
Fターム[4B065AA79]の内容
微生物、その培養処理 (127,014) | 微生物の種類 (32,496) | 菌類 (5,543) | 酵母 (2,459) | サッカロミセス (688)
Fターム[4B065AA79]の下位に属するFターム
サッカロミセス セレビッシェ (513)
サッカロミセス カールスベンゲンシス (7)
Fターム[4B065AA79]に分類される特許
141 - 160 / 168
ハロゲナーゼを含有する核酸断片およびベクター、ならびに化合物のハロゲン化方法
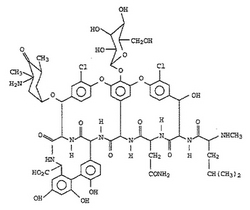
【課題】ハロゲナーゼの存在下で化合物をハロゲン化することを特徴とするハロゲン化法の提供。
【解決手段】ハロゲナーゼが、 (a) 特定の配列または遺伝暗号の縮重に基づいて該配列から誘導される配列にコードされるもの、または、 (b) (a)に記載されたものの機能的断片をコードする核酸配列にコードされるもの、または、 (c) (a)または(b)に記載された配列と標準的な条件下でハイブリダイズする配列にコードされるもの、または、 (d) (a)で特定される配列に対して30%を超える同一性もしくは60%を超える類似性を有する配列にコードされるもの、である、上記方法に関する。
(もっと読む)
セラミドの抽出方法
【課題】 真菌類(酵母、カビ、キノコ)の乾燥物からセラミドを抽出する際、乾燥物としては、製造コストの高い凍結乾燥品が使用されているが、その代替となる乾燥物を開発する。
【解決手段】 本発明によって、セラミドを抽出する対象物として、比表面積が7,000〜11,000cm2/g、平均粒径が400〜620μm、及び水分が2〜15wt%である真菌類の乾燥物が新たに開発された。この新規乾燥物は、攪拌流動層乾燥法等凍結乾燥法とは異なる乾燥方法によって製造することができ、得られた乾燥物は常法にしたがって有機溶媒抽出することにより効率的にセラミドを抽出することができる。
(もっと読む)
ファルネソール生産性酵母
【課題】新規なerg9-2変異遺伝子を有するファルネソール(FOH)生産性酵母を創製し、これを用いて通常の培養しやすい条件でFOHを効率よく大量生産でき、これによってFOHを含有する清酒や食品を工業的に製造できるようにすることである。
【解決手段】スクアレン合成酵素をコードするERG9遺伝子が、その塩基配列の714番目のgをaに置換されerg9-2遺伝子に変異したサッカロミセス・セレビシエ(Saccharomyces cerevisiae)であるFOH生産性酵母。FOH生産性酵母は、サッカロミセス・セレビシエ(Saccharomyces cerevisiae)HL296株[FERM AP−20132菌株]を使用できる。FOH生産性酵母と清酒酵母(協会7号系酵母)の一倍体株と交雑させ、胞子形成させた後、子嚢解剖を行い得られた一倍体株で高濃度のFOHを生産する酵母とすることもできる。
(もっと読む)
バイオリアクター
【課題】流動床型のバイオリアクターを提供することを課題とする。また、比較的簡易な構造で、連続発酵などの処理ができるバイオリアクター装置を提供することを課題とする。
【解決手段】管体の下方から上方に向かって水流を生じさせる水流発生手段が具備され、且つ前記管体の上方に、生体触媒によって処理された処理液が取り出される取出口が設けられており、該水流発生手段によって生体触媒が流動し被処理液を処理するバイオリアクターであって、前記管体の上方内部には、筒状の隔壁が管体の内周面に対して間隔を空けて設けられており、前記隔壁の下端部が取出口の下方に延出されていることを特徴とするバイオリアクター。
(もっと読む)
遺伝子およびポリペプチド配列
本発明は、(i)(a)分泌プレ配列、および(b)以下のモチーフ:−X1−X2−X3−X4−X5−(式中、X1はフェニルアラニン、トリプトファンまたはチロシンであり、X2はイソロイシン、ロイシン、バリン、アラニンまたはメチオニンであり、X3はロイシン、バリン、アラニンまたはメチオニンであり、X4はセリンまたはトレオニンであり、かつ、X5はイソロイシン、バリン、アラニンまたはメチオニンである)を含んでなるリーダー配列、ならびに(ii)そのリーダー配列に対して異種の目的タンパク質を含んでなるポリペプチドを提供する。本発明のポリペプチドは、リーダー配列の一部として、分泌プロ配列をさらに含んでなってよい。本発明はまた、本発明のポリペプチドをコードする配列を含んでなるポリヌクレオチドおよび該ポリヌクレオチドを含んでなる細胞、好ましくは、酵母細胞を提供する。  (もっと読む)
(もっと読む)
1,4−ブタンジアミンの生化学合成
本発明は、生来のレベルのオルニチンデカルボキシラーゼ活性と比較して増加したレベルのオルニチンデカルボキシラーゼ活性(増加したODC活性)を有する微生物における1,4−ブタンジアミンの生化学合成のためのプロセスであって、ここで、増加したODC活性は、増加した翻訳および/または転写効率によるオルニチンデカルボキシラーゼをコードする遺伝子の過剰発現によって得られ、そして微生物において産生される1,4−ブタンジアミンは、発酵ブロスに分泌され、発酵ブロスから回収される、プロセスに関する。好適な実施形態では、(i)アルギニンデカルボキシラーゼをコードする遺伝子speAおよびアグマチナーゼをコードする遺伝子speB;または(ii)アルギニンデカルボキシラーゼをコードする遺伝子speAおよびアグマチンイミノヒドロラーゼをコードする遺伝子aguA、およびN−カルバモイルプトレシンアミドヒドロラーゼをコードする遺伝子aguB、ならびに場合によりまた、アグマチナーゼをコードする遺伝子speBのいずれかの過剰発現によっても増加した酵素活性が得られる。本発明はまた、増加したレベルの活性で、1つもしくはそれ以上の上記の酵素活性を担持するベクター、プラスミドおよび宿主に関する。 (もっと読む)
細菌ロドバクタースフェロイデスから得られるチロシンアンモニアリアーゼ酵素のDNAおよびアミノ酸配列
細菌ロドバクタースフェロイデス(Rhodobacter sphaeroides)において、新規のチロシンアンモニアリアーゼ酵素が同定された。この酵素は、フェニルアラニンに対してよりもチロシンに対してより高い活性を有し、チロシンから直接パラ−ヒドロキシケイ皮酸を生成するために有用である。この酵素をコードする遺伝子が、ゲノムDNAを用いる直接的増幅によってクローン化され、大腸菌中で発現された。 (もっと読む)
ポリペプチド製造用シグナルペプチド
本発明は、シグナルペプチドを使用することを含んでなるポリペプチドを製造する方法、シグナルペプチドをコードする第1ヌクレオチド配列と第1ヌクレオチド配列に対して外来であるポリペプチドをコードする第2 ヌクレオチド配列とを含んでなる核酸構築物に関する。さらに、本発明は、また、前記核酸構築物を含んでなる発現ベクターおよび宿主細胞に関する。 (もっと読む)
キシロースで生育する非組換えサッカロマイセス株
本発明は、キシロースを唯一の炭素源として望ましい生育速度(例えば、少なくとも48時間あたり一世代)で生育し得るサッカロマイセス株の作製方法、そのような方法により作製される株、並びに非組換え手法によって作製される、生育のための唯一の炭素源としてキシロースを用いて少なくとも48時間あたり一世代の生育速度で生育するサッカロマイセス株に関する。 (もっと読む)
4−アミノ−4−デオキシコリスミ酸(ADC)および[3R,4R]−4−アミノ−3−ヒドロキシシクロヘキサ−1,5−ジエン−1−カルボン酸(3,4−CHA)の生合成生産
本発明は、高められたレベルの活性で、4−アミノ−4−デオキシコリスミ酸シンターゼ、好ましくはPabAB二部分タンパク質(それは融合タンパク質であってもよい)を使ってインビボで発酵により行われ、それによって、回収されるADCおよび4−アミノ−4−デオキシプレフェネート(ADP)を含む培養液を得る、4−アミノ−4−デオキシコリスミ酸(ADC)の生合成生産方法に関する。本発明はまた、p−アミノフェニルアラニンへのADPのさらなる転化方法にも関する。本発明はさらに、3,4−CHAの回収を含む、かかる4−アミノ−4−デオキシコリスミ酸シンターゼのおよびイソコリメートを[5S,6S]−5,6−ジヒドロキシシクロヘキサ−1,3−ジエン−1カルボン酸(2,3−CHD)へ転化することができる酵素、好ましくはフェナジン生合成タンパク質PhzDの共同作用による[3R,4R]−4−アミノ−3−ヒドロキシシクロヘキサ−1,5−ジエン−1−カルボン酸(3,4−CHA)の生合成生産に関する。本発明はまた、かかる方法の任意のものでの使用のための発現ベクターおよびホスト細胞にも関する。本発明はさらに、触媒活性生成物としての、特にキラル触媒としての3,4−CHAの使用に関する。そして本発明は最後に3,4−CHAからのリン酸オセルタミビルの合成に関する。 (もっと読む)
糖タンパク質の作製においてアルファ−マンノシダーゼ抵抗性グリカンを減少させるか又は排除する方法
本発明は、酵母における糖タンパク質上のa−マンノシダーゼ抵抗性グリカンを減少させるか又は排除する方法を提供する。糖タンパク質上のa−マンノシダーゼ抵抗性グリカンの減少又は排除は、新たに単離されたβ1,2−マンノシルトランスフェラーゼをコードするP.パストリスAMR2遺伝子の破壊に起因する。本発明は、グリカンにおけるa−マンノシダーゼ抵抗性に関する新規の遺伝子、ポリペプチド、抗体、ベクター、及び宿主細胞も開示する。  (もっと読む)
(もっと読む)
新規アルカリプロテアーゼおよび、該新規アルカリプロテアーゼを含有する洗浄剤および清浄剤
本発明は、互いに類似しており、DNAが土壌サンプルから得られる2つの新規アルカリプロテアーゼ(配列番号:4および7)に関し、C末端が削除され同様のタンパク質分解活性を示すそのフラグメント(配列番号:5および8)、配列番号:4と少なくとも90%までまたは配列番号:7と少なくとも87.5%まで類似している全てのアルカリプロテアーゼ、および配列番号:4および配列番号:7から得られるコンセンサス配列(配列番号9)としてまとめられ得るものに関する。さらに、関連核酸(配列番号:3および6)と少なくとも85%同一の相同性を持つ全ての核酸または該関連フラグメントに関する。さらに、これらのプロテアーゼに対する使用の技術的可能性を定義し、特に洗浄剤および清浄剤におけるそれらの使用を記載する。 (もっと読む)
下等真核生物におけるガラクトシル化された糖タンパク質の産生
本発明は、末端β−ガラクトース残基を有すること及びフコース残基及びシアル酸残基を本質的に欠失していることを特徴とするヒト様糖タンパク質を産生する新規下等真核宿主細胞を提供する。本発明は、治療用糖タンパク質として使用できる、組換え型下等真核宿主細胞における受容体基質への、UDP−ガラクトースからのガラクトース残基の転移を触媒するための方法も提供する。 (もっと読む)
酵母におけるHPV52L1の最適化された発現
HPV 52 L1タンパク質をコードする合成DNA分子が提供される。具体的には、本発明はHPV 52 L1タンパク質をコードするポリヌクレオチドを提供し、このポリヌクレオチドは酵母細胞内において高レベル発現をするようコドン最適化されている。本発明のもう1つの実施形態では、合成分子のヌクレオチド配列は酵母によって認識される転写終結シグナルを除去するよう改変されている。合成分子はHPV 52ウイルス様粒子(VLPs)を産生するため、およびHPV 52 VLPsを含むワクチンおよび医薬組成物を生産するために使用され得る。本発明のワクチンは、中和抗体および細胞媒介性免疫反応を通してパピローマウイルス感染に対する有効な免疫学的予防を提供し、また既存のHPV感染の治療にも有効であり得る。  (もっと読む)
(もっと読む)
真菌および酵母におけるシチジンモノホスフェート−シアル酸合成経路を操作する方法
本発明は、内因性CMP−シアル酸を欠く非ヒト宿主において、CMP−シアル酸を生成するための方法を提供する。この方法は、この宿主に、細菌、哺乳動物またはハイブリッドのCMP−シアル酸生合成経路に由来する、CMP−シアル酸合成に関与する酵素を提供することによる。シアル化糖タンパク質の生成のためにCMP−シアル酸生合成経路を発現する新規な真菌宿主も提供される。この経路は、内因性CMP−シアル酸を欠く非ヒト宿主細胞における糖タンパク質のシアル化に特に有用である。  (もっと読む)
(もっと読む)
個別のペレットからなる凍結乳酸菌培養物
少なくとも50gの重量の凍結物質を有する商業用パッケージ中の凍結乳酸菌(LAB)培養物であって、ここで当該凍結培養物が、個別のペレットの形態で存在し、-46℃で7〜14日間貯蔵されたとき、凍結培養物の個別のペレットが互いにくっ付かず、その結果、実質的に個別の粒子として残るということにより特徴付けられる。 (もっと読む)
組み換えタンパク質生産用タンパク質融合因子の超高速選別方法及びこれによって選別されたタンパク質融合因子
本発明は、従来の組み換え生産方法で生産することが難しい難発現性タンパク質を発現及び分泌生産することが可能な適合型タンパク質融合因子(TFP)を多様な遺伝子源から超高速で選別する方法、及びこれから得られたタンパク質分泌誘導タンパク質融合因子を開示する。 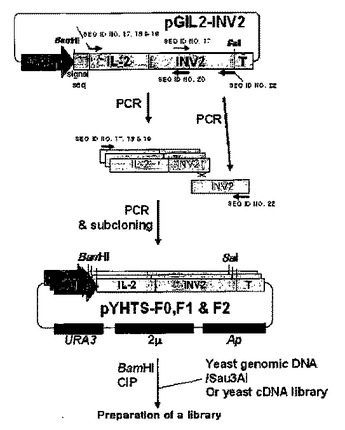 (もっと読む)
(もっと読む)
遺伝子発現法
本発明により、異種タンパク質を製造する方法であって、(a)シャペロンタンパク質の配列を含むタンパク質をコードしている遺伝子と、異種タンパク質をコードしている遺伝子とを含む2μm系プラスミドを含む宿主細胞を準備すること;(b)その宿主細胞を、培養培地中、シャペロンタンパク質をコードしている遺伝子と、異種タンパク質をコードしている遺伝子の発現を可能とする条件下で培養すること;および(c)発現した異種タンパク質を培養宿主細胞または培養培地から精製することを含んでなる方法が提供される。 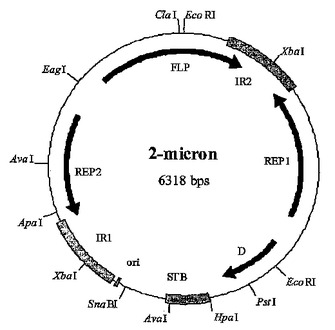 (もっと読む)
(もっと読む)
エンドグルカナーゼSTCEおよびそれを含むセルラーゼ調製物
スタフィロトリクム・ココスポラム(Staphylotrichum coccosporum)由来の新規エンドグルカナーゼ、前記エンドグルカナーゼをコードするポリヌクレオチド、及び前記エンドグルカナーゼを含むセルラーゼ調製物を開示する。
前記エンドグルカナーゼ又はセルラーゼ調製物は、セルロース含有繊維の色の澄明化、毛羽立ちの低減、肌触り及び外観の改善、色の局所的変化、ごわつきの低減などを目的とした洗剤用、並びに繊維加工用途に有用である。
(もっと読む)
改良されたグルコースデヒドロゲナーゼポリペプチドおよび関連ポリヌクレオチド
本発明は、中心的野生型グルコースデヒドロゲナーゼポリペプチドと比較して、増強されたGDH活性および/または熱安定性を有する、グルコースデヒドロゲナーゼ(GDH)に関する。さらに、本発明は、本発明のGDHポリペプチドをコードするポリヌクレオチド、上記ポリヌクレオチドを含有する核酸配列、プロモーターに作動可能に連結したポリヌクレオチドを包含する発現ベクター、GDHポリペプチドを発現するように形質転換された宿主細胞および本発明のGDHポリペプチドを産生する方法。 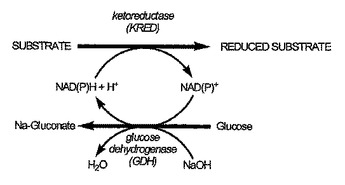 (もっと読む)
(もっと読む)
141 - 160 / 168
[ Back to top ]