細胞もしくは宿主細胞。
【請求項43】
遺伝子送達もしくはタンパク質送達のための薬の製造における、請求項1〜37のいずれか1項に記載の形質転換された宿主細胞もしくは宿主細胞の使用。
【請求項44】
目的の遺伝子を患者に送達する方法であって、該方法は、請求項3〜11もしくは請求項13〜37のいずれか1項に記載の形質転換された宿主細胞を該患者に投与する工程を包含する、方法。
【請求項45】
レシピエント生物体内にプラスミドを維持する方法であって、該方法は、請求項1〜11もしくは請求項13〜37のいずれか1項に記載の形質転換された宿主細胞を該生物体内へ導入する工程を包含し、該形質転換された宿主細胞内の前記染色体遺伝子は、インビボでの細胞増殖のために必須である、方法。
【請求項1】
形質転換された宿主細胞であって、
i)細胞増殖を阻害する染色体遺伝子;および
ii)アンチセンス配列をコードするプラスミド、
を含み、該プラスミドによってコードされるアンチセンス配列は、該染色体遺伝子の作用を阻害し、それによって細胞増殖を許容する、形質転換された宿主細胞。
【請求項2】
前記プラスミドは目的の遺伝子の挿入のためのクローニング部位を含む、請求項1に記載の形質転換された宿主細胞。
【請求項3】
前記プラスミドはさらに目的の遺伝子を含む、請求項1もしくは請求項2に記載の形質転換された宿主細胞。
【請求項4】
前記プラスミドによってコードされるアンチセンス配列は、前記染色体遺伝子に結合することによって該染色体遺伝子の作用を阻害する、請求項1〜3のいずれか1項に記載の形質転換された宿主細胞。
【請求項5】
前記プラスミドによってコードされるアンチセンス配列は、前記染色体遺伝子から転写されるmRNAに結合することによって該染色体遺伝子の作用を阻害する、請求項1〜3のいずれか1項に記載の形質転換された宿主細胞。
【請求項6】
調節配列が前記染色体遺伝子に作動的に連結される、請求項1〜5のいずれか1項に記載の形質転換された宿主細胞。
【請求項7】
前記プラスミドによってコードされるアンチセンス配列は、前記調節配列に結合することによって前記染色体遺伝子の作用を阻害する、請求項6に記載の形質転換された宿主細胞。
【請求項8】
前記プラスミドによってコードされるアンチセンス配列は、前記調節配列から転写されるmRNAに結合することによって前記染色体遺伝子の作用を阻害する、請求項6に記載の形質転換された宿主細胞。
【請求項9】
前記プラスミドによってコードされるアンチセンス配列は、該プラスミドの複製起点によってコードされる、請求項8に記載の形質転換された宿主細胞。
【請求項10】
前記プラスミドによってコードされるアンチセンス配列は、RNAIもしくはRNAIの一部であり、前記染色体遺伝子に作動的に連結される調節配列は、RNAIIもしくはRNAIIの一部をコードする、請求項9に記載の形質転換された宿主細胞。
【請求項11】
前記プラスミドによってコードされるアンチセンス配列は、RNAIIもしくはRNAIIの一部であり、前記染色体遺伝子に作動的に連結される調節配列は、RNAIもしくはRNAIの一部である、請求項10に記載の形質転換された宿主細胞。
【請求項12】
細胞増殖を阻害する染色体遺伝子を含む宿主細胞であって、該染色体遺伝子は、該染色体遺伝子の上流に位置する調節配列に作動的に連結されており、該調節配列は、RNAI遺伝子もしくはRNAI遺伝子の一部、またはRNAII遺伝子もしくはRNAII遺伝子の一部をコードする、宿主細胞。
【請求項13】
前記細胞はインビトロで培養中である、請求項1〜11のいずれか1項に記載の形質転換された宿主細胞、もしくは請求項12に記載の宿主細胞。
【請求項14】
原核細胞である、請求項1〜11のいずれか1項に記載の形質転換された宿主細胞、もしくは請求項12および請求項13に記載の宿主細胞。
【請求項15】
細菌細胞である、請求項14に記載の形質転換された宿主細胞もしくは宿主細胞。
【請求項16】
前記細胞はグラム陰性細菌細胞である、請求項15に記載の形質転換された宿主細胞もしくは宿主細胞。
【請求項17】
前記細胞はE.coli細胞もしくはSalmonella細胞である、請求項16に記載の形質転換された宿主細胞もしくは宿主細胞。
【請求項18】
前記細胞はグラム陽性細菌細胞である、請求項15に記載の形質転換された宿主細胞もしくは宿主細胞。
【請求項19】
前記細胞はBacillus細胞である、請求項18に記載の形質転換された宿主細胞もしくは宿主細胞。
【請求項20】
弱毒化された細胞である、請求項15〜19のいずれか1項に記載の形質転換された宿主細胞もしくは宿主細胞。
【請求項21】
前記細胞は真核細胞である、請求項1〜13のいずれか1項に記載の形質転換された宿主細胞もしくは宿主細胞。
【請求項22】
前記細胞は酵母のような真菌である、請求項21に記載の形質転換された宿主細胞。
【請求項23】
前記細胞は植物細胞である、請求項21に記載の形質転換された宿主細胞。
【請求項24】
前記細胞は、哺乳類細胞もしくは昆虫細胞のような動物細胞である、請求項21に記載の形質転換された宿主細胞。
【請求項25】
前記染色体遺伝子はトキシン遺伝子である、請求項1〜24のいずれか1項に記載の形質転換された宿主細胞もしくは宿主細胞。
【請求項26】
前記トキシン遺伝子はsacBである、請求項25に記載の形質転換された宿主細胞もしくは宿主細胞。
【請求項27】
前記染色体遺伝子は、細胞増殖のために必須な第二の染色体遺伝子の発現を阻害するリプレッサータンパク質をコードする、請求項1〜24のいずれか1項に記載の形質転換された宿主細胞もしくは宿主細胞。
【請求項28】
前記第二の染色体遺伝子は、細胞増殖のために条件的に必須である、請求項27に記載の形質転換された宿主細胞もしくは宿主細胞。
【請求項29】
前記染色体遺伝子はリプレッサーlacIをコードし、前記第二の染色体遺伝子はlacIのオペレーターおよびプロモーターに作動的に連結される、請求項27もしくは請求項28に記載の形質転換された宿主細胞もしくは宿主細胞。
【請求項30】
前記染色体遺伝子はdapDもしくはfabAである、請求項27もしくは請求項28に記載の形質転換された宿主細胞もしくは宿主細胞。
【請求項31】
前記染色体遺伝子は、細胞増殖のために必須な第二の染色体遺伝子の発現を阻害するアンチセンス配列をコードする、請求項1〜24のいずれか1項に記載の形質転換された宿主細胞もしくは宿主細胞。
【請求項32】
前記染色体遺伝子によってコードされるアンチセンス配列は、前記第二の染色体遺伝子に結合することによって該第二の染色体遺伝子の発現を阻害する、請求項31に記載の形質転換された宿主細胞もしくは宿主細胞。
【請求項33】
前記染色体遺伝子によってコードされるアンチセンス配列は、前記第二の染色体遺伝子から転写されるmRNAに結合することによって該第二の染色体遺伝子の発現を阻害する、請求項31に記載の形質転換された宿主細胞もしくは宿主細胞。
【請求項34】
前記第二の染色体遺伝子は、細胞増殖のために条件的に必須である、請求項31〜33のいずれか1項に記載の形質転換された宿主細胞もしくは宿主細胞。
【請求項35】
前記第二の染色体遺伝子は、dapDもしくはfabAである、請求項31〜33のいずれか1項に記載の形質転換された宿主細胞もしくは宿主細胞。
【請求項36】
前記染色体遺伝子もしくは前記調節配列−染色体遺伝子融合体は、構成的プロモーターの制御下にある、請求項1〜35のいずれか1項に記載の形質転換された宿主細胞もしくは宿主細胞。
【請求項37】
前記染色体遺伝子、もしくは前記調節配列−染色体遺伝子融合体は、誘導性プロモーターの制御下にある、請求項1〜35のいずれか1項に記載の形質転換された宿主細胞もしくは宿主細胞。
【請求項38】
インビトロで宿主細胞内にプラスミドを維持する方法であって、該方法は、請求項1〜11のいずれか1項もしくは請求項13〜37のいずれか1項に記載の形質転換された宿主細胞を、該細胞が増殖することを許容するために十分な条件下で培養する工程を包含する、方法。
【請求項39】
プラスミドDNAを産生する方法であって、該方法は、形質転換された宿主細胞を請求項38に記載の方法に従って培養する工程、および該プラスミドDNAを単離する工程を包含する、方法。
【請求項40】
組換えタンパク質を産生する方法であって、該方法は、目的のタンパク質をコードするプラスミドを含む形質転換された宿主細胞を請求項38に記載の方法に従って培養する工程、および該タンパク質を該細胞から単離する工程を包含する、方法。
【請求項41】
薬学的組成物であって、該薬学的組成物は、請求項1〜37のいずれか1項に記載の形質転換された宿主細胞もしくは宿主細胞を、薬学的に受容可能な賦形剤、希釈剤、もしくは緩衝剤と共に含む、薬学的組成物。
【請求項42】
治療用途のための、請求項1〜37のいずれか1項に記載の形質転換された宿主細胞もしくは宿主細胞。
【請求項43】
遺伝子送達もしくはタンパク質送達のための薬の製造における、請求項1〜37のいずれか1項に記載の形質転換された宿主細胞もしくは宿主細胞の使用。
【請求項44】
目的の遺伝子を患者に送達する方法であって、該方法は、請求項3〜11もしくは請求項13〜37のいずれか1項に記載の形質転換された宿主細胞を該患者に投与する工程を包含する、方法。
【請求項45】
レシピエント生物体内にプラスミドを維持する方法であって、該方法は、請求項1〜11もしくは請求項13〜37のいずれか1項に記載の形質転換された宿主細胞を該生物体内へ導入する工程を包含し、該形質転換された宿主細胞内の前記染色体遺伝子は、インビボでの細胞増殖のために必須である、方法。
【図1】

【図2】

【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【公表番号】特表2007−511236(P2007−511236A)
【公表日】平成19年5月10日(2007.5.10)
【国際特許分類】
【出願番号】特願2006−540601(P2006−540601)
【出願日】平成16年11月22日(2004.11.22)
【国際出願番号】PCT/GB2004/004929
【国際公開番号】WO2005/052167
【国際公開日】平成17年6月9日(2005.6.9)
【出願人】(506170591)コブラ バイオロジックス リミテッド (2)
【Fターム(参考)】
【公表日】平成19年5月10日(2007.5.10)
【国際特許分類】
【出願日】平成16年11月22日(2004.11.22)
【国際出願番号】PCT/GB2004/004929
【国際公開番号】WO2005/052167
【国際公開日】平成17年6月9日(2005.6.9)
【出願人】(506170591)コブラ バイオロジックス リミテッド (2)
【Fターム(参考)】
[ Back to top ]