
ウイルスの病原性の阻害方法
本発明は、ウイルスerb-Bリガンドによる細胞の活性化を阻害する方法、およびゲノムにウイルスerb-Bリガンドをコードする核酸配列を含むウイルスに感染しているか、感染すると考えられるか、または感染するリスクがある動物(たとえば、ヒト被験者)の免疫応答を増強する方法を特徴とする。
【発明の詳細な説明】
【技術分野】
【0001】
技術分野
本発明は、ウイルス感染の制御に、より詳細にはポックスウイルス(たとえば、天然痘)感染の制御に関する。
【背景技術】
【0002】
背景
B型肝炎などのウイルス病によって生じるヒトの経済的、および社会的な荒廃並びに天然痘などの疾患のバイオテロリストの利用に対する恐るべき可能性の点からみて、このような感染症に対する有効な治療薬および/または予防薬が開発されることが必須である。天然痘は、20年以上前に人類から根絶されたと考えられていたので、原因となるウイルス(痘瘡)のワクチン接種は、その時に廃止した。したがって、現在のヒト集団の比較的多くの割合が、これまでに疾患に対するワクチン接種がなされていない。さらに、ワクチン接種の廃止前に生産された限られた天然痘ワクチンの供給だけが現在利用可能である。
【発明の開示】
【0003】
概要
本発明者らは、erb-Bタンパク質チロシンキナーゼを阻害する化合物が、ポックスウイルス感染症を阻害するのに有効であることを発見し、したがって、これらの化合物は、有効な抗ウイルス治療薬および予防薬である可能性が高い。
【0004】
実施例2は、哺乳類タンパク質エピレギュリン(エピレギュリン:EPI)に対して、特にこれらの上皮性因子(EGF)様のドメイン内において、比較的高い相同性を有するポックスウイルス・タンパク質を記載してある。これらのポックスウイルス・タンパク質は、本明細書においてエピレギュリン様成長因子(ELGF)と称される。天然痘(痘瘡)ウイルスELGFは、指定を受けた天然痘成長因子(SPGF)であり、ワクシニアELGFは、指定を受けたワクシニア成長因子(VGF)である。
【0005】
本発明は、一つには、細胞表面erb-B1受容体に対するD4R(大痘瘡のバングラデシュ株に由来するSPGF)の組換え上皮細胞成長因子(EGF)様ドメインの結合が、(a)これらを発現する細胞におけるこのような受容体のチロシンキナーゼ活性の活性化;(b)このような細胞の増殖および/または生存の増強;(c)erb-B1受容体の迅速なインターナリゼーション;並びに(d)複製および付随する炎症の増強によって生じるウイルス病原性を生じるという観察に基づいている。これらの知見は、SPGFが(たとえば、痘瘡ビリオンの表面で、または痘瘡ビリオンから分断されて)、痘瘡ウイルスの宿主細胞への侵入、およびその中での複製、およびその後の炎症において中心的役割を果たすことを示す。
【0006】
加えて、本発明者らは、erb-B1受容体を経た細胞に対するSPGFの結合によって刺激されるタンパク質チロシンキナーゼ活性およびSPGFによるライゲーション後のerb-B1受容体のインターナリゼーションが、erb-Bタンパク質チロシンキナーゼ阻害剤によって阻害されることを見いだした。このerb-B1インターナリゼーションの阻害は、明らかにc-Cblとerb-B1の間の誘導性の結合の阻害によるものであった。これらの知見と一致して、本発明者らも、キナゾリン・タンパク質チロシンキナーゼ阻害剤CI-1033が、ワクシニアウイルスに感染した動物における臨床症状を阻害し、ウイルス負荷を減少したことを観察した。このCI-1033の処置効果は、中和ワクシニア特異的モノクローナル抗体(mAb)(抗L1R mAb)の同時投与によって増強された。加えて、本発明者らは、erb-Bチロシンキナーゼ阻害剤が1つの細胞から別の細胞への痘瘡ウイルスの伝播を減少することを見いだした。
【0007】
面白いことに、CI-1033は、また、感染動物におけるワクシニアウイルスに対するT細胞性免疫を増強したこと、およびこの増強は、感染動物に対して抗L1R mAbを同時投与することによってさらに増大することが判明した。さらに、本発明者らは、CI-1033と同様に、VGF(SPGFのワクシニア相同分子種)に結合するmAbがワクシニアに感染した動物におけるワクシニアウイルスに対するT細胞免疫を増強したこと、およびこの効果が、動物に対する抗L1R mAbの投与によってさらに増大されたことを見いだした。
【0008】
これらの知見は、erb-Bチロシン・タンパク質キナーゼ活性の阻害剤、またはタンパク質キナーゼ活性の活性化の阻害剤が、動物(たとえば、ヒト被験者)のポックスウイルス感染症に対する有効な薬剤であり得ることを示す。これらの阻害剤は、少なくとも、細胞におけるウイルス複製を阻害すること、および細胞からウイルスが出ることによって、並びにウイルスに対する免疫の増強によって作用すると思われる。また、阻害剤は、細胞へのウイルス侵入を減少させるかもしれない。
【0009】
より詳細には、本発明は:(a)ウイルスerb-Bリガンドをコードする遺伝子を含むウイルスに曝露された可能性があるか、または暴露されている可能性がある動物を同定する工程;および、(b)erb-Bチロシンキナーゼの活性を阻害するか、またはerb-Bチロシンキナーゼの活性化を阻害する化合物で動物を処置する工程、を含む方法を特徴とする。動物は、工程(b)の前に、間に、または後に、ウイルスに感染していたと同定される。ウイルスerb-Bリガンドは、オルトポックスウイルス(たとえば、大痘瘡、小痘瘡、サル痘ウイルス、またはワクシニア)erb-Bリガンドなどのポックスウイルスerb-Bリガンドであることができる。erb-Bリガンドは、天然痘成長因子(SPGF)またはワクシニア成長因子(VGF)などのエピレギュリン様の成長因子(ELGF)であることができ、erb-Bチロシンキナーゼは、erb-B1、erb-B2、またはerb-B4チロシンキナーゼであることができる。
【0010】
化合物は、たとえばerb-Bチロシンキナーゼと結合する非アゴニスト抗体、または非アゴニストerb-Bリガンド、またはerb-Bリガンドの非アゴニスト断片であることができる。または、化合物は、たとえば、キナゾリンに基づいた化合物(たとえば、4-アニリノ・キナゾリン)、ピリドピリミジンに基づいた化合物、キノリン-3-カルボニトリルに基づいた化合物、またはピロロピリミジンに基づいた化合物などの小分子erb-Bチロシンキナーゼ阻害剤であることができる。小分子erb-Bチロシンキナーゼ阻害剤は、erb-Bチロシンキナーゼ活性の不可逆的または可逆的な阻害剤であることができる。キナゾリンに基づいた化合物は、たとえば、CI-1033、PD168393、PD160678、PD160879、PD174265、PD153035、ZD1839、GW572016、GW974、OSI-774、またはAG1478であることができる。ピリドピリミジンに基づく化合物は、たとえばPD69896、PD153717、またはPD158780であることができる。キノリン-3-カルボニトリルに基づいた化合物は、たとえばEKB-569であることができ、およびピロロピリミジンに基づいた化合物は、たとえばCGP59326Aであることができる。
【0011】
本方法では、動物は、ヒトであることができる。本方法は、ウイルスの1つもしくは複数の形態を実質的に中和する能力を有する抗体、たとえばオルトポックスウイルス(たとえば、大痘瘡、小痘瘡、ワクシニア、またはサル痘ウイルス)の細胞内成熟ウイルス粒子(IMV)形態、細胞外外膜ウイルス(EEV)形態、および/または細胞結合エンベロープ・ウイルス(CEV)形態と結合する抗体を動物に投与することをさらに含むことができる。
【0012】
本方法において、化合物での処置は、動物におけるウイルスに対する免疫応答を増強することができる。免疫応答は、T細胞応答、たとえばCD8+ T細胞応答またはCD4+ T細胞応答であることができ、T細胞応答は、インターフェロン-γ(IFN-γ)産生T細胞応答であることができる。または、免疫応答は、抗体産生B細胞応答であることができる。
【0013】
本方法の一つの態様において、ウイルスは、ポックスウイルス発現ベクターであることができ、かつ(a)免疫原をコードする異種核酸配列;および、(b)異種核酸配列に動作可能に連結される、転写調節性のエレメント(TRE)をさらに含むことができる。ポックスウイルス発現ベクターは、ワクシニアウイルス発現ベクター、弱毒ワクシニアウイルス発現ベクター、カナリア痘瘡ウイルスベクター、または鶏痘ウイルスベクターであることができる。
【0014】
また、本発明は:(a)erb-Bリガンドをコードする遺伝子を含むウイルスによる感染に感受性の動物を同定する工程;並びに、(b)動物を、(i)ウイルスに対する免疫応答を刺激するウイルスまたはワクチンの1つもしくは複数の形態を実質的に中和する抗体、および(ii)erb-Bチロシンキナーゼを阻害するか、またはerb-Bチロシンキナーゼの活性化を阻害する化合物で処置することと、を含む方法を特徴とする。ウイルス、動物、erb-Bリガンド、化合物、およびウイルスの1つまたは複数の形態を実質的に中和する抗体は、上記したもののいずれであることもできる。そのうえ、本方法は、上記で詳述した免疫応答のいずれを増強することもでき、ウイルスは、上記ポックスウイルス発現ベクターのいずれであることもできる。
【0015】
本発明のもう一つの態様は:(a)erb-Bチロシンキナーゼと結合し、かつerb-Bチロシンキナーゼの活性または受容体のチロシンキナーゼの活性化を阻害する単離された化合物を提供する工程;(b)erb-Bチロシンキナーゼを発現する細胞と化合物を接触させる工程;および、(c)工程(b)の前に、と同時に、もしくは後に、ウイルスerb-Bリガンドまたはリガンドの機能的な断片と細胞を接触させる工程を含むインビトロ方法である。化合物は、リガンドまたは断片による細胞上のerb-Bチロシンキナーゼの活性化を減少させることができる。または、本方法は、化合物が、リガンドまたは断片による細胞上のerb-Bチロシンキナーゼの活性化を減少させるかどうか決定することをさらに含むことができる。ウイルスerb-Bリガンド、erb-Bチロシンキナーゼ、および化合物は、上記したもののいずれであることもできる。
【0016】
また、本発明は、化合物が抗ウイルス性の化合物であるかどうかを決定する方法であって:(a)erb-Bチロシンキナーゼの活性を阻害するか、またはerb-Bチロシンキナーゼの活性化を阻害する化合物を提供する工程;(b)erb-Bリガンドをコードする遺伝子を含むウイルスに対する感染に感受性の動物に対して化合物を投与する工程;(c)工程(b)の前に、の間に、または後に、ウイルスに対して動物を曝露する工程;および、(d)化合物が動物のウイルス感染の症状を減少させるかどうか決定する工程を含む方法を提供する。ウイルスは、ポックスウイルス、たとえば大痘瘡、小痘瘡、サル痘ウイルス、またはワクシニアなどのオルソポックスウイルスであることができる。動物、erb-Bリガンド、および化合物は、上記したもののいずれであることもできる。
【0017】
さらに本発明による特徴とされるもう1つの方法は:(a)erb-Bリガンドをコードする遺伝子を含むウイルスによる感染に感受性の動物を同定する工程;並びに、(b)動物を、(i)リガンドに対する抗体、および(ii)ウイルスに対する免疫応答を刺激するウイルスまたはワクチンの1つもしくは複数の形態を実質的に中和する抗体で処置する工程を含む。動物は、工程(b)の前に、の間に、または後に、ウイルスに感染していたと同定することができる。処置によりウイルスに対する免疫応答を増強することができ、免疫応答は、上記で列記したもののいずれであることもできる。ウイルスは、ポックスウイルス発現ベクターであることができ、かつ(a)免疫原をコードする異種核酸配列;および、(b)異種核酸配列に動作可能に連結される、転写調節性のエレメント(TRE)をさらに含むことができる。ポックスウイルス発現ベクターは、ワクシニアウイルス発現ベクター、弱毒ワクシニアウイルス発現ベクター、カナリア痘瘡ウイルスベクター、または鶏痘ウイルスベクターであることができる。リガンドと結合する抗体は、13E8モノクローナル抗体(ATCCアクセッション番号PTA-5040)または11D7モノクローナル抗体(ATCCアクセッション番号PTA-5039)であることができる。ウイルス、動物、erb-Bリガンド、化合物、およびウイルスの1つまたは複数の形態を実質的に中和する抗体は、上記したもののいずれであることもできる。
【0018】
「ポリペプチド」および「タンパク質」は、交換可能に使用され、長さまたは翻訳後の修飾に関係なく、何らかのペプチド結合されたアミノ酸鎖を意味する。本発明に使用されるポリペプチドは、保存的置換を有するものを含む。保存的置換は、典型的には以下の群内の置換を含む:グリシンおよびアラニン;バリン、イソロイシン、およびロイシン;アスパラギン酸およびグルタミン酸;アスパラギン、グルタミン、セリン、およびスレオニン;リジン、ヒスチジン、およびアルギニン;並びにフェニルアラニンおよびチロシン。一般に、保存的置換を有する変種ポリペプチドは、40未満(たとえば、35;30;25;20;15;13;11;10;9;8;7;6;5;4;3;2;または1つ未満)の保存的置換を含む。変種ポリペプチドは、野生型ポリペプチドの活性の少なくとも20%(たとえば、少なくとも:30%;40%;50%;60%;70%;80%;90% ;95%;98%;99%;99.5%;99.8%;99.9%;または100%以上)を有するということが、必要とされる全てである。
【0019】
本明細書に使用されるものとして「細胞の活性化」は、細胞増殖の増強または細胞の生存の増強をトリガーすることを意味する。「細胞増殖の増強または細胞の生存の増強」は、(a)低レベルの細胞増殖または細胞生存;または(b)実質的に細胞増殖または細胞生存がないこと、と比較してして増大されることを意味する。
【0020】
本明細書に使用されるものとして「免疫応答を増強する」ことは、(a)より低い免疫応答;または(b)実質的に免疫反応がないこと、と比較してして免疫応答を増大することを意味する。
【0021】
「単離された化合物」の用語は、本明細書に使用されるものとして、天然に存在する対応物を有さないか、または天然にこれを伴う成分、たとえば膵臓、肝臓、脾臓、卵巣、精巣、筋肉、関節組織、神経組織、胃腸組織、もしくは腫瘍組織などの組織、または血液、血清、もしくは尿などの体液中の成分から分離されず、または精製されていない化合物(たとえば、タンパク質)をいう。典型的には、天然に存在する生物学的化合物は、これが、乾燥重量で、天然に付随しているタンパク質およびその他の天然に存在する有機分子が少なくとも70%であるときに、「単離された」とみなされる。好ましくは、本発明に使用される化合物の標品は、その化合物の乾燥重量で、少なくとも80%、より好ましくは、少なくとも90%、および最も好ましくは少なくとも99%である。単離または純度の程度は、任意の適切な方法によって、たとえばカラムクロマトグラフィー、ポリアクリルアミドゲル電気泳動、またはHPLC(高速液体クロマトグラフィー)解析によって測定することができる。化学的に合成された化合物(たとえば、タンパク質)は、その性質上、天然にこれに付随する成分から分離されており、合成化合物は、定義上「単離され」ている。
【0022】
単離された化合物、および本発明のために有用なさらなる薬剤は、たとえば以下によって得ることができる:(i)天然の供与源から(たとえば、組織または体液から)の抽出;(ii)化合物がタンパク質である場合は、タンパク質をコードする組換え核酸の発現により;または(iii)当業者に公知である標準的な化学的合成法により。これが天然に由来する供与源とは異なる細胞の系で産生されるタンパク質は、これが天然にそれを伴う成分を必然的に含まないので、「単離され」ている。
【0023】
他に定義されない限り、本明細書に使用される全ての技術的および科学的な用語は、本発明が属する当該技術分野において当業者によって共通に理解されるものと同じ意味を有する。矛盾する場合には、定義を含む本明細書が対照となる。好ましい方法および材料は、以下に記載してあるが、本明細書に記載されているものと同じか、または均等な方法および材料も、本発明の実施または試験に使用することができる。全ての刊行物、特許出願、特許、および本明細書において述べられるその他の参照は、これらの全体が参照として組み入れられる。本明細書に開示された材料、方法、および実施例は、例示するだけであり、限定されることは企図されない。
【0024】
たとえば、細胞の活性化を阻害することを含む、本発明のその他の特徴および利点は、以下の明細書から、図面から、および特許請求の範囲から明らかであろう。
【0025】
詳細な説明
上記の概要の節において概説した実験の知見は、erb-B1タンパク質チロシンキナーゼ活性、またはerb-B1タンパク質チロシンキナーゼ活性の刺激のいずれかを阻害する化合物が、オルソポックスウイルスを媒介した病気の発生(たとえば、天然痘またはワクシニア感染症)の治療、および/またはこれに対する予防に有効であり得ることを示す。本発明は、いずれの特定の作用機構によっても限定されないが、このような化合物は、宿主細胞へのウイルス粒子の侵入、細胞内のウイルス複製、および/または細胞からのウイルス放出を阻害することによって、並びにウイルスに対する適応性T細胞免疫を増強することによって作用するのであろうと考えられる。オルトポックスウイルスのゲノムは、免疫抑制因子をコードする遺伝子を含むという点で、ウイルス複製の阻害および適応免疫の増強に機能的に大いに関連する可能性がある。たとえば、B型肝炎ウイルス、EBウイルス、RNA型腫瘍ウイルス(たとえば、ニワトリ赤芽球症ウイルス)、サイトメガロウイルス(CMV)、およびレオウイルスによる細胞の感染におけるerbB受容体の役割を示す以前の知見は(実施例7の参照を参照されたい)、本観察を考慮して、erb-Bタンパク質チロシンキナーゼの阻害剤が複数のポックスウイルスだけでなく、多種多様なその他のウイルスに対しても有効な治療薬および/または予防薬である可能性が高いことを示唆する。
【0026】
天然痘(大痘瘡および小痘瘡)および牛痘(ワクシニア)ウイルスでは、特に興味がもたれる。これらのウイルスは、感染細胞の細胞質で複製する大きなニ本鎖DNAウイルスである。痘瘡ウイルスは、気道を介して被検者に入り;ヒト-ヒトの伝達は、通常経口咽頭分泌物中のウイルスの咳嗽の結果として生じる。7〜19日のインキュベーション期間に続いて、熱、頭痛、および背中の痛みがある。2〜3日後に、熱が下がり、顔、躯幹、および四肢に発疹が現れ、小胞、膿疱、および痂皮に進行して数週間の間続く。大痘瘡感染症は、ワクチン非接種人間の約40%では致命的である。死は、主に内出血(播種性血管内凝固)および血管虚脱によるものである。小痘瘡に感染すると、致死率の減少したあまり重症型ではない天然痘を生じる。
【0027】
1980年頃の天然痘の国際根絶により、ワクチン接種は廃止した。従って、天然痘の防護は、米国において約20年間提供されてこなかった。その結果、かなりの個体群が、天然痘に対してこれまでワクチン接種がなされなかった。さらに、米国には、現在、天然痘ワクチン(弱毒化ワクシニアウイルス)の適当な蓄えがなく、利用できるものは、免疫不全(たとえば、AIDSおよび癌)患者に有害な影響を及ぼす能力がある生きたワクシニアウイルスを産した十分に制御されていないプロセスによって産生されるものである。加えて、10〜20の第二世代の場合は、一人のヒトの痘瘡感染から生じる可能性がある。これらの要因により、痘瘡ウイルス、特に大痘瘡ウイルスは、米国人口に対する生物兵器としての明白な選択肢となる。したがって、ウイルスに対する効率的な、安全な治療的な、および/または予防的な方法論ができる限り速やかに開発されることが不可避である。
【0028】
痘瘡は、最も大ききく、かつ最も複雑なニ本鎖DNAウイルスの1つであり、光学顕微鏡によって見える。配列データは、大痘瘡ウイルスおよび小痘瘡ウイルスについて入手可能である。GenBankの痘瘡の現在の配列データには、以下を含む:(a)大痘瘡の5つのヌクレオチド配列の登録および626アミノ酸配列の登録;並びに(b)小痘瘡の3つのヌクレオチド配列の登録および619アミノ酸配列の登録。2つの完全な大痘瘡ゲノム・ヌクレオチド配列があり、インド株1967(GI:9627521;185,578 bp)およびバングラデシュ株(GI:623595;186,103 bp)に対応しており、小痘瘡株アラストリムの1つの完全なゲノム・ヌクレオチド配列(GI:5830555;186,986 bp)がGenBankで入手可能である。大痘瘡および小痘瘡ゲノムの比較では、これらが97%以上、全ゲノムの全体にわたって同一であることを示す。同様に、いずれの痘瘡ゲノムのワクシニアのものとの比較でも(GI:9790357;191,737 bp)、これらが〜95%同一であることを示す。
【0029】
大痘瘡インド株1967のゲノムには、197個の遺伝子がある(GI:9627521)。これらの機能およびウイルスのライフサイクルの関連については、ほとんど知られていない。痘瘡ウイルス(大痘瘡インド株)のゲノムのコンピュータに基づいた解析からのデータでは、天然痘に対するワクチンとして、および有効な受動免疫保護剤と考えられる抗体(同時係属中のU.S. Application Serial No. 10/429,685を参照されたい(この開示はその全体が参照として本明細書に組み入れられる))を作製するために潜在的に有用であるゲノム内の遺伝子によってコードされる種々のポリペプチドを同定した。この解析によって同定されたポリペプチドのうちの1つは、D1Lであり、これは、上皮細胞成長因子(EGF)様膜結合型タンパク質であり、その他の痘瘡株の対応する分子に対して≧97%の相同性、対応するワクシニア・タンパク質と86〜89%の同一性、およびヒト・エピレギュリンに対して30%の同一性を有する。上記のように、これらのポックスウイルスのEGF様膜結合タンパク質は、本明細書において、ひとまとめにしてエピレギュリン様成長因子(ELGF)と称し;痘瘡株由来のELGFは、本明細書において、天然痘成長因子(SPGF)と称し、およびワクシニア株由来の由来のものは、ワクシニア成長因子(VGF)と称される。図3AおよびBでは、2つの痘瘡株由来のSPGF、すなわち、大痘瘡インド由来のD1L(図3A;SEQ ID N0:1)および大痘瘡バングラデシュ由来のD4R(図3B;SEQ ID NO:2)のアミノ酸配列を示してある。これらの2つのSPGFは、2つのアミノ酸部位(図3AおよびBにおいて太字で示し、下線を引いた)のみが異なる点に留意されたい。
【0030】
本発明の種々の態様を以下に記載してある。
【0031】
細胞の活性化を阻害する方法
本発明は、細胞表面erb-Bタンパク質チロシンキナーゼ分子を発現する細胞の活性化を阻害する方法を含む。これらの方法では、erbBタンパク質チロシンキナーゼ活性を阻害するか、またはerb-Bタンパク質チロシンキナーゼ活性の活性化を阻害する1つまたは複数の単離された化合物が、関心対象の細胞に送達される。1つまたは複数の化合物の細胞に対するデリバリーは、ウイルスerb-Bリガンド(たとえば、ELGF)または機能的な(すなわち、erb-Bに結合し、活性化する)erb-Bリガンドの断片と細胞の接触前に、同時に、もしくは後に生じさせることができる。本方法は、デリバリー工程の前に、erb-Bタンパク質チロシンキナーゼを発現する細胞を同定することを含むことができる。細胞は、erb-Bタンパク質チロシンキナーゼ分子を発現する任意の細胞、たとえば:任意の腫の上皮細胞、例えば肺上皮細胞などの肺上皮細胞、胃腸上皮細胞(たとえば、胃または結腸の上皮細胞)、皮膚上皮細胞(たとえば、ケラチノサイト)、または泌尿生殖系の上皮細胞(たとえば、膀胱または子宮上皮細胞);または線維芽細胞であることができる。細胞は、正常細胞または悪性細胞であることができる。erbBタンパク質チロシンキナーゼは、erb-B1、erb-B2、もしくはerb-B4タンパク質チロシンキナーゼ、これらの3つのタンパク質のいずれかのホモ二量体、またはerb-B1、erb-B2、erb-B3、およびerb-B4タンパク質チロシンキナーゼの任意の二つの間のヘテロ二量体であることができる。
【0032】
ウイルスerb-Bリガンド(EBL)は、ゲノムがerb-Bタンパク質チロシンキナーゼと結合するタンパク質をコードする核酸(DNAまたはRNA)を含む任意のウイルスに由来することができる。たとえば、ウイルスEBLは、本明細書に一覧を示した任意のウイルスであることができる。たとえば、ポックスウイルス由来であることができる。関心対象のポックスウイルス(ポックスウイルス科)は、オルソポックスウイルス属(たとえば、痘瘡、ワクシニア、およびサル痘)、トリポックスウイルス属(Avipoxviruses)(たとえば、鶏痘およびカナリア痘瘡)、カプリポックスウイルス属(たとえば、ヒツジ・ポックスウイルス)、レオポリポックスウイルス属(Leoporipoxviruses)(たとえば、粘液腫およびショープウイルス)、パラポックスウイルス属(たとえば、オルフウイルスおよび豚痘)、軟体動物ポックスウイルス属(Molluscipoxviruses)(たとえば、伝染性軟属腫ウイルス)、およびヤタポックスウイルス属(Yatapoxviruses)(たとえば、ヤタサル腫瘍ウイルス)を含むが、これらに限定されない。痘瘡(大痘瘡および小痘瘡)およびワクシニアウイルスは、特に関心がもたれる。ポックスウイルスELGFは、たとえばSPGF(たとえば、大痘瘡インド株由来のD1Lタンパク質または大痘瘡バングラデシュ株由来のD4Rタンパク質)またはVGF(ワクシニアウイルス由来)であることができる。ウイルスEBLの機能的な断片は、本明細書に使用されるものとして、断片は、全長の野生型の成熟タンパク質よりも短いが、野生型の成熟タンパク質がerb-B受容体の1つまたは複数(野生型の全長タンパク質が結合する)と結合する能力の少なくとも20%(たとえば、少なくとも:30%;40%;50%;60%;70%;80%;90%;95%;97%;98%;99%;99.5%;99.7%;99.8%;99.9%、または100%以上)を有する。リガンドが受容体と結合する能力を測定する方法は、当該技術分野において既知である(このような方法の実施例については以下を参照されたい)。機能的な断片は、たとえばEGF様ドメイン、たとえばSPGF D1L(SEQ ID NO:1)、もしくはD4R(SEQ ID NO:2)のアミノ酸40〜90であるか、またはこれを含むことができる。配列の整列により(図2を参照されたい)、当業者であれば、関心対象のEBLの領域がEGF様ドメインを含むか、またはEGF様ドメインであることを理解することができるであろう。
【0033】
細胞表面erb-Bタンパク質チロシンキナーゼに接触するウイルスEBLは、ウイルス粒子の成分であることができ、実際に、細胞表面erb-B分子に対するこの形態のウイルスEBLの結合は、ウイルス粒子による細胞の感染の少なくとも1つの初期工程であることができる。または、ウイルスEBL(またはこれらの機能的な断片)は、ウイルスに感染した細胞によって分泌されるか、または細胞外ウイルス粒子から分断することができる。加えて、ウイルスEBL(またはこれらの機能的な断片)は、たとえばウイルスから部分的もしくは完全に精製されたか、または化学的もしくは組換えで産生された分子であることができる。
【0034】
erb-Bタンパク質チロシンキナーゼを阻害する化合物は、erb-Bタンパク質と結合する抗体であることができる。このような抗体は、非アゴニスト抗体、すなわちそれが結合する受容体を介したシグナルを実質的に活性化しない抗体であることができる。erb-Bの外部ドメインに特異的な抗体は、リガンド結合に関して競合して、erb-B1シグナリングをブロックするものであると記載されており;さらに、これらの抗体は、細胞周期停止および/または細胞死を生じさせることが示された[Arteaga (2001) J. Clin Oncol. 19 (18) (September 15 supplement): 32s-40s、およびその中で引用される参照文献]。
【0035】
関連した化合物のもう一つのクラスは、ウイルスEBLに結合する抗体、たとえば3D4R-13E8および3D4R-11D7 mAbsなどのSPGFまたはVGFと結合するmAb(それぞれ13E8および11D7 mAbsとも称される)を含む。
【0036】
さらに、本発明の方法のために有用な化合物は、非アゴニストerb-Bリガンドまたはerb-Bタンパク質と結合するerb-Bリガンドの非アゴニスト断片であることができる。関心対象の受容体と結合する化合物がアゴニストまたはその受容体の非アゴニストであるかどうかを決定するための方法は、当該技術分野において既知である。erb-Bの哺乳類リガンドは、EGF、エピレギュリン、トランスフォーミング成長因子α(TGFα)、アンフィレギュリン(amphiregulin)、ヘパリン結合EGF、ベータセルリン(betacellulin)、およびヘレギュリン(heregulin)を含むが、これらに限定されない[Arteaga et al.(2001) J. Clin. Oncol. 19: 32s-40s]。その他の適切な化合物は、ウイルスタンパク質、たとえばELGF(例えばSPGFまたはVGF)の非アゴニスト断片を含む。
【0037】
小分子erb-Bチロシンキナーゼ阻害剤
特に興味がもたれる化合物は、erb-Bタンパク質チロシンキナーゼ活性の小分子阻害剤である。多くのこのような化合物が記載されている。このような化合物には、本質的に3つのカテゴリーがある。カテゴリーAには、erb-Bタンパク質チロシンキナーゼの触媒ドメインのMg-ATP結合部位に競合することによって実質的に作用するように見える化合物がある[Al-Abeidi (2000) Oncogene 18:5690-5701]。カテゴリーBには、erb-Bタンパク質チロシンキナーゼ上のチロシン部位に競合することによって明らかに実質的に作用する化合物がある[Posner et al. (1994) Mol. Pharmacol. 45:673-683、その全体が参照として本明細書に組み入れられる]。カテゴリーCには、両方の機構によって作用するように見える化合物がある[Posner et al. (1994) Mol. Pharmacol. 45:673-683]。
【0038】
カテゴリーAは、二環式のキナゾリンに基づいた化合物(たとえば、4-アニリノキナゾリン)、ピリドピラミジンに基づいた化合物、キノリン-3-カルボニトリルに基づいた化合物、ピロロピリミジンに基づいた化合物、ピラゾロピリミジンに基づいた化合物、並びにキナゾリンの三環系の誘導体(イミダゾキナゾリン、ピロロキナゾリン、およびピラゾロキナゾリン)、並びに上記の一覧に記載されたその他の二環式化合物を含むが、これらに限定されない[Fry et al. (2003) Exp. Cell Res. 284:131-139、その全体が参照として本明細書に組み入れられる]。
【0039】
キナゾリンに基づいた化合物の例は、以下を含む:ZD1839/イレッサ[Fry et al.(2003)];GW572016(N-{3-クロロ-4-[3-フルオロベンジル)オキシ]フェニル}-6-[5-({[2-メチルスルホニル)エチル]アミノ}メチル)-2-フリル]-4-キナゾリン[Xia et al. (2002) Oncogene 21:6255-6263;および Cockerill et al. (2001) Bioorg. Med. Chem. Lett. 11:1401-1405、両方ともその全体が参照として本明細書に組み入れられる];GW974[Cockerill et al. (2001);および Rusnak et al. (2001) Cancer Res. (2001) 61:7196-7203、その全体が参照として本明細書に組み入れられる];PD168393[Fry et al. (1998) Proc. Natl. Acad. Sci. USA 95:12022-12027、その全体が参照として本明細書に組み入れられる;図12]、PD153035/ZM252868/AG1517[Fry et al. (1994) Science 265:1093-1095、その全体が参照として本明細書に組み入れられる;図12]、およびCI-1033[Pfizer;図12];OSI-774/CP-358、774/Tarceva[Fry et al.(2003);および Stamos et al. (2002) J. Biol. Chem. 277:46265-46272、その全体が参照として本明細書に組み入れられる];AG1478[Levitzki et al. (1995) Science 267:1782-1788、両方ともその全体が参照として本明細書に組み入れられる];並びにPD160678、PD160879、PD168393、およびPD174265[Fry et al. (1998) Proc. Natl. Acad. Sci. USA 95:12022-12027]。
【0040】
有用なピリドピリミジンに基づいた化合物の例はPD69896、PD153717、およびPD158780[Fry et al. (1997) Biochem. Pharmacol. 54 (8):877-887、その全体が参照として本明細書に組み入れられる]を含む。有用なキノリン-3-カルボニトリルは、EKB-569[Fry et al. (2003)]であり、有用なピロロピリミジンは、CGP59326A/PKI-166[Lydon et al. (1998) Int. J. Cancer 76 (1):154-163、その全体が参照として本明細書に組み入れられる]であり、および有用なピラゾロピリミジンは、たとえば、Traxler et al. [(1997) J. Med. Chem. 40:3601-3616、その全体が参照として本明細書に組み入れられる]の化合物9および11を含む。
【0041】
広範囲にわたる置換基を有する化合物のこれらのクラスの全ての複数の誘導体が、erb-Bチロシンキナーゼ阻害活性を示すことが明らかであり、したがって、本発明の方法の候補である可能性が高い[たとえば、

を参照されたい、これらの全てが、参照としてこれらの全体が本明細書に組み入れられる]。
【0042】
いくつかの上記の化合物(たとえば、CI-1033、PD168393、EKB-569、PD1606678、PD 160879、およびPD 174265)は、アクリルアミド基によって、たとえば、erb-B1のCys773残基をアルキル化することによって、不可逆阻害剤として作用する。一連のこのようなアクリルアミド誘導体の相対的な有効性の比較は、Smaill et al [(1999) J. Med. Chem. 42(10):803-1815、その全体が参照として本明細書に組み入れられる]に記載されている。加えて、これらの化合物をアルキルアミンおよびアルコキシアミン側鎖によって置換すると(たとえば、キナゾリンおよびピリドピリミジン化合物の7位置で)、水性溶媒の溶解性の問題を解決するために役立つことが示された[Smaill et al. (2000) J. Med. Chem. 43 (7) 1380-1397、その全体が参照をそばに本明細書において組み入れられる]。
【0043】
また、カテゴリーAには、化合物DAPH 1(4,5-ジアニリノフタリミド)、およびその誘導体[Buchdunger et al.(1994)Proc. Natl. Acad. Sci. USA 91:2334-238、その全体が参照として本明細書に組み入れられる]、並びに真菌産物、ケルセチン、ゲニステイン、およびラベンズスチン(lavendustin)A[Levitzki et al. (1995) Science 267:1782-1788、その全体が参照として本明細書に組み入れられる]がある。
【0044】
カテゴリーBには、化合物AG537およびAG538[Posner et al. (1994) Mol. Pharmacol. 45:673-683]、並びにCushman et al. [(1994) J. Med. Chem. 37:3353-3362、その全体が参照として本明細書に組み入れられる)の化合物10a、10c、および10mなどのフラボン誘導体がある。
【0045】
カテゴリーCには:Geissler et al. [(1990) J. Biol. Chem. 265 (36):22255-22261、その全体が参照として本明細書に組み入れられる]の化合物1、2、および3などのチアゾリジンジオン;Showalter et al. [(1997) J. Med. Chem. 40:413-426、その全体が参照として本明細書に組み入れられる]の化合物33および34などの3-置換された2,2'-ジセレノビス(1H-インドール);Thompson et al. [(1993) J. Med. Chem. 36:2459-2469、その全体が参照として本明細書に組み入れられる]の化合物9、32、および33などの2,3-ジヒドロ2-チオキソ-1H-インドール-3-アルカノン酸および2,2'-ジヒドロビス(1H-インドール-3-アルカノン酸);並びにTraxler et al. [(1995) J. Med. Chem. 38:2441-2448、その全体が参照として本明細書に組み入れられる]の化合物5、17、18、および19などの[(アルキルアミノ)メチル]アクリロフェノンおよび(アルキルアミノ)プロピオフェノンがある。
【0046】
「小分子erb-Bチロシンキナーゼ阻害剤」の用語は、本明細書に使用されるものとして、表題「小分子erb-Bチロシンキナーゼ阻害剤」の下で上記した全ての化合物をいう。
【0047】
大分子(たとえば、抗体)および小分子の両方の化合物は、単独で細胞に、または2以上、たとえば3、4、5、6、7、8、9、10、15、または20を組み合わせて送達することができる。このような組み合わせは、上記の阻害剤のクラスの1つだけ、または1つ以上(たとえば、2、3、または4)を含む組み合わせを含むことができる。さらに、上記の化合物の効果を増強するために、補充薬を細胞または細胞の環境、たとえば細胞を含む培養液または動物に送達することができる。使用することができるそのような補充薬は、関心対象のEBLをコードする遺伝子を含むウイルスの1つまたは複数の形態と結合して、ウイルスを実質的に中和する抗体である。本明細書で使用されるものとして、「実質的に中和する」とは、少なくとも5倍、たとえば少なくとも10倍;20倍;50倍;100倍;1,000倍;10,000倍;100,000倍;1,000,000倍、またはそれ以上ウイルスの力価を減少させる能力を有することを意味する。このような抗体の例は、ポックスウイルスの細胞内成熟ウイルス粒子(IMV)(たとえば、L1Rワクシニア・タンパク質に、またはその痘瘡相同分子種に特異的な抗体)、細胞外外膜ウイルス(EEV)、または細胞結合される外膜ウイルス(CEV)と結合する中和抗体を含む。
【0048】
一定のインビボ態様において、被検者に中和抗体を投与する代わりに、被検者には、インサイチュウで中和抗体を産生させるために、適切な抗原の供与源をワクチン接種することができる。免疫化の適切な抗原および方法は、下記の免疫応答の増強法の節に記載してある。
【0049】
上記した全ての抗体は、ポリクローナル抗体またはmAbであることができ、任意の広範な腫、たとえばヒト、非ヒト霊長類(たとえば、サルまたはチンパンジー)、ウシ、ウマ、ヤギ、ヒツジ、ブタ、ネコ、イヌ、ウサギ、モルモット、ハムスター、スナネズミ、ラット、マウス、またはニワトリに由来するものであることができる。
【0050】
本明細書で使用されるものとして、「抗体」の用語は、完全な抗体(たとえば、IgM、IgG、IgA、IgD、またはIgE)分子だけではなく、抗原結合性断片、たとえばFab、F(ab')2、Fv、および単鎖のFv(scFv)断片もいう。scFv断片は、scFvが誘導される抗体の重鎖および軽鎖の可変領域の両方を含む単一のポリペプチド鎖である。また、キメラ抗体、たとえばヒト化抗体も含まれる。
【0051】
分子の結合ドメインを含む抗体断片は、既知の技術によって作製することができる。たとえば:F(ab')2断片は、抗体分子のペプシン消化によって作製することができ;およびFab断片は、F(ab')2断片のジスルフィド架橋を還元させることによって、またはパパインおよび還元剤で抗体分子を処置することによって作製することができる。たとえば、National Institutes of Health, 1 Current Protocols In Immunology, Coligan et al., ed. 2.8, 2.10 (Wiley Interscience, 1991)を参照されたい。scFv断片は、たとえばU.S. Patent No. 4,642,334(その全体が参照として本明細書に組み入れられる)に記載されているように作製することができる。
【0052】
たとえば、キメラ・ヒト化モノクローナル抗体は、Robinson et al., 国際特許出願第PCT/US86/02269号;Akira et al., 欧州特許出願第184,187号;Taniguchi, 欧州特許出願第171,496号;Morrison et al., 欧州特許出願第173,494号;Neuberger et al., PCT出願 国際公開公報第86/01533号;Cabilly et al., 米国特許第4,816,567号;Cabilly et al., 欧州特許出願第125,023号;Better et al. (1988) Science 240,1041-43 ; Liu et al. (1987) J. Immunol. 139, 3521-26;Sun et al. (1987) PNAS 84, 214-18;Nishimura et al. (1987) Canc. Res. 47,999-1005;Wood et al. (1985) Nature 314, 446-49;Shaw et al. (1988) J. Natl. Cancer Inst. 80, 1553-59;Morrison, (1985) Science 229, 1202-07;Oi et al. (1986) BioTechniques 4, 214;Winter, 米国特許第5,225,539号;Jones et al.(1986) Nature 321, 552-25;Veroeyan et al. (1988) Science 239, 1534;および、Beidler et al. (1988) J. Immunol. 141, 4053-60に記載されている方法を使用して当該技術分野において既知の組換えDNA技術によって作製することができる。
【0053】
完全ヒト抗体(ポリクローナルまたはモノクローナル)は、全てのヒト免疫グロブリン(すなわち、可変、接合、多様性、および定常)領域をコードする遺伝子セグメントをコードする遺伝子セグメントを含むトランスジェニック動物(たとえば、マウス)を免疫することによって産生することができる(たとえば、U.S. Patent Nos.5,545,806および5,569,825を参照されたい)。
【0054】
出願人は、アメリカン・タイプ・カルチャー・コレクション(ATCC), Rockville, MD 20852, U.S.Aで3D4R-11D7および3D4R-13E8ハイブリドーマをブダペスト条約の下で寄託した。3D4R-11D7ハイブリドーマには、ATCCアクセッション番号PTA-5039が、および3D4R-13E8ハイブリドーマにはATCCアクセッション番号PTA-5040が割り当てられた。ATCCで寄託したハイブリドーマは、本出願の優先日の前から Dana Farber Cancer Institute, Inc.によって維持された寄託から得られる。ハイブリドーマの寄託により、30年の期間、またはごく最近の要請後5年の期間、または有効な特許存続期間の間は、どんなに長くても、ATCC寄託所に制限なしで維持され、その期間の間に寄託が生育不能になった場合は交換される。
【0055】
細胞の活性化を阻害する方法は、インビトロまたはインビボであることができる。
【0056】
本発明の方法のインビトロでの適用は、ウイルス感染およびウイルス感染に対する細胞抵抗の機構に対する基本的科学的研究に有用であろう。本発明のインビトロ方法において、化合物は、関心対象の細胞(上記のものを参照)および上記で詳述した任意のウイルスEBLの形態と共に培養することができる。たとえば、ウイルス力価、細胞増殖/生存のレベル、および/またはタンパク質リン酸化の測定は、本明細書において記載され、および当該技術分野において既知の方法を使用して、種々のインキュベーション時間後に行うことができる。
【0057】
さらに、このような方法は、試験化合物を、これらが細胞のウイルス感染を阻害する能力についてスクリーニングするために有用であろう。このようなアッセイ法では、細胞と共に試験化合物を培養することによって、erb-B受容体の関心対象に結合する試験化合物をerb-B受容体を発現する細胞と接触させることができる。接触させる前、同時に、もしくは後に、細胞をウイルスEBLまたはEBLの機能的な断片と接触させることができる。次いで試験化合物が、リガンドまたは機能的な断片によってerb-B受容体の活性化が減少されるかどうかの決定がなされる。erb-B受容体の活性化は、当該技術分野において既知の方法によって、たとえば(a)活性化されたerb-B受容体によって直接、もしくはerb-B受容体の活性化に続いてカスケード様式で活性化されるタンパク質によって間接的にリン酸化されるerb-B受容体または下流の細胞内タンパク質のチロシン・リン酸化を検出または測定すること;(b)erb-B受容体のインターナリゼーションを検出または測定すること;または、(c)細胞の増殖および/または生存を検出または測定することによって測定することができる。
【0058】
また、使用する化合物がerb-B受容体を活性化することが既知である際の本発明のインビトロ方法は、ウイルスEBLによる細胞の活性化を阻害する能力を有する化合物についての上記スクリーニングアッセイ法の「ポジティブ対照」となり得る。
【0059】
本発明の方法は、好ましくはインビボで行われる。これらの適用は、関連したウイルス病の治療および予防に有用であろう。また、これらは、生きているウイルス(たとえば、牛痘)のワクチン接種の副作用を減弱させるためにも有用であろう。感染動物(たとえば、ヒト天然痘患者)、EBL発現ウイルスに感染するであろう動物(たとえば、ワクシニアウイルスでワクチン接種されたヒト被験者)、もしくは感染するリスクのある動物(たとえば、予想されるバイオテロリストの攻撃の間に大痘瘡ウイルスに感染するリスクのあるヒト被験者)に関心対象の化合物を投与することによって、ウイルスによって生じる臨床症状からの治療または予防を達成することができる。
【0060】
本明細書で使用されるものとして、「予防」は、疾患の症状の完全な予防、疾患の症状の発症の遅延、またはその後に発生する疾患症状の重篤さを少なくすることを意味することができる。本明細書で使用されるものとして、「治療」は、疾患の症状の完全な消滅または疾患の症状の重篤さの減少を意味することができる。
【0061】
本発明の上記のインビボの方法の修飾は、関心対象のウイルスに対する有効な予防薬および/または治療薬である化合物のスクリーニングアッセイ法として使用することができる。このような方法では、erb-Bチロシンキナーゼの活性を阻害するか、またはerb-Bチロシンキナーゼの活性化を阻害する試験化合物が、EBLをコードする遺伝子を含むウイルスに対する感染に感受性の動物に投与される。動物は、投与の前に、の間、またはの後、ウイルスに曝露される。次いで、化合物の有効性の決定は、たとえば体液(たとえば、血液、尿、または粘膜)、器官もしくは組織(たとえば、肺、脾臓、または肝臓組織)、または洗浄液(たとえば、肺、腸、膀胱、または膣の洗浄液)中のウイルスの力価の変化を検出または測定することによってなされる。または、ウイルス感染の1つまたは複数の症状(たとえば、熱、発疹、肢麻痺)の減少を検出または測定することができる。
【0062】
、ウマ、牛、ブタ、ヒツジ、ヤギ、イヌ、ネコ、ウサギ、モルモット、ハムスター、ラット、マウス、並びにニワトリ、シチメンチョウ、およびカナリアなどのトリに適用することができる。
【0063】
インビボ・アプローチ
好ましいインビボ・アプローチにおいて、単離された化合物自体が被検者に投与される。通常、化合物は、薬学的に許容されるキャリア(たとえば、生理食塩水)に懸濁して、経口で、もしくは静脈内(i.v.)注入によって投与されるか、または皮下に、筋肉内に、くも膜下腔内に、腹腔内に、内直腸に、腟内に、鼻腔内に、胃内に、気管内に、または肺内に注射される。これらは、たとえば、感染の部位、たとえば感染症が肺の中である場合は、肺内に直接送達することができる。必要とされる用量は、投与の経路の選択;製剤の性質;患者の疾病の性質;被検者のサイズ、重量、表面積、年齢、および性;投与されている他剤;並びに主治医の判断に依存する。適切な用量は、0.0001〜100.0mg/kgの範囲である。必要とされる用量の広範なバリエーションは、利用できる化合物の多様性および種々の投与経路での異なる効率から考えて予想される。たとえば、経口投与は、i.v.注射による投与よりも高い用量が必要であると思われる。これらの用量レベルのバリエーションは、当該技術分野において周知であるように、最適化のための標準的な経験的ルーチンを使用して調整することができる。投与は、一回または複数回(たとえば、2、3、4、6、8、10、20、50、100、150、またはそれ以上の回数)であることができる。適切なデリバリー媒体(たとえば、重合体の微小粒子または移植可能な装置)に化合物をカプセル化することにより、特に経口デリバリーに関してのデリバリーの効率を増大するであろう。
【0064】
有用な化合物がタンパク質である場合、これらは、このようなタンパク質をコードする核酸を含む発現ベクターを、またはこのような発現ベクターで形質転換された細胞(たとえば、動物に由来する細胞)を、細胞が存在する動物に対してインビボで投与することによって細胞に送達することができることが理解される。このような方法は、当該技術分野において周知である。
【0065】
免疫応答を増強する方法
本発明は、免疫応答を増強する方法を特徴とする。増強される免疫応答は、CD8+またはCD4+ T細胞応答であることができる。T細胞応答の増強は、サイトカイン/リンホカイン産生性または細胞障害性Tリンパ球(CTL)の反応であることができる。サイトカイン/リンホカイン産生性反応は、好ましくはTh1タイプのサイトカイン/リンホカイン反応、たとえばインターロイキン(IL)-12およびIFN-γを産生する反応である。しかし、また、一定の状況下で(たとえば、抗体対細胞免疫反応が好ましい)、インターロイキン(IL)-2、IL-3、IL-4、IL-10、IL-13、IL-15、顆粒球コロニー刺激因子(G-CSF)、マクロファージコロニー刺激因子(M-CSF)、もしくは顆粒球マクロファージ-コロニー刺激因子(GM-CSF)などのサイトカインまたはリンホカインの産生を含む反応を活性化することも望ましいであろう。(1)本発明の方法が、CD4+ T細胞応答並びにCD8+ T細胞応答を増強することが示されており(実施例6および7)、かつ(2)B細胞(抗体産生性)応答には、一般にCD4+ヘルパーT細胞の活性が必要である:が故に、本発明の方法によって増強することができる免疫応答は、B細胞応答を含む。
【0066】
本発明のこれらの方法は、インビボであることができ、上記細胞の活性化を阻害するインビボ法と本質的に同様である。加えて、これらは、後者の方法を上で詳述したのと同じ被検者に適用することができる。本方法は、以下のさらなる工程を含むことができる:1つまたは複数の治療を行う工程、動物における免疫応答について試験する工程、および/または当該技術分野においてなじみがある方法によって、反応を、たとえばT細胞応答、CD4+ T細胞応答、CD8+ T細胞応答、または抗体産生B細胞応答として特徴付ける工程。
【0067】
免疫応答性を増強することを要求される場合、関心対象のウイルスの1つまたは複数の形態を実質的に中和する抗体(上記参照)に加えて、補充薬を使用することができる。特に関心対象の免疫応答を増強する方法は、ウイルスEBL(たとえば、mAbs 13E8および11D7)に結合する上記の抗体の1つまたは複数、および関心対象のウイルスの1つまたは複数の形態を実質的に中和する1つまたは複数の抗体、たとえば抗L1R抗体を投与することを含む。
【0068】
免疫応答を増強するために有用なさらなる補充薬は、たとえば適切なウイルス抗原を含む。抗原は、被検者が感染するか、するであろうか、またはするリスクがあり、かつ投与された化合物がウイルス複製増強効果を阻害するウイルスEBLの供与源であるウイルスの成分であることができ、たとえば免疫を増強する方法は、生きているウイルスでのワクチン接種の効率を増強するために使用する場合である(たとえば、ワクシニアウイルスでの種痘に対するワクチン接種)。しかし、さらなる抗原を投与することもできる。このような抗原は、たとえば死滅したウイルス、ウイルスタンパク質、またはウイルスタンパク質のペプチド断片であることができる。抗原は、単独で、またはアジュバント、たとえばコレラ毒素(CT)、大腸菌熱易動性毒素(LT)、変異体CT(MCT)[Yamamoto et al.(1997) J. Exp. Med. 185:1203-1210]、および変異体大腸菌熱不安定性毒素(MLT)[Di Tommaso et al. (1996) Infect. Immunity 64:974-979]と共に投与することができる。MCTおよびMLTは、親分子のものと比較して実質的にアジュバント活性を損なうことなく実質的に毒性を減弱させる点突然変異を含む。その他の有用なアジュバントは、ミョウバン、フロイント完全および不完全アジュバント、並びにRIBIを含む。
【0069】
さらに免疫を増強するために用いることができるその他の補充薬は、MIP-1α、MIP-3β、RANTES、IL-1、IL-2、IL-3、IL-4、IL-5、IL-6、IL-7、IL-8、IL-10、IL-12、IL-13、IL-15、インターフェロン(IFN)-α、β、もしくはγ、顆粒球コロニー刺激因子(G-CSF)、マクロファージコロニー刺激因子(M-CSF)、または顆粒球マクロファージ-コロニー刺激因子(GM-CSF)などのケモカイン、リンホカイン、およびサイトカインを含む。加えて、関心対象の免疫応答を阻害する補充薬、ケモカイン、リンホカイン、またはサイトカインに特異的な抗体を使用することができる。従って、たとえば、それは細胞の免疫応答を増強することが要求される場合、IL-4、またはIL-10に特異的な抗体を補充薬として使用してもよい。一方では、抗体産生B細胞応答を増強することが要求される場合、IL-12またはIFN-γに特異的な抗体を補充薬として使用してもよい。
【0070】
細胞の活性化を阻害する方法の節において上記した化合物のデリバリーおよび投与の全ての方法論(または、これらの明らかなバリエーション)は、本明細書に記載されている全ての補充薬のデリバリーおよび投与のために使用することができることが理解される。これらの補充薬は、化合物の前に、後に、または同時に投与することができる。これらは、化合物と同じか、もしくは異なる経路によって、および同じか、もしくは異なる頻度で投与することができる。
【0071】
本発明のこれらの方法は、遺伝子の免疫化に適用することができる。このような方法では、関心対象の免疫原をコードする異種核酸配列を含むポックスウイルス発現ベクターが、適切な動物(たとえば、ヒト、非ヒト霊長類(たとえば、サルまたはチンパンジー)、ウシ、ウマ、ヤギ、ヒツジ、ブタ、ネコ、イヌ、ウサギ、モルモット、ハムスター、スナネズミ、ラット、マウス、ニワトリ、または本明細書において詳述した任意のその他の種)に対して、当該技術分野において既知の方法によって有効であることが決定された頻度で投与される。ベクター内では、異種核酸配列が転写調節性のエレメント(TRE)に対して動作可能に連結される。TREは、プロモーターおよびエンハンサーを含み、当該技術分野において既知である。本明細書で使用されるものとして、コード配列に「動作可能に連結された」発現制御配列(たとえば、TRE)は、これがコード配列の発現を効率的に制御するように、遺伝子構築物内に組み入れられる。関心対象のポックスウイルス・ベクターは、ワクシニア・ベクター、弱毒ワクシニア・ベクター、カナリア痘瘡ベクター、および伝染性上皮腫ベクターを含む。
【0072】
投与の経路は、好ましくは皮膚(たとえば、乱切によって)、筋肉内、皮下、静脈内、または腹腔内、または粘膜(たとえば、鼻腔内もしくは直腸内)であろう。ベクターの投与の前に、同時に、または後に、上記したerb-B抑制化合物の1つまたは複数が動物に投与される。任意の上記した補充薬を動物に投与することもできる。また、化合物および補充薬の投与の頻度は、当該技術分野における技術のうちの1つによって容易に決定することができる。
【0073】
erb-Bチロシンキナーゼ阻害剤の投与により、ポックスウイルスに感染した動物のCD4+およびCD8+免疫応答を増強したので(実施例6および7を参照されたい)、ポックスウイルス・ベクターを使用する上記の遺伝子免疫化ストラテジーは、免疫化に使用したポックスウイルス・ベクター内の異種核酸配列によってコードされる免疫原に対する細胞性応答(CD8+およびCD4+ T細胞)またはB細胞媒介応答の両方の増強に有効であろう。これらは、多種多様な免疫原に対する治療的、予防的、または非治療的/非予防的な免疫応答を生じさせるために使用することができる。これらの方法論により、複製能のあるポックスウイルス・ベクター(たとえば、ヒトにおけるワクシニア・ベクター)を使用することができ、非複製ポックスウイルス・ベクター(たとえば、ヒトにおいてカナリア痘瘡ベクター)よりも強力な反応を誘導するが、遺伝的免疫化に対して、宿主動物(たとえば、免疫無防備状態の患者)の感染が制御不能となる可能性の懸念が生じる。従って、関心対象の免疫原をコードする配列を含む複製能のあるポックスウイルス・ベクターが被検者に投与される。十分な時間ベクターを十分に複製させて、被検者の強力な免疫応答を開始するために十分な免疫原を生じさせることができる。有効な時間を確立するために、当該技術分野において周知の方法を使用することができる。次いで、1つまたは複数のerb-B抑制化合物が(1つもしくは複数の補充薬を伴って、または伴わずに)被検者に投与される。この投与は、被検者におけるウイルスベクターの力価を減少させるため、および免疫原に対する免疫応答を増強するための両方のために役立つ。
【0074】
関心対象の免疫原は、たとえば微生物(たとえば、ウイルス、細菌、真菌、酵母、または原生動物)抗原、寄生虫(たとえば、線虫)由来抗原、腫瘍抗原、または研究的、治療的、予防的、もしくは診断の目的のための抗体(ポリクローナルまたはモノクローナル)を作製することが要求されるものに対するタンパク質であることができる。
【0075】
得ることができる免疫原をコードする核酸配列に由来する関連した微生物の例は、結核菌(Mycobacteria tuberculosis)、腸炎菌(Salmonella enteriditis)、リステリア菌、癩病菌(M. leprae)、黄色ブドウ球菌(Staphylococcus aureus)、大腸菌(Escherichia coli)、肺炎連鎖球菌(Streptococcus pneumoniae)、ボレリア・ブルグドルフェリ(Borrelia burgdorferi)、肺疫菌(Actinobacillus pleuropneumoniae)、ピロリ菌(Helicobacter pylori)、髄膜炎菌、エンテロコリチカ菌(Yersinia enterocolitica)、百日咳菌(Bordetella pertussis)、ポルフィノモナス・ギンギバリス(Porphyromonas gingivalis)、マイコプラズマ、ヒストプラスマ・カプスラーツム(Histoplasma capsulatum)、クリプトコッカス・ネオフォルマンス(Cryptococcus neoformans)、トラコーマ病原体(Chlamydia trachomatis)、鵞口瘡カンジダ(Candida albicans)、熱帯熱マラリア原虫(Plasmodium falciparum)、三日熱マラリア(Plasmodium vivax)、四日熱マラリア原虫(Plasmodium malariae)、赤痢アメーバ(Entamoeba histolytica)、トキソプラズマ・ブルセイ(Toxoplasma brucei)、トキソプラズマ・ゴンジ(Toxoplasma gondii)、リーシュマニア・メジャー(Leishmania major)、ヒト免疫不全症ウイルス1および2、インフルエンザウイルス、はしかウイルス(measles virus)、狂犬病ウイルス(rabies virus)、肝炎ウイルスA、B、およびC、ロタウイルス、パピローマウイルス、呼吸器合胞体ウイルス、ネコ免疫不全ウイルス、ネコ白血病ウイルス、サル免疫不全ウイルス、および本明細書において詳述した全てのポックスウイルスを含むが、これらに限定されない。関連した微生物免疫原の例は、大腸菌の熱易動性エンテロトキシン[Konieczny et al. (2000) FEMS Immunol. Med. Microbiol. 27 (4):321-332]のBサブユニット、熱ショックタンパク質、たとえばエンテロコリチカ菌(Y enterocolitica)の熱ショックタンパク質60[Konieczny et al. (2000) supra; Mertz et al. (2000) J. Immunol. 164 (3):1529-1537]、および結核菌熱ショックタンパク質hsp60およびhsp70;トラコーマ病原体外膜タンパク質[Ortiz et al. (2000) Infect. Immun. 68 (3):1719-1723];B.ブルグドルフェリ(B. burgdorferi)外面タンパク質[Chen et al. (1999) Arthritis Rheum. 42 (9): 1813-1823];L.メジャー(L. major)GP63[White et al. (1999) Vaccine 17 (17):2150-2161 (およびVaccine 17 (20-21):2755の公表された正誤表)];髄膜炎菌(N. meningitidis)の髄膜炎菌血清型15のPorBタンパク質[Delvig et al. (1997) Clin. Immunol. Immunopathol. 85 (2):134-142];P.ギンギバリス(P. gingivalis)381海馬采タンパク質[Ogawa, (1994) J. Med. Microbiol. 41 (5):349-358];大腸菌外膜タンパク質F[Williams et al. (2000) Infect. Immun. 68 (5):2535-2545];インフルエンザウイルス血球凝集素およびノイラミニダーゼ;レトロウイルス(たとえば、HIV)表面糖タンパク質(たとえば、HIV gp160/120)、またはレトロウイルスのtatもしくはgagタンパク質を含むが、これに限定されない。
【0076】
ポックスウイルス・ベクターに挿入される免疫原をコードする核酸を得ることができる腫瘍の例は、神経組織癌、黒色腫、乳癌、肺癌、胃腸の癌、卵嚢癌、精巣癌、肺癌、前立腺癌、子宮頚癌、膀胱癌、膣癌、肝臓癌、腎癌、骨癌、血液細胞癌、および血管組織癌を含む。関連した腫瘍抗原の例は、前立腺特異的膜抗原(PSMA)[Israeli et al. (1993) Cancer Res. 53 (2):227-230]、MUC-1などのムチン、または腫瘍細胞の表面上に発現されるその他の任意の抗原を含む。
【0077】
ポックスウイルス・ベクターの異種核酸配列は、全長未成熟のタンパク質、全長成熟したタンパク質、免疫原性エピトープを含むタンパク質のセグメント、またはT細胞エピトープを含むペプチド断片をコードすることができる。これらは、単一のポリペプチド配列または複数(たとえば、2、3、4、5、6、7、8、9、10、11、12、15、18、20、25、30以上)のポリペプチドをコードすることができる。異種核酸配列が複数のポリペプチドをコードする場合、ポリペプチドは、1つもしくは複数(たとえば、2、3、4、5、6、7、8、9、または10)の微生物の生物体および/または腫瘍に由来することができる。
【0078】
上記のポックスウイルス・ベクター/erb-B阻害剤のストラテジーでは、遺伝的免疫化に限定されないと理解されるが、免疫原の代わりに、たとえばサイトカインもしくは成長因子(本明細書において列記したもののいずれかなど)、薬物もしくはプロドラッグ、または関心対象の被検者では欠損した酵素、たとえばアデノシンデアミナーゼなどの治療的分子をコードする異種核酸配列を、ポックスウイルス・ベクター遺伝子治療法に一般に適用することができる。
【0079】
ポックスウイルスおよびこれらの使用は、Paoletti [(1996) Proc. Natl. Acad. Sci. USA 93:11349-11353]に更に詳細に記載されており、その全体が参照として本明細書に組み入れられる。
【0080】
特定の処方計画が、特定の疾患に対して治療的、または予防的かどうかを試験するための方法は、当該技術分野において既知である。治療効果が試験される場合、疾患の症状を示す試験集団(たとえば、天然痘を有するヒトまたは実験動物)を任意の上記したストラテジーを含む試験処方計画で治療する。疾患の症状を示す対照群も、偽薬または異なる処方計画で処置する。試験被検者における疾患症状の消滅または減少は、試験処方計画が有効な治療法であることを示すであろう。
【0081】
疾患症状の発症前に同じストラテジーを被検者に適用することによって(たとえば、ワクシニアウイルスでの作為的な感染前の実験動物)、試験処方計画を予防法としての有効性について試験することができる。この状況では、疾患症状の発症の予防、または遅延が試験される。
【0082】
以下の実施例は、本発明を例証することを意味し、限定しない。
【0083】
実施例
実施例1.材料および方法
SPGF D4Rの組換えEGF様ドメインの産生
SPGF D4RのEGF様ドメインを含むポリペプチド(SEQ ID NO:2のアミノ酸残基40〜90)をE.coli DL21細胞において封入体として発現させて、生じるタンパク質をヒトEGFおよびマウスエピレギュリン(EPI)から作製された類似のEGF様ドメインと比較した。便宜のために、これらのEGF様ドメインをそれぞれrecSPGF、recEGF、およびrecEPIと称する。封入体は、6Mグアニジン-HCl溶液に溶解し、ポリペプチドを、6つのヒスチジン残基を含むポリペプチドのN末端のタグ配列を利用してNi2+-NTAカラムクロマトグラフィーによって精製した。部分的に精製したEGF様ドメインをレドックス緩衝液(2mM還元および1mM酸化グルタチオン)中で再び折り畳ませて、ポリペプチドをRP-HPLC(逆相高速液体クロマトグラフィー)を使用してさらに精製した。RecSPGF(8kDa)を15%のナトリウムSDS-PAGEによってrecEGF(6kDA)およびrecEPI(5kDa)と比較し;タンパク質をクーマシーブルーで染色した(図1)。
【0084】
ヒト・ケラチノサイトおよび線維芽細胞の分裂促進因子アッセイ法
正常な初代ヒト真皮線維芽細胞株R2F[Tubo et al.(1987)Oncogene Re.1:407-421]は、15%の子ウシ血清(HyClone, Inc., Logan, UT)および10ng/mlのEGFを補ったDMEM/F12培地(Life Technologies, Inc., Carlsbad, CA)からなる「完全線維芽細胞培地」中で培養した。正常な初代ヒト表皮ケラチノサイト株N[Schon et al. J. Invest. Dermatol. 107:428-438]は、前述したように[Rheinwald et al. (2002) Mol. Cell Biol. 22:5157-5172]、30μg/mlウシ下垂体抽出物(BPE)、0.2ng/mlのEGF、および0.3mMのCacl2を補ったGIBCOケラチノサイト無血清培地(Life Technologies, Inc.)からなる「完全ケラチノサイト培地」中で培養した。
【0085】
分裂促進アッセイ法では、添加したerb-Bリガンドの濃度範囲に応答するこれらの増殖を評価することができるように、EGFを含まず、かつ血清の量を減少した(およびN細胞については、BPEの量を減少して)以外は、関連培地中に低密度で細胞をまくことによって行った。
【0086】
3000個のR2F細胞を9cm2の培養ウェルに、1%の子ウシ血清を含むDMEM/F12培地中にまいた。その翌日、およびプレートにまいて4日後に、ウェルには、recEGFまたはrecSPGF(種々の濃度で)を補った培地か、または完全線維芽細胞培地を与えた。
【0087】
1000個のN細胞を9cm2の培養ウェルに、BPE(15μg/ml)および0.3mMのCacl2を含むGIBCOケラチノサイト無血清培地にまいた。その翌日、およびプレートにまいて4日後に、ウェルには、recEGFまたはrecSPGF(種々の濃度で)を補った培地か、または完全ケラチノサイト培地を与えた。
【0088】
プレートにまいて6日後に、細胞をトリプシン処理することによって培養ウェルの底から剥がして計数した。増殖速度は、1日あたりの集団倍加:[log2(6日後の細胞数/まいた細胞の数)]/6として測定した。
【0089】
チロシンキナーゼ阻害実験については、ヒト包皮線維芽細胞(SC-J)を6ウェルの組織培養プレートにまいて、2%のFBS(ウシ胎児血清)DMEM培地中で一晩「不足させた(starved)」。細胞を37℃で1時間チロシンキナーゼ阻害剤で前処理し、次いで異なる量のrecSPGFで刺激した。18時間後に、細胞を集めて、80%の氷冷エタノール中で、4℃で1時間固定した。リン酸緩衝食塩水(PBS)で一回洗浄した後、細胞を50μg/ml RNase Aを含む2.5μg/mlヨウ化プロピジウムの溶液で37℃において30分間染色した。細胞周期データをFFC解析によって得て、Cell Quest(登録商標)(Becton Dickinson, San Jose, CA)およびModfit(登録商標)(Becton Dickinson)ソフトウェアを使用して解析した。
【0090】
RecSPGFのビオチン化および上皮細胞に対する結合をFFCによって解析した
RecSPGFは、製造業者のプロトコルに従ってECLタンパク質ビオチン化モジュール(Amersham Biosciences, Piscataway, NJ)を使用してビオチン化した。1×105 MB468(またはMB453)上皮細胞をそれぞれの試験試料のために使用した。直接結合アッセイ法のために、1つの細胞の一定分量を1μgの抗erb-B1ブロッキング抗体(mAb 528;Santa Cruz Biotechnology, Inc., Santa Cruz, CA)と共に4℃で1時間インキュベートした。異なる濃度のビオチン化されたrecSPGFの50μl FFC緩衝液(1×PBS/2.5%のFCS/0.02%のNaN3)溶液を試料に添加し、次いで、4℃で45分間さらにインキュベートした。競合アッセイ法のためには、細胞をラベルされていない異なる量のrecSPGF、recEPI、またはrecEGFを含む50μl FFC緩衝液と共に4℃で20分間インキュベートした。ビオチン化されたrecSPGF(25ng)を全ての試料に添加し、次いでこれを4℃で30分間インキュベートした。細胞をFFC緩衝液中で洗浄し、結合したビオチン化recSPGFをフィコエリトリン(PE)結合ストレプトアビジン(Molecular Probes, Eugene, OR)でFFCによって検出した。収集されるデータは、平均蛍光強度(MFI)であった。リガンド結合の阻害は、MFIexpが実験試料で検出されるMFIである場合、(MFIexp-MFImin/MFImax-MFImin)の割合として表してあり、MFIminは、ビオチン化されたrecSPGFが添加されなかったネガティブ対照試料で検出されるMFIであり、MFImaxは、ビオチン化されたrecSPGFが添加されたが、阻害剤が添加されなかったポジティブ対照試料で検出されるMFIである。
【0091】
細胞可溶化物の調製、免疫沈降、およびウエスタン・ブロッティング
10cmの培養ディッシュ内のHeLa細胞(80%〜90%コンフルエント)を37℃で10分間50ng/mlの濃度の種々のerb-B1リガンドで刺激した。ディッシュを氷冷PBSで一度洗浄した。次いで、細胞を1mlの溶解緩衝液(25mMのトリスpH7.4、150mMのNaCl、1%のTritonX-100、1mMのフェニルメチルスルホニルフルオライド、0.35トリプシン阻害剤ユニット/mlアプロチニン、5μg/mlのロイペプチン、10mMのNaF、10mMのβ-グリセロリン酸、および1mMのNa3VO4)でディッシュ内において直接4℃で30分間溶解した。可溶化液をミクロチューブに移し、遠心分離後、0.5mlの可溶化液上清を1μgのヤギ抗EGFRポリクローナル抗体(Santa Cruz Biotechnology, Inc.)および10μlのガンマ結合プラス(商標)ビーズ(Amersham Biosciences)で4℃において一晩に免疫沈降した。ビーズを3回洗浄し、タンパク質を2×SDS-PAGE充填緩衝液に直接ビーズから溶出させた。全細胞の可溶化液または免疫沈降された試料の一定分量を7.5%のSDS-PAGEで分解し、生じるゲルをPVDF(ポリビニリデンフルオライド)膜にブロットした。膜を2.5%のウシ血清アルブミンを含むトリス-緩衝食塩水/Tween-20(TBST)(erb-B受容体に関して)または2%のゼラチンを含むTBST(4G10に関して)中で37℃において少なくとも30分間ブロックした。膜は、製造業者の説明書に従って一次抗体と共に4℃で一晩インキュベートし、洗浄し、1:10,000抗ヤギIgG(erb-B受容体のために)または抗マウスIgG2b(4G10のために)HRPO(西洋ワサビペルオキシダーゼ)抱合体と共に室温で1時間インキュベートした。広範な洗浄後に、膜を化学発光試薬キット(Perkin Elmer Biosystems, Wellesley, MA)およびMRフィルム(Eastman Kodak Co., Rochester, NY)を使用して発色させた。
【0092】
チロシンキナーゼ阻害実験では、HeLa細胞を37℃で30分間50nMで阻害剤で処置し、続いてrecSPGF(50ng/ml)で刺激した。細胞可溶化物を調製し、標準的な方法を使用して、ヤギ抗erb-B1ポリクローナル抗体(Santa Cruz Biotechnology, Inc.)およびガンマ結合プラスビーズまたは抗c-Cbl抗体被覆ビーズ(Santa Cruz Biotechnology, Inc.)で4℃で一晩免疫沈降(上記の通り)に供した。ビーズを洗浄し、2×SDS-PAGE充填緩衝液に溶出した。全細胞の可溶化液または免疫沈降物を抗EGFRまたは抗c-Cblポリクローナル抗体でのウエスタン・ブロッティングによって解析した。
【0093】
recSPGFおよびerb-B受容体の化学的架橋
2.25×106のMB468またはMB453細胞を、受容体インターナリゼーションを阻害するために4℃で30min間0.04%のNaN3で前処理した。次いで、細胞をビオチン化されたrecSPGF(1μg/ml)と共に4℃で30分間インキュベートした。培養ディッシュを氷冷DMEMで洗浄し、ビス(スルホスクシニミジル)-スベラート(BS3)架橋剤を1mMの濃度で添加した。反応混合物を随時振盪しながらさらに1時間4℃でインキュベートした。氷冷PBSで2回洗浄した後、1mlの溶解緩衝液(25mMのトリスpH7.4、150mMのNaCl、1%のTritonX-100、1mMのフェニルメチルスルホニルフルオライド、0.35トリプシン阻害剤ユニット/mlアプロチニン、および5μg/mlロイペプチン)を直接ディッシュに添加し、続いて4℃で30分間振盪した。可溶化液をミクロチューブに移し、遠心分離後、可溶化液の上清を1μg抗erb-B1、抗erb-B3、もしくは抗erb-B4ヤギ・ポリクローナル抗体(Santa Cruz Biotechnology)、または1:40希釈抗erb-B2(Cell Signaling Technology, Beverly, MA)ウサギ・ポリクローナル抗体、および10μlのガンマ結合プラスビーズでの免疫沈降(IP)に供した。試料を4℃で5時間回転させた後に、ビーズを1×TBS/1%のTriton X-100で4回洗浄し、次いで2×SDS-PAGE充填緩衝液中で煮沸した。ビオチン化されたrecSPGFを検出するために、SDS-PAGEおよびストレプトアビジン-HRPOでのウエスタン・ブロッティングを使用した。
【0094】
EGFRに結合するrecSPGFの親和性
recSPGFの125I-ラベリングのためにラクトペルオキシダーゼ触媒法を使用し、recEGFの125I-ラベリングのために、クロラミンT触媒法を使用した。ラベルされたrecSPGFの比活性は、0.54pm/cpmであり、recEGFのものは、0.34pm/cpmであった。MB468細胞は、3×104/0.1ml/96ウェルの濃度でまいた。125IのラベルされたrecSPGFまたはrecEGFを種々の濃度で細胞の50μlの結合培地溶液(0.1%のNaN3を含むL15培地)に4℃で添加し、5時間インキュベートした。上清を回収し、細胞を60μlの氷冷結合培地で迅速に2回洗浄した。次いで、上清および洗浄液を合わせた。50μlの0.5N NaOHをそれぞれのウェルに添加し、プレートを室温で1h間インキュベートして接着細胞を溶解し、さらに60μlの0.5N NaOHで一度洗浄した。可溶化液および洗浄液を合わせた。γカウントを使用して、培養液上清中の遊離リガンドの量および細胞可溶化物中の結合したリガンドの量を決定した。受容体数および親和性は、スキャッチャード解析によって決定した(たとえば、Current Protocols in Immunology. Eds. John E. Coligan et al., John Wiley & Sons. (2001))。
【0095】
Erb-B1インターナリゼーション
HeLa細胞をチャンバ・スライド上で培養しErb-B1リガンド(50ng/ml)によって37℃で10分間刺激した。次いで、細胞を3.7%のホルムアルデヒドで10分間固定して、0.1%のTriton X-100で室温において5min間透過化処理した。細胞を室温で30分間、1%のBSA/PBSのmAb(Santa Cruz Biotechnology, Inc.)、1μgの抗erb-B1とインキュベートした。結合した抗erb-B1 mAbを抗マウスIg-FITC(フルオレッセインイソチオシアネート)抱合体で染色することによって検出した。
【0096】
A431表皮癌細胞を50nM阻害剤によって4℃で30分間前処理した。細胞をビオチン化されたrecSPGF(100ng)と共に4℃で30分間インキュベートした。洗浄後、細胞を37℃で5分間インキュベートし、ホルムアルデヒド中で室温において5分間固定した。ストレプトアビジン-PE染色の後に、種々の試料のMFIsをFFCによって得た。
【0097】
細胞およびウイルス
ワクシニアウイルス株WRを、L929細胞内で増殖させて、ウイルス力価アッセイ法を前述したようにVero細胞において行った[Selin et al. (1994) J. Exp. Med. 179:1933-1943; Selin et al. (1998) J. Exp. Med. 188:1705-1715]。全ての動物感染のために、組織培養上清からの未精製のワクシニアウイルスを使用した。
【0098】
ワクシニアウイルス感染
6〜8週齢の雄C57BL/6(B6)マウスには、200μgの試験抗体および/または1mgのCI-1033のPBS溶液を腹腔内注射した(i.p.)。抗体は一度だけ投与したが、CI-1033は、実験の6〜10日の期間毎日与えた。抗体および最初のCI-1033投与の6時間後に、マウスを4×104 PFU(プラーク形成単位)のワクシニアウイルス鼻腔内に感染させた。毎日マウスの重量を測定し、感染の症状を観察した(1=正常、2=乱れた毛皮;3=2プラス背中を丸めた姿勢;4=3プラスほとんど移動しない;5=4プラスの無気力な最小限の応答)。ウイルス力価は、示した時点で決定した。
【0099】
RNase保護サイトカインアッセイ法
ワクシニアウイルスでの感染の種々の日数後に屠殺したそれぞれのマウスからの肺の一方の葉を、18ゲージ針を通してRLT緩衝液(Qiagen, Valencia, CA)中で破壊した。可溶化液を、QIAshredder(商標)モジュール(Qiagen)を使用してさらにホモジナイズして、製造業者の説明書に従ってRNeasyミニ(商標)・キット(Qiagen)を使用して精製した。同量の総RNA(2μg/マウス)をそれぞれの時点で(4、6、および8日)3匹のマウスからプールした。6μgのRNAを、製造業者の説明書に従ってmCK-2b(商標)テンプレートセット(BD Biosciences-Pharmingen)から作製したプローブとハイブリダイズさせ、保護された断片をDNAシーケンシング・ゲルで分解した。ゲルをX線フィルムに曝露することによってオートラジオグラムを作製した。
【0100】
細胞内サイトカイン染色
単個細胞浮遊液を、種々の方法で処置した感染マウスの脾臓から調製し、0.84%のNH4Cl溶液を使用して溶解することによって赤血球を除去した。ワクシニアウイルス特異的な、IFN-γ産生性の、CD8+ T細胞を、ワクシニアウイルス感染線維芽細胞(MC57G)、非感染対照MC57G線維芽細胞、またはプレートに結合した抗CD3(145-2C11)mAbでインビトロで刺激した後に、5μg/mlでCytofix/CytopermキットPlus(商標)(GolgiPlug(商標), BD Biosciences-Pharmingen)を使用してFFCによって検出した。
【0101】
簡単には、MC57G細胞を5×106 PFUのウイルスで2h間インキュベーションすることによってワクシニアウイルスのWR株を感染させた(M.O.I(感染効率)= 1.2)。その後、実験マウスからの1〜2×106の脾細胞を、96ウェルプレートにおいて2.5×105ワクシニア感染MC57G(VV-MC57G)細胞または非感染MC57G細胞と共に、10 U/mlヒト組換えIL-2(BD Biosciences)、および0.2μlのGolgiPlug(商標)試薬の存在下において37℃で5時間インキュベートした。全てが最近活性化されたCD8+ T細胞を検出するための対照培養には、抗CD3 mAbを含めた。これらの培養において、脾細胞を培養ウェルの底に被覆した精製した抗マウスCD3ε mAb(145-2C11)(5μg/mlの濃度で)と共にインキュベートした(上記で記載されている同じ条件下で)。
【0102】
100μlのFFC緩衝液(HBBS(ハンク平行塩溶液)、2%のFCS、0.1%のNaN3)を含む96ウェルプレート内の1μLのFcブロック(商標)試薬(2.4G2 mAb)と共にプレインキュベーションした後、細胞をCD8α(mAb 53-6.7)およびCD44(mAb IM7)に特異的な蛍光色素ラベルしたmAbを組み合わせて染色した(20分、4℃)。その後に、細胞の固定および透過化処理を行って、抗IFNγ mAbを細胞内に到達させた(XMG1.2; BD Biosciences)。新たに染色された試料は、Becton Dickinson FACS CaliburおよびCellQuestソフトウェア(San Jose, CA)を使用して解析した。
【0103】
痘瘡プラークおよびコメット形成に対するCI-1033の効果の試験
コンフルエントなBSC-40細胞の単層を種々の濃度のCI-1033のRPMI +2%のFBS(RPMI-2%)溶液で3回、室温で30分間前処理するか、またはモックを同一の培養条件下で前処理した。7つの濃度のCI-1033を評価して、CI-1033の濃度につき3つのモック前処理対照を7つの個々の6ウェル組織培養プレートで行った。単層には、痘瘡株ソライメン(Solaimen)の懸濁液を感染させ、その結果それぞれのウェルで50プラークが観察された。プレートを6%のCO2雰囲気において35℃で1時間インキュベートし、単層の均一な感染を確実にするために15分の間隔で揺らした。種菌を除去し、単層を1×RPMI-2%でリンスした。単層をCI-1033を含まないか、または適切な濃度で含む培地で覆い、6%のCO2雰囲気において35℃でで4日間インキュベートした。プラークを免疫組織化学的染色によって解析した。コメットは、大きなプラークとコメット型に結合した2つの連続する小さなプラークよりも大きいものとして定義した。異なる濃度のCI-1033の存在下におけるプラークおよびコメットの数をウィルコクソン順位和検定を用いて対照プレートと比較した。7つのランダムに選択したモック処置したウェル(各々の7枚のプレートから選択したもの)を比較のために使用した。
【0104】
実施例2
SPGFのEGF様ドメインは、細胞増殖および/または細胞生存を刺激する
図2は、ヒトおよびマウスのエピレギュリンのアミノ酸配列の整列、並びにそれぞれオルソポックスウイルス相同分子種D1L、痘瘡、ラクダ痘、ワクシニア、およびサル痘ウイルスに由来するCMP11R、C11R、およびD3Rを示す。哺乳類タンパク質と〜30%の同一性を有するウイルスタンパク質間の高い相同性(〜90%以上)に留意されたい。EGF様ドメイン自体の中では、ウイルスタンパク質配列の事実上の同一性が明らかである(図2)。51残基のEGF様ドメインでは(D1L(SEQ ID NO:1)およびD4R(SEQ ID NO:2)の残基40〜90)、D4Rは、3残基だけがワクシニア相同分子種と異なる(C11R;SEQ ID NO:4)。大痘瘡インド株由来のD1L(図3A;SEQ ID NO:1)は、大痘瘡バングラデシュ株由来の対応する分子の(図3B;SEQ ID NO:2)D4Rとは2つのアミノ酸だけが異なる。
【0105】
一連の機能研究をrecSPGF、recEPI、およびrecEGFで行った。以下の実験によって示されるように、recSPGFは、細胞増殖または細胞生存を刺激する。ヒト初代線維芽細胞(SC-J)を2%のウシ胎児血清(FBS)を含む培地中で6時間不足(starve)させ、次いで種々の濃度のrecSPGF、recEPI、またはrecEGFで18時間刺激した。細胞周期解析は、ヨウ化プロピジウム染色FFCを使用して行った。FFCデータから、細胞周期のS期の細胞の割合をそれぞれの試料について算出し、リガンド濃度の関数としてプロットした(図4)。recSPGFは、recEGFまたはrecEPI(S期に誘導される細胞の最大のパーセントによって評価したもの)よりも低い能力を示したが、recSPGF活性は、その他の成長因子よりも低いモル濃度でプラトーに達した。
【0106】
図5は、SPGFが初代ヒト・ケラチノサイト(N細胞)およびヒト線維芽細胞(R2F)の両方の長期の成長因子であることを示す。細胞を10%のFBS(「完全」)を含む標準的な組織培養培地において、または2%のFBSおよび示した濃度のrecSPGF(「SPGF」)またはrecEGF(「EGF」)を含む組織培養培地において7日間培養し、合計生細胞のカウントを毎日行った。データは、1日あたりの平均集団倍加として表してある。
【0107】
実施例3
SPGFは、Erb-B1受容体と結合する
EGFRの4つのサブタイプがある:erb-B1、erb-B2、erb-B3、およびerb-B4。どのerb-B受容体がSPGFに結合するかについて決定するために、2つの結合実験と、MB453(erb-B1-、-B2+.-B3+、-B4+)およびMB468(erb-B1+、-B2-、-B3+、-B4-)上皮細胞株を使用してビオチン化されたrecSPGFによって行った。第1の実験は架橋解析であった。MB453またはMB468細胞をビオチン化されたrecSPGFと共にインキュベートした。recSPGFとerb-B受容体の架橋は、架橋剤BS3で行った。次いで、可溶化液を調製して免疫沈降に供し、これをHRPO結合ストレプトアビジン(「Sav-HRPO」)(図6、上段パネル)で、または各々のerb-B受容体-特異抗体(図6、中段パネル)でのウエスタン・ブロッティングによって行った。これらの2つの細胞株のerb-B受容体1〜4の発現は、図6の下段パネルに示した表に示してある。図6の上段パネルは、recSPGFがerb-B1のみに検出可能な程度に結合することを証明する。
【0108】
化学的架橋の結果と一致して、図7は、抗erb-B1 mAbが、MB468上皮細胞に対するrecSPGFの結合をブロックすることを示す。MB453細胞は、erb-B1を発現しないので、これらに対するrecSPGFの結合は検出されなかった(図7)。
【0109】
結合阻害実験では、erb-B1に対するSPGFの結合は、EGFよりもわずかに弱いが、エピレギュリンよりも強いことを示す(図8)。
【0110】
EGFR1発現細胞に対するrecSPGF結合の親和性の測定は、スキャッチャード解析によって得られた(図9)。MB468細胞を96ウェルマイクロタイター組織培養プレートのウェルにまいた。125IラベルされたrecSPGFを種々の濃度で添加して、プレートを4℃で5時間インキュベートした。親和性(Kd)および受容体数は、実施例1に記載したように算出した。この解析では、MB468細胞あたり約5.5×104高い親和性(Kd = 0.14nM)のerb-B1受容体および8.8×106の低い親和性(Kd =258 nM)のerb-B1受容体があることを示した。
【0111】
erb-B1受容体は、刺激されていない細胞をerb-B1(図10A)に特異的な抗体で染色したときに得られるリニアな蛍光のほとんど均一なパターンによって証明されるとおり、通常上皮細胞の表面にかなり一様に分布した。しかし、recSPGF、recEPI、またはrecEGFの添加の10分後に、受容体は迅速に内部に取り入れられた(それぞれ、図1OB、1OC、および図10D)。
【0112】
recSPGFによって活性化されるerb-B1の迅速インターナリゼーション、およびチロシン・プロテインキナーゼ・リン酸化がアクチンおよびエズリンを含む細胞突出の形成を経てIMVの侵入を活性化する能力[Locker et al. (2000) Mol. Biol. Cell 11:2497-2511]は、両方とも細胞に対する痘瘡ビリオンの侵入を容易にするために貢献する。天然には、侵入前の細胞表面に対する痘瘡ビリオンの最初の結合は、少なくとも一部において、SPGF-erb-B1の相互作用を経ることができる。細胞に対するビリオンの侵入を可能にすることに加えて、上記の活性化イベントは、宿主細胞をより効率的なウイルス複製「工場」にする。
【0113】
実施例4
SPGFのEGF様ドメインは、細胞におけるタンパク質チロシン・リン酸化を刺激する
RecSPGFは、EGF受容体およびこれらの細胞基質のリン酸化を刺激した。無処置(-)またはrecSPGF(「SPGF」)、recEPI(「EPI」)、もしくはrecEGF(「EGF」)で処置したHeLa細胞からの細胞全体の可溶化液を4G10抗ホスホチロシン・モノクローナル抗体でのウエスタンブロット法(WB)に供した(図11A)。加えて、HeLa細胞可溶化液を抗EGFRポリクローナル抗体で免疫沈降(IP)し、抗ホスホチロシン抗体でのWBに供した(図11B)。EGFR自体およびその種々の細胞基質の両者のチロシン残基は、試験した3つの成長因子によって、同一でない場合でも、同様の様式でリン酸化された。
【0114】
実施例5
SPGFのEGF様ドメインによるタンパク質チロシン・リン酸化の刺激は、キナゾリンに基づいた化合物によって阻害される
SPGFで刺激される活性に対するいくつかのキナゾリンに基づいたチロシンキナーゼ阻害剤の効果を評価するために、一連の実験を行った。図12は、 3つの4-アニリノキナゾリンチロシンキナーゼ阻害剤(PD 153035、PD 168393、およびCI-1033)の化学構造を、同様に一般的な4-アニリノキナゾリン構造を示す。erb-B1受容体キナーゼ・ドメイン単独での、およびそのような阻害剤の1つとの複合体の構造は、これらの4-アニリノキナゾリンがどのようにキナーゼ・ドメインのATP結合ポケットと結合するかを示している[Stamos et al. (2002) J. Biol. Chem. 277:46265-46272]。図12に示すように、PD153035およびPD168393は、同じ4-(3'-ブロモ-アニリン)環を有するが、キナゾリン環に付着されるこれらのR3およびR4基が異なる。特に、PD168393は、erb-B1 Cys773をアルキル化することができる6位にアクリルアミドを有し、その結果、阻害剤は、1:1のモル比で不可逆的にerb-B1と結合する。チロシンキナーゼ活性なerb-B2およびerb-B4分子は、それぞれCys784およびCys778に相当するシステイン残基を有しており、修飾のためにターゲットすることができる。対照的に、PD153035は、主に疎水性の力を経て可逆的な様式で結合する。CI-1033は、PD168393同様に、6位(R3)にアクリルアミド付加物を有し、したがって、PD168393同様、Cys773を経て不可逆的にerb-B1と結合する。CI-1033は、加えて、7位(R4)に可溶性のモルフォリン側鎖を有する。CI-1033は、erb-B1、erb-B2、およびerb-B4に対して、それぞれ0.8、19、および7nMのIC50値を示す[Allen et al. (2002) Sem. Oncol. 29:11-21]。
【0115】
第1の実験では、2つのキナゾリンに基づいた化合物(PD168393およびPD153035)およびピリドピリミジンに基づいた化合物(PD158780)を、これらがチロシン・リン酸化を阻害する能力について試験した。全3つの化合物は、Calbiochem, San Diego, CAから得られ、Calbiochemのカタログでは、これらの化学構造を示している。HeLa細胞を24ウェル組織培養プレートのウェルにまき、阻害剤化合物を50nMの終濃度に添加した。細胞を30分間培養し、recSPGF(50ng/ml)をネガティブ対照試料(図13において(-)と示す)以外の全てのウェルに添加した;図13に(+)として示した試料は、recSPGFを受けたが、阻害剤を受けさせなかった。次いで、細胞をさらに10分間37℃で培養し、次いで洗浄した後に溶解した。細胞可溶化物を10%のSDS-PAGEによって分離して、SDS-PAGEゲルをPVDF膜にブロットした。膜を4G10抗ホスホチロシンmAbに曝露し、その結合をHRPO結合抗マウスIgG2bポリクローナル抗体によって検出した。膜をECL(化学発光の増強)によって発色させた。図13のデータは、recSPGFによって活性化されるチロシン・リン酸化が全3つの化合物(PD168393、PD153035、およびPD158780)によって阻害されることを示す。
【0116】
recSPGFを媒介した細胞増殖および/または細胞生存の増大に対するチロシンキナーゼ阻害剤の効果を試験した。図14Aに示すように、種々の濃度のrecSPGFでヒト線維芽細胞を一晩刺激した後、S期に入っている細胞の数は、8%に近づいた。50nM濃度の示した阻害剤で37℃において1時間細胞を前処理すると、このDNA合成の増加をブロックする。図14Bは、erb-B1のチロシン・リン酸化(150kDバンド)並びにそのリン酸化のさらなる基質(120kD、80kD、60kDおよび55kD)を阻害したものと同じ化合物が、HeLa細胞においてrecSPGFによって活性化されたことを示す。SrcファミリーPTK特異的阻害剤PP2は、10μMで使用したときでも、これらのerb-B1基質のリン酸化をブロックする能力をほとんど有さなかった。対照的に、50nMの濃度でPD168393またはCI-1033を添加すると、主にrecSPGFでトリガーされるリン酸化を防止したが、PD153035で観察される効果は、部分的なものだけであった。これらの結果は、キナーゼ活性の可逆的阻害剤が、試験される50nM濃度において不可逆阻害剤と比較してタンパク質チロシン・リン酸化のブロックにあまり効率的ではないことを示唆する。チロシンキナーゼ・リン酸化の同様の阻害は、ワクシニアウイルス成長因子(VGF)によって活性化されるリン酸化でも観察された(データ示さず)。
【0117】
図10Bは、recSPGFと37℃で15分間インキュベーションする前および後に、免疫蛍光顕微鏡観察によって検出されるヒトHeLa上皮細胞のerb-B1受容体の分布を示す。成長因子の非存在下では、erb-B1の分布は、主に膜に限定される。対照的に、SPGF曝露後では、erb-B1分子は迅速にインターナライズされて、点状の細胞内蛍光の凝集として現れる。チロシンキナーゼ阻害剤がSPGFの能力に影響を及ぼしてerb-B1をダウンモジュレートするかどうか判断するために、A431細胞を阻害剤で前処理し、ビオチン化されたrecSPGFと共にインキュベートし、次いで固定した。次いで、細胞表面に結合したerb-B1リガンドをストレプトアビジン-PEによって視覚化し、蛍光をFFC解析によって定量化した。図14Cは、阻害剤の非存在下では、MFIは、〜110であったが;対照的に、不可逆的erb-B阻害剤は、MFIを〜3倍(300〜350)に増大し、可逆的erb-B1阻害剤PD153035は、MFIを240に増大したことを示す。srcファミリーキナーゼ阻害剤PP2は、わずかにerb-B1表面のコピー数に影響を及ぼすだけであった。
【0118】
erb-B受容体は、2つの経路によって除去される:1)リガンド依存的なエンドサイトーシス、およびc-Cblユビキチンリガーゼ機構を含む分解[Levkowitz et al. (1999) Mol. Cell 4:1029-1040]、並びに2)ストレスで誘導されるerb-B受容体と結合したシャペロンおよび関与するプロテアソームのプロテイナーゼのシャフリング[Xu et al. (2001) J. Bio. Chem. 276:3702-3708]。最近の研究は、CI-1033によるerb-B2キナーゼの阻害が、第2のプロセスを経てerb-B2のダウンレギュレーションを促進することを示す[Citri et al.(2002) EMBO J. 21:2407-2417]。しかし、CI-1033がerb-B1の表面発現を増強することを考えると、この知見は、リガンド特異的なSPGF erb-B1発現に対するCI-1033の効果は、構成的erb-B2発現に対するものと類似しないことを示唆する。
【0119】
4-アニリノキナゾリンがc-Cblのerb-B受容体との相互作用に影響するかどうかを試験するために、阻害剤を前処理したか、または無処置のrecSPGF刺激したHeLa細胞を溶解し、可溶化液を抗erb-B1抗体で免疫沈降した。その後、erb-B1分子の発現を定量化するために抗erb-B抗血清で、またはerb-B1結合に対する化合物の効果を評価するために抗c-Cbl抗体でのいずれかでウエスタン・ブロッティングを行った。図14Dに示すように、刺激されていない細胞(-)と比較して、recSPGFでトリガーされたHeLa細胞(+)は、迅速な細胞内分解のために合計erb-B1が少なくなる。FFC解析(図14C)と一致して、PP2は、このプロセスに対してほとんど影響を有さなかった。他方、PD153035、PD168393、およびCI-1033の前処置では、recSPGFの添加にもかかわらず、免疫沈降される合計erb-B1タンパク質が増大し;recSPGF刺激されていない対照細胞から得られたもの以上にerb-B1の数倍の増加を示した。後者の結果は、阻害剤によるerb-B1の構成的インターナリゼーションまたは分解の封鎖を意味する。さらに重要なことに、recSPGFで誘導されるc-Cblのerb-Bとの結合は、4-アニリノキナゾリンによってブロックされたが、PP2によってブロックされなかった。c-Cbl/erb-B1複合体形成の減少は、平行したc-Cbl免疫沈降およびウエスタン・ブロッティングによって示される合計細胞c-Cblレベルの減少に対して二次的であった。その代わりに、これらのerb-Bキナーゼ阻害剤は、SPGFの結合後の誘導性のc-Cblのerb-B1との結合を防げた。
【0120】
実施例6
チロシンキナーゼ阻害剤のインビボでの抗ウイルス効果
ワクシニアWRのVGF遺伝子の破壊は、インビボでワクシニアウイルスの病原性を減少させ、ウイルス増殖を〜10,000倍まで緩徐化することを示した[Buller et al. (1988) J. Virol. 62:866-874]。上に示したように、CI-1033は、SPGF刺激されたerb-B1で駆動される細胞増殖または生存、受容体を媒介したチロシン・リン酸化、インターナリゼーション、および分解をブロックするので、erb-B1キナーゼ阻害剤は、インビボでのオルソポックス成長因子活性を減らすことができるようであった。この可能性を試験するために、致死的に鼻腔内にワクシニアWRを与えたB6マウスの臨床経過に対するCI-1033の効果を調査した[Chen et al. (2001) Nat. Immunol. 2:1067-1076]。図15Aに示したように、全ての無処置の動物は、劇症性の急性肺炎による感染の7日後までに死んだ。対照的に、感染の6h前に開始して毎日50mg/kgでCI-1033をi.p.投与すると死を防止した。本療法は、ワクシニア細胞内成熟ウイルス粒子(IMV)を中和することが既知の抗L1RワクシニアウイルスmAb(7D11)の一回の200μgのi.p.用量の投与と同程度に有効であった[Hooper et al. (2000) Virol. 266:329-339]。CI-1033と抗L1Rの併用投与でも、保護された。感染後の動物の臨床モニタリングでは、抗L1R mAbまたはCI-1033処置が有意に総合的症状を減少させたが、併用療法ほど効率的ではなかったことを示した(図15B)。
【0121】
動物のコホートを感染の6日後に屠殺し、これらの肺を検査した。処理群間の総肺重量の違いを図15Cに示してある。未処理の動物では、多発性出血、および正常な、非感染性のB6マウスの約2倍の重量を有する浮腫状の肺を有した。この病状は、抗L1RまたはCI-1033単独での処置によっていくらか減弱されたが、CI-1033プラス抗L1Rの併用によって劇的に改善された。病理学的な所見と一致して、肺におけるワクシニアウイルスの力価は、8日目にアッセイしたときに、抗L1RまたはCI-1033単独によってやや減少されたが、併用治療によっては本質的に除去された。無処置の動物は、全て8日までに死んだので、7日のウイルス肺力価を比較のために使用した。また、図15Dは:(i)抗SPGF mAb(13E8)(これは、VGFと交差反応する(出願中のU.S. Application Serial No. 10/429,685を参照されたい))での免疫療法;および(ii)抗L1Rプラス抗SPGF mAbsでの二重免疫療法の結果を図示する。抗L1RプラスCI-1033処置で観察されるウイルス力価の、さらなる2つの経過記録の減少(抗L1R mAbプラス抗SPGF mAb処置と比較して)は、mAbの可逆的な結合と比較して、CI-1033の不可逆的結合の結果によるであろう[Fry et al. (1998) Proc. Natl. Acad. Sci. USA 95:12022-12027]。阻害剤の不可逆的結合により、阻害剤の可逆的結合よりも完全なerb-B1経路の封鎖が生じるのであろう。
【0122】
併用した抗L1R mAbプラス抗SPGF mAb免疫療法実験(出願中のU.S. Application Serial No. 10/429,685号を参照されたい)では、ウイルス力価が感染の後6日まで減退しなかった。この緩徐な時間経過は、抗体のウイルス中和効果ではなくむしろウイルスに対して適応性T細胞応答が発生していることを示唆した。図示はしていないが、同様の動態は、抗L1R mAbプラスCI-1033の併用療法によっても観察された。これらの知見は、CI-1033が、単独でまたは抗L1R mAbと併用して、T細胞応答を増大するであろうことを示唆する。この概念を試験するために、肺におけるT細胞で調節されるサイトカイン産生を感染の4、6、および8日後にRNase保護解析によって試験した。図16Aに示したように、CI-1033を単独で与えられたマウスでは、インターロイキン-1β(IL-1β)、インターロイキン-1受容体アンタゴニスト(IL-1Ra)、およびIFN-γ mRNAのレベルが、無処置のマウスと比較して増大し、6日にピークに達した。たとえば、IFN-γ RNAのレベルは、6日目で、対照感染動物のものと比較して、CI-1033処置したマウスの肺のよりも4.1倍大きかった。しかし、インターロイキン6(IL-6)、インターロイキン10(IL-10)、インターロイキン12(IL-12)、またはマクロファージ遊走阻止因子(MIF)mRNAのレベルに対する効果は、CI-1033を与えらたマウスの肺においても全く見られなかった。
【0123】
肺サイトカインのレベルと一致して、VV-MC57G細胞に対する全身性の抗原特異的T細胞応答は、脾臓のCD8+CD44+ T細胞の細胞内IFN-γ産生によって評価すると、対照の無処置のマウスと比較して、CI-1033処置したマウスにおいて実質的に増強された(図16B)。VV-MC57G細胞によって活性化されるIFN-γ産生CD8+CD44+ T細胞の割合は、感染8日後に10倍に増大した。この増加は、VV-MC57G細胞によって活性化されるIFN-γ産生CD8+CD44+ T細胞の絶対数の21倍の増加に対応した(1.85×105対8.5×103細胞);この効果は、無処置のワクシニアウイルス感染マウスにおけるリンパ球減少によるものであった。同じ実験において、抗L1Rおよび抗L1R + CI-1033処置したマウスについて、それぞれ22.7および48倍の増大が観察された。抗L1R処置により、抗L1R + CI-1033の二重療法よりも肺におけるIFN-γサイトカインRNAを生じたことから、前者の処置での持続的なウイルス負荷を示しているのであろう(図15D)。マウスの抗L1Rおよび抗L1R + CI-1033処置後のIFN-γ産生CD4+CD44+ T細胞の比率に、よりわずかな、しかし有意な増大が見られた(データ示さず)。
【0124】
実施例7
VGFに特異的な抗体によるワクシニアウイルスに対するインビボ免疫の増強
同時継続中のU.S. Application No. 10/429,685では、CI-1033と同様に、痘瘡SPGF D4Rに特異的で、かつVGFと交差反応を起こすmAb(13E8)は、抗L1R mAbの保護効果を増強したことが示された。本明細書において、CI-1033(実施例6を参照されたい)に関し、mAb 13E8がインビボでワクシニアウイルスに対するT細胞免疫を増強し、インビボで抗L1R mAbによって得られるT細胞免疫の増強を増大することが示される。
【0125】
ワクシニアウイルスは、IFN-γ産生T細胞によってインビボで制御されるので、そのレベルは、感染の6〜8日後にピークに達し[Ruby et al.(1991) Lymphokine Cytokine Res. 10:353-358]、13E8および抗L1R mAbで見られるウイルス減少の動態(同時継続中のU.S. Application No. 10/429,685)は、適応免疫が関与することを示唆した。従って、CD8+ T細胞応答を、ワクシニアウイルスに感染したマウスを以下によって処置した7日目に検査した:(a)対照1A3 mAb、(b)13E8 mAb、(c)抗L1R mAb;または(d)13E8および抗L1R mAbsの組み合わせ。VV-MC57Gスティミュレーター細胞で、または抗CD3 mAbで(これらは最近活性化された全てのT細胞を刺激する)、5h間インビトロで刺激した新たに単離された脾臓T細胞において、細胞内IFN-γレベルを解析した。実験では、ナイーブ非感染マウスと比較して、以下の順序で、ワクシニアに感染した処理群のCD44+CD8+ T細胞のIFN-γ反応を増大することを示した:1A3 mAb(対照)、13E8 mAb、抗L1R mAb、および抗L1R mAb + 13E8 mAb(図17C)。この実験およびその他の実験において、脾臓は、抗L1R mAbおよび抗L1R mAb + 13E8 mAb群において拡大し、その結果、たとえば抗L1R mAb + 13E8 mAbを組合せて処置したマウスの脾臓あたりのワクシニアウイルス特異的IFN-γ産生細胞の総数は、1A3処置したマウスよりも11倍高かった(0.1が×印を付ける3.3±1.4×105対0.3±0.1×105、群あたりn=3)。
【0126】
マウスの抗L1Rおよび抗L1R + 13E8 mAb処置後のIFN-γ産生CD4+CD44+ T細胞の比率には、より少ないが、しかし、有意な増大が見られた(データ示さず)。
【0127】
抗L1R + 13E8 mAb-処置マウスにおける活性化されたIFN-γ産生脾臓のT細胞のレベルの増強は、肺感染と戦うために肺に移動する能力がある細胞の貯蔵所を示唆し;CD44(T細胞刺激によってアップレギュレートされる活性化マーカー)は、炎症部位にホーミングする白血球に関与するリサイクル可能なヒアルロナンの受容体である点に留意されたい[DeGrendele et al. (1997) Science 278:672-675]。この可能性を試験するために、肺を非感染の、または感染7日、およびmAb処置したマウスから集めて、組織切片のヘマトキシリン-エオジン染色によって検査した。肺胞腔が容易に識別可能であり、細気管支の上皮は無処理であった非感染マウスからの肺と比較して、感染した対照mAb処置したマウスからの肺は、壊死性の細気管支上皮および重篤な肺胞浮腫によって特徴づけられた。これは、ワクシニアウイルスに感染した肺の重量が、非感染肺の約2倍であることを反映した(.35 ± .04 g対 .21 ± .01g、n = 3)。肺胞腔は、これらが好酸性物質で満たされたという点で、除去され;あるとしても、わずかな実質細胞の浸潤が存在した。細胞浸潤の程度は、13E8 mAb処置マウスにおいて幾分増大され、抗L1R mAb処置マウスにおいてより増大され、および組み合わせたmAbで処置したマウスにおいて最も劇的に増大された。二重mAb処置群において、肺胞腔は、開放性であり、細気管支上皮は、正常であった。細胞浸潤する細胞は、大部分がCD8+ T細胞であった。3つの抗L1R mAb処置したマウスのプールされた肺から単離された白血球は、25%のCD8+および5%のCD4+ T細胞であり、抗L1R mAbプラス13E8 mAb処置したマウスから単離されたものは、34%のCD8+および8%のCD4+ T細胞であった。従って、非中和性13E8 mAbは、脾臓および肺のT細胞応答の増大に付随して、ワクシニアウイルスのクリアランスを増強する。
【0128】
先天性免疫、いわゆる急性期反応の全身性の炎症性の成分は、生物体の統合性が破壊されたときに迅速に誘導される[Yoo et al. (2003) Proc. Natl. Acad. Sci USA 100:1157-1162]。上皮細胞を含むセンチネル細胞は、このプロセスの間に、IL-1βおよび/またはIL-6を産生する[Schluns et al. (1997) J. Immunol. 1158:2704-2712]。13E8 mAbによるerb-B1刺激の遮断は、ウイルスによってコードされる分子による、その後のサイトカイン調節不全、およびそれに伴う免疫抑制の回避を補助するであろう[Diehl et al. (2002) Mol. Immunol. 39:531-536; Tanaka et al. (2001) J. Mol. Cell Cardiol. 33:1627-1635]。さらに、EGFおよびTGFα(両方ともerb-Bリガンド)は、インビボで浸出液を誘導することができる[Ohmura et al. (1990) Cancer Res. 50:4915-4917]。SPGFおよびVGFなどのELGFは、類似の病態に寄与している可能性があり、抗ELGF抗体(たとえば、l3E8 mAb)によるその阻害は、このようなオルトポックスウイルス感染に対する抗体のさらなる治療活性を構成すると考えられる。
【0129】
実施例8
タンパク質チロシンキナーゼ阻害剤は、痘瘡ウイルスの細胞内形成および細胞間伝播を阻害する
痘瘡ウイルス増殖に対するCI-1033の効果を検査するために、BSC-40細胞のコンフルエントな単層を、種々の濃度のerb-B阻害剤の存在下または非存在下において痘瘡株ソライメン(Solaimen)約50プラーク形成単位に感染させて、インビトロで4日間培養した。図18Aに示すように、CI-1033の濃度を増大すると、個体のプラークのサイズおよびコメット形成を激減させるが、プラーク数に対しては最小限の効果を有した。プラーク・サイズおよびコメット形成に対する効果は、滴定可能であり(図18A、18B、および18C);CI-1033(10μM)の最も高い濃度では、極めて小さなプラークがかすかに見えるだけであった。それぞれの濃度でのプラークの総数は、7つのランダムに選択した対照ウエルと比較したときに有意に異ならなかった(図18Aおよび18B)。
【0130】
コメットは、細胞外外膜ウイルス(EEV)形成および播種を示す[Payne (1980) J. Gen. Virol. 50:89-100]。図18Cは、500nM以上のCI-1033濃度におけるコメット(p<0.05)の数の統計学的に有意な減少を示す。これらの集約した結果は、CI-1033が初感染ではなく、むしろEEVのウイルス形態形成をブロックすることを示唆する。特に、EEVの放出は、阻害に感受性である。
【0131】
プラーク・サイズに対してあまり重大ではない効果が見られ、より高い濃度のCI-1033で定性的に見えることから、細胞結合内外膜ウイルス(CEV)形成に対する効果を示している。オルトポックスウイルスは、出現するために微小管およびアクチンフィラメントの両方を使用し[Hollinshead et al. (2001) J. Cell Biol. 154:389-402; Smith et al. (2002) J. Gen. Virol.83:2915-2931]、CEVは、細胞からそれ自体が駆出するためのアクチン尾部を誘導する。erb-B1活性化により、アクチンミクロフィラメント系を再編成するので[Lynch et al. (2003) J. Biol. Chem. 278:21805-21813]、これは、細胞結合ウイルスおよび細胞外外膜ウイルスの両方の放出をブロックして、二次的なウイルスの細胞間の伝播を遅延させ、それ故、個体のプラークのサイズを制限している可能性が高いと思われる。
【0132】
EEVを経たウイルスの広範囲の伝播(コメット形成の阻害で示すとおり)に対するより重要な関連した効果は、さらなる妨害機構の可能性を示唆する。erb-Bエンドサイトーシスはまた、アクチン細胞骨格にリンクしているので[Lynch et al.]、erb-B1キナーゼ阻害も同様に、この機構を経て表面EGFR発現を維持するために役立っているかもしれない。細胞のerb-Bリガンドまたはその他の受容体を経たerb-B1のトランス刺激によるerb-B活性化、並びにその下流のポックスウイルス出現のために使用される細胞骨格エレメントに対する効果をブロックするCI-1033の能力は、抗SPGF mAbでは観察されないプラーク形態形成に対する重大な効果の根拠となる可能性が高い(データ示さず)。
【0133】
要するに、併用免疫療法/化学療法は、ウイルス力価の減少およびT細胞免疫の刺激に非常に有効である。本発明は、いずれの特定の作用機序によっても限定されないが、後者が抗炎症性のウイルス産物の生成、並びにウイルス成長因子に依存的な上皮細胞におけるウイルスのDNA複製後のラウンドを防げた結果であろうと推測することができる[Alcami (2003) Nat. Rev. Immunol. 3:36-50; Seet et al. (1998) Ann. Rev. Immunol. 21:377-423]。抗ウイルス化学療法のための標的としての宿主細胞シグナリング経路の遮断の使用は、典型的には、病原体それ自体に対して向けられたその他のアプローチとは別である。宿主にターゲットされたストラテジーの1つの利点は、薬物抵抗性を発生し得ないことである。さらに、抗SPGF mAbsとは異なり、その他の密接に関連した病原性因子(すなわち、VGF対SPGF)との交差反応性が保証されない場合に、erb-Bチロシンキナーゼ阻害剤は、種々のerb-Bタンパク質チロシン・ファミリー数に対して免疫学的に異なったリガンドを潜在的にブロックすることができる。たとえば、VGFおよび粘液腫ウイルス成長因子(MGF)は、異なったerb-B受容体に結合するので[Tzahar et al. (1998) EMBO J. 17:5948-5963]、このより広範な標的活性が関連している。
【0134】
CI-1033の単独投与が、オルソポックス(たとえば、天然の、またはバイオテロリストが蔓延させた天然痘)による「実際の」感染を遅延させるために十分であろう可能性が高いと思われ、オルソポックス粒子の吸入の用量は、本研究において使用したものよりも有意に少ない可能性が高い。そのCI-1033は、T細胞応答を増大すると共に、感染の総合的症状も減少させることから:(a)ワクシニアワクチン接種と併用した免疫刺激薬として;または(b)ワクチン接種の合併症の治療における使用も示唆される。
【0135】
ポックスウイルスおよびこれらのerb-Bに対する成長因子に加えて、B型肝炎ウイルスおよびEBウイルスも、erb-B1転写を調節不全にする産物をコードする遺伝子を有する[Menzo et al. (1993) Virol 196:878-882; Miller et al. (1995) J. Virol. 69: 4390-4398]。さらに、ニワトリ赤芽球症ウイルスなどのRNA型腫瘍ウイルスも、erb-B1シグナリングを利用する[Adelsman et al. (1996) J. Virol. 70:2533-2544]。さらに、機能的なEGF受容体は、宿主細胞の効果的なレオウイルスおよびヒトサイトメガロウイルス(CMV)感染症に必要なことが報告された[Strong et al. (1993) Virol. 197:405-411; Wang et al. (2003) Nature 424:456-461]。これらの知見は、ひとまとめにして、複数のウイルス感染が、erb-Bを媒介したシグナル・ネットワークに結合しているであろうことを示唆する。erb-B受容体を構成的にまたは誘導性に発現する上皮細胞は、体における生得的なセンチネル細胞系の成分である[Yoo et al.(2003) Proc. Natl. Acad. Sci USA 100:1157-1162]。erb-B阻害剤は、オルトポックスウイルス感染よりも感染症においてより広範に適用することができる。ウイルスの病原性因子によって利用される細胞シグナリング経路の化学阻害剤の開発により、一般に感染症の制御に対して新たなアプローチがもたらされるであろう。
【0136】
本発明の多くの態様を記載した。それにもかかわらず、本発明の精神と範囲から逸脱することなく、種々の修飾を行ってもよいことが理解されるであろう。従って、その他の態様も、請求項の範囲内にある。
【図面の簡単な説明】
【0137】
【図1】マウスのエピレギュリン(「Epi」)の上皮細胞成長因子(EGF)様ドメイン、ヒトEGF、および大痘瘡バングラデシュ株天然痘成長因子(「SPGF」)の組換え形態のクーマシーブルー染色したSDS-PAGE(ドデシル硫酸ナトリウム・ポリアクリルアミドゲル電気泳動)ゲルの写真である。これらの組換えタンパク質は、本明細書において、それぞれrecEGF、recEPI、およびrecSPGFと称される。分子量(MW)マーカーのゲルの位置は、写真の左上に示してあり、値は、MW×10-3を示してある。
【図2】ヒト・エピレギュリン(SEQ ID NO:6)およびマウスエピレギュリン(SEQ ID NO:7)、並びにそれぞれ大痘瘡インド株、ラクダ痘、ワクシニア、およびサル痘ウイルス由来のオルソポックスウイルス相同分子種D1L(SEQ ID NO:1)、CMP11R(SEQ ID NO:3)、C11R(SEQ ID NO:4)、およびD3R(SEQ ID NO:5)のアミノ酸配列の整列(最適な相同性のために)の描写である。リーダー配列および膜貫通領域に対応する領域を示してあり、EGF様ドメインで最も保存されたセグメントを枠に入れ、「EGF」をラベルした。
【図3】図3Aおよび3Bは、それぞれ大痘瘡インド株(D1L)(SEQ ID NO:1)および大痘瘡株バングラデシュ(D4R)(SEQ ID NO:2)のSPGFのアミノ酸配列の描写である。2つの配列が異なる位を太字で示して、下線を引いてある。
【図4】示した濃度のrecEGF(「EGF」)、recEPI(「EPI」)、およびrecSPGF(「SPGF」)の18時間の培養後での細胞周期のS期のヒト初代線維芽細胞(SC-J)の割合を示す線グラフである。
【図5】10%のウシ胎児血清(FBS)を含む標準的な組織培養培地中(「完全」)か、または2%のFBSおよび示した濃度のrecSPGF(「SPGF」)またはrecEGF(「EGF」)を含む組織培養培地中のいずれかでの培養の間の、初代ヒト・ケラチノサイト(「Nケラチノサイト」)および初代ヒト線維芽細胞(「R2F線維芽細胞」)の増殖を示す棒グラフである。
【図6】MB453細胞(「453」)またはMB468細胞(「468」)からの免疫沈降物のウエスタンブロットの一対の写真である。免疫沈降物は、ビオチン化されたrecSPGFと共に細胞をインキュベートすることによって調製して、架橋剤BS3に細胞を曝露し(細胞表面上のerb-B受容体に結合したビオチン化されたrecSPGF分子を架橋するため)、細胞を溶解して、および4つのerb-B受容体、すなわちerb-B1(「B1」)、erb-B2(「B2」)、erb-B3(「B3」)、またはerb-B4(「B4」)に特異的な抗体で可溶化液の一定分量を免疫沈降(「IP」)することによって調製した。2つの2回のウエスタンブロットを免疫沈降物から作製した;一方は、西洋ワサビペルオキシダーゼ複合ストレプトアビジン(「Sav-HRPO」)(図6、上段パネル)で染色し、他方は、各々のerb-B受容体-特異抗体(図6は、中段パネル)で染色した。図6の下段パネルは、MB453(「453」)およびMB468(「468」)細胞による4erb-B受容体の発現を示す。
【図7】erb-B1に特異的な阻止抗体の非存在下(白丸)および存在下(「+mAb」;黒丸)での、ビオチン化されたrecSPGF(フィコエリトリン(PE)結合ストレプトアビジンによって検出)(「SPGF」)の示された量(「ng」)のMB468細胞(「対468」)に対する結合を示す線グラフである。データは、平均蛍光強度(「MFI」)として示してあり、これは蛍光フローサイトメトリー(FFC)によって決定した。また、erb-B1を発現しないMB453細胞(「対453」)に対するrecSPGFの結合を試験した(「対453」;白三角)。
【図8】示した濃度のrecSPGF(「SPGF」)、recEPI(「EPI」)、およびrecEGF(「EGF」)がMB468細胞に対するビオチン化されたrecSPGFの結合を阻害する相対的な能力(%阻害として表した)を示す線グラフである。
【図9】MB468細胞に対するrecSPGF結合のスキャッチャードプロットである。解析は、細胞上のSPGFについて、約5.5×104高親和性受容体(「KD= 0.14nM」)および約8.8×106の低親和性受容体(「KD= 258nM」)の細胞上での存在を示した。
【図10】図10A-Dは、刺激されていないか(「(-)」;図1A)、またはrecSPGF(「SPGF」;図10B)、recEPI(「EPI」:図10C)、もしくはrecEGF(「EGF」;図10D)のいずれかで10分間刺激されたかいずれかのHeLa細胞上のerb-B1受容体(erb-B1受容体に特異的なマウス抗体およびフルオレッセインイソチオシアネート(FITC)結合抗マウスIg抗体によって検出されるもの)の細胞分布を示す一連の蛍光顕微写真である。
【図11】図11Aは、無処置であるか(「(-)」)、またはrecSPGF(「SPGF」)、recEPI(「EPI」)、もしくはrecEGF(「EGF」)で刺激したかいずれかのHeLa細胞の細胞全体の可溶化液のウエスタンブロットの写真である。ウエスタンブロットは、ホスホチロシン残基に特異的なmAb(「WB:4G10」)で染色した。分子量(MW)マーカーのゲル上の位置を写真の左上に示してあり、値は、MW ×10-3であるように示した。 図11Bは、無処置であるか(「(-)」)、またはrecSPGF(「SPGF」)、recEPI(「EPI」)、もしくはrecEGF(「EGF」)で刺激したかいずれかのHeLa細胞の細胞可溶化物の免疫沈降物のウエスタンブロットの写真である。免疫沈降物は、erb-B1(「IP:EGFR」)に対して特異的な抗体によって調製した。ウエスタンブロットは、ホスホチロシン残基に特異的なmAb(「WB:4G10」)で染色した。
【図12】図12A-12Dは、関連した原子の番号付けを示す4-アニリノキナゾリン化合物の一般的な化学構造(「一般的な4-アニリノキナゾリン」;図1D);並びに3つの特異的な4-アニリノキナゾリン化合物、PD153035(図12A)、PD168393(図12B)、およびCI-1033(図12C)の描写である。
【図13】非処理(「(-)」)であるか、またはerb-Bタンパク質チロシンキナーゼ阻害剤に対して事前に露光していないか、もしくは3つのerb-Bタンパク質チロシンキナーゼ阻害剤、PD153035、PD158780、およびPD168393のうちの1つに対して事前に露光した、いずれかrecSPGF(「SPGF」)で刺激したHeLa細胞の細胞全体の可溶化液のウエスタンブロットの写真である。ウエスタンブロットは、ホスホチロシン残基に特異的なmAbで染色した。分子量(MW)マーカーのゲル上の位置を写真の左上に示してあり、値は、MW ×10-3であるように示した。
【図14】図14Aは、前処理なし(「SPGF単独」)か、またはPD153035、CI-1033、もしくはPS168393;および示した濃度のrecSPGF(「ng/ml」)で一晩刺激;での前処理後の細胞周期のS期のヒト初代線維芽細胞(SC-J)の割合を示す線グラフである。 図14Bは、非処理(「(-)」)であるか、またはerb-Bタンパク質チロシンキナーゼ阻害剤(「(+)」)に対して事前に露光していないか、またはSrcファミリー・タンパク質チロシンキナーゼ阻害剤PP2もしくはerb-Bタンパク質チロシンキナーゼ阻害剤、PD153035、PD16839、もしくはCI-1033に対して事前に露光した、recSPGF(「SPGF」)で刺激したかいずれかのHeLa細胞の細胞全体の可溶化液のウエスタンブロットの写真である。ウエスタンブロットは、ホスホチロシン残基に特異的なmAb(「WB:P-Y(4G10)」)で染色した。分子量(MW)マーカーのゲル上の位置を写真の左上に示してあり、値は、MW ×10-3であるように示した。写真の右上の矢印は、erb-B1タンパク質チロシンキナーゼ阻害によって影響を受けたタンパク質リン酸化の位置を示す。 図14Cは、以下の後でのA431細胞上のerb-B1受容体の発現レベル(FFCによって決定されるMFIの)を示す棒グラフである:阻害剤なしでの前処理、またはPP2、PD153035、PD168393、もしくはCI-1033での前処理;およびビオチン化されたrecSPGFとの5分間のインキュベーション。 図14Dは、前処理していないか(「(+)」)、またはPP2、PD153035、PD168393、もしくはCI-1033のいずれかで前処理し、次いでrecSPGFで5分間刺激したHeLa細胞の細胞可溶化物の免疫沈降物(「IP」)のウエスタンブロットの一連の写真である。細胞を前処理しないか、または刺激していないrecSPGF(「(-)」)対照でも行った。免疫沈降物(「IP」)は、erb-B1(「EGFR」)に特異的な抗体またはc-Cbl(「c-Cbl」)に特異的な抗体のいずれかによって調製した。ウエスタンブロットは、erb-B1(「EGFW」;上段パネル)またはc-Cbl(中段および下段パネル)に特異的なmAbsで染色した(「WB」)。
【図15】図15Aは、ワクシニアウイルスで鼻腔内にチャレンジし、かつ無処置か(「対照」;白丸)、抗L1R mAbで一度前処理したか(「抗L1R」;黒丸)、CI-1033で毎日処置したか(「CI-1033」;白三角)、または抗L1R mAbで一度前処理して、CI-1033で毎日処置したか(「CI-1033 +抗L1R」;黒逆三角)のいずれかのマウスの、全時間にわたった生存を(種々の処理群のマウスの割合で)示す線グラフである。 図15Bは、図15Aに記載されているマウス実験群の臨床スコア(実施例1に記載したものに由来する)を示す線グラフである。図15Aに使用する同じシンボルを種々の実験群を示すために使用してある。 図15Cは、ワクシニアウイルスでの致死チャレンジ6日後の、処置なし(「対照」)、抗L1R mAbで前処理(「抗L1R」)、CI-1033で毎日処理(「CI-1033」)、または抗L1R mAbで前処理して、CI-1033で毎日処理したもの(「CI-1033 + 抗L1R」)のいずれかのマウスからの全肺の平均重量を示す棒グラフである。 図15Dは、ワクシニアウイルスに対する感染の8日後の、抗L1R mAbで前処理(「抗L1R」)、CI-1033で毎日処理(「CI-1033」)、13E8 mAbで前処理したもの、抗L1R mAbおよび13E8 mAbで前処理したもの、または抗L1R mAbで前処理して、CI-1033で毎日処理したもの(「CI-1033 + 抗L1R」)のいずれかのマウスの肺における平均ウイルス力価を示す棒グラフである(「Log10PFU」で;PFU(プラーク形成単位))。本実験の対照群(「なし」;斜線の棒)(ウイルスを感染させたが、前処理または処置を受けなかったマウス)のマウスの全てが、実験8日目に死んだので、対照群は、7日めに屠殺したマウスの別々の実験に由来する。
【図16】図16Aは、感染していない(「0」);ワクシニアウイルスに対する感染の示した日数(「4」、「6」または「8」)後の:マウスの肺から調製したRNAを使用するRnase保護アッセイ法からのオートラジオグラムの写真である。感染動物は、無処置であるか(「対照」)、抗L1R mAb(「抗L1R」)で前処理したか、CI-1033(「CI-1033」)で毎日処置したか、または抗L1R mAbで一旦前処理して、CI-1033(「CI-1033 + 抗L1R」)で毎日処置したものであった。写真の左上に示したように、肺RNAは、mCK-2bテンプレートセット(BD Biosciences-Pharmingen, San Diego, CA)のサイトカイン特異的なプローブとハイブリダイズさせた。保護断片をDNAシーケンシング・ゲル上で溶解し、次いでこれをX線写真に曝露した。CI-1033(単独で、または抗L1R mAbと共に)によってレベルが増大されたサイトカインのmRNAを写真の右上の矢印によって示してある。 図16Bは、インビトロにおいて:正常MC57G細胞(「MC57G」);またはワクシニアウイルスに感染したMC57G細胞(「VV感染MC57G」)で刺激されたマウス脾臓CD8+細胞内の細胞内インターフェロン-γのレベル(「IFN-γ(log)」)、または細胞表面CD44(「CD44(log)」)のレベルを示す一連の二次元FFCプロフィールである。実験群は、ワクシニアウイルスに感染して(屠殺し、アッセイする5日前)無処置(「対照」)か、抗L1R mAbで一回前処理した(「抗L1R」)か、CI-1033で毎日処置した(「CI-1033」)か、または抗L1R mAbで一回前処理してCI-1033で毎日処置した(「CI-1033」+「抗L1R」)からなる。示したデータは、マウスのそれぞれの実験群からの代表的マウスからのものである。
【図17】正常MC57G細胞(「MC57G」);ワクシニアウイルス感染MC57G細胞(「VV感染MC57G」);または抗CD 3mAb(「抗CD3」)でインビトロで刺激されたマウス脾臓CD8+細胞の細胞内インターフェロン-γ(「IFN-γ(log)」)のレベル、および細胞表面CD44(「CD44(log)」)のレベルを示す一連の二次元FFCプロフィールである。実験群は、無処置のマウス(「ナイーブ」)またはワクシニアウイルスに感染して(屠殺し、アッセイする5日前)無処置(「対照」)か、抗L1R mAbで一回前処理した(「抗L1R」)か、13E8 mAbで一回前処理した(「13E8」)か、または抗L1R mAbおよび13E8 mAbで一回前処理した(「抗L1R+13E8」)マウスからなる。示したデータは、マウスのそれぞれの実験群からの代表的マウスからのものである。
【図18】図18Aは、示した濃度のCI-1033の非存在下(「-」)または存在下(「+」)のいずれかにおいて、BSC-40細胞単層を培養することによって得た痘瘡株ソライメンの免疫染色を示す一連の写真である。 図18Bは、示した濃度のCI-1033の非存在下(「0」)または存在下(「CI-1033」)のいずれかにおいて、BSC-40細胞単層を培養することによって得たプラークの数に対するCI-1033の効果を示す棒グラフである。 図18Cは、示した濃度のCI-1033の非存在下(「0」)または存在下(「CI-1033」)のいずれかにおいてBSC-40細胞単層を培養することによって得たコメットの数を示す棒グラフである。
【技術分野】
【0001】
技術分野
本発明は、ウイルス感染の制御に、より詳細にはポックスウイルス(たとえば、天然痘)感染の制御に関する。
【背景技術】
【0002】
背景
B型肝炎などのウイルス病によって生じるヒトの経済的、および社会的な荒廃並びに天然痘などの疾患のバイオテロリストの利用に対する恐るべき可能性の点からみて、このような感染症に対する有効な治療薬および/または予防薬が開発されることが必須である。天然痘は、20年以上前に人類から根絶されたと考えられていたので、原因となるウイルス(痘瘡)のワクチン接種は、その時に廃止した。したがって、現在のヒト集団の比較的多くの割合が、これまでに疾患に対するワクチン接種がなされていない。さらに、ワクチン接種の廃止前に生産された限られた天然痘ワクチンの供給だけが現在利用可能である。
【発明の開示】
【0003】
概要
本発明者らは、erb-Bタンパク質チロシンキナーゼを阻害する化合物が、ポックスウイルス感染症を阻害するのに有効であることを発見し、したがって、これらの化合物は、有効な抗ウイルス治療薬および予防薬である可能性が高い。
【0004】
実施例2は、哺乳類タンパク質エピレギュリン(エピレギュリン:EPI)に対して、特にこれらの上皮性因子(EGF)様のドメイン内において、比較的高い相同性を有するポックスウイルス・タンパク質を記載してある。これらのポックスウイルス・タンパク質は、本明細書においてエピレギュリン様成長因子(ELGF)と称される。天然痘(痘瘡)ウイルスELGFは、指定を受けた天然痘成長因子(SPGF)であり、ワクシニアELGFは、指定を受けたワクシニア成長因子(VGF)である。
【0005】
本発明は、一つには、細胞表面erb-B1受容体に対するD4R(大痘瘡のバングラデシュ株に由来するSPGF)の組換え上皮細胞成長因子(EGF)様ドメインの結合が、(a)これらを発現する細胞におけるこのような受容体のチロシンキナーゼ活性の活性化;(b)このような細胞の増殖および/または生存の増強;(c)erb-B1受容体の迅速なインターナリゼーション;並びに(d)複製および付随する炎症の増強によって生じるウイルス病原性を生じるという観察に基づいている。これらの知見は、SPGFが(たとえば、痘瘡ビリオンの表面で、または痘瘡ビリオンから分断されて)、痘瘡ウイルスの宿主細胞への侵入、およびその中での複製、およびその後の炎症において中心的役割を果たすことを示す。
【0006】
加えて、本発明者らは、erb-B1受容体を経た細胞に対するSPGFの結合によって刺激されるタンパク質チロシンキナーゼ活性およびSPGFによるライゲーション後のerb-B1受容体のインターナリゼーションが、erb-Bタンパク質チロシンキナーゼ阻害剤によって阻害されることを見いだした。このerb-B1インターナリゼーションの阻害は、明らかにc-Cblとerb-B1の間の誘導性の結合の阻害によるものであった。これらの知見と一致して、本発明者らも、キナゾリン・タンパク質チロシンキナーゼ阻害剤CI-1033が、ワクシニアウイルスに感染した動物における臨床症状を阻害し、ウイルス負荷を減少したことを観察した。このCI-1033の処置効果は、中和ワクシニア特異的モノクローナル抗体(mAb)(抗L1R mAb)の同時投与によって増強された。加えて、本発明者らは、erb-Bチロシンキナーゼ阻害剤が1つの細胞から別の細胞への痘瘡ウイルスの伝播を減少することを見いだした。
【0007】
面白いことに、CI-1033は、また、感染動物におけるワクシニアウイルスに対するT細胞性免疫を増強したこと、およびこの増強は、感染動物に対して抗L1R mAbを同時投与することによってさらに増大することが判明した。さらに、本発明者らは、CI-1033と同様に、VGF(SPGFのワクシニア相同分子種)に結合するmAbがワクシニアに感染した動物におけるワクシニアウイルスに対するT細胞免疫を増強したこと、およびこの効果が、動物に対する抗L1R mAbの投与によってさらに増大されたことを見いだした。
【0008】
これらの知見は、erb-Bチロシン・タンパク質キナーゼ活性の阻害剤、またはタンパク質キナーゼ活性の活性化の阻害剤が、動物(たとえば、ヒト被験者)のポックスウイルス感染症に対する有効な薬剤であり得ることを示す。これらの阻害剤は、少なくとも、細胞におけるウイルス複製を阻害すること、および細胞からウイルスが出ることによって、並びにウイルスに対する免疫の増強によって作用すると思われる。また、阻害剤は、細胞へのウイルス侵入を減少させるかもしれない。
【0009】
より詳細には、本発明は:(a)ウイルスerb-Bリガンドをコードする遺伝子を含むウイルスに曝露された可能性があるか、または暴露されている可能性がある動物を同定する工程;および、(b)erb-Bチロシンキナーゼの活性を阻害するか、またはerb-Bチロシンキナーゼの活性化を阻害する化合物で動物を処置する工程、を含む方法を特徴とする。動物は、工程(b)の前に、間に、または後に、ウイルスに感染していたと同定される。ウイルスerb-Bリガンドは、オルトポックスウイルス(たとえば、大痘瘡、小痘瘡、サル痘ウイルス、またはワクシニア)erb-Bリガンドなどのポックスウイルスerb-Bリガンドであることができる。erb-Bリガンドは、天然痘成長因子(SPGF)またはワクシニア成長因子(VGF)などのエピレギュリン様の成長因子(ELGF)であることができ、erb-Bチロシンキナーゼは、erb-B1、erb-B2、またはerb-B4チロシンキナーゼであることができる。
【0010】
化合物は、たとえばerb-Bチロシンキナーゼと結合する非アゴニスト抗体、または非アゴニストerb-Bリガンド、またはerb-Bリガンドの非アゴニスト断片であることができる。または、化合物は、たとえば、キナゾリンに基づいた化合物(たとえば、4-アニリノ・キナゾリン)、ピリドピリミジンに基づいた化合物、キノリン-3-カルボニトリルに基づいた化合物、またはピロロピリミジンに基づいた化合物などの小分子erb-Bチロシンキナーゼ阻害剤であることができる。小分子erb-Bチロシンキナーゼ阻害剤は、erb-Bチロシンキナーゼ活性の不可逆的または可逆的な阻害剤であることができる。キナゾリンに基づいた化合物は、たとえば、CI-1033、PD168393、PD160678、PD160879、PD174265、PD153035、ZD1839、GW572016、GW974、OSI-774、またはAG1478であることができる。ピリドピリミジンに基づく化合物は、たとえばPD69896、PD153717、またはPD158780であることができる。キノリン-3-カルボニトリルに基づいた化合物は、たとえばEKB-569であることができ、およびピロロピリミジンに基づいた化合物は、たとえばCGP59326Aであることができる。
【0011】
本方法では、動物は、ヒトであることができる。本方法は、ウイルスの1つもしくは複数の形態を実質的に中和する能力を有する抗体、たとえばオルトポックスウイルス(たとえば、大痘瘡、小痘瘡、ワクシニア、またはサル痘ウイルス)の細胞内成熟ウイルス粒子(IMV)形態、細胞外外膜ウイルス(EEV)形態、および/または細胞結合エンベロープ・ウイルス(CEV)形態と結合する抗体を動物に投与することをさらに含むことができる。
【0012】
本方法において、化合物での処置は、動物におけるウイルスに対する免疫応答を増強することができる。免疫応答は、T細胞応答、たとえばCD8+ T細胞応答またはCD4+ T細胞応答であることができ、T細胞応答は、インターフェロン-γ(IFN-γ)産生T細胞応答であることができる。または、免疫応答は、抗体産生B細胞応答であることができる。
【0013】
本方法の一つの態様において、ウイルスは、ポックスウイルス発現ベクターであることができ、かつ(a)免疫原をコードする異種核酸配列;および、(b)異種核酸配列に動作可能に連結される、転写調節性のエレメント(TRE)をさらに含むことができる。ポックスウイルス発現ベクターは、ワクシニアウイルス発現ベクター、弱毒ワクシニアウイルス発現ベクター、カナリア痘瘡ウイルスベクター、または鶏痘ウイルスベクターであることができる。
【0014】
また、本発明は:(a)erb-Bリガンドをコードする遺伝子を含むウイルスによる感染に感受性の動物を同定する工程;並びに、(b)動物を、(i)ウイルスに対する免疫応答を刺激するウイルスまたはワクチンの1つもしくは複数の形態を実質的に中和する抗体、および(ii)erb-Bチロシンキナーゼを阻害するか、またはerb-Bチロシンキナーゼの活性化を阻害する化合物で処置することと、を含む方法を特徴とする。ウイルス、動物、erb-Bリガンド、化合物、およびウイルスの1つまたは複数の形態を実質的に中和する抗体は、上記したもののいずれであることもできる。そのうえ、本方法は、上記で詳述した免疫応答のいずれを増強することもでき、ウイルスは、上記ポックスウイルス発現ベクターのいずれであることもできる。
【0015】
本発明のもう一つの態様は:(a)erb-Bチロシンキナーゼと結合し、かつerb-Bチロシンキナーゼの活性または受容体のチロシンキナーゼの活性化を阻害する単離された化合物を提供する工程;(b)erb-Bチロシンキナーゼを発現する細胞と化合物を接触させる工程;および、(c)工程(b)の前に、と同時に、もしくは後に、ウイルスerb-Bリガンドまたはリガンドの機能的な断片と細胞を接触させる工程を含むインビトロ方法である。化合物は、リガンドまたは断片による細胞上のerb-Bチロシンキナーゼの活性化を減少させることができる。または、本方法は、化合物が、リガンドまたは断片による細胞上のerb-Bチロシンキナーゼの活性化を減少させるかどうか決定することをさらに含むことができる。ウイルスerb-Bリガンド、erb-Bチロシンキナーゼ、および化合物は、上記したもののいずれであることもできる。
【0016】
また、本発明は、化合物が抗ウイルス性の化合物であるかどうかを決定する方法であって:(a)erb-Bチロシンキナーゼの活性を阻害するか、またはerb-Bチロシンキナーゼの活性化を阻害する化合物を提供する工程;(b)erb-Bリガンドをコードする遺伝子を含むウイルスに対する感染に感受性の動物に対して化合物を投与する工程;(c)工程(b)の前に、の間に、または後に、ウイルスに対して動物を曝露する工程;および、(d)化合物が動物のウイルス感染の症状を減少させるかどうか決定する工程を含む方法を提供する。ウイルスは、ポックスウイルス、たとえば大痘瘡、小痘瘡、サル痘ウイルス、またはワクシニアなどのオルソポックスウイルスであることができる。動物、erb-Bリガンド、および化合物は、上記したもののいずれであることもできる。
【0017】
さらに本発明による特徴とされるもう1つの方法は:(a)erb-Bリガンドをコードする遺伝子を含むウイルスによる感染に感受性の動物を同定する工程;並びに、(b)動物を、(i)リガンドに対する抗体、および(ii)ウイルスに対する免疫応答を刺激するウイルスまたはワクチンの1つもしくは複数の形態を実質的に中和する抗体で処置する工程を含む。動物は、工程(b)の前に、の間に、または後に、ウイルスに感染していたと同定することができる。処置によりウイルスに対する免疫応答を増強することができ、免疫応答は、上記で列記したもののいずれであることもできる。ウイルスは、ポックスウイルス発現ベクターであることができ、かつ(a)免疫原をコードする異種核酸配列;および、(b)異種核酸配列に動作可能に連結される、転写調節性のエレメント(TRE)をさらに含むことができる。ポックスウイルス発現ベクターは、ワクシニアウイルス発現ベクター、弱毒ワクシニアウイルス発現ベクター、カナリア痘瘡ウイルスベクター、または鶏痘ウイルスベクターであることができる。リガンドと結合する抗体は、13E8モノクローナル抗体(ATCCアクセッション番号PTA-5040)または11D7モノクローナル抗体(ATCCアクセッション番号PTA-5039)であることができる。ウイルス、動物、erb-Bリガンド、化合物、およびウイルスの1つまたは複数の形態を実質的に中和する抗体は、上記したもののいずれであることもできる。
【0018】
「ポリペプチド」および「タンパク質」は、交換可能に使用され、長さまたは翻訳後の修飾に関係なく、何らかのペプチド結合されたアミノ酸鎖を意味する。本発明に使用されるポリペプチドは、保存的置換を有するものを含む。保存的置換は、典型的には以下の群内の置換を含む:グリシンおよびアラニン;バリン、イソロイシン、およびロイシン;アスパラギン酸およびグルタミン酸;アスパラギン、グルタミン、セリン、およびスレオニン;リジン、ヒスチジン、およびアルギニン;並びにフェニルアラニンおよびチロシン。一般に、保存的置換を有する変種ポリペプチドは、40未満(たとえば、35;30;25;20;15;13;11;10;9;8;7;6;5;4;3;2;または1つ未満)の保存的置換を含む。変種ポリペプチドは、野生型ポリペプチドの活性の少なくとも20%(たとえば、少なくとも:30%;40%;50%;60%;70%;80%;90% ;95%;98%;99%;99.5%;99.8%;99.9%;または100%以上)を有するということが、必要とされる全てである。
【0019】
本明細書に使用されるものとして「細胞の活性化」は、細胞増殖の増強または細胞の生存の増強をトリガーすることを意味する。「細胞増殖の増強または細胞の生存の増強」は、(a)低レベルの細胞増殖または細胞生存;または(b)実質的に細胞増殖または細胞生存がないこと、と比較してして増大されることを意味する。
【0020】
本明細書に使用されるものとして「免疫応答を増強する」ことは、(a)より低い免疫応答;または(b)実質的に免疫反応がないこと、と比較してして免疫応答を増大することを意味する。
【0021】
「単離された化合物」の用語は、本明細書に使用されるものとして、天然に存在する対応物を有さないか、または天然にこれを伴う成分、たとえば膵臓、肝臓、脾臓、卵巣、精巣、筋肉、関節組織、神経組織、胃腸組織、もしくは腫瘍組織などの組織、または血液、血清、もしくは尿などの体液中の成分から分離されず、または精製されていない化合物(たとえば、タンパク質)をいう。典型的には、天然に存在する生物学的化合物は、これが、乾燥重量で、天然に付随しているタンパク質およびその他の天然に存在する有機分子が少なくとも70%であるときに、「単離された」とみなされる。好ましくは、本発明に使用される化合物の標品は、その化合物の乾燥重量で、少なくとも80%、より好ましくは、少なくとも90%、および最も好ましくは少なくとも99%である。単離または純度の程度は、任意の適切な方法によって、たとえばカラムクロマトグラフィー、ポリアクリルアミドゲル電気泳動、またはHPLC(高速液体クロマトグラフィー)解析によって測定することができる。化学的に合成された化合物(たとえば、タンパク質)は、その性質上、天然にこれに付随する成分から分離されており、合成化合物は、定義上「単離され」ている。
【0022】
単離された化合物、および本発明のために有用なさらなる薬剤は、たとえば以下によって得ることができる:(i)天然の供与源から(たとえば、組織または体液から)の抽出;(ii)化合物がタンパク質である場合は、タンパク質をコードする組換え核酸の発現により;または(iii)当業者に公知である標準的な化学的合成法により。これが天然に由来する供与源とは異なる細胞の系で産生されるタンパク質は、これが天然にそれを伴う成分を必然的に含まないので、「単離され」ている。
【0023】
他に定義されない限り、本明細書に使用される全ての技術的および科学的な用語は、本発明が属する当該技術分野において当業者によって共通に理解されるものと同じ意味を有する。矛盾する場合には、定義を含む本明細書が対照となる。好ましい方法および材料は、以下に記載してあるが、本明細書に記載されているものと同じか、または均等な方法および材料も、本発明の実施または試験に使用することができる。全ての刊行物、特許出願、特許、および本明細書において述べられるその他の参照は、これらの全体が参照として組み入れられる。本明細書に開示された材料、方法、および実施例は、例示するだけであり、限定されることは企図されない。
【0024】
たとえば、細胞の活性化を阻害することを含む、本発明のその他の特徴および利点は、以下の明細書から、図面から、および特許請求の範囲から明らかであろう。
【0025】
詳細な説明
上記の概要の節において概説した実験の知見は、erb-B1タンパク質チロシンキナーゼ活性、またはerb-B1タンパク質チロシンキナーゼ活性の刺激のいずれかを阻害する化合物が、オルソポックスウイルスを媒介した病気の発生(たとえば、天然痘またはワクシニア感染症)の治療、および/またはこれに対する予防に有効であり得ることを示す。本発明は、いずれの特定の作用機構によっても限定されないが、このような化合物は、宿主細胞へのウイルス粒子の侵入、細胞内のウイルス複製、および/または細胞からのウイルス放出を阻害することによって、並びにウイルスに対する適応性T細胞免疫を増強することによって作用するのであろうと考えられる。オルトポックスウイルスのゲノムは、免疫抑制因子をコードする遺伝子を含むという点で、ウイルス複製の阻害および適応免疫の増強に機能的に大いに関連する可能性がある。たとえば、B型肝炎ウイルス、EBウイルス、RNA型腫瘍ウイルス(たとえば、ニワトリ赤芽球症ウイルス)、サイトメガロウイルス(CMV)、およびレオウイルスによる細胞の感染におけるerbB受容体の役割を示す以前の知見は(実施例7の参照を参照されたい)、本観察を考慮して、erb-Bタンパク質チロシンキナーゼの阻害剤が複数のポックスウイルスだけでなく、多種多様なその他のウイルスに対しても有効な治療薬および/または予防薬である可能性が高いことを示唆する。
【0026】
天然痘(大痘瘡および小痘瘡)および牛痘(ワクシニア)ウイルスでは、特に興味がもたれる。これらのウイルスは、感染細胞の細胞質で複製する大きなニ本鎖DNAウイルスである。痘瘡ウイルスは、気道を介して被検者に入り;ヒト-ヒトの伝達は、通常経口咽頭分泌物中のウイルスの咳嗽の結果として生じる。7〜19日のインキュベーション期間に続いて、熱、頭痛、および背中の痛みがある。2〜3日後に、熱が下がり、顔、躯幹、および四肢に発疹が現れ、小胞、膿疱、および痂皮に進行して数週間の間続く。大痘瘡感染症は、ワクチン非接種人間の約40%では致命的である。死は、主に内出血(播種性血管内凝固)および血管虚脱によるものである。小痘瘡に感染すると、致死率の減少したあまり重症型ではない天然痘を生じる。
【0027】
1980年頃の天然痘の国際根絶により、ワクチン接種は廃止した。従って、天然痘の防護は、米国において約20年間提供されてこなかった。その結果、かなりの個体群が、天然痘に対してこれまでワクチン接種がなされなかった。さらに、米国には、現在、天然痘ワクチン(弱毒化ワクシニアウイルス)の適当な蓄えがなく、利用できるものは、免疫不全(たとえば、AIDSおよび癌)患者に有害な影響を及ぼす能力がある生きたワクシニアウイルスを産した十分に制御されていないプロセスによって産生されるものである。加えて、10〜20の第二世代の場合は、一人のヒトの痘瘡感染から生じる可能性がある。これらの要因により、痘瘡ウイルス、特に大痘瘡ウイルスは、米国人口に対する生物兵器としての明白な選択肢となる。したがって、ウイルスに対する効率的な、安全な治療的な、および/または予防的な方法論ができる限り速やかに開発されることが不可避である。
【0028】
痘瘡は、最も大ききく、かつ最も複雑なニ本鎖DNAウイルスの1つであり、光学顕微鏡によって見える。配列データは、大痘瘡ウイルスおよび小痘瘡ウイルスについて入手可能である。GenBankの痘瘡の現在の配列データには、以下を含む:(a)大痘瘡の5つのヌクレオチド配列の登録および626アミノ酸配列の登録;並びに(b)小痘瘡の3つのヌクレオチド配列の登録および619アミノ酸配列の登録。2つの完全な大痘瘡ゲノム・ヌクレオチド配列があり、インド株1967(GI:9627521;185,578 bp)およびバングラデシュ株(GI:623595;186,103 bp)に対応しており、小痘瘡株アラストリムの1つの完全なゲノム・ヌクレオチド配列(GI:5830555;186,986 bp)がGenBankで入手可能である。大痘瘡および小痘瘡ゲノムの比較では、これらが97%以上、全ゲノムの全体にわたって同一であることを示す。同様に、いずれの痘瘡ゲノムのワクシニアのものとの比較でも(GI:9790357;191,737 bp)、これらが〜95%同一であることを示す。
【0029】
大痘瘡インド株1967のゲノムには、197個の遺伝子がある(GI:9627521)。これらの機能およびウイルスのライフサイクルの関連については、ほとんど知られていない。痘瘡ウイルス(大痘瘡インド株)のゲノムのコンピュータに基づいた解析からのデータでは、天然痘に対するワクチンとして、および有効な受動免疫保護剤と考えられる抗体(同時係属中のU.S. Application Serial No. 10/429,685を参照されたい(この開示はその全体が参照として本明細書に組み入れられる))を作製するために潜在的に有用であるゲノム内の遺伝子によってコードされる種々のポリペプチドを同定した。この解析によって同定されたポリペプチドのうちの1つは、D1Lであり、これは、上皮細胞成長因子(EGF)様膜結合型タンパク質であり、その他の痘瘡株の対応する分子に対して≧97%の相同性、対応するワクシニア・タンパク質と86〜89%の同一性、およびヒト・エピレギュリンに対して30%の同一性を有する。上記のように、これらのポックスウイルスのEGF様膜結合タンパク質は、本明細書において、ひとまとめにしてエピレギュリン様成長因子(ELGF)と称し;痘瘡株由来のELGFは、本明細書において、天然痘成長因子(SPGF)と称し、およびワクシニア株由来の由来のものは、ワクシニア成長因子(VGF)と称される。図3AおよびBでは、2つの痘瘡株由来のSPGF、すなわち、大痘瘡インド由来のD1L(図3A;SEQ ID N0:1)および大痘瘡バングラデシュ由来のD4R(図3B;SEQ ID NO:2)のアミノ酸配列を示してある。これらの2つのSPGFは、2つのアミノ酸部位(図3AおよびBにおいて太字で示し、下線を引いた)のみが異なる点に留意されたい。
【0030】
本発明の種々の態様を以下に記載してある。
【0031】
細胞の活性化を阻害する方法
本発明は、細胞表面erb-Bタンパク質チロシンキナーゼ分子を発現する細胞の活性化を阻害する方法を含む。これらの方法では、erbBタンパク質チロシンキナーゼ活性を阻害するか、またはerb-Bタンパク質チロシンキナーゼ活性の活性化を阻害する1つまたは複数の単離された化合物が、関心対象の細胞に送達される。1つまたは複数の化合物の細胞に対するデリバリーは、ウイルスerb-Bリガンド(たとえば、ELGF)または機能的な(すなわち、erb-Bに結合し、活性化する)erb-Bリガンドの断片と細胞の接触前に、同時に、もしくは後に生じさせることができる。本方法は、デリバリー工程の前に、erb-Bタンパク質チロシンキナーゼを発現する細胞を同定することを含むことができる。細胞は、erb-Bタンパク質チロシンキナーゼ分子を発現する任意の細胞、たとえば:任意の腫の上皮細胞、例えば肺上皮細胞などの肺上皮細胞、胃腸上皮細胞(たとえば、胃または結腸の上皮細胞)、皮膚上皮細胞(たとえば、ケラチノサイト)、または泌尿生殖系の上皮細胞(たとえば、膀胱または子宮上皮細胞);または線維芽細胞であることができる。細胞は、正常細胞または悪性細胞であることができる。erbBタンパク質チロシンキナーゼは、erb-B1、erb-B2、もしくはerb-B4タンパク質チロシンキナーゼ、これらの3つのタンパク質のいずれかのホモ二量体、またはerb-B1、erb-B2、erb-B3、およびerb-B4タンパク質チロシンキナーゼの任意の二つの間のヘテロ二量体であることができる。
【0032】
ウイルスerb-Bリガンド(EBL)は、ゲノムがerb-Bタンパク質チロシンキナーゼと結合するタンパク質をコードする核酸(DNAまたはRNA)を含む任意のウイルスに由来することができる。たとえば、ウイルスEBLは、本明細書に一覧を示した任意のウイルスであることができる。たとえば、ポックスウイルス由来であることができる。関心対象のポックスウイルス(ポックスウイルス科)は、オルソポックスウイルス属(たとえば、痘瘡、ワクシニア、およびサル痘)、トリポックスウイルス属(Avipoxviruses)(たとえば、鶏痘およびカナリア痘瘡)、カプリポックスウイルス属(たとえば、ヒツジ・ポックスウイルス)、レオポリポックスウイルス属(Leoporipoxviruses)(たとえば、粘液腫およびショープウイルス)、パラポックスウイルス属(たとえば、オルフウイルスおよび豚痘)、軟体動物ポックスウイルス属(Molluscipoxviruses)(たとえば、伝染性軟属腫ウイルス)、およびヤタポックスウイルス属(Yatapoxviruses)(たとえば、ヤタサル腫瘍ウイルス)を含むが、これらに限定されない。痘瘡(大痘瘡および小痘瘡)およびワクシニアウイルスは、特に関心がもたれる。ポックスウイルスELGFは、たとえばSPGF(たとえば、大痘瘡インド株由来のD1Lタンパク質または大痘瘡バングラデシュ株由来のD4Rタンパク質)またはVGF(ワクシニアウイルス由来)であることができる。ウイルスEBLの機能的な断片は、本明細書に使用されるものとして、断片は、全長の野生型の成熟タンパク質よりも短いが、野生型の成熟タンパク質がerb-B受容体の1つまたは複数(野生型の全長タンパク質が結合する)と結合する能力の少なくとも20%(たとえば、少なくとも:30%;40%;50%;60%;70%;80%;90%;95%;97%;98%;99%;99.5%;99.7%;99.8%;99.9%、または100%以上)を有する。リガンドが受容体と結合する能力を測定する方法は、当該技術分野において既知である(このような方法の実施例については以下を参照されたい)。機能的な断片は、たとえばEGF様ドメイン、たとえばSPGF D1L(SEQ ID NO:1)、もしくはD4R(SEQ ID NO:2)のアミノ酸40〜90であるか、またはこれを含むことができる。配列の整列により(図2を参照されたい)、当業者であれば、関心対象のEBLの領域がEGF様ドメインを含むか、またはEGF様ドメインであることを理解することができるであろう。
【0033】
細胞表面erb-Bタンパク質チロシンキナーゼに接触するウイルスEBLは、ウイルス粒子の成分であることができ、実際に、細胞表面erb-B分子に対するこの形態のウイルスEBLの結合は、ウイルス粒子による細胞の感染の少なくとも1つの初期工程であることができる。または、ウイルスEBL(またはこれらの機能的な断片)は、ウイルスに感染した細胞によって分泌されるか、または細胞外ウイルス粒子から分断することができる。加えて、ウイルスEBL(またはこれらの機能的な断片)は、たとえばウイルスから部分的もしくは完全に精製されたか、または化学的もしくは組換えで産生された分子であることができる。
【0034】
erb-Bタンパク質チロシンキナーゼを阻害する化合物は、erb-Bタンパク質と結合する抗体であることができる。このような抗体は、非アゴニスト抗体、すなわちそれが結合する受容体を介したシグナルを実質的に活性化しない抗体であることができる。erb-Bの外部ドメインに特異的な抗体は、リガンド結合に関して競合して、erb-B1シグナリングをブロックするものであると記載されており;さらに、これらの抗体は、細胞周期停止および/または細胞死を生じさせることが示された[Arteaga (2001) J. Clin Oncol. 19 (18) (September 15 supplement): 32s-40s、およびその中で引用される参照文献]。
【0035】
関連した化合物のもう一つのクラスは、ウイルスEBLに結合する抗体、たとえば3D4R-13E8および3D4R-11D7 mAbsなどのSPGFまたはVGFと結合するmAb(それぞれ13E8および11D7 mAbsとも称される)を含む。
【0036】
さらに、本発明の方法のために有用な化合物は、非アゴニストerb-Bリガンドまたはerb-Bタンパク質と結合するerb-Bリガンドの非アゴニスト断片であることができる。関心対象の受容体と結合する化合物がアゴニストまたはその受容体の非アゴニストであるかどうかを決定するための方法は、当該技術分野において既知である。erb-Bの哺乳類リガンドは、EGF、エピレギュリン、トランスフォーミング成長因子α(TGFα)、アンフィレギュリン(amphiregulin)、ヘパリン結合EGF、ベータセルリン(betacellulin)、およびヘレギュリン(heregulin)を含むが、これらに限定されない[Arteaga et al.(2001) J. Clin. Oncol. 19: 32s-40s]。その他の適切な化合物は、ウイルスタンパク質、たとえばELGF(例えばSPGFまたはVGF)の非アゴニスト断片を含む。
【0037】
小分子erb-Bチロシンキナーゼ阻害剤
特に興味がもたれる化合物は、erb-Bタンパク質チロシンキナーゼ活性の小分子阻害剤である。多くのこのような化合物が記載されている。このような化合物には、本質的に3つのカテゴリーがある。カテゴリーAには、erb-Bタンパク質チロシンキナーゼの触媒ドメインのMg-ATP結合部位に競合することによって実質的に作用するように見える化合物がある[Al-Abeidi (2000) Oncogene 18:5690-5701]。カテゴリーBには、erb-Bタンパク質チロシンキナーゼ上のチロシン部位に競合することによって明らかに実質的に作用する化合物がある[Posner et al. (1994) Mol. Pharmacol. 45:673-683、その全体が参照として本明細書に組み入れられる]。カテゴリーCには、両方の機構によって作用するように見える化合物がある[Posner et al. (1994) Mol. Pharmacol. 45:673-683]。
【0038】
カテゴリーAは、二環式のキナゾリンに基づいた化合物(たとえば、4-アニリノキナゾリン)、ピリドピラミジンに基づいた化合物、キノリン-3-カルボニトリルに基づいた化合物、ピロロピリミジンに基づいた化合物、ピラゾロピリミジンに基づいた化合物、並びにキナゾリンの三環系の誘導体(イミダゾキナゾリン、ピロロキナゾリン、およびピラゾロキナゾリン)、並びに上記の一覧に記載されたその他の二環式化合物を含むが、これらに限定されない[Fry et al. (2003) Exp. Cell Res. 284:131-139、その全体が参照として本明細書に組み入れられる]。
【0039】
キナゾリンに基づいた化合物の例は、以下を含む:ZD1839/イレッサ[Fry et al.(2003)];GW572016(N-{3-クロロ-4-[3-フルオロベンジル)オキシ]フェニル}-6-[5-({[2-メチルスルホニル)エチル]アミノ}メチル)-2-フリル]-4-キナゾリン[Xia et al. (2002) Oncogene 21:6255-6263;および Cockerill et al. (2001) Bioorg. Med. Chem. Lett. 11:1401-1405、両方ともその全体が参照として本明細書に組み入れられる];GW974[Cockerill et al. (2001);および Rusnak et al. (2001) Cancer Res. (2001) 61:7196-7203、その全体が参照として本明細書に組み入れられる];PD168393[Fry et al. (1998) Proc. Natl. Acad. Sci. USA 95:12022-12027、その全体が参照として本明細書に組み入れられる;図12]、PD153035/ZM252868/AG1517[Fry et al. (1994) Science 265:1093-1095、その全体が参照として本明細書に組み入れられる;図12]、およびCI-1033[Pfizer;図12];OSI-774/CP-358、774/Tarceva[Fry et al.(2003);および Stamos et al. (2002) J. Biol. Chem. 277:46265-46272、その全体が参照として本明細書に組み入れられる];AG1478[Levitzki et al. (1995) Science 267:1782-1788、両方ともその全体が参照として本明細書に組み入れられる];並びにPD160678、PD160879、PD168393、およびPD174265[Fry et al. (1998) Proc. Natl. Acad. Sci. USA 95:12022-12027]。
【0040】
有用なピリドピリミジンに基づいた化合物の例はPD69896、PD153717、およびPD158780[Fry et al. (1997) Biochem. Pharmacol. 54 (8):877-887、その全体が参照として本明細書に組み入れられる]を含む。有用なキノリン-3-カルボニトリルは、EKB-569[Fry et al. (2003)]であり、有用なピロロピリミジンは、CGP59326A/PKI-166[Lydon et al. (1998) Int. J. Cancer 76 (1):154-163、その全体が参照として本明細書に組み入れられる]であり、および有用なピラゾロピリミジンは、たとえば、Traxler et al. [(1997) J. Med. Chem. 40:3601-3616、その全体が参照として本明細書に組み入れられる]の化合物9および11を含む。
【0041】
広範囲にわたる置換基を有する化合物のこれらのクラスの全ての複数の誘導体が、erb-Bチロシンキナーゼ阻害活性を示すことが明らかであり、したがって、本発明の方法の候補である可能性が高い[たとえば、
を参照されたい、これらの全てが、参照としてこれらの全体が本明細書に組み入れられる]。
【0042】
いくつかの上記の化合物(たとえば、CI-1033、PD168393、EKB-569、PD1606678、PD 160879、およびPD 174265)は、アクリルアミド基によって、たとえば、erb-B1のCys773残基をアルキル化することによって、不可逆阻害剤として作用する。一連のこのようなアクリルアミド誘導体の相対的な有効性の比較は、Smaill et al [(1999) J. Med. Chem. 42(10):803-1815、その全体が参照として本明細書に組み入れられる]に記載されている。加えて、これらの化合物をアルキルアミンおよびアルコキシアミン側鎖によって置換すると(たとえば、キナゾリンおよびピリドピリミジン化合物の7位置で)、水性溶媒の溶解性の問題を解決するために役立つことが示された[Smaill et al. (2000) J. Med. Chem. 43 (7) 1380-1397、その全体が参照をそばに本明細書において組み入れられる]。
【0043】
また、カテゴリーAには、化合物DAPH 1(4,5-ジアニリノフタリミド)、およびその誘導体[Buchdunger et al.(1994)Proc. Natl. Acad. Sci. USA 91:2334-238、その全体が参照として本明細書に組み入れられる]、並びに真菌産物、ケルセチン、ゲニステイン、およびラベンズスチン(lavendustin)A[Levitzki et al. (1995) Science 267:1782-1788、その全体が参照として本明細書に組み入れられる]がある。
【0044】
カテゴリーBには、化合物AG537およびAG538[Posner et al. (1994) Mol. Pharmacol. 45:673-683]、並びにCushman et al. [(1994) J. Med. Chem. 37:3353-3362、その全体が参照として本明細書に組み入れられる)の化合物10a、10c、および10mなどのフラボン誘導体がある。
【0045】
カテゴリーCには:Geissler et al. [(1990) J. Biol. Chem. 265 (36):22255-22261、その全体が参照として本明細書に組み入れられる]の化合物1、2、および3などのチアゾリジンジオン;Showalter et al. [(1997) J. Med. Chem. 40:413-426、その全体が参照として本明細書に組み入れられる]の化合物33および34などの3-置換された2,2'-ジセレノビス(1H-インドール);Thompson et al. [(1993) J. Med. Chem. 36:2459-2469、その全体が参照として本明細書に組み入れられる]の化合物9、32、および33などの2,3-ジヒドロ2-チオキソ-1H-インドール-3-アルカノン酸および2,2'-ジヒドロビス(1H-インドール-3-アルカノン酸);並びにTraxler et al. [(1995) J. Med. Chem. 38:2441-2448、その全体が参照として本明細書に組み入れられる]の化合物5、17、18、および19などの[(アルキルアミノ)メチル]アクリロフェノンおよび(アルキルアミノ)プロピオフェノンがある。
【0046】
「小分子erb-Bチロシンキナーゼ阻害剤」の用語は、本明細書に使用されるものとして、表題「小分子erb-Bチロシンキナーゼ阻害剤」の下で上記した全ての化合物をいう。
【0047】
大分子(たとえば、抗体)および小分子の両方の化合物は、単独で細胞に、または2以上、たとえば3、4、5、6、7、8、9、10、15、または20を組み合わせて送達することができる。このような組み合わせは、上記の阻害剤のクラスの1つだけ、または1つ以上(たとえば、2、3、または4)を含む組み合わせを含むことができる。さらに、上記の化合物の効果を増強するために、補充薬を細胞または細胞の環境、たとえば細胞を含む培養液または動物に送達することができる。使用することができるそのような補充薬は、関心対象のEBLをコードする遺伝子を含むウイルスの1つまたは複数の形態と結合して、ウイルスを実質的に中和する抗体である。本明細書で使用されるものとして、「実質的に中和する」とは、少なくとも5倍、たとえば少なくとも10倍;20倍;50倍;100倍;1,000倍;10,000倍;100,000倍;1,000,000倍、またはそれ以上ウイルスの力価を減少させる能力を有することを意味する。このような抗体の例は、ポックスウイルスの細胞内成熟ウイルス粒子(IMV)(たとえば、L1Rワクシニア・タンパク質に、またはその痘瘡相同分子種に特異的な抗体)、細胞外外膜ウイルス(EEV)、または細胞結合される外膜ウイルス(CEV)と結合する中和抗体を含む。
【0048】
一定のインビボ態様において、被検者に中和抗体を投与する代わりに、被検者には、インサイチュウで中和抗体を産生させるために、適切な抗原の供与源をワクチン接種することができる。免疫化の適切な抗原および方法は、下記の免疫応答の増強法の節に記載してある。
【0049】
上記した全ての抗体は、ポリクローナル抗体またはmAbであることができ、任意の広範な腫、たとえばヒト、非ヒト霊長類(たとえば、サルまたはチンパンジー)、ウシ、ウマ、ヤギ、ヒツジ、ブタ、ネコ、イヌ、ウサギ、モルモット、ハムスター、スナネズミ、ラット、マウス、またはニワトリに由来するものであることができる。
【0050】
本明細書で使用されるものとして、「抗体」の用語は、完全な抗体(たとえば、IgM、IgG、IgA、IgD、またはIgE)分子だけではなく、抗原結合性断片、たとえばFab、F(ab')2、Fv、および単鎖のFv(scFv)断片もいう。scFv断片は、scFvが誘導される抗体の重鎖および軽鎖の可変領域の両方を含む単一のポリペプチド鎖である。また、キメラ抗体、たとえばヒト化抗体も含まれる。
【0051】
分子の結合ドメインを含む抗体断片は、既知の技術によって作製することができる。たとえば:F(ab')2断片は、抗体分子のペプシン消化によって作製することができ;およびFab断片は、F(ab')2断片のジスルフィド架橋を還元させることによって、またはパパインおよび還元剤で抗体分子を処置することによって作製することができる。たとえば、National Institutes of Health, 1 Current Protocols In Immunology, Coligan et al., ed. 2.8, 2.10 (Wiley Interscience, 1991)を参照されたい。scFv断片は、たとえばU.S. Patent No. 4,642,334(その全体が参照として本明細書に組み入れられる)に記載されているように作製することができる。
【0052】
たとえば、キメラ・ヒト化モノクローナル抗体は、Robinson et al., 国際特許出願第PCT/US86/02269号;Akira et al., 欧州特許出願第184,187号;Taniguchi, 欧州特許出願第171,496号;Morrison et al., 欧州特許出願第173,494号;Neuberger et al., PCT出願 国際公開公報第86/01533号;Cabilly et al., 米国特許第4,816,567号;Cabilly et al., 欧州特許出願第125,023号;Better et al. (1988) Science 240,1041-43 ; Liu et al. (1987) J. Immunol. 139, 3521-26;Sun et al. (1987) PNAS 84, 214-18;Nishimura et al. (1987) Canc. Res. 47,999-1005;Wood et al. (1985) Nature 314, 446-49;Shaw et al. (1988) J. Natl. Cancer Inst. 80, 1553-59;Morrison, (1985) Science 229, 1202-07;Oi et al. (1986) BioTechniques 4, 214;Winter, 米国特許第5,225,539号;Jones et al.(1986) Nature 321, 552-25;Veroeyan et al. (1988) Science 239, 1534;および、Beidler et al. (1988) J. Immunol. 141, 4053-60に記載されている方法を使用して当該技術分野において既知の組換えDNA技術によって作製することができる。
【0053】
完全ヒト抗体(ポリクローナルまたはモノクローナル)は、全てのヒト免疫グロブリン(すなわち、可変、接合、多様性、および定常)領域をコードする遺伝子セグメントをコードする遺伝子セグメントを含むトランスジェニック動物(たとえば、マウス)を免疫することによって産生することができる(たとえば、U.S. Patent Nos.5,545,806および5,569,825を参照されたい)。
【0054】
出願人は、アメリカン・タイプ・カルチャー・コレクション(ATCC), Rockville, MD 20852, U.S.Aで3D4R-11D7および3D4R-13E8ハイブリドーマをブダペスト条約の下で寄託した。3D4R-11D7ハイブリドーマには、ATCCアクセッション番号PTA-5039が、および3D4R-13E8ハイブリドーマにはATCCアクセッション番号PTA-5040が割り当てられた。ATCCで寄託したハイブリドーマは、本出願の優先日の前から Dana Farber Cancer Institute, Inc.によって維持された寄託から得られる。ハイブリドーマの寄託により、30年の期間、またはごく最近の要請後5年の期間、または有効な特許存続期間の間は、どんなに長くても、ATCC寄託所に制限なしで維持され、その期間の間に寄託が生育不能になった場合は交換される。
【0055】
細胞の活性化を阻害する方法は、インビトロまたはインビボであることができる。
【0056】
本発明の方法のインビトロでの適用は、ウイルス感染およびウイルス感染に対する細胞抵抗の機構に対する基本的科学的研究に有用であろう。本発明のインビトロ方法において、化合物は、関心対象の細胞(上記のものを参照)および上記で詳述した任意のウイルスEBLの形態と共に培養することができる。たとえば、ウイルス力価、細胞増殖/生存のレベル、および/またはタンパク質リン酸化の測定は、本明細書において記載され、および当該技術分野において既知の方法を使用して、種々のインキュベーション時間後に行うことができる。
【0057】
さらに、このような方法は、試験化合物を、これらが細胞のウイルス感染を阻害する能力についてスクリーニングするために有用であろう。このようなアッセイ法では、細胞と共に試験化合物を培養することによって、erb-B受容体の関心対象に結合する試験化合物をerb-B受容体を発現する細胞と接触させることができる。接触させる前、同時に、もしくは後に、細胞をウイルスEBLまたはEBLの機能的な断片と接触させることができる。次いで試験化合物が、リガンドまたは機能的な断片によってerb-B受容体の活性化が減少されるかどうかの決定がなされる。erb-B受容体の活性化は、当該技術分野において既知の方法によって、たとえば(a)活性化されたerb-B受容体によって直接、もしくはerb-B受容体の活性化に続いてカスケード様式で活性化されるタンパク質によって間接的にリン酸化されるerb-B受容体または下流の細胞内タンパク質のチロシン・リン酸化を検出または測定すること;(b)erb-B受容体のインターナリゼーションを検出または測定すること;または、(c)細胞の増殖および/または生存を検出または測定することによって測定することができる。
【0058】
また、使用する化合物がerb-B受容体を活性化することが既知である際の本発明のインビトロ方法は、ウイルスEBLによる細胞の活性化を阻害する能力を有する化合物についての上記スクリーニングアッセイ法の「ポジティブ対照」となり得る。
【0059】
本発明の方法は、好ましくはインビボで行われる。これらの適用は、関連したウイルス病の治療および予防に有用であろう。また、これらは、生きているウイルス(たとえば、牛痘)のワクチン接種の副作用を減弱させるためにも有用であろう。感染動物(たとえば、ヒト天然痘患者)、EBL発現ウイルスに感染するであろう動物(たとえば、ワクシニアウイルスでワクチン接種されたヒト被験者)、もしくは感染するリスクのある動物(たとえば、予想されるバイオテロリストの攻撃の間に大痘瘡ウイルスに感染するリスクのあるヒト被験者)に関心対象の化合物を投与することによって、ウイルスによって生じる臨床症状からの治療または予防を達成することができる。
【0060】
本明細書で使用されるものとして、「予防」は、疾患の症状の完全な予防、疾患の症状の発症の遅延、またはその後に発生する疾患症状の重篤さを少なくすることを意味することができる。本明細書で使用されるものとして、「治療」は、疾患の症状の完全な消滅または疾患の症状の重篤さの減少を意味することができる。
【0061】
本発明の上記のインビボの方法の修飾は、関心対象のウイルスに対する有効な予防薬および/または治療薬である化合物のスクリーニングアッセイ法として使用することができる。このような方法では、erb-Bチロシンキナーゼの活性を阻害するか、またはerb-Bチロシンキナーゼの活性化を阻害する試験化合物が、EBLをコードする遺伝子を含むウイルスに対する感染に感受性の動物に投与される。動物は、投与の前に、の間、またはの後、ウイルスに曝露される。次いで、化合物の有効性の決定は、たとえば体液(たとえば、血液、尿、または粘膜)、器官もしくは組織(たとえば、肺、脾臓、または肝臓組織)、または洗浄液(たとえば、肺、腸、膀胱、または膣の洗浄液)中のウイルスの力価の変化を検出または測定することによってなされる。または、ウイルス感染の1つまたは複数の症状(たとえば、熱、発疹、肢麻痺)の減少を検出または測定することができる。
【0062】
、ウマ、牛、ブタ、ヒツジ、ヤギ、イヌ、ネコ、ウサギ、モルモット、ハムスター、ラット、マウス、並びにニワトリ、シチメンチョウ、およびカナリアなどのトリに適用することができる。
【0063】
インビボ・アプローチ
好ましいインビボ・アプローチにおいて、単離された化合物自体が被検者に投与される。通常、化合物は、薬学的に許容されるキャリア(たとえば、生理食塩水)に懸濁して、経口で、もしくは静脈内(i.v.)注入によって投与されるか、または皮下に、筋肉内に、くも膜下腔内に、腹腔内に、内直腸に、腟内に、鼻腔内に、胃内に、気管内に、または肺内に注射される。これらは、たとえば、感染の部位、たとえば感染症が肺の中である場合は、肺内に直接送達することができる。必要とされる用量は、投与の経路の選択;製剤の性質;患者の疾病の性質;被検者のサイズ、重量、表面積、年齢、および性;投与されている他剤;並びに主治医の判断に依存する。適切な用量は、0.0001〜100.0mg/kgの範囲である。必要とされる用量の広範なバリエーションは、利用できる化合物の多様性および種々の投与経路での異なる効率から考えて予想される。たとえば、経口投与は、i.v.注射による投与よりも高い用量が必要であると思われる。これらの用量レベルのバリエーションは、当該技術分野において周知であるように、最適化のための標準的な経験的ルーチンを使用して調整することができる。投与は、一回または複数回(たとえば、2、3、4、6、8、10、20、50、100、150、またはそれ以上の回数)であることができる。適切なデリバリー媒体(たとえば、重合体の微小粒子または移植可能な装置)に化合物をカプセル化することにより、特に経口デリバリーに関してのデリバリーの効率を増大するであろう。
【0064】
有用な化合物がタンパク質である場合、これらは、このようなタンパク質をコードする核酸を含む発現ベクターを、またはこのような発現ベクターで形質転換された細胞(たとえば、動物に由来する細胞)を、細胞が存在する動物に対してインビボで投与することによって細胞に送達することができることが理解される。このような方法は、当該技術分野において周知である。
【0065】
免疫応答を増強する方法
本発明は、免疫応答を増強する方法を特徴とする。増強される免疫応答は、CD8+またはCD4+ T細胞応答であることができる。T細胞応答の増強は、サイトカイン/リンホカイン産生性または細胞障害性Tリンパ球(CTL)の反応であることができる。サイトカイン/リンホカイン産生性反応は、好ましくはTh1タイプのサイトカイン/リンホカイン反応、たとえばインターロイキン(IL)-12およびIFN-γを産生する反応である。しかし、また、一定の状況下で(たとえば、抗体対細胞免疫反応が好ましい)、インターロイキン(IL)-2、IL-3、IL-4、IL-10、IL-13、IL-15、顆粒球コロニー刺激因子(G-CSF)、マクロファージコロニー刺激因子(M-CSF)、もしくは顆粒球マクロファージ-コロニー刺激因子(GM-CSF)などのサイトカインまたはリンホカインの産生を含む反応を活性化することも望ましいであろう。(1)本発明の方法が、CD4+ T細胞応答並びにCD8+ T細胞応答を増強することが示されており(実施例6および7)、かつ(2)B細胞(抗体産生性)応答には、一般にCD4+ヘルパーT細胞の活性が必要である:が故に、本発明の方法によって増強することができる免疫応答は、B細胞応答を含む。
【0066】
本発明のこれらの方法は、インビボであることができ、上記細胞の活性化を阻害するインビボ法と本質的に同様である。加えて、これらは、後者の方法を上で詳述したのと同じ被検者に適用することができる。本方法は、以下のさらなる工程を含むことができる:1つまたは複数の治療を行う工程、動物における免疫応答について試験する工程、および/または当該技術分野においてなじみがある方法によって、反応を、たとえばT細胞応答、CD4+ T細胞応答、CD8+ T細胞応答、または抗体産生B細胞応答として特徴付ける工程。
【0067】
免疫応答性を増強することを要求される場合、関心対象のウイルスの1つまたは複数の形態を実質的に中和する抗体(上記参照)に加えて、補充薬を使用することができる。特に関心対象の免疫応答を増強する方法は、ウイルスEBL(たとえば、mAbs 13E8および11D7)に結合する上記の抗体の1つまたは複数、および関心対象のウイルスの1つまたは複数の形態を実質的に中和する1つまたは複数の抗体、たとえば抗L1R抗体を投与することを含む。
【0068】
免疫応答を増強するために有用なさらなる補充薬は、たとえば適切なウイルス抗原を含む。抗原は、被検者が感染するか、するであろうか、またはするリスクがあり、かつ投与された化合物がウイルス複製増強効果を阻害するウイルスEBLの供与源であるウイルスの成分であることができ、たとえば免疫を増強する方法は、生きているウイルスでのワクチン接種の効率を増強するために使用する場合である(たとえば、ワクシニアウイルスでの種痘に対するワクチン接種)。しかし、さらなる抗原を投与することもできる。このような抗原は、たとえば死滅したウイルス、ウイルスタンパク質、またはウイルスタンパク質のペプチド断片であることができる。抗原は、単独で、またはアジュバント、たとえばコレラ毒素(CT)、大腸菌熱易動性毒素(LT)、変異体CT(MCT)[Yamamoto et al.(1997) J. Exp. Med. 185:1203-1210]、および変異体大腸菌熱不安定性毒素(MLT)[Di Tommaso et al. (1996) Infect. Immunity 64:974-979]と共に投与することができる。MCTおよびMLTは、親分子のものと比較して実質的にアジュバント活性を損なうことなく実質的に毒性を減弱させる点突然変異を含む。その他の有用なアジュバントは、ミョウバン、フロイント完全および不完全アジュバント、並びにRIBIを含む。
【0069】
さらに免疫を増強するために用いることができるその他の補充薬は、MIP-1α、MIP-3β、RANTES、IL-1、IL-2、IL-3、IL-4、IL-5、IL-6、IL-7、IL-8、IL-10、IL-12、IL-13、IL-15、インターフェロン(IFN)-α、β、もしくはγ、顆粒球コロニー刺激因子(G-CSF)、マクロファージコロニー刺激因子(M-CSF)、または顆粒球マクロファージ-コロニー刺激因子(GM-CSF)などのケモカイン、リンホカイン、およびサイトカインを含む。加えて、関心対象の免疫応答を阻害する補充薬、ケモカイン、リンホカイン、またはサイトカインに特異的な抗体を使用することができる。従って、たとえば、それは細胞の免疫応答を増強することが要求される場合、IL-4、またはIL-10に特異的な抗体を補充薬として使用してもよい。一方では、抗体産生B細胞応答を増強することが要求される場合、IL-12またはIFN-γに特異的な抗体を補充薬として使用してもよい。
【0070】
細胞の活性化を阻害する方法の節において上記した化合物のデリバリーおよび投与の全ての方法論(または、これらの明らかなバリエーション)は、本明細書に記載されている全ての補充薬のデリバリーおよび投与のために使用することができることが理解される。これらの補充薬は、化合物の前に、後に、または同時に投与することができる。これらは、化合物と同じか、もしくは異なる経路によって、および同じか、もしくは異なる頻度で投与することができる。
【0071】
本発明のこれらの方法は、遺伝子の免疫化に適用することができる。このような方法では、関心対象の免疫原をコードする異種核酸配列を含むポックスウイルス発現ベクターが、適切な動物(たとえば、ヒト、非ヒト霊長類(たとえば、サルまたはチンパンジー)、ウシ、ウマ、ヤギ、ヒツジ、ブタ、ネコ、イヌ、ウサギ、モルモット、ハムスター、スナネズミ、ラット、マウス、ニワトリ、または本明細書において詳述した任意のその他の種)に対して、当該技術分野において既知の方法によって有効であることが決定された頻度で投与される。ベクター内では、異種核酸配列が転写調節性のエレメント(TRE)に対して動作可能に連結される。TREは、プロモーターおよびエンハンサーを含み、当該技術分野において既知である。本明細書で使用されるものとして、コード配列に「動作可能に連結された」発現制御配列(たとえば、TRE)は、これがコード配列の発現を効率的に制御するように、遺伝子構築物内に組み入れられる。関心対象のポックスウイルス・ベクターは、ワクシニア・ベクター、弱毒ワクシニア・ベクター、カナリア痘瘡ベクター、および伝染性上皮腫ベクターを含む。
【0072】
投与の経路は、好ましくは皮膚(たとえば、乱切によって)、筋肉内、皮下、静脈内、または腹腔内、または粘膜(たとえば、鼻腔内もしくは直腸内)であろう。ベクターの投与の前に、同時に、または後に、上記したerb-B抑制化合物の1つまたは複数が動物に投与される。任意の上記した補充薬を動物に投与することもできる。また、化合物および補充薬の投与の頻度は、当該技術分野における技術のうちの1つによって容易に決定することができる。
【0073】
erb-Bチロシンキナーゼ阻害剤の投与により、ポックスウイルスに感染した動物のCD4+およびCD8+免疫応答を増強したので(実施例6および7を参照されたい)、ポックスウイルス・ベクターを使用する上記の遺伝子免疫化ストラテジーは、免疫化に使用したポックスウイルス・ベクター内の異種核酸配列によってコードされる免疫原に対する細胞性応答(CD8+およびCD4+ T細胞)またはB細胞媒介応答の両方の増強に有効であろう。これらは、多種多様な免疫原に対する治療的、予防的、または非治療的/非予防的な免疫応答を生じさせるために使用することができる。これらの方法論により、複製能のあるポックスウイルス・ベクター(たとえば、ヒトにおけるワクシニア・ベクター)を使用することができ、非複製ポックスウイルス・ベクター(たとえば、ヒトにおいてカナリア痘瘡ベクター)よりも強力な反応を誘導するが、遺伝的免疫化に対して、宿主動物(たとえば、免疫無防備状態の患者)の感染が制御不能となる可能性の懸念が生じる。従って、関心対象の免疫原をコードする配列を含む複製能のあるポックスウイルス・ベクターが被検者に投与される。十分な時間ベクターを十分に複製させて、被検者の強力な免疫応答を開始するために十分な免疫原を生じさせることができる。有効な時間を確立するために、当該技術分野において周知の方法を使用することができる。次いで、1つまたは複数のerb-B抑制化合物が(1つもしくは複数の補充薬を伴って、または伴わずに)被検者に投与される。この投与は、被検者におけるウイルスベクターの力価を減少させるため、および免疫原に対する免疫応答を増強するための両方のために役立つ。
【0074】
関心対象の免疫原は、たとえば微生物(たとえば、ウイルス、細菌、真菌、酵母、または原生動物)抗原、寄生虫(たとえば、線虫)由来抗原、腫瘍抗原、または研究的、治療的、予防的、もしくは診断の目的のための抗体(ポリクローナルまたはモノクローナル)を作製することが要求されるものに対するタンパク質であることができる。
【0075】
得ることができる免疫原をコードする核酸配列に由来する関連した微生物の例は、結核菌(Mycobacteria tuberculosis)、腸炎菌(Salmonella enteriditis)、リステリア菌、癩病菌(M. leprae)、黄色ブドウ球菌(Staphylococcus aureus)、大腸菌(Escherichia coli)、肺炎連鎖球菌(Streptococcus pneumoniae)、ボレリア・ブルグドルフェリ(Borrelia burgdorferi)、肺疫菌(Actinobacillus pleuropneumoniae)、ピロリ菌(Helicobacter pylori)、髄膜炎菌、エンテロコリチカ菌(Yersinia enterocolitica)、百日咳菌(Bordetella pertussis)、ポルフィノモナス・ギンギバリス(Porphyromonas gingivalis)、マイコプラズマ、ヒストプラスマ・カプスラーツム(Histoplasma capsulatum)、クリプトコッカス・ネオフォルマンス(Cryptococcus neoformans)、トラコーマ病原体(Chlamydia trachomatis)、鵞口瘡カンジダ(Candida albicans)、熱帯熱マラリア原虫(Plasmodium falciparum)、三日熱マラリア(Plasmodium vivax)、四日熱マラリア原虫(Plasmodium malariae)、赤痢アメーバ(Entamoeba histolytica)、トキソプラズマ・ブルセイ(Toxoplasma brucei)、トキソプラズマ・ゴンジ(Toxoplasma gondii)、リーシュマニア・メジャー(Leishmania major)、ヒト免疫不全症ウイルス1および2、インフルエンザウイルス、はしかウイルス(measles virus)、狂犬病ウイルス(rabies virus)、肝炎ウイルスA、B、およびC、ロタウイルス、パピローマウイルス、呼吸器合胞体ウイルス、ネコ免疫不全ウイルス、ネコ白血病ウイルス、サル免疫不全ウイルス、および本明細書において詳述した全てのポックスウイルスを含むが、これらに限定されない。関連した微生物免疫原の例は、大腸菌の熱易動性エンテロトキシン[Konieczny et al. (2000) FEMS Immunol. Med. Microbiol. 27 (4):321-332]のBサブユニット、熱ショックタンパク質、たとえばエンテロコリチカ菌(Y enterocolitica)の熱ショックタンパク質60[Konieczny et al. (2000) supra; Mertz et al. (2000) J. Immunol. 164 (3):1529-1537]、および結核菌熱ショックタンパク質hsp60およびhsp70;トラコーマ病原体外膜タンパク質[Ortiz et al. (2000) Infect. Immun. 68 (3):1719-1723];B.ブルグドルフェリ(B. burgdorferi)外面タンパク質[Chen et al. (1999) Arthritis Rheum. 42 (9): 1813-1823];L.メジャー(L. major)GP63[White et al. (1999) Vaccine 17 (17):2150-2161 (およびVaccine 17 (20-21):2755の公表された正誤表)];髄膜炎菌(N. meningitidis)の髄膜炎菌血清型15のPorBタンパク質[Delvig et al. (1997) Clin. Immunol. Immunopathol. 85 (2):134-142];P.ギンギバリス(P. gingivalis)381海馬采タンパク質[Ogawa, (1994) J. Med. Microbiol. 41 (5):349-358];大腸菌外膜タンパク質F[Williams et al. (2000) Infect. Immun. 68 (5):2535-2545];インフルエンザウイルス血球凝集素およびノイラミニダーゼ;レトロウイルス(たとえば、HIV)表面糖タンパク質(たとえば、HIV gp160/120)、またはレトロウイルスのtatもしくはgagタンパク質を含むが、これに限定されない。
【0076】
ポックスウイルス・ベクターに挿入される免疫原をコードする核酸を得ることができる腫瘍の例は、神経組織癌、黒色腫、乳癌、肺癌、胃腸の癌、卵嚢癌、精巣癌、肺癌、前立腺癌、子宮頚癌、膀胱癌、膣癌、肝臓癌、腎癌、骨癌、血液細胞癌、および血管組織癌を含む。関連した腫瘍抗原の例は、前立腺特異的膜抗原(PSMA)[Israeli et al. (1993) Cancer Res. 53 (2):227-230]、MUC-1などのムチン、または腫瘍細胞の表面上に発現されるその他の任意の抗原を含む。
【0077】
ポックスウイルス・ベクターの異種核酸配列は、全長未成熟のタンパク質、全長成熟したタンパク質、免疫原性エピトープを含むタンパク質のセグメント、またはT細胞エピトープを含むペプチド断片をコードすることができる。これらは、単一のポリペプチド配列または複数(たとえば、2、3、4、5、6、7、8、9、10、11、12、15、18、20、25、30以上)のポリペプチドをコードすることができる。異種核酸配列が複数のポリペプチドをコードする場合、ポリペプチドは、1つもしくは複数(たとえば、2、3、4、5、6、7、8、9、または10)の微生物の生物体および/または腫瘍に由来することができる。
【0078】
上記のポックスウイルス・ベクター/erb-B阻害剤のストラテジーでは、遺伝的免疫化に限定されないと理解されるが、免疫原の代わりに、たとえばサイトカインもしくは成長因子(本明細書において列記したもののいずれかなど)、薬物もしくはプロドラッグ、または関心対象の被検者では欠損した酵素、たとえばアデノシンデアミナーゼなどの治療的分子をコードする異種核酸配列を、ポックスウイルス・ベクター遺伝子治療法に一般に適用することができる。
【0079】
ポックスウイルスおよびこれらの使用は、Paoletti [(1996) Proc. Natl. Acad. Sci. USA 93:11349-11353]に更に詳細に記載されており、その全体が参照として本明細書に組み入れられる。
【0080】
特定の処方計画が、特定の疾患に対して治療的、または予防的かどうかを試験するための方法は、当該技術分野において既知である。治療効果が試験される場合、疾患の症状を示す試験集団(たとえば、天然痘を有するヒトまたは実験動物)を任意の上記したストラテジーを含む試験処方計画で治療する。疾患の症状を示す対照群も、偽薬または異なる処方計画で処置する。試験被検者における疾患症状の消滅または減少は、試験処方計画が有効な治療法であることを示すであろう。
【0081】
疾患症状の発症前に同じストラテジーを被検者に適用することによって(たとえば、ワクシニアウイルスでの作為的な感染前の実験動物)、試験処方計画を予防法としての有効性について試験することができる。この状況では、疾患症状の発症の予防、または遅延が試験される。
【0082】
以下の実施例は、本発明を例証することを意味し、限定しない。
【0083】
実施例
実施例1.材料および方法
SPGF D4Rの組換えEGF様ドメインの産生
SPGF D4RのEGF様ドメインを含むポリペプチド(SEQ ID NO:2のアミノ酸残基40〜90)をE.coli DL21細胞において封入体として発現させて、生じるタンパク質をヒトEGFおよびマウスエピレギュリン(EPI)から作製された類似のEGF様ドメインと比較した。便宜のために、これらのEGF様ドメインをそれぞれrecSPGF、recEGF、およびrecEPIと称する。封入体は、6Mグアニジン-HCl溶液に溶解し、ポリペプチドを、6つのヒスチジン残基を含むポリペプチドのN末端のタグ配列を利用してNi2+-NTAカラムクロマトグラフィーによって精製した。部分的に精製したEGF様ドメインをレドックス緩衝液(2mM還元および1mM酸化グルタチオン)中で再び折り畳ませて、ポリペプチドをRP-HPLC(逆相高速液体クロマトグラフィー)を使用してさらに精製した。RecSPGF(8kDa)を15%のナトリウムSDS-PAGEによってrecEGF(6kDA)およびrecEPI(5kDa)と比較し;タンパク質をクーマシーブルーで染色した(図1)。
【0084】
ヒト・ケラチノサイトおよび線維芽細胞の分裂促進因子アッセイ法
正常な初代ヒト真皮線維芽細胞株R2F[Tubo et al.(1987)Oncogene Re.1:407-421]は、15%の子ウシ血清(HyClone, Inc., Logan, UT)および10ng/mlのEGFを補ったDMEM/F12培地(Life Technologies, Inc., Carlsbad, CA)からなる「完全線維芽細胞培地」中で培養した。正常な初代ヒト表皮ケラチノサイト株N[Schon et al. J. Invest. Dermatol. 107:428-438]は、前述したように[Rheinwald et al. (2002) Mol. Cell Biol. 22:5157-5172]、30μg/mlウシ下垂体抽出物(BPE)、0.2ng/mlのEGF、および0.3mMのCacl2を補ったGIBCOケラチノサイト無血清培地(Life Technologies, Inc.)からなる「完全ケラチノサイト培地」中で培養した。
【0085】
分裂促進アッセイ法では、添加したerb-Bリガンドの濃度範囲に応答するこれらの増殖を評価することができるように、EGFを含まず、かつ血清の量を減少した(およびN細胞については、BPEの量を減少して)以外は、関連培地中に低密度で細胞をまくことによって行った。
【0086】
3000個のR2F細胞を9cm2の培養ウェルに、1%の子ウシ血清を含むDMEM/F12培地中にまいた。その翌日、およびプレートにまいて4日後に、ウェルには、recEGFまたはrecSPGF(種々の濃度で)を補った培地か、または完全線維芽細胞培地を与えた。
【0087】
1000個のN細胞を9cm2の培養ウェルに、BPE(15μg/ml)および0.3mMのCacl2を含むGIBCOケラチノサイト無血清培地にまいた。その翌日、およびプレートにまいて4日後に、ウェルには、recEGFまたはrecSPGF(種々の濃度で)を補った培地か、または完全ケラチノサイト培地を与えた。
【0088】
プレートにまいて6日後に、細胞をトリプシン処理することによって培養ウェルの底から剥がして計数した。増殖速度は、1日あたりの集団倍加:[log2(6日後の細胞数/まいた細胞の数)]/6として測定した。
【0089】
チロシンキナーゼ阻害実験については、ヒト包皮線維芽細胞(SC-J)を6ウェルの組織培養プレートにまいて、2%のFBS(ウシ胎児血清)DMEM培地中で一晩「不足させた(starved)」。細胞を37℃で1時間チロシンキナーゼ阻害剤で前処理し、次いで異なる量のrecSPGFで刺激した。18時間後に、細胞を集めて、80%の氷冷エタノール中で、4℃で1時間固定した。リン酸緩衝食塩水(PBS)で一回洗浄した後、細胞を50μg/ml RNase Aを含む2.5μg/mlヨウ化プロピジウムの溶液で37℃において30分間染色した。細胞周期データをFFC解析によって得て、Cell Quest(登録商標)(Becton Dickinson, San Jose, CA)およびModfit(登録商標)(Becton Dickinson)ソフトウェアを使用して解析した。
【0090】
RecSPGFのビオチン化および上皮細胞に対する結合をFFCによって解析した
RecSPGFは、製造業者のプロトコルに従ってECLタンパク質ビオチン化モジュール(Amersham Biosciences, Piscataway, NJ)を使用してビオチン化した。1×105 MB468(またはMB453)上皮細胞をそれぞれの試験試料のために使用した。直接結合アッセイ法のために、1つの細胞の一定分量を1μgの抗erb-B1ブロッキング抗体(mAb 528;Santa Cruz Biotechnology, Inc., Santa Cruz, CA)と共に4℃で1時間インキュベートした。異なる濃度のビオチン化されたrecSPGFの50μl FFC緩衝液(1×PBS/2.5%のFCS/0.02%のNaN3)溶液を試料に添加し、次いで、4℃で45分間さらにインキュベートした。競合アッセイ法のためには、細胞をラベルされていない異なる量のrecSPGF、recEPI、またはrecEGFを含む50μl FFC緩衝液と共に4℃で20分間インキュベートした。ビオチン化されたrecSPGF(25ng)を全ての試料に添加し、次いでこれを4℃で30分間インキュベートした。細胞をFFC緩衝液中で洗浄し、結合したビオチン化recSPGFをフィコエリトリン(PE)結合ストレプトアビジン(Molecular Probes, Eugene, OR)でFFCによって検出した。収集されるデータは、平均蛍光強度(MFI)であった。リガンド結合の阻害は、MFIexpが実験試料で検出されるMFIである場合、(MFIexp-MFImin/MFImax-MFImin)の割合として表してあり、MFIminは、ビオチン化されたrecSPGFが添加されなかったネガティブ対照試料で検出されるMFIであり、MFImaxは、ビオチン化されたrecSPGFが添加されたが、阻害剤が添加されなかったポジティブ対照試料で検出されるMFIである。
【0091】
細胞可溶化物の調製、免疫沈降、およびウエスタン・ブロッティング
10cmの培養ディッシュ内のHeLa細胞(80%〜90%コンフルエント)を37℃で10分間50ng/mlの濃度の種々のerb-B1リガンドで刺激した。ディッシュを氷冷PBSで一度洗浄した。次いで、細胞を1mlの溶解緩衝液(25mMのトリスpH7.4、150mMのNaCl、1%のTritonX-100、1mMのフェニルメチルスルホニルフルオライド、0.35トリプシン阻害剤ユニット/mlアプロチニン、5μg/mlのロイペプチン、10mMのNaF、10mMのβ-グリセロリン酸、および1mMのNa3VO4)でディッシュ内において直接4℃で30分間溶解した。可溶化液をミクロチューブに移し、遠心分離後、0.5mlの可溶化液上清を1μgのヤギ抗EGFRポリクローナル抗体(Santa Cruz Biotechnology, Inc.)および10μlのガンマ結合プラス(商標)ビーズ(Amersham Biosciences)で4℃において一晩に免疫沈降した。ビーズを3回洗浄し、タンパク質を2×SDS-PAGE充填緩衝液に直接ビーズから溶出させた。全細胞の可溶化液または免疫沈降された試料の一定分量を7.5%のSDS-PAGEで分解し、生じるゲルをPVDF(ポリビニリデンフルオライド)膜にブロットした。膜を2.5%のウシ血清アルブミンを含むトリス-緩衝食塩水/Tween-20(TBST)(erb-B受容体に関して)または2%のゼラチンを含むTBST(4G10に関して)中で37℃において少なくとも30分間ブロックした。膜は、製造業者の説明書に従って一次抗体と共に4℃で一晩インキュベートし、洗浄し、1:10,000抗ヤギIgG(erb-B受容体のために)または抗マウスIgG2b(4G10のために)HRPO(西洋ワサビペルオキシダーゼ)抱合体と共に室温で1時間インキュベートした。広範な洗浄後に、膜を化学発光試薬キット(Perkin Elmer Biosystems, Wellesley, MA)およびMRフィルム(Eastman Kodak Co., Rochester, NY)を使用して発色させた。
【0092】
チロシンキナーゼ阻害実験では、HeLa細胞を37℃で30分間50nMで阻害剤で処置し、続いてrecSPGF(50ng/ml)で刺激した。細胞可溶化物を調製し、標準的な方法を使用して、ヤギ抗erb-B1ポリクローナル抗体(Santa Cruz Biotechnology, Inc.)およびガンマ結合プラスビーズまたは抗c-Cbl抗体被覆ビーズ(Santa Cruz Biotechnology, Inc.)で4℃で一晩免疫沈降(上記の通り)に供した。ビーズを洗浄し、2×SDS-PAGE充填緩衝液に溶出した。全細胞の可溶化液または免疫沈降物を抗EGFRまたは抗c-Cblポリクローナル抗体でのウエスタン・ブロッティングによって解析した。
【0093】
recSPGFおよびerb-B受容体の化学的架橋
2.25×106のMB468またはMB453細胞を、受容体インターナリゼーションを阻害するために4℃で30min間0.04%のNaN3で前処理した。次いで、細胞をビオチン化されたrecSPGF(1μg/ml)と共に4℃で30分間インキュベートした。培養ディッシュを氷冷DMEMで洗浄し、ビス(スルホスクシニミジル)-スベラート(BS3)架橋剤を1mMの濃度で添加した。反応混合物を随時振盪しながらさらに1時間4℃でインキュベートした。氷冷PBSで2回洗浄した後、1mlの溶解緩衝液(25mMのトリスpH7.4、150mMのNaCl、1%のTritonX-100、1mMのフェニルメチルスルホニルフルオライド、0.35トリプシン阻害剤ユニット/mlアプロチニン、および5μg/mlロイペプチン)を直接ディッシュに添加し、続いて4℃で30分間振盪した。可溶化液をミクロチューブに移し、遠心分離後、可溶化液の上清を1μg抗erb-B1、抗erb-B3、もしくは抗erb-B4ヤギ・ポリクローナル抗体(Santa Cruz Biotechnology)、または1:40希釈抗erb-B2(Cell Signaling Technology, Beverly, MA)ウサギ・ポリクローナル抗体、および10μlのガンマ結合プラスビーズでの免疫沈降(IP)に供した。試料を4℃で5時間回転させた後に、ビーズを1×TBS/1%のTriton X-100で4回洗浄し、次いで2×SDS-PAGE充填緩衝液中で煮沸した。ビオチン化されたrecSPGFを検出するために、SDS-PAGEおよびストレプトアビジン-HRPOでのウエスタン・ブロッティングを使用した。
【0094】
EGFRに結合するrecSPGFの親和性
recSPGFの125I-ラベリングのためにラクトペルオキシダーゼ触媒法を使用し、recEGFの125I-ラベリングのために、クロラミンT触媒法を使用した。ラベルされたrecSPGFの比活性は、0.54pm/cpmであり、recEGFのものは、0.34pm/cpmであった。MB468細胞は、3×104/0.1ml/96ウェルの濃度でまいた。125IのラベルされたrecSPGFまたはrecEGFを種々の濃度で細胞の50μlの結合培地溶液(0.1%のNaN3を含むL15培地)に4℃で添加し、5時間インキュベートした。上清を回収し、細胞を60μlの氷冷結合培地で迅速に2回洗浄した。次いで、上清および洗浄液を合わせた。50μlの0.5N NaOHをそれぞれのウェルに添加し、プレートを室温で1h間インキュベートして接着細胞を溶解し、さらに60μlの0.5N NaOHで一度洗浄した。可溶化液および洗浄液を合わせた。γカウントを使用して、培養液上清中の遊離リガンドの量および細胞可溶化物中の結合したリガンドの量を決定した。受容体数および親和性は、スキャッチャード解析によって決定した(たとえば、Current Protocols in Immunology. Eds. John E. Coligan et al., John Wiley & Sons. (2001))。
【0095】
Erb-B1インターナリゼーション
HeLa細胞をチャンバ・スライド上で培養しErb-B1リガンド(50ng/ml)によって37℃で10分間刺激した。次いで、細胞を3.7%のホルムアルデヒドで10分間固定して、0.1%のTriton X-100で室温において5min間透過化処理した。細胞を室温で30分間、1%のBSA/PBSのmAb(Santa Cruz Biotechnology, Inc.)、1μgの抗erb-B1とインキュベートした。結合した抗erb-B1 mAbを抗マウスIg-FITC(フルオレッセインイソチオシアネート)抱合体で染色することによって検出した。
【0096】
A431表皮癌細胞を50nM阻害剤によって4℃で30分間前処理した。細胞をビオチン化されたrecSPGF(100ng)と共に4℃で30分間インキュベートした。洗浄後、細胞を37℃で5分間インキュベートし、ホルムアルデヒド中で室温において5分間固定した。ストレプトアビジン-PE染色の後に、種々の試料のMFIsをFFCによって得た。
【0097】
細胞およびウイルス
ワクシニアウイルス株WRを、L929細胞内で増殖させて、ウイルス力価アッセイ法を前述したようにVero細胞において行った[Selin et al. (1994) J. Exp. Med. 179:1933-1943; Selin et al. (1998) J. Exp. Med. 188:1705-1715]。全ての動物感染のために、組織培養上清からの未精製のワクシニアウイルスを使用した。
【0098】
ワクシニアウイルス感染
6〜8週齢の雄C57BL/6(B6)マウスには、200μgの試験抗体および/または1mgのCI-1033のPBS溶液を腹腔内注射した(i.p.)。抗体は一度だけ投与したが、CI-1033は、実験の6〜10日の期間毎日与えた。抗体および最初のCI-1033投与の6時間後に、マウスを4×104 PFU(プラーク形成単位)のワクシニアウイルス鼻腔内に感染させた。毎日マウスの重量を測定し、感染の症状を観察した(1=正常、2=乱れた毛皮;3=2プラス背中を丸めた姿勢;4=3プラスほとんど移動しない;5=4プラスの無気力な最小限の応答)。ウイルス力価は、示した時点で決定した。
【0099】
RNase保護サイトカインアッセイ法
ワクシニアウイルスでの感染の種々の日数後に屠殺したそれぞれのマウスからの肺の一方の葉を、18ゲージ針を通してRLT緩衝液(Qiagen, Valencia, CA)中で破壊した。可溶化液を、QIAshredder(商標)モジュール(Qiagen)を使用してさらにホモジナイズして、製造業者の説明書に従ってRNeasyミニ(商標)・キット(Qiagen)を使用して精製した。同量の総RNA(2μg/マウス)をそれぞれの時点で(4、6、および8日)3匹のマウスからプールした。6μgのRNAを、製造業者の説明書に従ってmCK-2b(商標)テンプレートセット(BD Biosciences-Pharmingen)から作製したプローブとハイブリダイズさせ、保護された断片をDNAシーケンシング・ゲルで分解した。ゲルをX線フィルムに曝露することによってオートラジオグラムを作製した。
【0100】
細胞内サイトカイン染色
単個細胞浮遊液を、種々の方法で処置した感染マウスの脾臓から調製し、0.84%のNH4Cl溶液を使用して溶解することによって赤血球を除去した。ワクシニアウイルス特異的な、IFN-γ産生性の、CD8+ T細胞を、ワクシニアウイルス感染線維芽細胞(MC57G)、非感染対照MC57G線維芽細胞、またはプレートに結合した抗CD3(145-2C11)mAbでインビトロで刺激した後に、5μg/mlでCytofix/CytopermキットPlus(商標)(GolgiPlug(商標), BD Biosciences-Pharmingen)を使用してFFCによって検出した。
【0101】
簡単には、MC57G細胞を5×106 PFUのウイルスで2h間インキュベーションすることによってワクシニアウイルスのWR株を感染させた(M.O.I(感染効率)= 1.2)。その後、実験マウスからの1〜2×106の脾細胞を、96ウェルプレートにおいて2.5×105ワクシニア感染MC57G(VV-MC57G)細胞または非感染MC57G細胞と共に、10 U/mlヒト組換えIL-2(BD Biosciences)、および0.2μlのGolgiPlug(商標)試薬の存在下において37℃で5時間インキュベートした。全てが最近活性化されたCD8+ T細胞を検出するための対照培養には、抗CD3 mAbを含めた。これらの培養において、脾細胞を培養ウェルの底に被覆した精製した抗マウスCD3ε mAb(145-2C11)(5μg/mlの濃度で)と共にインキュベートした(上記で記載されている同じ条件下で)。
【0102】
100μlのFFC緩衝液(HBBS(ハンク平行塩溶液)、2%のFCS、0.1%のNaN3)を含む96ウェルプレート内の1μLのFcブロック(商標)試薬(2.4G2 mAb)と共にプレインキュベーションした後、細胞をCD8α(mAb 53-6.7)およびCD44(mAb IM7)に特異的な蛍光色素ラベルしたmAbを組み合わせて染色した(20分、4℃)。その後に、細胞の固定および透過化処理を行って、抗IFNγ mAbを細胞内に到達させた(XMG1.2; BD Biosciences)。新たに染色された試料は、Becton Dickinson FACS CaliburおよびCellQuestソフトウェア(San Jose, CA)を使用して解析した。
【0103】
痘瘡プラークおよびコメット形成に対するCI-1033の効果の試験
コンフルエントなBSC-40細胞の単層を種々の濃度のCI-1033のRPMI +2%のFBS(RPMI-2%)溶液で3回、室温で30分間前処理するか、またはモックを同一の培養条件下で前処理した。7つの濃度のCI-1033を評価して、CI-1033の濃度につき3つのモック前処理対照を7つの個々の6ウェル組織培養プレートで行った。単層には、痘瘡株ソライメン(Solaimen)の懸濁液を感染させ、その結果それぞれのウェルで50プラークが観察された。プレートを6%のCO2雰囲気において35℃で1時間インキュベートし、単層の均一な感染を確実にするために15分の間隔で揺らした。種菌を除去し、単層を1×RPMI-2%でリンスした。単層をCI-1033を含まないか、または適切な濃度で含む培地で覆い、6%のCO2雰囲気において35℃でで4日間インキュベートした。プラークを免疫組織化学的染色によって解析した。コメットは、大きなプラークとコメット型に結合した2つの連続する小さなプラークよりも大きいものとして定義した。異なる濃度のCI-1033の存在下におけるプラークおよびコメットの数をウィルコクソン順位和検定を用いて対照プレートと比較した。7つのランダムに選択したモック処置したウェル(各々の7枚のプレートから選択したもの)を比較のために使用した。
【0104】
実施例2
SPGFのEGF様ドメインは、細胞増殖および/または細胞生存を刺激する
図2は、ヒトおよびマウスのエピレギュリンのアミノ酸配列の整列、並びにそれぞれオルソポックスウイルス相同分子種D1L、痘瘡、ラクダ痘、ワクシニア、およびサル痘ウイルスに由来するCMP11R、C11R、およびD3Rを示す。哺乳類タンパク質と〜30%の同一性を有するウイルスタンパク質間の高い相同性(〜90%以上)に留意されたい。EGF様ドメイン自体の中では、ウイルスタンパク質配列の事実上の同一性が明らかである(図2)。51残基のEGF様ドメインでは(D1L(SEQ ID NO:1)およびD4R(SEQ ID NO:2)の残基40〜90)、D4Rは、3残基だけがワクシニア相同分子種と異なる(C11R;SEQ ID NO:4)。大痘瘡インド株由来のD1L(図3A;SEQ ID NO:1)は、大痘瘡バングラデシュ株由来の対応する分子の(図3B;SEQ ID NO:2)D4Rとは2つのアミノ酸だけが異なる。
【0105】
一連の機能研究をrecSPGF、recEPI、およびrecEGFで行った。以下の実験によって示されるように、recSPGFは、細胞増殖または細胞生存を刺激する。ヒト初代線維芽細胞(SC-J)を2%のウシ胎児血清(FBS)を含む培地中で6時間不足(starve)させ、次いで種々の濃度のrecSPGF、recEPI、またはrecEGFで18時間刺激した。細胞周期解析は、ヨウ化プロピジウム染色FFCを使用して行った。FFCデータから、細胞周期のS期の細胞の割合をそれぞれの試料について算出し、リガンド濃度の関数としてプロットした(図4)。recSPGFは、recEGFまたはrecEPI(S期に誘導される細胞の最大のパーセントによって評価したもの)よりも低い能力を示したが、recSPGF活性は、その他の成長因子よりも低いモル濃度でプラトーに達した。
【0106】
図5は、SPGFが初代ヒト・ケラチノサイト(N細胞)およびヒト線維芽細胞(R2F)の両方の長期の成長因子であることを示す。細胞を10%のFBS(「完全」)を含む標準的な組織培養培地において、または2%のFBSおよび示した濃度のrecSPGF(「SPGF」)またはrecEGF(「EGF」)を含む組織培養培地において7日間培養し、合計生細胞のカウントを毎日行った。データは、1日あたりの平均集団倍加として表してある。
【0107】
実施例3
SPGFは、Erb-B1受容体と結合する
EGFRの4つのサブタイプがある:erb-B1、erb-B2、erb-B3、およびerb-B4。どのerb-B受容体がSPGFに結合するかについて決定するために、2つの結合実験と、MB453(erb-B1-、-B2+.-B3+、-B4+)およびMB468(erb-B1+、-B2-、-B3+、-B4-)上皮細胞株を使用してビオチン化されたrecSPGFによって行った。第1の実験は架橋解析であった。MB453またはMB468細胞をビオチン化されたrecSPGFと共にインキュベートした。recSPGFとerb-B受容体の架橋は、架橋剤BS3で行った。次いで、可溶化液を調製して免疫沈降に供し、これをHRPO結合ストレプトアビジン(「Sav-HRPO」)(図6、上段パネル)で、または各々のerb-B受容体-特異抗体(図6、中段パネル)でのウエスタン・ブロッティングによって行った。これらの2つの細胞株のerb-B受容体1〜4の発現は、図6の下段パネルに示した表に示してある。図6の上段パネルは、recSPGFがerb-B1のみに検出可能な程度に結合することを証明する。
【0108】
化学的架橋の結果と一致して、図7は、抗erb-B1 mAbが、MB468上皮細胞に対するrecSPGFの結合をブロックすることを示す。MB453細胞は、erb-B1を発現しないので、これらに対するrecSPGFの結合は検出されなかった(図7)。
【0109】
結合阻害実験では、erb-B1に対するSPGFの結合は、EGFよりもわずかに弱いが、エピレギュリンよりも強いことを示す(図8)。
【0110】
EGFR1発現細胞に対するrecSPGF結合の親和性の測定は、スキャッチャード解析によって得られた(図9)。MB468細胞を96ウェルマイクロタイター組織培養プレートのウェルにまいた。125IラベルされたrecSPGFを種々の濃度で添加して、プレートを4℃で5時間インキュベートした。親和性(Kd)および受容体数は、実施例1に記載したように算出した。この解析では、MB468細胞あたり約5.5×104高い親和性(Kd = 0.14nM)のerb-B1受容体および8.8×106の低い親和性(Kd =258 nM)のerb-B1受容体があることを示した。
【0111】
erb-B1受容体は、刺激されていない細胞をerb-B1(図10A)に特異的な抗体で染色したときに得られるリニアな蛍光のほとんど均一なパターンによって証明されるとおり、通常上皮細胞の表面にかなり一様に分布した。しかし、recSPGF、recEPI、またはrecEGFの添加の10分後に、受容体は迅速に内部に取り入れられた(それぞれ、図1OB、1OC、および図10D)。
【0112】
recSPGFによって活性化されるerb-B1の迅速インターナリゼーション、およびチロシン・プロテインキナーゼ・リン酸化がアクチンおよびエズリンを含む細胞突出の形成を経てIMVの侵入を活性化する能力[Locker et al. (2000) Mol. Biol. Cell 11:2497-2511]は、両方とも細胞に対する痘瘡ビリオンの侵入を容易にするために貢献する。天然には、侵入前の細胞表面に対する痘瘡ビリオンの最初の結合は、少なくとも一部において、SPGF-erb-B1の相互作用を経ることができる。細胞に対するビリオンの侵入を可能にすることに加えて、上記の活性化イベントは、宿主細胞をより効率的なウイルス複製「工場」にする。
【0113】
実施例4
SPGFのEGF様ドメインは、細胞におけるタンパク質チロシン・リン酸化を刺激する
RecSPGFは、EGF受容体およびこれらの細胞基質のリン酸化を刺激した。無処置(-)またはrecSPGF(「SPGF」)、recEPI(「EPI」)、もしくはrecEGF(「EGF」)で処置したHeLa細胞からの細胞全体の可溶化液を4G10抗ホスホチロシン・モノクローナル抗体でのウエスタンブロット法(WB)に供した(図11A)。加えて、HeLa細胞可溶化液を抗EGFRポリクローナル抗体で免疫沈降(IP)し、抗ホスホチロシン抗体でのWBに供した(図11B)。EGFR自体およびその種々の細胞基質の両者のチロシン残基は、試験した3つの成長因子によって、同一でない場合でも、同様の様式でリン酸化された。
【0114】
実施例5
SPGFのEGF様ドメインによるタンパク質チロシン・リン酸化の刺激は、キナゾリンに基づいた化合物によって阻害される
SPGFで刺激される活性に対するいくつかのキナゾリンに基づいたチロシンキナーゼ阻害剤の効果を評価するために、一連の実験を行った。図12は、 3つの4-アニリノキナゾリンチロシンキナーゼ阻害剤(PD 153035、PD 168393、およびCI-1033)の化学構造を、同様に一般的な4-アニリノキナゾリン構造を示す。erb-B1受容体キナーゼ・ドメイン単独での、およびそのような阻害剤の1つとの複合体の構造は、これらの4-アニリノキナゾリンがどのようにキナーゼ・ドメインのATP結合ポケットと結合するかを示している[Stamos et al. (2002) J. Biol. Chem. 277:46265-46272]。図12に示すように、PD153035およびPD168393は、同じ4-(3'-ブロモ-アニリン)環を有するが、キナゾリン環に付着されるこれらのR3およびR4基が異なる。特に、PD168393は、erb-B1 Cys773をアルキル化することができる6位にアクリルアミドを有し、その結果、阻害剤は、1:1のモル比で不可逆的にerb-B1と結合する。チロシンキナーゼ活性なerb-B2およびerb-B4分子は、それぞれCys784およびCys778に相当するシステイン残基を有しており、修飾のためにターゲットすることができる。対照的に、PD153035は、主に疎水性の力を経て可逆的な様式で結合する。CI-1033は、PD168393同様に、6位(R3)にアクリルアミド付加物を有し、したがって、PD168393同様、Cys773を経て不可逆的にerb-B1と結合する。CI-1033は、加えて、7位(R4)に可溶性のモルフォリン側鎖を有する。CI-1033は、erb-B1、erb-B2、およびerb-B4に対して、それぞれ0.8、19、および7nMのIC50値を示す[Allen et al. (2002) Sem. Oncol. 29:11-21]。
【0115】
第1の実験では、2つのキナゾリンに基づいた化合物(PD168393およびPD153035)およびピリドピリミジンに基づいた化合物(PD158780)を、これらがチロシン・リン酸化を阻害する能力について試験した。全3つの化合物は、Calbiochem, San Diego, CAから得られ、Calbiochemのカタログでは、これらの化学構造を示している。HeLa細胞を24ウェル組織培養プレートのウェルにまき、阻害剤化合物を50nMの終濃度に添加した。細胞を30分間培養し、recSPGF(50ng/ml)をネガティブ対照試料(図13において(-)と示す)以外の全てのウェルに添加した;図13に(+)として示した試料は、recSPGFを受けたが、阻害剤を受けさせなかった。次いで、細胞をさらに10分間37℃で培養し、次いで洗浄した後に溶解した。細胞可溶化物を10%のSDS-PAGEによって分離して、SDS-PAGEゲルをPVDF膜にブロットした。膜を4G10抗ホスホチロシンmAbに曝露し、その結合をHRPO結合抗マウスIgG2bポリクローナル抗体によって検出した。膜をECL(化学発光の増強)によって発色させた。図13のデータは、recSPGFによって活性化されるチロシン・リン酸化が全3つの化合物(PD168393、PD153035、およびPD158780)によって阻害されることを示す。
【0116】
recSPGFを媒介した細胞増殖および/または細胞生存の増大に対するチロシンキナーゼ阻害剤の効果を試験した。図14Aに示すように、種々の濃度のrecSPGFでヒト線維芽細胞を一晩刺激した後、S期に入っている細胞の数は、8%に近づいた。50nM濃度の示した阻害剤で37℃において1時間細胞を前処理すると、このDNA合成の増加をブロックする。図14Bは、erb-B1のチロシン・リン酸化(150kDバンド)並びにそのリン酸化のさらなる基質(120kD、80kD、60kDおよび55kD)を阻害したものと同じ化合物が、HeLa細胞においてrecSPGFによって活性化されたことを示す。SrcファミリーPTK特異的阻害剤PP2は、10μMで使用したときでも、これらのerb-B1基質のリン酸化をブロックする能力をほとんど有さなかった。対照的に、50nMの濃度でPD168393またはCI-1033を添加すると、主にrecSPGFでトリガーされるリン酸化を防止したが、PD153035で観察される効果は、部分的なものだけであった。これらの結果は、キナーゼ活性の可逆的阻害剤が、試験される50nM濃度において不可逆阻害剤と比較してタンパク質チロシン・リン酸化のブロックにあまり効率的ではないことを示唆する。チロシンキナーゼ・リン酸化の同様の阻害は、ワクシニアウイルス成長因子(VGF)によって活性化されるリン酸化でも観察された(データ示さず)。
【0117】
図10Bは、recSPGFと37℃で15分間インキュベーションする前および後に、免疫蛍光顕微鏡観察によって検出されるヒトHeLa上皮細胞のerb-B1受容体の分布を示す。成長因子の非存在下では、erb-B1の分布は、主に膜に限定される。対照的に、SPGF曝露後では、erb-B1分子は迅速にインターナライズされて、点状の細胞内蛍光の凝集として現れる。チロシンキナーゼ阻害剤がSPGFの能力に影響を及ぼしてerb-B1をダウンモジュレートするかどうか判断するために、A431細胞を阻害剤で前処理し、ビオチン化されたrecSPGFと共にインキュベートし、次いで固定した。次いで、細胞表面に結合したerb-B1リガンドをストレプトアビジン-PEによって視覚化し、蛍光をFFC解析によって定量化した。図14Cは、阻害剤の非存在下では、MFIは、〜110であったが;対照的に、不可逆的erb-B阻害剤は、MFIを〜3倍(300〜350)に増大し、可逆的erb-B1阻害剤PD153035は、MFIを240に増大したことを示す。srcファミリーキナーゼ阻害剤PP2は、わずかにerb-B1表面のコピー数に影響を及ぼすだけであった。
【0118】
erb-B受容体は、2つの経路によって除去される:1)リガンド依存的なエンドサイトーシス、およびc-Cblユビキチンリガーゼ機構を含む分解[Levkowitz et al. (1999) Mol. Cell 4:1029-1040]、並びに2)ストレスで誘導されるerb-B受容体と結合したシャペロンおよび関与するプロテアソームのプロテイナーゼのシャフリング[Xu et al. (2001) J. Bio. Chem. 276:3702-3708]。最近の研究は、CI-1033によるerb-B2キナーゼの阻害が、第2のプロセスを経てerb-B2のダウンレギュレーションを促進することを示す[Citri et al.(2002) EMBO J. 21:2407-2417]。しかし、CI-1033がerb-B1の表面発現を増強することを考えると、この知見は、リガンド特異的なSPGF erb-B1発現に対するCI-1033の効果は、構成的erb-B2発現に対するものと類似しないことを示唆する。
【0119】
4-アニリノキナゾリンがc-Cblのerb-B受容体との相互作用に影響するかどうかを試験するために、阻害剤を前処理したか、または無処置のrecSPGF刺激したHeLa細胞を溶解し、可溶化液を抗erb-B1抗体で免疫沈降した。その後、erb-B1分子の発現を定量化するために抗erb-B抗血清で、またはerb-B1結合に対する化合物の効果を評価するために抗c-Cbl抗体でのいずれかでウエスタン・ブロッティングを行った。図14Dに示すように、刺激されていない細胞(-)と比較して、recSPGFでトリガーされたHeLa細胞(+)は、迅速な細胞内分解のために合計erb-B1が少なくなる。FFC解析(図14C)と一致して、PP2は、このプロセスに対してほとんど影響を有さなかった。他方、PD153035、PD168393、およびCI-1033の前処置では、recSPGFの添加にもかかわらず、免疫沈降される合計erb-B1タンパク質が増大し;recSPGF刺激されていない対照細胞から得られたもの以上にerb-B1の数倍の増加を示した。後者の結果は、阻害剤によるerb-B1の構成的インターナリゼーションまたは分解の封鎖を意味する。さらに重要なことに、recSPGFで誘導されるc-Cblのerb-Bとの結合は、4-アニリノキナゾリンによってブロックされたが、PP2によってブロックされなかった。c-Cbl/erb-B1複合体形成の減少は、平行したc-Cbl免疫沈降およびウエスタン・ブロッティングによって示される合計細胞c-Cblレベルの減少に対して二次的であった。その代わりに、これらのerb-Bキナーゼ阻害剤は、SPGFの結合後の誘導性のc-Cblのerb-B1との結合を防げた。
【0120】
実施例6
チロシンキナーゼ阻害剤のインビボでの抗ウイルス効果
ワクシニアWRのVGF遺伝子の破壊は、インビボでワクシニアウイルスの病原性を減少させ、ウイルス増殖を〜10,000倍まで緩徐化することを示した[Buller et al. (1988) J. Virol. 62:866-874]。上に示したように、CI-1033は、SPGF刺激されたerb-B1で駆動される細胞増殖または生存、受容体を媒介したチロシン・リン酸化、インターナリゼーション、および分解をブロックするので、erb-B1キナーゼ阻害剤は、インビボでのオルソポックス成長因子活性を減らすことができるようであった。この可能性を試験するために、致死的に鼻腔内にワクシニアWRを与えたB6マウスの臨床経過に対するCI-1033の効果を調査した[Chen et al. (2001) Nat. Immunol. 2:1067-1076]。図15Aに示したように、全ての無処置の動物は、劇症性の急性肺炎による感染の7日後までに死んだ。対照的に、感染の6h前に開始して毎日50mg/kgでCI-1033をi.p.投与すると死を防止した。本療法は、ワクシニア細胞内成熟ウイルス粒子(IMV)を中和することが既知の抗L1RワクシニアウイルスmAb(7D11)の一回の200μgのi.p.用量の投与と同程度に有効であった[Hooper et al. (2000) Virol. 266:329-339]。CI-1033と抗L1Rの併用投与でも、保護された。感染後の動物の臨床モニタリングでは、抗L1R mAbまたはCI-1033処置が有意に総合的症状を減少させたが、併用療法ほど効率的ではなかったことを示した(図15B)。
【0121】
動物のコホートを感染の6日後に屠殺し、これらの肺を検査した。処理群間の総肺重量の違いを図15Cに示してある。未処理の動物では、多発性出血、および正常な、非感染性のB6マウスの約2倍の重量を有する浮腫状の肺を有した。この病状は、抗L1RまたはCI-1033単独での処置によっていくらか減弱されたが、CI-1033プラス抗L1Rの併用によって劇的に改善された。病理学的な所見と一致して、肺におけるワクシニアウイルスの力価は、8日目にアッセイしたときに、抗L1RまたはCI-1033単独によってやや減少されたが、併用治療によっては本質的に除去された。無処置の動物は、全て8日までに死んだので、7日のウイルス肺力価を比較のために使用した。また、図15Dは:(i)抗SPGF mAb(13E8)(これは、VGFと交差反応する(出願中のU.S. Application Serial No. 10/429,685を参照されたい))での免疫療法;および(ii)抗L1Rプラス抗SPGF mAbsでの二重免疫療法の結果を図示する。抗L1RプラスCI-1033処置で観察されるウイルス力価の、さらなる2つの経過記録の減少(抗L1R mAbプラス抗SPGF mAb処置と比較して)は、mAbの可逆的な結合と比較して、CI-1033の不可逆的結合の結果によるであろう[Fry et al. (1998) Proc. Natl. Acad. Sci. USA 95:12022-12027]。阻害剤の不可逆的結合により、阻害剤の可逆的結合よりも完全なerb-B1経路の封鎖が生じるのであろう。
【0122】
併用した抗L1R mAbプラス抗SPGF mAb免疫療法実験(出願中のU.S. Application Serial No. 10/429,685号を参照されたい)では、ウイルス力価が感染の後6日まで減退しなかった。この緩徐な時間経過は、抗体のウイルス中和効果ではなくむしろウイルスに対して適応性T細胞応答が発生していることを示唆した。図示はしていないが、同様の動態は、抗L1R mAbプラスCI-1033の併用療法によっても観察された。これらの知見は、CI-1033が、単独でまたは抗L1R mAbと併用して、T細胞応答を増大するであろうことを示唆する。この概念を試験するために、肺におけるT細胞で調節されるサイトカイン産生を感染の4、6、および8日後にRNase保護解析によって試験した。図16Aに示したように、CI-1033を単独で与えられたマウスでは、インターロイキン-1β(IL-1β)、インターロイキン-1受容体アンタゴニスト(IL-1Ra)、およびIFN-γ mRNAのレベルが、無処置のマウスと比較して増大し、6日にピークに達した。たとえば、IFN-γ RNAのレベルは、6日目で、対照感染動物のものと比較して、CI-1033処置したマウスの肺のよりも4.1倍大きかった。しかし、インターロイキン6(IL-6)、インターロイキン10(IL-10)、インターロイキン12(IL-12)、またはマクロファージ遊走阻止因子(MIF)mRNAのレベルに対する効果は、CI-1033を与えらたマウスの肺においても全く見られなかった。
【0123】
肺サイトカインのレベルと一致して、VV-MC57G細胞に対する全身性の抗原特異的T細胞応答は、脾臓のCD8+CD44+ T細胞の細胞内IFN-γ産生によって評価すると、対照の無処置のマウスと比較して、CI-1033処置したマウスにおいて実質的に増強された(図16B)。VV-MC57G細胞によって活性化されるIFN-γ産生CD8+CD44+ T細胞の割合は、感染8日後に10倍に増大した。この増加は、VV-MC57G細胞によって活性化されるIFN-γ産生CD8+CD44+ T細胞の絶対数の21倍の増加に対応した(1.85×105対8.5×103細胞);この効果は、無処置のワクシニアウイルス感染マウスにおけるリンパ球減少によるものであった。同じ実験において、抗L1Rおよび抗L1R + CI-1033処置したマウスについて、それぞれ22.7および48倍の増大が観察された。抗L1R処置により、抗L1R + CI-1033の二重療法よりも肺におけるIFN-γサイトカインRNAを生じたことから、前者の処置での持続的なウイルス負荷を示しているのであろう(図15D)。マウスの抗L1Rおよび抗L1R + CI-1033処置後のIFN-γ産生CD4+CD44+ T細胞の比率に、よりわずかな、しかし有意な増大が見られた(データ示さず)。
【0124】
実施例7
VGFに特異的な抗体によるワクシニアウイルスに対するインビボ免疫の増強
同時継続中のU.S. Application No. 10/429,685では、CI-1033と同様に、痘瘡SPGF D4Rに特異的で、かつVGFと交差反応を起こすmAb(13E8)は、抗L1R mAbの保護効果を増強したことが示された。本明細書において、CI-1033(実施例6を参照されたい)に関し、mAb 13E8がインビボでワクシニアウイルスに対するT細胞免疫を増強し、インビボで抗L1R mAbによって得られるT細胞免疫の増強を増大することが示される。
【0125】
ワクシニアウイルスは、IFN-γ産生T細胞によってインビボで制御されるので、そのレベルは、感染の6〜8日後にピークに達し[Ruby et al.(1991) Lymphokine Cytokine Res. 10:353-358]、13E8および抗L1R mAbで見られるウイルス減少の動態(同時継続中のU.S. Application No. 10/429,685)は、適応免疫が関与することを示唆した。従って、CD8+ T細胞応答を、ワクシニアウイルスに感染したマウスを以下によって処置した7日目に検査した:(a)対照1A3 mAb、(b)13E8 mAb、(c)抗L1R mAb;または(d)13E8および抗L1R mAbsの組み合わせ。VV-MC57Gスティミュレーター細胞で、または抗CD3 mAbで(これらは最近活性化された全てのT細胞を刺激する)、5h間インビトロで刺激した新たに単離された脾臓T細胞において、細胞内IFN-γレベルを解析した。実験では、ナイーブ非感染マウスと比較して、以下の順序で、ワクシニアに感染した処理群のCD44+CD8+ T細胞のIFN-γ反応を増大することを示した:1A3 mAb(対照)、13E8 mAb、抗L1R mAb、および抗L1R mAb + 13E8 mAb(図17C)。この実験およびその他の実験において、脾臓は、抗L1R mAbおよび抗L1R mAb + 13E8 mAb群において拡大し、その結果、たとえば抗L1R mAb + 13E8 mAbを組合せて処置したマウスの脾臓あたりのワクシニアウイルス特異的IFN-γ産生細胞の総数は、1A3処置したマウスよりも11倍高かった(0.1が×印を付ける3.3±1.4×105対0.3±0.1×105、群あたりn=3)。
【0126】
マウスの抗L1Rおよび抗L1R + 13E8 mAb処置後のIFN-γ産生CD4+CD44+ T細胞の比率には、より少ないが、しかし、有意な増大が見られた(データ示さず)。
【0127】
抗L1R + 13E8 mAb-処置マウスにおける活性化されたIFN-γ産生脾臓のT細胞のレベルの増強は、肺感染と戦うために肺に移動する能力がある細胞の貯蔵所を示唆し;CD44(T細胞刺激によってアップレギュレートされる活性化マーカー)は、炎症部位にホーミングする白血球に関与するリサイクル可能なヒアルロナンの受容体である点に留意されたい[DeGrendele et al. (1997) Science 278:672-675]。この可能性を試験するために、肺を非感染の、または感染7日、およびmAb処置したマウスから集めて、組織切片のヘマトキシリン-エオジン染色によって検査した。肺胞腔が容易に識別可能であり、細気管支の上皮は無処理であった非感染マウスからの肺と比較して、感染した対照mAb処置したマウスからの肺は、壊死性の細気管支上皮および重篤な肺胞浮腫によって特徴づけられた。これは、ワクシニアウイルスに感染した肺の重量が、非感染肺の約2倍であることを反映した(.35 ± .04 g対 .21 ± .01g、n = 3)。肺胞腔は、これらが好酸性物質で満たされたという点で、除去され;あるとしても、わずかな実質細胞の浸潤が存在した。細胞浸潤の程度は、13E8 mAb処置マウスにおいて幾分増大され、抗L1R mAb処置マウスにおいてより増大され、および組み合わせたmAbで処置したマウスにおいて最も劇的に増大された。二重mAb処置群において、肺胞腔は、開放性であり、細気管支上皮は、正常であった。細胞浸潤する細胞は、大部分がCD8+ T細胞であった。3つの抗L1R mAb処置したマウスのプールされた肺から単離された白血球は、25%のCD8+および5%のCD4+ T細胞であり、抗L1R mAbプラス13E8 mAb処置したマウスから単離されたものは、34%のCD8+および8%のCD4+ T細胞であった。従って、非中和性13E8 mAbは、脾臓および肺のT細胞応答の増大に付随して、ワクシニアウイルスのクリアランスを増強する。
【0128】
先天性免疫、いわゆる急性期反応の全身性の炎症性の成分は、生物体の統合性が破壊されたときに迅速に誘導される[Yoo et al. (2003) Proc. Natl. Acad. Sci USA 100:1157-1162]。上皮細胞を含むセンチネル細胞は、このプロセスの間に、IL-1βおよび/またはIL-6を産生する[Schluns et al. (1997) J. Immunol. 1158:2704-2712]。13E8 mAbによるerb-B1刺激の遮断は、ウイルスによってコードされる分子による、その後のサイトカイン調節不全、およびそれに伴う免疫抑制の回避を補助するであろう[Diehl et al. (2002) Mol. Immunol. 39:531-536; Tanaka et al. (2001) J. Mol. Cell Cardiol. 33:1627-1635]。さらに、EGFおよびTGFα(両方ともerb-Bリガンド)は、インビボで浸出液を誘導することができる[Ohmura et al. (1990) Cancer Res. 50:4915-4917]。SPGFおよびVGFなどのELGFは、類似の病態に寄与している可能性があり、抗ELGF抗体(たとえば、l3E8 mAb)によるその阻害は、このようなオルトポックスウイルス感染に対する抗体のさらなる治療活性を構成すると考えられる。
【0129】
実施例8
タンパク質チロシンキナーゼ阻害剤は、痘瘡ウイルスの細胞内形成および細胞間伝播を阻害する
痘瘡ウイルス増殖に対するCI-1033の効果を検査するために、BSC-40細胞のコンフルエントな単層を、種々の濃度のerb-B阻害剤の存在下または非存在下において痘瘡株ソライメン(Solaimen)約50プラーク形成単位に感染させて、インビトロで4日間培養した。図18Aに示すように、CI-1033の濃度を増大すると、個体のプラークのサイズおよびコメット形成を激減させるが、プラーク数に対しては最小限の効果を有した。プラーク・サイズおよびコメット形成に対する効果は、滴定可能であり(図18A、18B、および18C);CI-1033(10μM)の最も高い濃度では、極めて小さなプラークがかすかに見えるだけであった。それぞれの濃度でのプラークの総数は、7つのランダムに選択した対照ウエルと比較したときに有意に異ならなかった(図18Aおよび18B)。
【0130】
コメットは、細胞外外膜ウイルス(EEV)形成および播種を示す[Payne (1980) J. Gen. Virol. 50:89-100]。図18Cは、500nM以上のCI-1033濃度におけるコメット(p<0.05)の数の統計学的に有意な減少を示す。これらの集約した結果は、CI-1033が初感染ではなく、むしろEEVのウイルス形態形成をブロックすることを示唆する。特に、EEVの放出は、阻害に感受性である。
【0131】
プラーク・サイズに対してあまり重大ではない効果が見られ、より高い濃度のCI-1033で定性的に見えることから、細胞結合内外膜ウイルス(CEV)形成に対する効果を示している。オルトポックスウイルスは、出現するために微小管およびアクチンフィラメントの両方を使用し[Hollinshead et al. (2001) J. Cell Biol. 154:389-402; Smith et al. (2002) J. Gen. Virol.83:2915-2931]、CEVは、細胞からそれ自体が駆出するためのアクチン尾部を誘導する。erb-B1活性化により、アクチンミクロフィラメント系を再編成するので[Lynch et al. (2003) J. Biol. Chem. 278:21805-21813]、これは、細胞結合ウイルスおよび細胞外外膜ウイルスの両方の放出をブロックして、二次的なウイルスの細胞間の伝播を遅延させ、それ故、個体のプラークのサイズを制限している可能性が高いと思われる。
【0132】
EEVを経たウイルスの広範囲の伝播(コメット形成の阻害で示すとおり)に対するより重要な関連した効果は、さらなる妨害機構の可能性を示唆する。erb-Bエンドサイトーシスはまた、アクチン細胞骨格にリンクしているので[Lynch et al.]、erb-B1キナーゼ阻害も同様に、この機構を経て表面EGFR発現を維持するために役立っているかもしれない。細胞のerb-Bリガンドまたはその他の受容体を経たerb-B1のトランス刺激によるerb-B活性化、並びにその下流のポックスウイルス出現のために使用される細胞骨格エレメントに対する効果をブロックするCI-1033の能力は、抗SPGF mAbでは観察されないプラーク形態形成に対する重大な効果の根拠となる可能性が高い(データ示さず)。
【0133】
要するに、併用免疫療法/化学療法は、ウイルス力価の減少およびT細胞免疫の刺激に非常に有効である。本発明は、いずれの特定の作用機序によっても限定されないが、後者が抗炎症性のウイルス産物の生成、並びにウイルス成長因子に依存的な上皮細胞におけるウイルスのDNA複製後のラウンドを防げた結果であろうと推測することができる[Alcami (2003) Nat. Rev. Immunol. 3:36-50; Seet et al. (1998) Ann. Rev. Immunol. 21:377-423]。抗ウイルス化学療法のための標的としての宿主細胞シグナリング経路の遮断の使用は、典型的には、病原体それ自体に対して向けられたその他のアプローチとは別である。宿主にターゲットされたストラテジーの1つの利点は、薬物抵抗性を発生し得ないことである。さらに、抗SPGF mAbsとは異なり、その他の密接に関連した病原性因子(すなわち、VGF対SPGF)との交差反応性が保証されない場合に、erb-Bチロシンキナーゼ阻害剤は、種々のerb-Bタンパク質チロシン・ファミリー数に対して免疫学的に異なったリガンドを潜在的にブロックすることができる。たとえば、VGFおよび粘液腫ウイルス成長因子(MGF)は、異なったerb-B受容体に結合するので[Tzahar et al. (1998) EMBO J. 17:5948-5963]、このより広範な標的活性が関連している。
【0134】
CI-1033の単独投与が、オルソポックス(たとえば、天然の、またはバイオテロリストが蔓延させた天然痘)による「実際の」感染を遅延させるために十分であろう可能性が高いと思われ、オルソポックス粒子の吸入の用量は、本研究において使用したものよりも有意に少ない可能性が高い。そのCI-1033は、T細胞応答を増大すると共に、感染の総合的症状も減少させることから:(a)ワクシニアワクチン接種と併用した免疫刺激薬として;または(b)ワクチン接種の合併症の治療における使用も示唆される。
【0135】
ポックスウイルスおよびこれらのerb-Bに対する成長因子に加えて、B型肝炎ウイルスおよびEBウイルスも、erb-B1転写を調節不全にする産物をコードする遺伝子を有する[Menzo et al. (1993) Virol 196:878-882; Miller et al. (1995) J. Virol. 69: 4390-4398]。さらに、ニワトリ赤芽球症ウイルスなどのRNA型腫瘍ウイルスも、erb-B1シグナリングを利用する[Adelsman et al. (1996) J. Virol. 70:2533-2544]。さらに、機能的なEGF受容体は、宿主細胞の効果的なレオウイルスおよびヒトサイトメガロウイルス(CMV)感染症に必要なことが報告された[Strong et al. (1993) Virol. 197:405-411; Wang et al. (2003) Nature 424:456-461]。これらの知見は、ひとまとめにして、複数のウイルス感染が、erb-Bを媒介したシグナル・ネットワークに結合しているであろうことを示唆する。erb-B受容体を構成的にまたは誘導性に発現する上皮細胞は、体における生得的なセンチネル細胞系の成分である[Yoo et al.(2003) Proc. Natl. Acad. Sci USA 100:1157-1162]。erb-B阻害剤は、オルトポックスウイルス感染よりも感染症においてより広範に適用することができる。ウイルスの病原性因子によって利用される細胞シグナリング経路の化学阻害剤の開発により、一般に感染症の制御に対して新たなアプローチがもたらされるであろう。
【0136】
本発明の多くの態様を記載した。それにもかかわらず、本発明の精神と範囲から逸脱することなく、種々の修飾を行ってもよいことが理解されるであろう。従って、その他の態様も、請求項の範囲内にある。
【図面の簡単な説明】
【0137】
【図1】マウスのエピレギュリン(「Epi」)の上皮細胞成長因子(EGF)様ドメイン、ヒトEGF、および大痘瘡バングラデシュ株天然痘成長因子(「SPGF」)の組換え形態のクーマシーブルー染色したSDS-PAGE(ドデシル硫酸ナトリウム・ポリアクリルアミドゲル電気泳動)ゲルの写真である。これらの組換えタンパク質は、本明細書において、それぞれrecEGF、recEPI、およびrecSPGFと称される。分子量(MW)マーカーのゲルの位置は、写真の左上に示してあり、値は、MW×10-3を示してある。
【図2】ヒト・エピレギュリン(SEQ ID NO:6)およびマウスエピレギュリン(SEQ ID NO:7)、並びにそれぞれ大痘瘡インド株、ラクダ痘、ワクシニア、およびサル痘ウイルス由来のオルソポックスウイルス相同分子種D1L(SEQ ID NO:1)、CMP11R(SEQ ID NO:3)、C11R(SEQ ID NO:4)、およびD3R(SEQ ID NO:5)のアミノ酸配列の整列(最適な相同性のために)の描写である。リーダー配列および膜貫通領域に対応する領域を示してあり、EGF様ドメインで最も保存されたセグメントを枠に入れ、「EGF」をラベルした。
【図3】図3Aおよび3Bは、それぞれ大痘瘡インド株(D1L)(SEQ ID NO:1)および大痘瘡株バングラデシュ(D4R)(SEQ ID NO:2)のSPGFのアミノ酸配列の描写である。2つの配列が異なる位を太字で示して、下線を引いてある。
【図4】示した濃度のrecEGF(「EGF」)、recEPI(「EPI」)、およびrecSPGF(「SPGF」)の18時間の培養後での細胞周期のS期のヒト初代線維芽細胞(SC-J)の割合を示す線グラフである。
【図5】10%のウシ胎児血清(FBS)を含む標準的な組織培養培地中(「完全」)か、または2%のFBSおよび示した濃度のrecSPGF(「SPGF」)またはrecEGF(「EGF」)を含む組織培養培地中のいずれかでの培養の間の、初代ヒト・ケラチノサイト(「Nケラチノサイト」)および初代ヒト線維芽細胞(「R2F線維芽細胞」)の増殖を示す棒グラフである。
【図6】MB453細胞(「453」)またはMB468細胞(「468」)からの免疫沈降物のウエスタンブロットの一対の写真である。免疫沈降物は、ビオチン化されたrecSPGFと共に細胞をインキュベートすることによって調製して、架橋剤BS3に細胞を曝露し(細胞表面上のerb-B受容体に結合したビオチン化されたrecSPGF分子を架橋するため)、細胞を溶解して、および4つのerb-B受容体、すなわちerb-B1(「B1」)、erb-B2(「B2」)、erb-B3(「B3」)、またはerb-B4(「B4」)に特異的な抗体で可溶化液の一定分量を免疫沈降(「IP」)することによって調製した。2つの2回のウエスタンブロットを免疫沈降物から作製した;一方は、西洋ワサビペルオキシダーゼ複合ストレプトアビジン(「Sav-HRPO」)(図6、上段パネル)で染色し、他方は、各々のerb-B受容体-特異抗体(図6は、中段パネル)で染色した。図6の下段パネルは、MB453(「453」)およびMB468(「468」)細胞による4erb-B受容体の発現を示す。
【図7】erb-B1に特異的な阻止抗体の非存在下(白丸)および存在下(「+mAb」;黒丸)での、ビオチン化されたrecSPGF(フィコエリトリン(PE)結合ストレプトアビジンによって検出)(「SPGF」)の示された量(「ng」)のMB468細胞(「対468」)に対する結合を示す線グラフである。データは、平均蛍光強度(「MFI」)として示してあり、これは蛍光フローサイトメトリー(FFC)によって決定した。また、erb-B1を発現しないMB453細胞(「対453」)に対するrecSPGFの結合を試験した(「対453」;白三角)。
【図8】示した濃度のrecSPGF(「SPGF」)、recEPI(「EPI」)、およびrecEGF(「EGF」)がMB468細胞に対するビオチン化されたrecSPGFの結合を阻害する相対的な能力(%阻害として表した)を示す線グラフである。
【図9】MB468細胞に対するrecSPGF結合のスキャッチャードプロットである。解析は、細胞上のSPGFについて、約5.5×104高親和性受容体(「KD= 0.14nM」)および約8.8×106の低親和性受容体(「KD= 258nM」)の細胞上での存在を示した。
【図10】図10A-Dは、刺激されていないか(「(-)」;図1A)、またはrecSPGF(「SPGF」;図10B)、recEPI(「EPI」:図10C)、もしくはrecEGF(「EGF」;図10D)のいずれかで10分間刺激されたかいずれかのHeLa細胞上のerb-B1受容体(erb-B1受容体に特異的なマウス抗体およびフルオレッセインイソチオシアネート(FITC)結合抗マウスIg抗体によって検出されるもの)の細胞分布を示す一連の蛍光顕微写真である。
【図11】図11Aは、無処置であるか(「(-)」)、またはrecSPGF(「SPGF」)、recEPI(「EPI」)、もしくはrecEGF(「EGF」)で刺激したかいずれかのHeLa細胞の細胞全体の可溶化液のウエスタンブロットの写真である。ウエスタンブロットは、ホスホチロシン残基に特異的なmAb(「WB:4G10」)で染色した。分子量(MW)マーカーのゲル上の位置を写真の左上に示してあり、値は、MW ×10-3であるように示した。 図11Bは、無処置であるか(「(-)」)、またはrecSPGF(「SPGF」)、recEPI(「EPI」)、もしくはrecEGF(「EGF」)で刺激したかいずれかのHeLa細胞の細胞可溶化物の免疫沈降物のウエスタンブロットの写真である。免疫沈降物は、erb-B1(「IP:EGFR」)に対して特異的な抗体によって調製した。ウエスタンブロットは、ホスホチロシン残基に特異的なmAb(「WB:4G10」)で染色した。
【図12】図12A-12Dは、関連した原子の番号付けを示す4-アニリノキナゾリン化合物の一般的な化学構造(「一般的な4-アニリノキナゾリン」;図1D);並びに3つの特異的な4-アニリノキナゾリン化合物、PD153035(図12A)、PD168393(図12B)、およびCI-1033(図12C)の描写である。
【図13】非処理(「(-)」)であるか、またはerb-Bタンパク質チロシンキナーゼ阻害剤に対して事前に露光していないか、もしくは3つのerb-Bタンパク質チロシンキナーゼ阻害剤、PD153035、PD158780、およびPD168393のうちの1つに対して事前に露光した、いずれかrecSPGF(「SPGF」)で刺激したHeLa細胞の細胞全体の可溶化液のウエスタンブロットの写真である。ウエスタンブロットは、ホスホチロシン残基に特異的なmAbで染色した。分子量(MW)マーカーのゲル上の位置を写真の左上に示してあり、値は、MW ×10-3であるように示した。
【図14】図14Aは、前処理なし(「SPGF単独」)か、またはPD153035、CI-1033、もしくはPS168393;および示した濃度のrecSPGF(「ng/ml」)で一晩刺激;での前処理後の細胞周期のS期のヒト初代線維芽細胞(SC-J)の割合を示す線グラフである。 図14Bは、非処理(「(-)」)であるか、またはerb-Bタンパク質チロシンキナーゼ阻害剤(「(+)」)に対して事前に露光していないか、またはSrcファミリー・タンパク質チロシンキナーゼ阻害剤PP2もしくはerb-Bタンパク質チロシンキナーゼ阻害剤、PD153035、PD16839、もしくはCI-1033に対して事前に露光した、recSPGF(「SPGF」)で刺激したかいずれかのHeLa細胞の細胞全体の可溶化液のウエスタンブロットの写真である。ウエスタンブロットは、ホスホチロシン残基に特異的なmAb(「WB:P-Y(4G10)」)で染色した。分子量(MW)マーカーのゲル上の位置を写真の左上に示してあり、値は、MW ×10-3であるように示した。写真の右上の矢印は、erb-B1タンパク質チロシンキナーゼ阻害によって影響を受けたタンパク質リン酸化の位置を示す。 図14Cは、以下の後でのA431細胞上のerb-B1受容体の発現レベル(FFCによって決定されるMFIの)を示す棒グラフである:阻害剤なしでの前処理、またはPP2、PD153035、PD168393、もしくはCI-1033での前処理;およびビオチン化されたrecSPGFとの5分間のインキュベーション。 図14Dは、前処理していないか(「(+)」)、またはPP2、PD153035、PD168393、もしくはCI-1033のいずれかで前処理し、次いでrecSPGFで5分間刺激したHeLa細胞の細胞可溶化物の免疫沈降物(「IP」)のウエスタンブロットの一連の写真である。細胞を前処理しないか、または刺激していないrecSPGF(「(-)」)対照でも行った。免疫沈降物(「IP」)は、erb-B1(「EGFR」)に特異的な抗体またはc-Cbl(「c-Cbl」)に特異的な抗体のいずれかによって調製した。ウエスタンブロットは、erb-B1(「EGFW」;上段パネル)またはc-Cbl(中段および下段パネル)に特異的なmAbsで染色した(「WB」)。
【図15】図15Aは、ワクシニアウイルスで鼻腔内にチャレンジし、かつ無処置か(「対照」;白丸)、抗L1R mAbで一度前処理したか(「抗L1R」;黒丸)、CI-1033で毎日処置したか(「CI-1033」;白三角)、または抗L1R mAbで一度前処理して、CI-1033で毎日処置したか(「CI-1033 +抗L1R」;黒逆三角)のいずれかのマウスの、全時間にわたった生存を(種々の処理群のマウスの割合で)示す線グラフである。 図15Bは、図15Aに記載されているマウス実験群の臨床スコア(実施例1に記載したものに由来する)を示す線グラフである。図15Aに使用する同じシンボルを種々の実験群を示すために使用してある。 図15Cは、ワクシニアウイルスでの致死チャレンジ6日後の、処置なし(「対照」)、抗L1R mAbで前処理(「抗L1R」)、CI-1033で毎日処理(「CI-1033」)、または抗L1R mAbで前処理して、CI-1033で毎日処理したもの(「CI-1033 + 抗L1R」)のいずれかのマウスからの全肺の平均重量を示す棒グラフである。 図15Dは、ワクシニアウイルスに対する感染の8日後の、抗L1R mAbで前処理(「抗L1R」)、CI-1033で毎日処理(「CI-1033」)、13E8 mAbで前処理したもの、抗L1R mAbおよび13E8 mAbで前処理したもの、または抗L1R mAbで前処理して、CI-1033で毎日処理したもの(「CI-1033 + 抗L1R」)のいずれかのマウスの肺における平均ウイルス力価を示す棒グラフである(「Log10PFU」で;PFU(プラーク形成単位))。本実験の対照群(「なし」;斜線の棒)(ウイルスを感染させたが、前処理または処置を受けなかったマウス)のマウスの全てが、実験8日目に死んだので、対照群は、7日めに屠殺したマウスの別々の実験に由来する。
【図16】図16Aは、感染していない(「0」);ワクシニアウイルスに対する感染の示した日数(「4」、「6」または「8」)後の:マウスの肺から調製したRNAを使用するRnase保護アッセイ法からのオートラジオグラムの写真である。感染動物は、無処置であるか(「対照」)、抗L1R mAb(「抗L1R」)で前処理したか、CI-1033(「CI-1033」)で毎日処置したか、または抗L1R mAbで一旦前処理して、CI-1033(「CI-1033 + 抗L1R」)で毎日処置したものであった。写真の左上に示したように、肺RNAは、mCK-2bテンプレートセット(BD Biosciences-Pharmingen, San Diego, CA)のサイトカイン特異的なプローブとハイブリダイズさせた。保護断片をDNAシーケンシング・ゲル上で溶解し、次いでこれをX線写真に曝露した。CI-1033(単独で、または抗L1R mAbと共に)によってレベルが増大されたサイトカインのmRNAを写真の右上の矢印によって示してある。 図16Bは、インビトロにおいて:正常MC57G細胞(「MC57G」);またはワクシニアウイルスに感染したMC57G細胞(「VV感染MC57G」)で刺激されたマウス脾臓CD8+細胞内の細胞内インターフェロン-γのレベル(「IFN-γ(log)」)、または細胞表面CD44(「CD44(log)」)のレベルを示す一連の二次元FFCプロフィールである。実験群は、ワクシニアウイルスに感染して(屠殺し、アッセイする5日前)無処置(「対照」)か、抗L1R mAbで一回前処理した(「抗L1R」)か、CI-1033で毎日処置した(「CI-1033」)か、または抗L1R mAbで一回前処理してCI-1033で毎日処置した(「CI-1033」+「抗L1R」)からなる。示したデータは、マウスのそれぞれの実験群からの代表的マウスからのものである。
【図17】正常MC57G細胞(「MC57G」);ワクシニアウイルス感染MC57G細胞(「VV感染MC57G」);または抗CD 3mAb(「抗CD3」)でインビトロで刺激されたマウス脾臓CD8+細胞の細胞内インターフェロン-γ(「IFN-γ(log)」)のレベル、および細胞表面CD44(「CD44(log)」)のレベルを示す一連の二次元FFCプロフィールである。実験群は、無処置のマウス(「ナイーブ」)またはワクシニアウイルスに感染して(屠殺し、アッセイする5日前)無処置(「対照」)か、抗L1R mAbで一回前処理した(「抗L1R」)か、13E8 mAbで一回前処理した(「13E8」)か、または抗L1R mAbおよび13E8 mAbで一回前処理した(「抗L1R+13E8」)マウスからなる。示したデータは、マウスのそれぞれの実験群からの代表的マウスからのものである。
【図18】図18Aは、示した濃度のCI-1033の非存在下(「-」)または存在下(「+」)のいずれかにおいて、BSC-40細胞単層を培養することによって得た痘瘡株ソライメンの免疫染色を示す一連の写真である。 図18Bは、示した濃度のCI-1033の非存在下(「0」)または存在下(「CI-1033」)のいずれかにおいて、BSC-40細胞単層を培養することによって得たプラークの数に対するCI-1033の効果を示す棒グラフである。 図18Cは、示した濃度のCI-1033の非存在下(「0」)または存在下(「CI-1033」)のいずれかにおいてBSC-40細胞単層を培養することによって得たコメットの数を示す棒グラフである。
【特許請求の範囲】
【請求項1】
(a)ウイルスerb-Bリガンドをコードする遺伝子を含むウイルスに曝露された可能性があるか、または暴露されている可能性がある動物を同定する工程、および、
(b)erb-Bチロシンキナーゼの活性を阻害するか、またはerb-Bチロシンキナーゼの活性化を阻害する化合物で動物を処置する工程、
を含む方法。
【請求項2】
工程(b)の前に、間に、または後に、動物がウイルスに感染していたと同定される、請求項1記載の方法。
【請求項3】
ウイルスerb-Bリガンドがポックスウイルスerb-Bリガンドである、請求項1記載の方法。
【請求項4】
ポックスウイルスがオルソポックスウイルスである、請求項3記載の方法。
【請求項5】
オルトポックスウイルスが大痘瘡(variola major)である、請求項4記載の方法。
【請求項6】
オルトポックスウイルスが小痘瘡(variola minor)である、請求項4記載の方法。
【請求項7】
オルトポックスウイルスがサル痘ウイルスである、請求項4記載の方法。
【請求項8】
オルトポックスウイルスがワクシニアである、請求項4記載の方法。
【請求項9】
erb-Bリガンドがエピレギュリン(epiregulin)様の成長因子(ELGF)である、請求項1記載の方法。
【請求項10】
ELGFが天然痘成長因子(SPGF)である、請求項9記載の方法。
【請求項11】
ELGFがワクシニア成長因子(VGF)である、請求項9記載の方法。
【請求項12】
erb-Bチロシンキナーゼが、erb-B1チロシンキナーゼである、請求項1記載の方法。
【請求項13】
化合物が、erb-Bチロシンキナーゼと結合する非アゴニスト抗体である、請求項1記載の方法。
【請求項14】
化合物が、非アゴニストerb-Bリガンドまたはerb-Bリガンドの非アゴニスト断片である、請求項1記載の方法。
【請求項15】
化合物が、小分子erb-Bチロシンキナーゼ阻害剤である、請求項1記載の方法。
【請求項16】
小分子erb-Bチロシンキナーゼ阻害剤が、キナゾリンに基づいた化合物である、請求項15記載の方法。
【請求項17】
小分子erb-Bチロシンキナーゼ阻害剤が、ピリドピリミジンに基づいた化合物である、請求項15記載の方法。
【請求項18】
小分子erb-Bチロシンキナーゼ阻害剤が、キノリン-3-カルボニトリルに基づいた化合物である、請求項15記載の方法。
【請求項19】
小分子erb-Bチロシンキナーゼ阻害剤が、ピロロピリミジンに基づいた化合物である、請求項15記載の方法。
【請求項20】
キナゾリンに基づいた化合物が、4-アニリノキナゾリンである、請求項16記載の方法。
【請求項21】
小分子erb-Bチロシンキナーゼ阻害剤が、erb-Bチロシンキナーゼ活性の不可逆阻害剤である、請求項15記載の方法。
【請求項22】
不可逆阻害剤が、CI-1033である、請求項21記載の方法。
【請求項23】
キナゾリンに基づいた化合物が、PD168393、PD160678、PD160879、またはPD174265である、請求項16記載の方法。
【請求項24】
キナゾリンに基づいた化合物が、PD153035、ZD1839、GW572016、GW974、OSI-774、またはAG1478である、請求項16記載の方法。
【請求項25】
ピリドピリミジンに基づいた化合物が、PD69896、PD153717、またはPD158780である、請求項17記載の方法。
【請求項26】
キノリン-3-カルボニトリルに基づいた化合物が、EKB-569である、請求項18記載の方法。
【請求項27】
ピロロピリミジンに基づいた化合物が、CGP59326Aである、請求項19記載の方法。
【請求項28】
動物がヒトである、請求項1記載の方法。
【請求項29】
実質的にウイルスの1つまたは複数の形態を中和する能力を有する抗体を動物に投与する工程をさらに含む、請求項1記載の方法。
【請求項30】
抗体が、オルトポックスウイルスの、細胞内成熟ウイルス粒子(IMV)形態、細胞外外膜ウイルス(EEV)形態、または細胞結合エンベロープ・ウイルス(CEV)形態と結合する、請求項29記載の方法。
【請求項31】
オルトポックスウイルスが大痘瘡である、請求項30記載の方法。
【請求項32】
オルトポックスウイルスが小痘瘡である、請求項30記載の方法。
【請求項33】
オルトポックスウイルスがワクシニアである、請求項30記載の方法。
【請求項34】
オルトポックスウイルスがサル痘ウイルスである、請求項30記載の方法。
【請求項35】
ウイルスが、ポックスウイルス発現ベクターであり、かつさらに以下を含む、請求項1記載の方法:(a)免疫原をコードする異種核酸配列;および、(b)異種核酸配列に動作可能に連結される、転写調節性のエレメント(TRE)。
【請求項36】
ポックスウイルス発現ベクターがワクシニアウイルス発現ベクターまたは弱毒ワクシニアウイルス発現ベクターである、請求項35記載の方法。
【請求項37】
ポックスウイルス発現ベクターが、カナリア痘瘡ウイルスまたは鶏痘ウイルスベクターである、請求項35記載の方法。
【請求項38】
処置が、動物におけるウイルスに対する免疫応答を増強する、請求項2記載の方法。
【請求項39】
免疫応答がT細胞応答である、請求項38記載の方法。
【請求項40】
T細胞応答がCD8+ T細胞応答である、請求項39記載の方法。
【請求項41】
T細胞応答がCD4+ T細胞応答である、請求項39記載の方法。
【請求項42】
T細胞応答がインターフェロン-γ産生T細胞応答である、請求項39記載の方法。
【請求項43】
免疫応答が抗体産生B細胞応答である、請求項38記載の方法。
【請求項44】
(a)erb-Bリガンドをコードする遺伝子を含むウイルスによる感染に感受性の動物を同定する工程;並びに、
(b)動物を、(i)ウイルスに対する免疫応答を刺激するウイルスまたはワクチンの1つもしくは複数の形態を実質的に中和する抗体、および(ii)erb-Bチロシンキナーゼを阻害するか、またはerb-Bチロシンキナーゼの活性化を阻害する化合物で処置する工程、
を含む方法。
【請求項45】
(a)erb-Bチロシンキナーゼと結合し、かつチロシンキナーゼ活性の活性または受容体のチロシンキナーゼの活性化を阻害する単離された化合物を提供する工程と;
(b)erb-Bチロシンキナーゼを発現する細胞と化合物を接触させる工程;および、
(c)工程(b)の前に、と同時に、もしくは後に、ウイルスerb-Bリガンドまたはリガンドの機能的な断片と細胞を接触させる工程、
を含むインビトロ方法。
【請求項46】
化合物が、リガンドまたは断片による細胞上のerb-Bチロシンキナーゼの活性化を減少させる、請求項45記載の方法。
【請求項47】
化合物が、リガンドまたは断片による細胞上のerb-Bチロシンキナーゼの活性化を減少させるかどうか決定する工程をさらに含む、請求項44記載の方法。
【請求項48】
化合物が、抗ウイルス性の化合物であるかどうかを決定する方法であって、以下の工程を含む方法:
(a)erb-Bチロシンキナーゼ活性を阻害する化合物を提供する工程;
(b)erb-Bリガンドをコードする遺伝子を含むウイルスに対する感染に感受性の動物に対して化合物を投与する工程;
(c)工程(b)の前に、間に、または後に、ウイルスに対して動物を曝露する工程;および、
(d)化合物が動物のウイルス感染の症状を減少させるかどうかを決定する工程。
【請求項49】
(a)erb-Bリガンドをコードする遺伝子を含むウイルスによる感染に感受性の動物を同定する工程;および、
(b)動物を、(i)リガンドに結合する抗体、および(ii)ウイルスに対する免疫応答を刺激するウイルスまたはワクチンの1つもしくは複数の形態を実質的に中和する抗体で処置する工程、
を含む方法。
【請求項50】
工程(b)の前に、間に、または後に、動物がウイルスに感染していたと同定される、請求項49記載の方法。
【請求項51】
処置がウイルスに対する免疫応答を増強する、請求項50記載の方法。
【請求項52】
免疫応答がT細胞応答である、請求項51記載の方法。
【請求項53】
T細胞応答がCD8+ T細胞応答である、請求項52記載の方法。
【請求項54】
T細胞応答がCD4+ T細胞応答である、請求項52記載の方法。
【請求項55】
T細胞応答が、インターフェロン-γ産生T細胞の免疫応答である、請求項52記載の方法。
【請求項56】
免疫応答が、抗体産生B細胞の応答である、請求項51記載の方法。
【請求項57】
ウイルスが、ポックスウイルス発現ベクターであり、かつさらに以下を含む、請求項50記載の方法:(a)免疫原をコードする異種核酸配列;および、(b)異種核酸配列に動作可能に連結される、転写調節性のエレメント(TRE)。
【請求項58】
ポックスウイルス発現ベクターが、ワクシニアウイルス発現ベクターまたは弱毒ワクシニアウイルス発現ベクターである、請求項57記載の方法。
【請求項59】
ポックスウイルス発現ベクターが、カナリア痘瘡ウイルスまたは鶏痘ウイルスベクターである、請求項56記載の方法。
【請求項60】
リガンドと結合する抗体が、13E8モノクローナル抗体(ATCCアクセッション番号PTA-5040)である、請求項49記載の方法。
【請求項61】
リガンドと結合する抗体が、11D7モノクローナル抗体(ATCCアクセッション番号PTA-5039)である、請求項49記載の方法。
【請求項1】
(a)ウイルスerb-Bリガンドをコードする遺伝子を含むウイルスに曝露された可能性があるか、または暴露されている可能性がある動物を同定する工程、および、
(b)erb-Bチロシンキナーゼの活性を阻害するか、またはerb-Bチロシンキナーゼの活性化を阻害する化合物で動物を処置する工程、
を含む方法。
【請求項2】
工程(b)の前に、間に、または後に、動物がウイルスに感染していたと同定される、請求項1記載の方法。
【請求項3】
ウイルスerb-Bリガンドがポックスウイルスerb-Bリガンドである、請求項1記載の方法。
【請求項4】
ポックスウイルスがオルソポックスウイルスである、請求項3記載の方法。
【請求項5】
オルトポックスウイルスが大痘瘡(variola major)である、請求項4記載の方法。
【請求項6】
オルトポックスウイルスが小痘瘡(variola minor)である、請求項4記載の方法。
【請求項7】
オルトポックスウイルスがサル痘ウイルスである、請求項4記載の方法。
【請求項8】
オルトポックスウイルスがワクシニアである、請求項4記載の方法。
【請求項9】
erb-Bリガンドがエピレギュリン(epiregulin)様の成長因子(ELGF)である、請求項1記載の方法。
【請求項10】
ELGFが天然痘成長因子(SPGF)である、請求項9記載の方法。
【請求項11】
ELGFがワクシニア成長因子(VGF)である、請求項9記載の方法。
【請求項12】
erb-Bチロシンキナーゼが、erb-B1チロシンキナーゼである、請求項1記載の方法。
【請求項13】
化合物が、erb-Bチロシンキナーゼと結合する非アゴニスト抗体である、請求項1記載の方法。
【請求項14】
化合物が、非アゴニストerb-Bリガンドまたはerb-Bリガンドの非アゴニスト断片である、請求項1記載の方法。
【請求項15】
化合物が、小分子erb-Bチロシンキナーゼ阻害剤である、請求項1記載の方法。
【請求項16】
小分子erb-Bチロシンキナーゼ阻害剤が、キナゾリンに基づいた化合物である、請求項15記載の方法。
【請求項17】
小分子erb-Bチロシンキナーゼ阻害剤が、ピリドピリミジンに基づいた化合物である、請求項15記載の方法。
【請求項18】
小分子erb-Bチロシンキナーゼ阻害剤が、キノリン-3-カルボニトリルに基づいた化合物である、請求項15記載の方法。
【請求項19】
小分子erb-Bチロシンキナーゼ阻害剤が、ピロロピリミジンに基づいた化合物である、請求項15記載の方法。
【請求項20】
キナゾリンに基づいた化合物が、4-アニリノキナゾリンである、請求項16記載の方法。
【請求項21】
小分子erb-Bチロシンキナーゼ阻害剤が、erb-Bチロシンキナーゼ活性の不可逆阻害剤である、請求項15記載の方法。
【請求項22】
不可逆阻害剤が、CI-1033である、請求項21記載の方法。
【請求項23】
キナゾリンに基づいた化合物が、PD168393、PD160678、PD160879、またはPD174265である、請求項16記載の方法。
【請求項24】
キナゾリンに基づいた化合物が、PD153035、ZD1839、GW572016、GW974、OSI-774、またはAG1478である、請求項16記載の方法。
【請求項25】
ピリドピリミジンに基づいた化合物が、PD69896、PD153717、またはPD158780である、請求項17記載の方法。
【請求項26】
キノリン-3-カルボニトリルに基づいた化合物が、EKB-569である、請求項18記載の方法。
【請求項27】
ピロロピリミジンに基づいた化合物が、CGP59326Aである、請求項19記載の方法。
【請求項28】
動物がヒトである、請求項1記載の方法。
【請求項29】
実質的にウイルスの1つまたは複数の形態を中和する能力を有する抗体を動物に投与する工程をさらに含む、請求項1記載の方法。
【請求項30】
抗体が、オルトポックスウイルスの、細胞内成熟ウイルス粒子(IMV)形態、細胞外外膜ウイルス(EEV)形態、または細胞結合エンベロープ・ウイルス(CEV)形態と結合する、請求項29記載の方法。
【請求項31】
オルトポックスウイルスが大痘瘡である、請求項30記載の方法。
【請求項32】
オルトポックスウイルスが小痘瘡である、請求項30記載の方法。
【請求項33】
オルトポックスウイルスがワクシニアである、請求項30記載の方法。
【請求項34】
オルトポックスウイルスがサル痘ウイルスである、請求項30記載の方法。
【請求項35】
ウイルスが、ポックスウイルス発現ベクターであり、かつさらに以下を含む、請求項1記載の方法:(a)免疫原をコードする異種核酸配列;および、(b)異種核酸配列に動作可能に連結される、転写調節性のエレメント(TRE)。
【請求項36】
ポックスウイルス発現ベクターがワクシニアウイルス発現ベクターまたは弱毒ワクシニアウイルス発現ベクターである、請求項35記載の方法。
【請求項37】
ポックスウイルス発現ベクターが、カナリア痘瘡ウイルスまたは鶏痘ウイルスベクターである、請求項35記載の方法。
【請求項38】
処置が、動物におけるウイルスに対する免疫応答を増強する、請求項2記載の方法。
【請求項39】
免疫応答がT細胞応答である、請求項38記載の方法。
【請求項40】
T細胞応答がCD8+ T細胞応答である、請求項39記載の方法。
【請求項41】
T細胞応答がCD4+ T細胞応答である、請求項39記載の方法。
【請求項42】
T細胞応答がインターフェロン-γ産生T細胞応答である、請求項39記載の方法。
【請求項43】
免疫応答が抗体産生B細胞応答である、請求項38記載の方法。
【請求項44】
(a)erb-Bリガンドをコードする遺伝子を含むウイルスによる感染に感受性の動物を同定する工程;並びに、
(b)動物を、(i)ウイルスに対する免疫応答を刺激するウイルスまたはワクチンの1つもしくは複数の形態を実質的に中和する抗体、および(ii)erb-Bチロシンキナーゼを阻害するか、またはerb-Bチロシンキナーゼの活性化を阻害する化合物で処置する工程、
を含む方法。
【請求項45】
(a)erb-Bチロシンキナーゼと結合し、かつチロシンキナーゼ活性の活性または受容体のチロシンキナーゼの活性化を阻害する単離された化合物を提供する工程と;
(b)erb-Bチロシンキナーゼを発現する細胞と化合物を接触させる工程;および、
(c)工程(b)の前に、と同時に、もしくは後に、ウイルスerb-Bリガンドまたはリガンドの機能的な断片と細胞を接触させる工程、
を含むインビトロ方法。
【請求項46】
化合物が、リガンドまたは断片による細胞上のerb-Bチロシンキナーゼの活性化を減少させる、請求項45記載の方法。
【請求項47】
化合物が、リガンドまたは断片による細胞上のerb-Bチロシンキナーゼの活性化を減少させるかどうか決定する工程をさらに含む、請求項44記載の方法。
【請求項48】
化合物が、抗ウイルス性の化合物であるかどうかを決定する方法であって、以下の工程を含む方法:
(a)erb-Bチロシンキナーゼ活性を阻害する化合物を提供する工程;
(b)erb-Bリガンドをコードする遺伝子を含むウイルスに対する感染に感受性の動物に対して化合物を投与する工程;
(c)工程(b)の前に、間に、または後に、ウイルスに対して動物を曝露する工程;および、
(d)化合物が動物のウイルス感染の症状を減少させるかどうかを決定する工程。
【請求項49】
(a)erb-Bリガンドをコードする遺伝子を含むウイルスによる感染に感受性の動物を同定する工程;および、
(b)動物を、(i)リガンドに結合する抗体、および(ii)ウイルスに対する免疫応答を刺激するウイルスまたはワクチンの1つもしくは複数の形態を実質的に中和する抗体で処置する工程、
を含む方法。
【請求項50】
工程(b)の前に、間に、または後に、動物がウイルスに感染していたと同定される、請求項49記載の方法。
【請求項51】
処置がウイルスに対する免疫応答を増強する、請求項50記載の方法。
【請求項52】
免疫応答がT細胞応答である、請求項51記載の方法。
【請求項53】
T細胞応答がCD8+ T細胞応答である、請求項52記載の方法。
【請求項54】
T細胞応答がCD4+ T細胞応答である、請求項52記載の方法。
【請求項55】
T細胞応答が、インターフェロン-γ産生T細胞の免疫応答である、請求項52記載の方法。
【請求項56】
免疫応答が、抗体産生B細胞の応答である、請求項51記載の方法。
【請求項57】
ウイルスが、ポックスウイルス発現ベクターであり、かつさらに以下を含む、請求項50記載の方法:(a)免疫原をコードする異種核酸配列;および、(b)異種核酸配列に動作可能に連結される、転写調節性のエレメント(TRE)。
【請求項58】
ポックスウイルス発現ベクターが、ワクシニアウイルス発現ベクターまたは弱毒ワクシニアウイルス発現ベクターである、請求項57記載の方法。
【請求項59】
ポックスウイルス発現ベクターが、カナリア痘瘡ウイルスまたは鶏痘ウイルスベクターである、請求項56記載の方法。
【請求項60】
リガンドと結合する抗体が、13E8モノクローナル抗体(ATCCアクセッション番号PTA-5040)である、請求項49記載の方法。
【請求項61】
リガンドと結合する抗体が、11D7モノクローナル抗体(ATCCアクセッション番号PTA-5039)である、請求項49記載の方法。
【図1】


【図2】

【図3】

【図4】
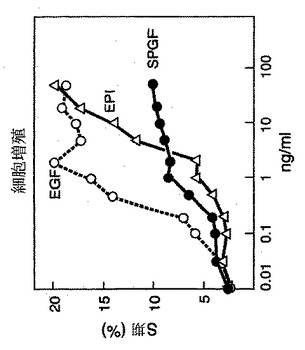
【図5】

【図6】
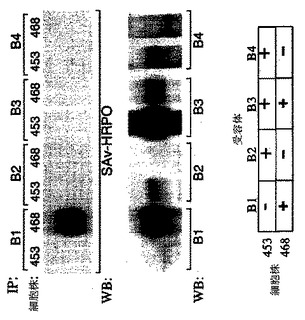
【図7】

【図8】

【図9】

【図10】

【図11】

【図12】

【図13】

【図14A】

【図14B】

【図14C】

【図14D】

【図15】

【図16A】

【図16B】

【図17】

【図18】

【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14A】
【図14B】
【図14C】
【図14D】
【図15】
【図16A】
【図16B】
【図17】
【図18】
【公表番号】特表2007−517767(P2007−517767A)
【公表日】平成19年7月5日(2007.7.5)
【国際特許分類】
【出願番号】特願2006−532321(P2006−532321)
【出願日】平成16年3月11日(2004.3.11)
【国際出願番号】PCT/US2004/007537
【国際公開番号】WO2005/003325
【国際公開日】平成17年1月13日(2005.1.13)
【出願人】(592090692)ダナ ファーバー キャンサー インスティテュート (20)
【Fターム(参考)】
【公表日】平成19年7月5日(2007.7.5)
【国際特許分類】
【出願日】平成16年3月11日(2004.3.11)
【国際出願番号】PCT/US2004/007537
【国際公開番号】WO2005/003325
【国際公開日】平成17年1月13日(2005.1.13)
【出願人】(592090692)ダナ ファーバー キャンサー インスティテュート (20)
【Fターム(参考)】
[ Back to top ]